
Abstract
Oxygenation in the Eastern Equatorial Pacific is responsive to ongoing climate change in the modern ocean, although whether the region saw a deglacial change in extent or position of the Oxygen Minimum Zone remains poorly constrained. Here, stable isotopes from the shells of an Oxygen Minimum Zone-dwelling planktic foraminifer are used to reassess the position of the mid-water Oxygen Minimum Zone relative to both the thermocline and benthos. Oxygen isotopes record a rapid shoaling of the Oxygen Minimum Zone towards the thermocline associated with Heinrich Stadial 1 and persisting through the deglaciation. Meanwhile, carbon isotope similarities between Oxygen Minimum Zone-dwelling Globorotaloides hexagonus and benthic Cibicidoides wuellerstorfi suggest a shared source water through the deglaciation. Results support a direct role for the Eastern Equatorial Pacific in venting carbon to the atmosphere through the deglaciation, a deglacial expansion of the Oxygen Minimum Zone, and a restructuring of mid-water oxygen and carbon dynamics from the glacial to Holocene intervals.
Similar content being viewed by others
Introduction
The oceans are currently experiencing a cascade of rapid changes due to release of carbon to the atmosphere. This encompasses shifts in temperature, stratification, productivity, and circulation, which together influence the availability and distribution of oxygen in the water column. As a result, the global expansion and intensification of Oxygen Minimum Zones (OMZs) has been observed over recent decades1,2,3,4,5. Changes in the extent or intensity of OMZs influence global nutrient cycling5,6,7 and define habitat range for many obligate aerobes8. They are simultaneously important reservoirs of marine carbon at intermediate depths9 due to low ventilation and high remineralization, both of which processes increase CO2 while decreasing O2. As a result, OMZ regions may act as sources of CO2 to the atmosphere.
The Eastern Pacific hosts the largest extent of modern OMZ waters, composed of Eastern Tropical North Pacific (ETNP), Eastern Tropical South Pacific, and Eastern Equatorial Pacific (EEP) components10,11, with the EEP currently an important year-round source of CO2 to the atmosphere12,13. The EEP may have played a similar role in the past, venting carbon from the intermediate or deep ocean into the atmosphere. This is a hypothesis supported by increasing evidence that the deep EEP Pacific was the site of a major carbon reservoir (and by the same drivers, O2 depletion) during the last glacial interval14,15,16,17,18,19,20. Support for the EEP as a glacial carbon reservoir, and subsequently a source of CO2 to the atmosphere during the deglaciation is evidenced by decreased deglacial δ13C of surface and thermocline-dwelling planktic foraminifera21,22, the apparent influence of ‘old’ (radiocarbon depleted) carbon at both thermocline14,23 and some intermediate water sites24 but not others25, and a decrease in near-surface pH across the equatorial Pacific26.
A scarcity of available intermediate depth core sites and limitations in proxies for the pelagic OMZ have made assessments of the deglacial evolution of intermediate to shallow reservoirs of EEP carbon more equivocal. While there is evidence that a mid-water OMZ persisted in some form, existing records have not coalesced into a single history for the EEP OMZ. Nitrogen isotope records extending from the EEP up the Northeastern Pacific margin indicate an increase in denitrification, implying an increase in the volume of anoxic waters in the ETNP, and perhaps EEP as well, since the Last Glacial Maximum27,28. Productivity proxies show a range of trends, but generally suggest an increase in export productivity, and potentially increased subsurface oxygen utilization in the easternmost EEP during peak deglaciation29,30,31,32. This would indicate a distinct history for the EEP compared to the ETNP where OMZs expanded from the deglaciation into the Holocene (e.g.,27,28,33,34,35,36). Concurrently, planktic foraminifera shells from the mixed-layer and thermocline at Cocos Ridge site ODP 1242 have persistently low I/Ca, interpreted as low oxygen near-surface waters from the Last Glacial Maximum through the deglacial16. Neighboring ODP Site 849 shells show an I/Ca minimum during the deglaciation16 and thus either a deep or absent OMZ in the Last Glacial Maximum. Despite this uncertainty, a change in extent of the OMZ from the last glacial to the modern ocean has important implications for Pacific carbon storage and cycling. For example, no change (or a contraction) in the vertical extent of the deglacial EEP OMZ would indicate a massive overall reduction in Pacific carbon storage. A vertical expansion of the OMZ from the glacial to modern EEP, would suggest a change in location (from deep to intermediate waters) of carbon storage and might indicate a smaller amount of overall CO2 released during the deglaciation.
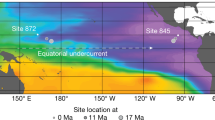
Here we contrast the oxygen and carbon isotope records of OMZ-dwelling Globorotaloides hexagonus shells from Galapagos Margin site TR163-23 (0° 24.6’ N, 92° 9.6’ W; 2,730 m depth; Fig. 1) with previously published records from mixed-layer species Globigerinoides ruber and thermocline-dwelling Neogloboquadrina dutertrei14, and with benthic Cibicidoides wuellerstorfi37. This is supported by records of O2-associated morphological traits in G. hexagonus, which together constrain deglacial changes in the vertical extent and intensity of the EEP OMZ.

a Shows O2 at 250 m depth and the location of TR163-23 (yellow star) as well as other core sites discussed here (blue circles). b Shows a section of O2 with depth along the 92°W meridian showing the location of TR163-23. Oxygen data was sourced from WOA1873.
Results
Morphological support for habitat tracking
In the modern ocean, G. hexagonus is found at sub-oxycline depths, with shell morphologies linked to environmental O2 concentrations across a relatively large range of low oxygen conditions38. Empirical correlations have been identified between porosity, shell length, and chambers in the final whorl of G. hexagonus shells and the oxygenation levels from which live shells were recovered38. None of these metrics are known to be susceptible to dissolution and were thus deemed suitable for analysis at TR163-23 despite suspected changes in deglacial bottom water saturation state14,37. A higher population-level number of chambers during Heinrich Stadial 1 and larger sizes during the Bølling-Allerød (p-value = 0.001 for both) may indicate greater occupation of very low O2 habitat during these deglacial intervals. However, there is no clear trend or any significant difference at the population-level (p-value < 0.05) apparent across multiple oxygen-associated morphological traits and all measurements, except maximum length, are within the range of those found in modern populations (Fig. 2). This supports the assumption that G. hexagonus has continued to live in a broadly defined sub-oxycline habitat through the deglaciation, similar to in the modern ocean.

Yellow arrows indicate the directions in which modern data from ETNP plankton tows38 exceed the range of fossil data in each trait.
Oxygen isotopes demonstrate deglacial shoaling of the EEP OMZ
The δ18O of planktic foraminifera is driven by both temperature and the δ18O of seawater, such that the shells of individuals living in deeper, colder habitats will record higher δ18O values than those living in shallower, warmer waters (Supplementary Fig. 1)39,40. As foraminifera track their preferred habitat within the water column41, comparing the δ18O values of foraminifera constrained to different habitats allows for a reconstruction of the relative vertical positioning of these habitats and thus water column structure through time. This approach has been used widely, including in the EEP42,43,44 focused on either thermocline or deep water column structure. Here we focus on the OMZ by comparing the δ18O and δ13C of G. hexagonus with published values for mixed layer species G. ruber, thermocline N. dutertrei, and benthic C. wuellerstorfi.
Globigerinoides ruber lives in the tropical to subtropical mixed layer, constrained to the photic zone by photosymbionts and is found at 25–60 m in the modern Panama Basin39. By contrast, N. dutertrei lives in or near the thermocline, likely tracking the chlorophyll maximum at 35–100 m depth in the Panama Basin39 with an apparent calcification depth of <70 m45. Finally, G. hexagonus, lives within sub-thermocline, low oxygen waters38,46,47, and has been recovered as deep as 1000 m in the Panama Basin39. Therefore, a change in the δ18O value of G. hexagonus shells relative to G. ruber and N. dutertrei should indicate a vertical migration of the OMZ (sub-oxycline) habitat relative to the mixed layer and thermocline. The relative positioning of the OMZ in this region is of particular interest due both to its potential importance for the global nitrogen budget as well as its intermediate positioning between thermocline/sub-thermocline upwelling source waters, and older, deeper water masses.
At site TR163-23 the spread of δ18O values between the three planktic species demonstrates a relatively large thermocline to OMZ gradient at the end of the last glacial interval (Fig. 3). An abrupt decrease in the δ18O values of G. hexagonus follows during Heinrich Stadial 1, both independent of and relative to mixed layer G. ruber and thermocline N. dutertrei (Figs. 3, 4). This is interpreted as a shallowing of the mean habitat depth of G. hexagonus and shoaling of the EEP OMZ during the deglaciation. The gradient between the δ18O of G. hexagonus and N. dutertrei may be an especially important indicator of how close the OMZ lies to the thermocline, and thus how accessible deep, O2-poor, carbon and nutrient-rich waters are to primary producers in the photic zone and to the atmosphere. A reduced thermocline-OMZ gradient persisted through the deglacial interval up to the Holocene onset (Figs. 3, 4). By the mid-Holocene, a more substantial gradient between the δ18O of the three species was restored, though with a smaller offset between N. dutertrei and G. hexagonus than during the glacial interval, suggestive of a shallower oxycline than was present during the glacial. Assuming both N. dutertrei and G. hexagonus incorporate δ18O close to equilibrium (or with similar offsets), the implication is an OMZ which shoaled nearly into the thermocline during much of the deglacial interval. This would be expected to result in increased connectivity between the OMZ, the mixed layer, and the atmosphere, and thus a direct venting of deep Pacific carbon to the atmosphere during the deglaciation.

The a δ18O records from G. ruber (pink), N. dutertrei (gray), G. hexagonus (yellow), and C. wuellerstorfi (blue) and b δ13C records from G. ruber (pink), N. dutertrei (gray), G. hexagonus (yellow), and C. wuellerstorfi (blue) are shown with a loess curve fit through each record. Below are schematic reconstructions of water column oxygen in the Holocene, deglacial interval, and late glacial/early deglacial interval. Data for G. hexagonus is supplemented with data from14,37. Species are ordered in the key by habitat depth. Measurement error is 0.05‰ for δ18O and 0.03‰ for δ13C, which is smaller than the points shown.

The a relative difference in δ18O between G. hexagonus and G. dutertrei (Δδ18Ohex-dut) with the interval indicating a shoaled OMZ in red, b relative difference in δ18O between C. wuellerstorfi and G. hexagonus (Δδ18Cwuel-hex) with the interval where that difference is near 0 in blue. The overlapping period of both shoaling and minimal δ13Cwuel-hex is shown in purple. These records are compared with c δ13C records of G. hexagonus and C. wuellerstorfi from ODP 124020,61 shown with 5-point moving averages through each, d organic carbon records74, e bulk δ15N records29, f I/Ca from N. dutertrei16, and g N. dutertrei bound δ15N records from neighboring sites60. Propagated measurement error is 0.07‰ for Δδ13Chex-dut values in a and 0.04‰ for Δδ13Cwuel-hex in b, which is smaller than the points shown.
A shallow OMZ is found coincident with the period of maximal nutrient delivery to surface and thermocline waters as well as export productivity. Such a pattern has been demonstrated consistently through the EEP30 and is supported by a decrease in δ13C of surface-dwelling foraminifera at this37 (Fig. 3) and nearby21 sites, sedimentary nitrogen records29,31,48, and foraminifera-bound nitrogen isotopes49 (Fig. 4). An increase in productivity has classically been associated with changes in source water composition21,22, decreased stratification50 evidenced by a shoaling of the thermocline42, and increased upwelling32 in the EEP. Results here are consistent with a decrease in stratification and shoaling of deep, nutrient-rich waters that may have allowed for more productive upwelling, with increased export and remineralization acting as a positive feedback to a shoaling OMZ. It is worth noting that the period of a shoaled OMZ appears to outlast deglacial productivity, which peaks in the Bølling-Allerød (Fig. 4).
Stable carbon isotope evidence for deglacial connectivity with bottom waters
Despite a convergence in δ18O space, the δ13C of G. hexagonus remains distinct from the two shallower-dwelling species. The δ13C values of G. hexagonus have consistently shown little or no offset from equilibrium with the δ13C of DIC46,51,52. Contrastingly, N. dutertrei and G. ruber both calcify out of equilibrium with seawater, with offsets of 0.9 reported in the positive direction in G. ruber and of −0.5 in N. dutertrei43. Thus, assuming near-equilibrium behavior in G. hexagonus, correction for δ13C in the two shallower dwelling species would still result in a substantial difference between the records. Given this and what is known about the habitat of G. hexagonus, it is likely that at least some of this offset is the result of G. hexagonus occupying a distinct, intermediate water mass throughout the record. Globorotaloides hexagonus δ13C records are in close agreement with those from nearby Panama Basin Site ODP 124053 (Fig. 4) which have been interpreted as recording a shift between greater contribution of nutrient-rich Glacial North Pacific Intermediate Water during Marine Isotope Stage 2 and less nutrient-rich southern sourced intermediate waters at present.
When the δ13C of G. hexagonus is compared with that of C. wuellerstorfi, the two records overlap through most the deglaciation. Previous work has shown that, like G. hexagonus, the δ13C signal of C. wuellerstorfi shells is near equilibrium with DIC54,55,56. Thus, the similarity between the two records through the deglacial interval can be interpreted as both species occupying a water mass with similar DIC and carbon source, extending from the upper oxycline to the benthos (2730 m). This is consistent with the presence of a single massive carbon reservoir during the late glacial and deglaciation16 (Fig. 2). Deglacial overlap between the δ13C of G. hexagonus (Max et al. 2017) and C. wuellorstorfi20 is also observed at ODP 1240 further east in the Panama Basin (2921 m depth), demonstrating that the similarity in intermediate and deep δ13C signal is not a feature unique to the Galapagos margin (Fig. 4).
Discussion
The records presented here are consistent with the presence of a deep carbon reservoir in the EEP at the termination of the last glacial interval19,57,58, which was partially ventilated through intermediate23,24 and thermocline/mixed layer21 waters in the EEP during the deglaciation. Oxygen isotope records from G. hexagonus corroborate previous findings in suggesting enhanced connectivity from near thermocline waters down to at least 2730 m depth, from 16.0 to 11.3 ka, and support that the EEP was likely an important source of CO2 during the deglaciation59. Moreover, while intermediate oxygen-depleted waters have been present since the last glacial interval, the oxycline was then substantially deeper than during the deglaciation or in the modern ocean. Finally, these results demonstrate that a shallow, intermediate water OMZ disconnected from deep waters in the EEP is a Holocene phenomenon, representing an oxygen and carbon profile distinct from either the last glacial or deglacial intervals.
This framework additionally provides an explanation for the mismatch between foraminifera-bound and bulk δ15N identified by60. At the end of the last glacial interval, a deep oxycline would have meant most denitrification was taking place too deep in the water column for a strong δ15N signal to reach productive waters, and thus to be exported to underlying sediments. A shoaling of the oxycline into or near the thermocline during the deglaciation would have allowed the isotopic signature of denitrification to be reflected in both foraminiferal-bound and bulk isotopic records. Upon reaching a modern state, the oxycline was shallow enough that some high δ15N, indicative of denitrification is available to upwelling centers and eventual export but much of this process is occurring below the thermocline and thus less represented in foraminifera-bound records.
At the end of the last glacial interval, G. hexagonus δ18O records are diagnostic of a relatively deep OMZ and greater formation and equatorward transport of well-oxygenated Glacial North Pacific Intermediate Water during the deglaciation extending into shallow-intermediate depths of the EEP58. A deep OMZ is also consistent with δ15N records suggesting reduced denitrification in intermediate to shallow EEP and ETNP waters during the last glacial interval27,28,60 and reduced I/Ca in surface waters at some sites16. While the available data does not place firm constraints on the depth of the upper oxycline, the deepest occurrence of modern day hexagonus at ~1000 m39 is likely a reasonable lower limit to depth, given the continued presence of G. hexagonus.
The shoaling of the oxycline observed during the deglaciation is consistent with decreased stratification42,50, increased productivity, and increased shallow denitrification49. The simultaneous shoaling of the OMZ and apparent connectivity of the mid-water OMZ with deep carbon reservoirs is suggestive of the degree to which shallow water masses may have been directly influenced by deep and intermediate water sources. Some evidence has pointed to a greater contribution of northern sources to deglacial EEP intermediate waters, and while G. hexagonus records at TR163-23 agree with δ13C records interpreted as such61,62, this data cannot settle that debate. Regardless of high-latitude contribution, this isotopic evidence leads to the conclusion that the deglacial EEP hosted a single massive carbon reservoir extending from the upper oxycline to the deep ocean, and that the region would have been a substantial source of carbon during the deglaciation59.
Differentiation between the δ13C of G. hexagonus and C. wuellerstorfi does not occur until the Holocene onset, when bottom waters at the site were ultimately ventilated14,37. This is at a substantial time-lag to the most dramatic shoaling of the OMZ (Fig. 3) and thus local venting is unlikely to have been the only cause of deep carbon retreat from the Galapagos Margin. A more complex ventilation history is consistent with the “missing” local carbonate maximum expected to cooccur with deglacial venting63. By the mid-Holocene, differentiation between the G. hexagonus and C. wuellerstorfi δ13C records is suggestive of distinct carbon environments, analogous to a modern water column structure, with a relatively shallow (<100 m in the modern ocean) OMZ and moderately oxygenated bottom waters. In the modern ocean, the EEP OMZ continues to act as a substantial reservoir for carbon at intermediate depths, with the potential for expansion and shoaling of the OMZ to increase connectivity between ocean carbon and the atmosphere in a warming world.
Methods
Oceanographic setting
The modern day EEP is typified by a strong and relatively shallow thermocline shoaling towards the east (< 100 m) and experiences upwelling at both the equator and along the eastern margins64,65. Core TR163-23 was recovered on the northwest margin of the Galapagos Islands. While the Galapagos currently sits within a zone of equatorial upwelling that is amplified by topography, TR163-23 is just to the north of the most intense upwelling and high primary productivity associated with the eastern island boundaries66. Below the pycnocline, this region sees Equatorial Undercurrent, a mix of high-latitude (primarily southern-sourced in the modern ocean) waters delivered via the western Pacific, from which most modern upwelling in the region is sourced65,67. Below that are Antarctic Intermediate Water between ~500 and 1000 m depth66. Due to the presence of the Equatorial Undercurrent, the equatorial region experiences a less intense and expansive OMZ than to either the north or south67, though a strong and relatively shallow OMZ is still present beneath the thermocline (Fig. 1; Supplementary Fig. 2).
Foraminiferal morphology
Planktic foraminifera of the species G. hexagonus were isolated from the > 150 μm fraction of sediments from core TR163-23 at a 5 cm resolution. Age models for this core were adopted from previous work14. Up to 32 individuals were selected at random for further morphometric analyses. Each individual was imaged on a Leica DM6000 light microscope at Yale University, with measurements of length made using the AutoMorph software68. Porosity is reported as the percentage of test surface area comprised of pores, derived from using the thresholding tool in the ImageJ Software69. Measurement accuracy of length measurements is ± 1 μm and repeated measurements of porosity on the same individuals give a measurement error of ± 2.3%.
Isotopic measurements
Foraminifera for isotopic measurements were selected from the entire > 150 μm size fraction. While a slight increase in δ18O values with size was previously noted46, a wide range of sizes were included to enable an adequate sample mass (> 20 μg) for analyses. Given the lack of systematic size change through the core (Fig. 2), any size effect on the δ18O records presented here is considered negligible. Where possible, 10 individuals were included in a measurement and no sample has fewer than 3 individuals. The δ18O and δ13C of all G. hexagonus was measured on a Kiel IV Carbonate Device with a Thermo MAT 253 in the Yale Analytical and Stable Isotope Center. The long-term standard deviation of standard reference materials is 0.05‰ for δ18O and 0.03‰ for δ13C. Data for both are reported relative to VPDB.
Statistical treatment
Statistics presented here were carried out using the R Software70. When testing for differences between morphological parameters, non-parametric statistics were favored to account for inherent non-normality in chamber number. For this reason, a Kruskal-Wallis test was used, followed by Dunn’s test with Bonferroni adjusted p-values. The deglacial intervals are delineated as the Last Glacial Maximum (>17.9 ka), the Heinrich Stadial 1 interval (17.9–14.7 ka), the Bølling-Allerød (14.7–12.9 ka), the Younger Dryas (12.9–11.7 ka) and the Holocene (<11.7 ka)71,72.
Data availability
All isotope and morphometric data shown in Figs. 2, 3, and 4 and needed to reproduce the analyses reported here have been deposited in the PANGAEA database (doi: 10.1594/PANGAEA.944997 and 10.1594/PANGAEA.944996) and are additionally available as a supplement to this manuscript (Supplementary Data 1 & 2). Datasets used or referenced from previously published works are cited in the text.
References
Stramma, L., Johnson, G.C., Firing, E. & Schmidtko, S. Eastern Pacific oxygen minimum zones: Supply paths and multidecadal changes. J. Geophys. Res. Oceans 115, https://doi.org/10.1029/2009JC005976 (2010).
Stramma, L., Johnson, G. C., Sprintall, J. & Mohrholz, V. Expanding oxygen minimum zones in the tropical oceans. Science 320, 655–658 (2008).
Levin, L. Manifestation, drivers, and emergence of open ocean deoxygenation. Ann. Rev. Mar. Sci. 10, 229–260 (2017).
Keeling, R. F., Körtzinger, A. & Gruber, N. Ocean deoxygenation in a warming world. Ann. Rev. Mar. Sci 2, 199–229 (2009).
Breitburg, D. et al. Declining oxygen in the global ocean and coastal waters. Science 359, 6371 (2018).
Gruber, N. The marine nitrogen cycle: overview and challenges. Nitrogen Marine Environ. 2, 1–50 (Elsevier, 2008).
DeVries, T., Deutsch, C., Primeau, F., Chang, B. & Devol, A. Global rates of water-column denitrification derived from nitrogen gas measurements. Nat. Geosci. 5, 547 (2012).
Stramma, L. et al. Expansion of oxygen minimum zones may reduce available habitat for tropical pelagic fishes. Nat. Clim. Change 2, 33–37 (2012).
Paulmier, A., Ruiz-Pino, D. & Garçon, V. CO2 maximum in the oxygen minimum zone (OMZ). Biogeosciences 8, 239–252 (2011).
Bianchi, D., Dunne, J.P., Sarmiento, J.L. & Galbraith, E.D. Data-based estimates of suboxia, denitrification, and N2O production in the ocean and their sensitivities to dissolved O2. Global Biogeochem. Cy. 26, https://doi.org/10.1029/2011gb004209 (2012).
Paulmier, A. & Ruiz-Pino, D. Oxygen minimum zones (OMZs) in the modern ocean. Prog. Oceanogr. 80, 113–128 (2009).
Takahashi, T. et al. Climatological mean and decadal change in surface ocean pCO2, and net sea–air CO2 flux over the global oceans. Deep Sea Res. Part II 56, 554–577 (2009).
Takahashi, T. et al. Global sea–air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects. Deep Sea Res. Part II 49, 1601–1622 (2002).
Umling, N. E. & Thunell, R. C. Synchronous deglacial thermocline and deep-water ventilation in the eastern equatorial Pacific. Nat. Commun. 8, 14203 (2017).
Umling, N. E., Thunell, R. C. & Bizimis, M. Deepwater expansion and enhanced remineralization in the Eastern Equatorial Pacific during the Last Glacial Maximum. Paleoceanogr. Paleoclim. 33, 563–578 (2018).
Hoogakker, B. A. A. et al. Glacial expansion of oxygen-depleted seawater in the eastern tropical Pacific. Nature 562, 410–413 (2018).
Jacobel, A. W. et al. Deep Pacific storage of respired carbon during the last ice age: Perspectives from bottom water oxygen reconstructions. Quat. Sci. Rev. 230, 106065 (2020).
Anderson, R. F. et al. Deep-sea oxygen depletion and ocean carbon sequestration during the Last Ice Age. Global Biogeochem. Cy. 33, 301–317 (2019).
de la Fuente, M., Skinner, L., Calvo, E., Pelejero, C. & Cacho, I. Increased reservoir ages and poorly ventilated deep waters inferred in the glacial Eastern Equatorial Pacific. Nat. Commun. 6, 7420 (2015).
de la Fuente, M. et al. The evolution of deep ocean chemistry and respired carbon in the Eastern Equatorial Pacific over the last deglaciation. Paleoceanography 32, 1371–1385 (2017).
Spero, H. J. & Lea, D. W. The cause of carbon isotope minimum events on glacial terminations. Science 296, 522–525 (2002).
Pena, L. D., Cacho, I., Ferretti, P. & Hall, M. A. El Niño–Southern Oscillation–like variability during glacial terminations and interlatitudinal teleconnections. Paleoceanography 23, 3 (2008).
Stott, L., Southon, J., Timmermann, A. & Koutavas, A. Radiocarbon age anomaly at intermediate water depth in the Pacific Ocean during the last deglaciation. Paleoceanography 24, 2 (2009).
Bova, S. C., Herbert, T. D. & Altabet, M. A. Ventilation of northern and southern sources of aged carbon in the Eastern Equatorial Pacific during the Younger Dryas rise in atmospheric CO2. Paleoceanogr. Paleoclim. 33, 1151–1168 (2018).
Chen, T. et al. Persistently well-ventilated intermediate-depth ocean through the last deglaciation. Nat. Geosci. 13, 733–738 (2020).
Kubota, K., Yokoyama, Y., Ishikawa, T., Obrochta, S. & Suzuki, A. Larger CO2 source at the equatorial Pacific during the last deglaciation. Sci. Rep. 4, 5261 (2014).
Emmer, E. & Thunell, R. C. Nitrogen isotope variations in Santa Barbara Basin sediments: Implications for denitrification in the eastern tropical North Pacific during the last 50,000 years. Paleoceanography 15, 377–387 (2000).
Kienast, S. S., Calvert, S. E. & Pedersen, T. F. Nitrogen isotope and productivity variations along the northeast Pacific margin over the last 120 kyr: Surface and subsurface paleoceanography. Paleoceanography 17, 7–11 (2002).
Robinson, R. S., Martinez, P., Pena, L. D. & Cacho, I. Nitrogen isotopic evidence for deglacial changes in nutrient supply in the eastern equatorial Pacific. Paleoceanography 24, 4 (2009).
Costa, K. M. et al. Productivity patterns in the equatorial Pacific over the last 30,000 years. Global Biogeochem. Cy. 31, 850–865 (2017).
Martinez, P. & Robinson, R. S. Increase in water column denitrification during the last deglaciation: the influence of oxygen demand in the eastern equatorial Pacific. Biogeosciences 7, 1–9 (2010).
Kienast, M. et al. Eastern Pacific cooling and Atlantic overturning circulation during the last deglaciation. Nature 443, 846–849 (2006).
Moffitt, S. E., Hill, T. M., Ohkushi, K., Kennett, J. P. & Behl, R. J. Vertical oxygen minimum zone oscillations since 20 ka in Santa Barbara Basin: A benthic foraminiferal community perspective. Paleoceanography 29, 44–57 (2014).
Ohkushi, K. et al. Quantified intermediate water oxygenation history of the NE Pacific: A new benthic foraminiferal record from Santa Barbara basin. Paleoceanography 28, 453–467 (2013).
Hendy, I. & Pedersen, T. Oxygen minimum zone expansion in the eastern tropical North Pacific during deglaciation. Geophys. Res. Lett. 33, 20 (2006).
Erdem, Z. et al. Bottom-water deoxygenation at the Peruvian margin during the last deglaciation recorded by benthic foraminifera. Biogeosciences 17, 3165–3182 (2020).
Umling, N. E. & Thunell, R. C. Mid-depth respired carbon storage and oxygenation of the eastern equatorial Pacific over the last 25,000 years. Quat. Sci. Rev. 189, 43–56 (2018).
Davis, C. V., Wishner, K., Renema, W. & Hull, P. M. Vertical distribution of planktic foraminifera through an oxygen minimum zone: How assemblages and test morphology reflect oxygen concentrations. Biogeosciences 18, 977–992 (2021).
Fairbanks, R. G., Sverdlove, M., Free, R., Wiebe, P. H. & Bé, A. W. Vertical distribution and isotopic fractionation of living planktonic foraminifera from the Panama Basin. Nature 298, 841–844 (1982).
Emiliani, C. Depth habitats of some species of pelagic foraminifera as indicated by oxygen isotope ratios. Am. J. Sci. 252, 149–158 (1954).
Rebotim, A. et al. Factors controlling the depth habitat of planktonic foraminifera in the subtropical eastern North Atlantic. Biogeosciences 14, 827–859 (2017).
Ford, H., McChesney, C., Hertzberg, J. & McManus, J. A deep eastern equatorial Pacific thermocline during the Last Glacial Maximum. Geophys. Res. Let. 45, 11,806–811 (2018).
Spero, H.J., Mielke, K.M., Kalve, E.M., Lea, D.W. & Pak, D.K. Multispecies approach to reconstructing eastern equatorial Pacific thermocline hydrography during the past 360 kyr. Paleoceanography 18, https://doi.org/10.1029/2002PA000814 (2003).
Patrick, A. & Thunell, R. C. Tropical Pacific sea surface temperatures and upper water column thermal structure during the Last Glacial Maximum. Paleoceanography 12, 649–657 (1997).
Mekik, A.F. & Anderson, R. Is the core top modern? Observations from the Eastern Equatorial Pacific. Quat. Sci. Rev. 186, https://doi.org/10.1016/j.quascirev.2018.01.020 (2018).
Birch, H., Coxall, H. K., Pearson, P. N., Kroon, D. & O’Regan, M. Planktonic foraminifera stable isotopes and water column structure: Disentangling ecological signals. Mar. Micropaleontol. 101, 127–145 (2013).
Ortiz, J. D. & Mix, A. C. The spatial distribution and seasonal succession of planktonic foraminifera in the California Current off Oregon, September 1987–September 1988. Geol. Soc. London Spec. Publ. 64, 197–213 (1992).
Martínez, I., Rincon, D., Yokoyama, Y. & Barrows, T. Foraminifera and coccolithophorid assemblage changes in the Panama Basin during the last deglaciation: Response to sea-surface productivity induced by a transient climate change. Palaeogeogr. Palaeoclim. Palaeoecol. 234, 114–126 (2006).
Studer, A. S. et al. Ice age-Holocene similarity of foraminifera-bound nitrogen isotope ratios in the eastern equatorial Pacific. Paleoceanogr. Paleoclimatol. 36, e2020PA004063 (2021).
Bova, S. C. et al. Links between eastern equatorial Pacific stratification and atmospheric CO2 rise during the last deglaciation. Paleoceanography 30, 1407–1424 (2015).
Ortiz, J. D., Mix, A., Rugh, W., Watkins, J. & Collier, R. Deep-dwelling planktonic foraminifera of the northeastern Pacific Ocean reveal environmental control of oxygen and carbon isotopic disequilibria. Geochim. Cosmochim. Acta. 60, 4509–4523 (1996).
Rippert, N. et al. Constraining foraminiferal calcification depths in the western Pacific warm pool. Mar. Micropaleontol. 128, 14–27 (2016).
Max, L. et al. Evidence for enhanced convection of North Pacific Intermediate Water to the low-latitude Pacific under glacial conditions. Paleoceanogr. 32, 41–55 (2017).
Duplessy, J.-C. et al. 13C Record of benthic foraminifera in the last interglacial ocean: Implications for the carbon cycle and the global deep water circulation. Quat. Res. 21, 225–243 (1984).
Belanger, P. E., Curry, W. B. & Matthews, R. K. Core-top evaluation of benthic foraminiferal isotopic ratios for paleo-oceanographic interpretations. Palaeogeogr. Palaeoclim. Palaeoecol. 33, 205–220 (1981).
Zahn, R., Winn, K. & Sarnthein, M. Benthic foraminiferal δ13C and accumulation rates of organic carbon: Uvigerina peregrina group and Cibicidoides wuellerstorfi. Paleoceanography 1, 27–42 (1986).
Keigwin, L. & Lehman, S. Radiocarbon evidence for a possible abyssal front near 3.1 km in the glacial equatorial Pacific Ocean. Earth Planet. Sci. Let. 425, 93–104 (2015).
Rae, J. W. et al. Overturning circulation, nutrient limitation, and warming in the Glacial North Pacific. Sci. Adv. 6, eabd1654 (2020).
Martínez-Botí, M. A. et al. Plio-Pleistocene climate sensitivity evaluated using high-resolution CO 2 records. Nature 518, 49–54 (2015).
Studer, A. S. et al. Ice age‐Holocene similarity of foraminifera‐bound nitrogen isotope ratios in the eastern equatorial Pacific. Paleoceanogr. Paleoclim. 36, e2020PA004063 (2021).
Max, L. et al. Evidence for enhanced convection of North Pacific Intermediate Water to the low‐latitude Pacific under glacial conditions. Paleoceanogr. Paleoclim. 32, 41–55 (2017).
Rippert, N. et al. Alternating influence of northern versus southern‐sourced water masses on the equatorial Pacific subthermocline during the past 240 ka. Paleoceanogr. 32, 1256–1274 (2017).
Mekik, A. F., Anderson, R., Loubere, P., François, R. & Richaud, M. The Mystery of the Missing Deglacial Carbonate Preservation Maximum. Quat. Sci. Rev. 39, 60–72 (2012).
Fiedler, P. C. & Talley, L. D. Hydrography of the eastern tropical Pacific: A review. Progr. Oceanogr. 69, 143–180 (2006).
Kessler, W. S. The circulation of the eastern tropical Pacific: A review. Progr. Oceanogr. 69, 181–217 (2006).
Pennington, J. T. et al. Primary production in the eastern tropical Pacific: A review. Progr. Oceanogr. 69, 285–317 (2006).
Goodman, P. J., Hazeleger, W., de Vries, P. & Cane, M. Pathways into the Pacific equatorial undercurrent: A trajectory analysis. J. Phys. Oceanogr. 35, 2134–2151 (2005).
Hsiang, A. Y. et al. AutoMorph: Accelerating morphometrics with automated 2D and 3D image processing and shape extraction. Methods Ecol. Evol. 9, 605–612 (2018).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 9, 671–675 (2012).
Team, R. C. (Vienna, Austria, 2013).
Clark, P. U. et al. Global climate evolution during the last deglaciation. Proc. Natl. Acad. Sci. 109, 1134–1142 (2012).
Rasmussen, K. et al. The sustainability of cassava-based bioethanol production in southern Mali. Geografisk Tidsskrift-Danish J, Geogr. 115, 14–26 (2015).
Garcia, H. et al. World Ocean Atlas 2018, Volume 3: Dissolved Oxygen, Apparent Oxygen Utilization, and Dissolved Oxygen Saturation. (2019).
Dubois, N. et al. Millennial-scale variations in hydrography and biogeochemistry in the Eastern Equatorial Pacific over the last 100 kyr. Quat. Sci. Rev. 30, 210–223 (2011).
Acknowledgements
Many thanks to N Umling for valuable discussions, P Hull for comments on an early draft, and B Erkkila and B Chang for laboratory assistance. This work was supported by NSF OCE 1851589 to CVD.
Author information
Authors and Affiliations
Contributions
C.V.D. designed the study, collected data, carried out analyses, and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The author declares no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Zunli Lu and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Joe Aslin.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Davis, C.V. Deglacial restructuring of the Eastern equatorial Pacific oxygen minimum zone. Commun Earth Environ 3, 145 (2022). https://doi.org/10.1038/s43247-022-00477-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-022-00477-8
This article is cited by
-
The Peruvian oxygen minimum zone was similar in extent but weaker during the Last Glacial Maximum than Late Holocene
Communications Earth & Environment (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
