
Abstract
The obesity epidemic continues to worsen worldwide, driving metabolic and chronic inflammatory diseases. Thiazolidinediones, such as rosiglitazone (Rosi), are PPARγ agonists that promote ‘M2-like’ adipose tissue macrophage (ATM) polarization and cause insulin sensitization. As ATM-derived small extracellular vesicles (ATM-sEVs) from lean mice are known to increase insulin sensitivity, we assessed the metabolic effects of ATM-sEVs from Rosi-treated obese male mice (Rosi-ATM-sEVs). Here we show that Rosi leads to improved glucose and insulin tolerance, transcriptional repolarization of ATMs and increased sEV secretion. Administration of Rosi-ATM-sEVs rescues obesity-induced glucose intolerance and insulin sensitivity in vivo without the known thiazolidinedione-induced adverse effects of weight gain or haemodilution. Rosi-ATM-sEVs directly increase insulin sensitivity in adipocytes, myotubes and primary mouse and human hepatocytes. Additionally, we demonstrate that the miRNAs within Rosi-ATM-sEVs, primarily miR-690, are responsible for these beneficial metabolic effects. Thus, using ATM-sEVs with specific miRNAs may provide a therapeutic path to induce insulin sensitization.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
Data availability
RNA sequencing data of SVCs have been deposited in the Gene Expression Omnibus under accession no. GSE234046. Source data are provided with this paper.
Code availability
No custom code was used.
References
Nianogo, R. A. & Arah, O. A. Forecasting obesity and type 2 diabetes incidence and burden: the ViLA-obesity simulation model. Front. Public Health 10, 818816 (2022).
Lee, Y. S. & Olefsky, J. Chronic tissue inflammation and metabolic disease. Genes Dev. 35, 307–328 (2021).
Rohm, T. V., Meier, D. T., Olefsky, J. M. & Donath, M. Y. Inflammation in obesity, diabetes, and related disorders. Immunity 55, 31–55 (2022).
Xu, H. et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 112, 1821–1830 (2003).
Weisberg, S. P. et al. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Invest. 112, 1796–1808 (2003).
Emont, M. P. et al. A single-cell atlas of human and mouse white adipose tissue. Nature 603, 926–933 (2022).
Sárvári, A. K. et al. Plasticity of epididymal adipose tissue in response to diet-induced obesity at single-nucleus resolution. Cell Metab. 33, 437–453 (2021).
Jaitin, D. A. et al. Lipid-associated macrophages control metabolic homeostasis in a Trem2-dependent manner. Cell 178, 686–698 (2019).
Kalluri, R. & LeBleu, V. S. The biology, function, and biomedical applications of exosomes. Science 367, eaau6977 (2020).
Doyle, L. M. & Wang, M. Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 8, 727 (2019).
Ying, W. et al. MiR-690, an exosomal-derived miRNA from M2-polarized macrophages, improves insulin sensitivity in obese mice. Cell Metab. 33, 781–790 (2021).
Ying, W. et al. Adipose tissue macrophage-derived exosomal miRNAs can modulate in vivo and in vitro insulin sensitivity. Cell 171, 372–384 (2017).
Isaac, R., Reis, F. C. G., Ying, W. & Olefsky, J. M. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab. 33, 1744–1762 (2021).
Chhoy, P., Brown, C. W., Amante, J. J. & Mercurio, A. M. Protocol for the separation of extracellular vesicles by ultracentrifugation from in vitro cell culture models. STAR Protoc. 2, 100303 (2021).
Gao, H. et al. MiR-690 treatment causes decreased fibrosis and steatosis and restores specific Kupffer cell functions in NASH. Cell Metab. 34, 978–990 (2022).
Kita, S., Maeda, N. & Shimomura, I. Interorgan communication by exosomes, adipose tissue, and adiponectin in metabolic syndrome. J. Clin. Invest. 129, 4041–4049 (2019).
Huang, Z. & Xu, A. Adipose extracellular vesicles in intercellular and inter-organ crosstalk in metabolic health and diseases. Front. Immunol. 12, 608680 (2021).
Ji, Y. et al. Hepatocyte-derived exosomes from early onset obese mice promote insulin sensitivity through miR-3075. Nat. Metab. 3, 1163–1174 (2021).
Mori, M. A., Ludwig, R. G., Garcia-Martin, R., Brandão, B. B. & Kahn, C. R. Extracellular miRNAs: from biomarkers to mediators of physiology and disease. Cell Metab. 30, 656–673 (2019).
Cariou, B., Charbonnel, B. & Staels, B. Thiazolidinediones and PPARγ agonists: time for a reassessment. Trends Endocrinol. Metab. 23, 205–215 (2012).
Paschoal, V. A. et al. Positive reinforcing mechanisms between GPR120 and PPARγ modulate insulin sensitivity. Cell Metab. 31, 1173–1188 (2020).
Colca, J. R. & Scherer, P. E. The metabolic syndrome, thiazolidinediones, and implications for intersection of chronic and inflammatory disease. Mol. Metab. 55, 101409 (2022).
Hauner, H. The mode of action of thiazolidinediones. Diabetes Metab. Res. Rev. 18, 10–15 (2002).
Kahn, C. R., Chen, L. & Cohen, S. E. Unraveling the mechanism of action of thiazolidinediones. J. Clin. Invest. 106, 1305–1307 (2000).
Xia, J. Y. et al. Acute loss of adipose tissue-derived adiponectin triggers immediate metabolic deterioration in mice. Diabetologia 61, 932–941 (2018).
Nawrocki, A. R. et al. Mice lacking adiponectin show decreased hepatic insulin sensitivity and reduced responsiveness to peroxisome proliferator-activated receptor γ agonists. J. Biol. Chem. 281, 2654–2660 (2006).
Kubota, N. et al. Pioglitazone ameliorates insulin resistance and diabetes by both adiponectin-dependent and -independent pathways. J. Biol. Chem. 281, 8748–8755 (2006).
Straub, L. G. & Scherer, P. E. Metabolic messengers: adiponectin. Nat. Metab. 1, 334–339 (2019).
Tang, W., Zeve, D., Seo, J., Jo, A.-Y. & Graff, J. M. Thiazolidinediones regulate adipose lineage dynamics. Cell Metab. 14, 116–122 (2011).
Chao, L. et al. Adipose tissue is required for the antidiabetic, but not for the hypolipidemic, effect of thiazolidinediones. J. Clin. Invest. 106, 1221–1228 (2000).
Cipolletta, D. et al. PPAR-γ is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature 486, 549–553 (2012).
Soccio, R. E., Chen, E. R. & Lazar, M. A. Thiazolidinediones and the promise of insulin sensitization in type 2 diabetes. Cell Metab. 20, 573–591 (2014).
Colca, J. R. The TZD insulin sensitizer clue provides a new route into diabetes drug discovery. Expert Opin. Drug Discov. 10, 1259–1270 (2015).
Spencer, M. et al. Pioglitazone treatment reduces adipose tissue inflammation through reduction of mast cell and macrophage number and by improving vascularity. PLoS ONE 9, e102190 (2014).
Stienstra, R. et al. Peroxisome proliferator-activated receptor γ activation promotes infiltration of alternatively activated macrophages into adipose tissue. J. Biol. Chem. 283, 22620–22627 (2008).
Yamamoto, S. et al. Atherosclerosis following renal injury is ameliorated by pioglitazone and losartan via macrophage phenotype. Atherosclerosis 242, 56–64 (2015).
Nolan, J. J., Ludvik, B., Beerdsen, P., Joyce, M. & Olefsky, J. Improvement in glucose tolerance and insulin resistance in obese subjects treated with troglitazone. N. Engl. J. Med. 331, 1188–1193 (1994).
Fonseca, V. Effect of thiazolidinediones on body weight in patients with diabetes mellitus. Am. J. Med. 115, 42–48 (2003).
Nesto, R. W. et al. Thiazolidinedione use, fluid retention, and congestive heart failure: a consensus statement from the American Heart Association and American Diabetes Association. Diabetes Care 27, 256–263 (2004).
Tontonoz, P. & Spiegelman, B. M. Fat and beyond: the diverse biology of PPARγ. Annu. Rev. Biochem. 77, 289–312 (2008).
Rohm, T. V. et al. Targeting colonic macrophages improves glycemic control in high-fat diet-induced obesity. Commun. Biol. 5, 370 (2022).
Bosch, A. J. T. et al. Lung versus gut exposure to air pollution particles differentially affect metabolic health in mice. Part. Fibre Toxicol. 20, 1–15 (2023).
Hill, D. A. et al. Distinct macrophage populations direct inflammatory versus physiological changes in adipose tissue. Proc. Natl Acad. Sci. USA 115, E5096–E5105 (2018).
Nakachi, Y. et al. Identification of novel PPARγ target genes by integrated analysis of ChIP-on-chip and microarray expression data during adipocyte differentiation. Biochem. Biophys. Res. Commun. 372, 362–366 (2008).
Daniel, B. et al. The nuclear receptor PPARγ controls progressive macrophage polarization as a ligand-insensitive epigenomic ratchet of transcriptional memory. Immunity 49, 615–626 (2018).
Li, P. et al. Adipocyte NCoR knockout decreases PPARγ phosphorylation and enhances PPARγ activity and insulin sensitivity. Cell 147, 815–826 (2011).
Hevener, A. L. et al. Macrophage PPARγ is required for normal skeletal muscle and hepatic insulin sensitivity and full antidiabetic effects of thiazolidinediones. J. Clin. Invest. 117, 1658–1669 (2007).
Tang, X. et al. Tumour-secreted Hsp90α on external surface of exosomes mediates tumour - stromal cell communication via autocrine and paracrine mechanisms. Sci. Rep. 9, 15108 (2019).
Marzec, M., Eletto, D. & Argon, Y. GRP94: an HSP90-like protein specialized for protein folding and quality control in the endoplasmic reticulum. Biochim. Biophys. Acta 1823, 774–787 (2012).
Song, M.-S., Alluin, J. & Rossi, J. J. The effect of dicer knockout on RNA interference using various dicer substrate small interfering RNA (DsiRNA) structures. Genes 13, 436 (2022).
Thum, T. et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 456, 980–984 (2008).
Krützfeldt, J. et al. Silencing of microRNAs in vivo with ‘antagomirs. Nature 438, 685–689 (2005).
Velu, C. S. & Grimes, H. L. Utilizing antagomir (antisense microRNA) to knock down microRNA in murine bone marrow cells. Methods Mol. Biol. 928, 185–195 (2012).
Kahn, B. B. Type 2 diabetes: when insulin secretion fails to compensate for insulin resistance. Cell 92, 593–596 (1998).
Donath, M. Y., Dinarello, C. A. & Mandrup-Poulsen, T. Targeting innate immune mediators in type 1 and type 2 diabetes. Nat. Rev. Immunol. 19, 734–746 (2019).
Pollack, R. M., Donath, M. Y., LeRoith, D. & Leibowitz, G. Anti-inflammatory agents in the treatment of diabetes and its vascular complications. Diabetes Care 39, S244–S252 (2016).
Raji, A., Seely, E. W., Bekins, S. A., Williams, G. H. & Simonson, D. C. Rosiglitazone improves insulin sensitivity and lowers blood pressure in hypertensive patients. Diabetes Care 26, 172–178 (2003).
Morris, D. L. et al. Adipose tissue macrophages: phenotypic plasticity and diversity in lean and obese states. Curr. Opin. Clin. Nutr. Metab. Care 14, 341–346 (2015).
Martinez, F. O. & Gordon, S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep. 6, 13 (2014).
Li, C. et al. Single cell transcriptomics based-MacSpectrum reveals novel macrophage activation signatures in diseases. JCI Insight 5, e126453 (2019).
Catrysse, L. et al. A20 deficiency in myeloid cells protects mice from diet-induced obesity and insulin resistance due to increased fatty acid metabolism. Cell Rep. 36, 109748 (2021).
Li, P. et al. Functional heterogeneity of CD11c-positive adipose tissue macrophages in diet-induced obese mice. J. Biol. Chem. 285, 15333–15345 (2010).
Gastaldelli, A. et al. Thiazolidinediones improve β-cell function in type 2 diabetic patients. Am. J. Physiol. Endocrinol. Metab. 292, 871–883 (2007).
Miyazaki, Y., Matsuda, M. & DeFronzo, R. A. Dose-response effect of pioglitazone on insulin sensitivity and insulin secretion in type 2 diabetes. Diabetes Care 25, 517–523 (2002).
Zeender, E. et al. Pioglitazone and sodium salicylate protect human β-cells against apoptosis and impaired function induced by glucose and interleukin-1β. J. Clin. Endocrinol. Metab. 89, 5059–5066 (2004).
Lupi, R. et al. Rosiglitazone prevents the impairment of human islet function induced by fatty acids: evidence for a role of PPARγ2 in the modulation of insulin secretion. Am. J. Physiol. Endocrinol. Metab. 286, 560–567 (2004).
Lin, C. Y., Gurlo, T., Haataja, L., Hsueh, W. A. & Butler, P. C. Activation of peroxisome proliferator-activated receptor-γ by rosiglitazone protects human islet cells against human islet amyloid polypeptide toxicity by a phosphatidylinositol 3′-kinase-dependent pathway. J. Clin. Endocrinol. Metab. 90, 6678–6686 (2005).
Ishida, H. et al. Pioglitazone improves insulin secretory capacity and prevents the loss of β-cell mass in obese diabetic db/db mice: possible protection of β cells from oxidative stress. Metabolism 53, 488–494 (2004).
Campbell, I. W. & Mariz, S. β-cell preservation with thiazolidinediones. Diabetes Res. Clin. Pract. 76, 163–176 (2007).
Diani, A. R., Sawada, G., Wyse, B., Murray, F. T. & Khan, M. Pioglitazone preserves pancreatic islet structure and insulin secretory function in three murine models of type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 286, 116–122 (2004).
Gao, H. et al. Accumulation of microbial DNAs promotes to islet inflammation and β cell abnormalities in obesity in mice. Nat. Commun. 13, 1–12 (2022).
Gao, H., Luo, Z., Jin, Z., Ji, Y. & Ying, W. Adipose tissue macrophages modulate obesity-associated β cell adaptations through secreted miRNA-containing extracellular vesicles. Cells 10, 6–10 (2021).
Yoshida, T., Asano, Y. & Ui-Tei, K. Modulation of microRNA processing by dicer via its associated dsRNA binding proteins. Non-coding RNA 7, 57 (2021).
Koscianska, E., Starega-Roslan, J. & Krzyzosiak, W. J. The role of Dicer protein partners in the processing of microRNA precursors. PLoS ONE 6, e28548 (2011).
Eto, K. et al. Role of NADH shuttle system in glucose-induced activation of mitochondrial metabolism and insulin secretion. Science 283, 981–985 (1999).
Xie, N. et al. NAD+ metabolism: pathophysiologic mechanisms and therapeutic potential. Signal Transduct. Target. Ther. 5, 227 (2020).
Plecitá-Hlavatá, L. et al. Mitochondrial superoxide production decreases on glucose-stimulated insulin secretion in pancreatic β cells due to decreasing mitochondrial matrix NADH/NAD+ ratio. Antioxid. Redox Signal. 33, 789–815 (2020).
Pettersson, U. S., Waldén, T. B., Carlsson, P.-O., Jansson, L. & Phillipson, M. Female mice are protected against high-fat diet induced metabolic syndrome and increase the regulatory T cell population in adipose tissue. PLoS ONE 7, e46057 (2012).
Medrikova, D. et al. Sex differences during the course of diet-induced obesity in mice: adipose tissue expandability and glycemic control. Int. J. Obes. 36, 262–272 (2012).
Ingvorsen, C., Karp, N. A. & Lelliott, C. J. The role of sex and body weight on the metabolic effects of high-fat diet in C57BL/6N mice. Nutr. Diabetes 7, e261 (2017).
Casimiro, I., Stull, N. D., Tersey, S. A. & Mirmira, R. G. Phenotypic sexual dimorphism in response to dietary fat manipulation in C57BL/6J mice. J. Diabetes Complications 35, 107795 (2021).
Mederacke, I., Dapito, D. H., Affò, S., Uchinami, H. & Schwabe, R. F. High-yield and high-purity isolation of hepatic stellate cells from normal and fibrotic mouse livers. Nat. Protoc. 10, 305–315 (2015).
Liu, X. et al. Isolation of primary human liver cells from normal and nonalcoholic steatohepatitis livers. STAR Protoc. 4, 102391 (2023).
Liu, X. et al. Primary alcohol-activated human and mouse hepatic stellate cells share similarities in gene-expression profiles. Hepatol. Commun. 4, 606–626 (2020).
Acknowledgements
This study was supported by grants from the Swiss National Science Foundation (P2BSP3_200177 to T.V.R.), the Larry L. Hillblom Foundation (2023-D-012-FEL to T.V.R., 2023-D-011-FEL to K.C.R) and by the US National Institute of Diabetes and Digestive and Kidney Diseases (P30DK063491 and R01DK101395 to J.M.O., DK124298 to Y.S.L.), the US National Institutes of Health awards (R00DK115998, R21HD107516, and R01DK125560 to W.Y.), UCLA LIFT-UP (Leveraging Institutional support for Talented, Underrepresented Physicians and/or Scientists to K.C.R.), the National Institutes of Health (DK099205, AA028550, DK101737, AA011999, DK120515, AA029019, DK091183 to T. Kisseleva) and a grant from Janssen Pharmaceuticals (to J.M.O.). Work at the Center for Epigenomics was supported by the University of California San Diego School of Medicine. We also thank T. Kisseleva and H. Y. Kim from the Department of Surgery, University of California San Diego School of Medicine for providing human hepatocytes, A. De Maio, D. M. Cauvi and D. Hawisher for sharing their tabletop ultracentrifuge and O. Osborn for advice and discussions. Additionally, we thank J. Park, M. Paszek, D. Zhang, J. Moon, I. Jeelani and K. Wang for their technical contributions and for sharing collected tissues. We also thank J. Pimentel and J. Pferdekamper for organizational support.
Author information
Authors and Affiliations
Contributions
T.V.R. and J.M.O. designed the experimental studies. T.V.R. performed all of the experiments. F.C.G.R. supported the BMDM-sEV studies by culturing and isolating BMDM-sEVs. F.C.G.R. and C. Murphy also helped to sacrifice in vivo cohorts and collect key organs, such as the adipose tissue, liver and muscle. R.I. performed liver perfusions. G.B. assisted in isolating liver NPCs and hepatocytes and in performing glucose production assays. H.G. and K.C.R. participated in conducting insulin injections into the vena cava to study in vivo AKT phosphorylation in insulin target tissues. K.C.R. also supported the miR-690 mimic and antagomir studies. C. Miciano and A.W. performed the scRNA-seq analysis. C. Murphy assisted in cell culture and in vivo animal studies, as well as performed PCRs and sEV isolations. A.M.L. assisted with feeding mice and collecting blood and tissues. R.C.Z. performed the MILLIPLEX Mouse Adipocytes Magnetic Bead Assay. All data analyses were performed by T.V.R. and the scRNA-seq analyses were conducted by C. Miciano and A.W. All figures were designed by T.V.R. T.V.R. and J.M.O. interpreted the data and co-wrote the paper. W.Y. and Y.S.L. gave key support in interpreting data and experimental design. T.V.R., F.C.G.R., R.C.Z., R.I., C. Murphy, K.C.R., W.Y., C. Miciano, A.W., Y.S.L., A.M.L. and J.M.O. edited the paper.
Corresponding authors
Ethics declarations
Competing interests
W.Y. and J.M.O. are co-investigators on a provisional patent covering the use of miR-690 as an insulin sensitizer. The other authors declare no competing interests.
Peer review
Peer review information
Nature Metabolism thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: Yanina-Yasmin Pesch, in collaboration with the Nature Metabolism team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Rosiglitazone reduces adipose tissue inflammation and increases the eosinophil percentage in both epididymal (eWAT) and subcutaneous (SubQ) fat.
WT mice were fed HFD for 4 months and treated for the last month with 3 mg kg-1 d-1 rosiglitazone (Rosi) compared to HFD and Chow control mice: a, Epididymal and subcutaneous fat mass (Chow, n = 22, HFD,n = 22, Rosi, n = 23, ***p < 0.0001). b, Number of sorted live (DAPI neg) CD45+CD11b+F4/80+ epidydimal (eWAT) ATMs per mouse (n = 30,40,27; Chow versus Rosi, P = 0.0083, Chow versus HFD, P < 0.0001,HFD versus Rosi, P = 0.0001). c, Percentage and absolute numbers of CD11b+SiglecF+ eWAT eosinophils (%, n = 21, #, n = 19,18,19; ***P = < 0.001, *P = 0.0325). d, e, Percentage and absolute numbers per gram (No. per g) of total subcutaneous adipose tissue macrophages (SubQ ATMs) (n = 25,28,26, %: Chow versus Rosi/HFD, P < 0.001, HFD versus Rosi, P = 0.0006, #: Chow versus Rosi, P = 0.0012, Chow versus HFD, P = 0.0147, HFD versus Rosi, P < 0.0001) (d) and ATM subsets: Double negative (DN), CD11c+CD206− monocyte-derived, CD11c+CD206low ‘M1-like’ and CD11c−CD206high ‘M2-like’ ATMs (Chow, n = 25, HFD, n = 28, Rosi, n=29, *P = 0.018, ***P < 0.0001) (e). f, g, Percentage and absolute numbers of CD9+ (%: *P = 0,0180,***P < 0.0001, #: *P = 0.0116, ***P < 0.0001) (f) or CD9+Trem2+ SubQ lipid-associated macrophages (LAMs, %: **P = 0.0034,***P < 0.0001, #: *P = 0.0130, ***P < 0.0001) (g) (n = 25,28,26). h, Percentage and absolute numbers of CD11b+SiglecF+ SubQ adipose tissue eosinophils (n = 25,28,26, %: Rosi versus Chow/HFD, P < 0.0001, Chow versus HFD, P = 0.0012, #: *P = 0.0143). Statistical data are expressed as mean ± s.e.m., with each data point representing a biologically independent mouse from three independent cohorts. *P < 0.05, **P < 0.01, ***P < 0.00, with one-way ANOVA, followed by Tukey′s multiple comparisons tests.
Extended Data Fig. 2 Rosiglitazone reduces pro-inflammatory cytokine gene expression and increases PPARy target and anti-inflammatory gene expression.
a, b, Relative gene expression of Pparg and its target gene Cd36 (*p = 0.0258, **p = 0.0037), ‘M2-like’ macrophage marker genes (Mrc1, Mgl1, *P = 0.0301,0.0232, **P = 0.0018) (a) and ‘M1-like’ macrophage marker genes (Tnf, Il1b, Infg, iNOS, *P = 0.0152, **P = 0.0065,0.0096) (b) in eWAT ATMs from Chow, HFD, and Rosi mice. c, d, Geometric mean fluorescence intensity (gMFI) of the M2-marker CD206 (P = 0.0043) (c) and the relative gene expression of the M2-marker gene Mgl1 (P < 0.0001), Pparg (P = 0.0008) and its target gene Cd36 (P = 0.0004), and M1-marker genes (Infg, Il1b, *P = 0.0350, **P = 0.0032) (d) in Rosi-treated M1-BMDMs compared to untreated M1-BMDMs. e, gMFI of CD206 (P = 0.0003) and Trem2 (P = 0.0286), and the percentage of Trem2 (P = 0.0286) from Rosi-treated M0 or M2-BMDMs compared to untreated M0- and M2-BMDMs. Statistical data are expressed as mean ± s.e.m., with each data point representing a biologically independent cell replicate, from three independent cohorts (a,b) or representative of one (e) or three (c,d) independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, with one-way ANOVA, followed by Tukey′s multiple comparisons tests (a,b,e) or an unpaired Mann-Whitney U-test with two-tailed distribution (c and d).
Extended Data Fig. 3 Rosiglitazone increases liver Kupffer cells and eosinophils.
WT mice were fed HFD for 4 months, with the last month on 3 mg kg-1 d-1 rosiglitazone (Rosi) compared to 4 months HFD and Chow control mice: a, Percentage and absolute numbers (No.) of CD11b+F4/80low recruited hepatic macrophages (RHMs, *P = 0.0257, ***P < 0.0001). b, Percentage, absolute numbers, and geometric mean fluorescence intensity (gMFI) of Clec4F from CD11b+F4/80high Kupffer cells (KCs, %, Rosi versus HFD, P = 0.0029, #, ***P < 0.0001, gMFI, *P = 0.0222, ***P = 0.0006). c-e, Percentage and absolute numbers of Clec4F+F4/80+ KCs (%, Rosi versus HFD, P = 0.0137, #, Chow versus Rosi, P = 0.0035, HFD versus Rosi, P = 0.0030) (c), Trem2+ KCs (%, Rosi versus HFD, P = 0.0185, #, **P = 0.0035, ***P = 0.0002) (d), and CD11c+ KCs (%, *P = 0.0217, Chow versus Rosi, P = 0.009, Chow versus HFD, P < 0.0001) (e). f, Percentage and absolute numbers of CD11c+CD9+ KCs and gMFI of CD9 from CD11b+F4/80high KCs (%, *P = 0.0055, **P = 0.0075, ***P < 0.0001, #, Chow versus HFD, P = 0.0003, Chow versus Rosi, P < 0.0001, gMFI, **P = 0.0035, Chow versus Rosi, P = 0.0003, Chow versus HFD, P = < 0.0001). g, Percentage and absolute numbers of Siglec F+ liver eosinophils (%, Chow versus HFD, P = 0.0008, HFD versus Rosi, P = 0.0038, ***P < 0.0001, # Chow versus HFD, P = 0.0003, HFD versus Chow/Rosi, P < 0.0001). Statistical data are expressed as mean ± s.e.m., with each data point representing a biologically independent mouse from two (gMFI and #, n = 8 per group) or three (%, n = 10 per group) independent cohorts. *P < 0.05, **P < 0.01, ***P < 0.00, while comparing only Rosi versus. HFD with an unpaired Mann-Whitney U-test with two-tailed distribution or comparing all three groups with one-way ANOVA, followed by Tukey′s multiple comparisons tests.
Extended Data Fig. 4 Cluster analysis of stromal vascular cells.
Single-cell RNA sequencing of epidydimal stromal vascular cells from WT 4 months HFD/obese mice treated for the last month with 3 mg kg-1 d-1 rosiglitazone (Rosi) compared to HFD control mice: a, Heat map showing scaled counts per million expression values of marker genes for broad clusters: macrophages, DCs, T cells, B cells, mast cells, endothelial cells, and adipocyte progenitors. b, Heat map showing genes involved in PPAR signalling pathways of broad macrophage cluster. c, d, Up- (c) or downregulated (d) GO Biological Process 2021 pathways after Rosi treatment compared to HFD for the macrophage cluster (FDR< = 0.1). Point sizes reflect the combined EnrichR score, and the colour represents the P-value for each annotated pathway analysed by two-tailed Wilcoxon rank-sum test. e, Heat map showing scaled counts per million expression values of macrophage subpopulation marker genes in each subcluster: Monocyte-derived (Mac5), ‘M1-like’ (Mac3), lipid-associated (LAMs/Mac2), and ‘M2-like’ ATMs (Mac1 and Mac4). Data are representative of three biologically independent mice from one experiment/cohort (n = 3 per group).
Extended Data Fig. 5 Rosi treatment alters the transcriptional profile of ATM subpopulations.
ScRNA-seq of epidydimal stromal vascular cells (SVCs) from WT 4 months HFD/obese mice treated for the last month with 3 mg kg-1 d-1 rosiglitazone (Rosi) compared to HFD control mice. a, Percentage of ATM subclusters, including ‘M1-like’ (Mac3) and ‘M2-like’ (Mac1 and Mac4) ATMs, of the total macrophage cluster. b, Volcano plots showing differential expression between Rosi diet and HFD for each ATM subcluster: Monocyte-derived macrophages (Mac5), ‘M1-like’ ATMs (Mac3), lipid-associated (LAMs/Mac2), and ‘M2-like’ ATMs (Mac1 and Mac4). Up- or downregulated genes, when significant at 0.05 % FDR threshold and absolute log2 fold change >0.58, analysed by two-tailed Wilcoxon rank-sum test. c, Up- (left) or downregulated (right) GO Biological Process 2021 pathways after Rosi treatment, compared to HFD for each macrophage subcluster (FDR< = 0.1). Point sizes reflect the combined EnrichR score, and the colour represents the P-value for each annotated pathway analysed by two-tailed Wilcoxon rank-sum test. Data are representative of three biologically independent mice from one experiment/cohort (n = 3 per group). Data (a) show mean ± s.e.m., and brackets indicate statistical significance; otherwise, nonsignificant (NS.) is stated. ‘log10(P)’’ is the enrichment P-value in log base 10.
Extended Data Fig. 6 Rosi causes the secretion of a greater number of smaller sEVs from ATMs.
a, Protein expression of exosome-associated markers (Alix, CD63, CD9, CD81) compared to heat shock protein HSP90 and the lysate control glucose-regulated protein 94 (Grp94). b, Mean sEV size of Rosi-ATM-sEVs compared to HFD- and Chow-ATM-sEVs and ratio of particle concentration and sorted CD11b+F4/80+ ATMs of visceral (Vis, size, P = 0.0051, ratio, *P = 0.0477, **P = 0.0043), epididymal (eWAT, size, P = 0.0399, ratio, P = 0.0344), or subcutaneous (SubQ, size, P = 0.0207, ratio, P = 0.0251) fat. c, NanoSight analysis of ATM-sEVs derived from Chow, HFD, and Rosi mice. d, NanoSight analysis of BMDM-sEVs derived from Rosi-treated M1-BMDMs compared to untreated M1, M2, and M0 BMDMs. Statistical data are expressed as mean ± s.e.m., with each data point (b) showing a biologically independent sEV sample replicate from three independent cohorts. *P < 0.05, **P < 0.01, with one-way ANOVA, followed by Tukey′s multiple comparisons tests.
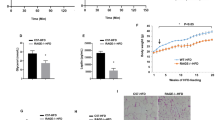
Extended Data Fig. 7 Rosi-ATM-sEVs enhance AKT phosphorylation in all insulin target tissues and reduce pro-inflammatory ATMs.
Obese mice fed 10 weeks HFD were treated for 4 weeks with Rosi-ATM-sEVs (Rosi-ATM-sEVs), HFD-ATM-sEVs, or PBS Ctrl liposomes compared to Rosi diet: a, Insulin-stimulated AKT phosphorylation in the epididymal adipose tissue (eWAT) and liver. b, Protein expression of PPARy (descending P = 0.0170,0.0164) and its inhibitory phosphorylation sites S273 (*P = 0.0342, descending ***P = < 0.0001,0.0002,0.0001) and S112 (*P = 0.0448, **P = 0.0017, descending ***P = 0.006, < 0.0001) in the eWAT. c-e, Absolute numbers of eWAT ATMs per gram (*P = 0.0340, ***P = 0.0007) (c) and their subpopulations: Double negative (DN), CD11c+CD206− monocyte-derived macrophages (p = 0.0010), CD11c+CD206low ‘M1-like’ (**P = 0.0020, ***P<0.0001), CD11c-CD206high ‘M2-like’ ATMs (d), and CD9+ (CD9+, descending P = 0.0179,0.0001,0.0178, Trem2-CD9+, PBS versus HFD-sEVs, P = 0.0015, HFD-sEVs versus Rosi-sEVs, P = 0.0015, ***P < 0.0001) or CD9+Trem2+ lipid-associated macrophages (LAMs, *P = 0.0387, **P = 0.0011) (e). f, Circulating leptin levels (P = 0.0301). g, Plasma particle concentration of Rosi-ATM-sEV-treated obese mice compared to Rosi mice measured by NanoSight analysis. Statistical data are expressed as mean ± s.e.m., with each data point representing a biologically independent mouse from one cohort. *P < 0.05, **P < 0.01, ***P < 0.001, with one-way ANOVA, followed by Tukey′s multiple comparisons tests (c-f) or an unpaired Mann-Whitney U-test with two-tailed distribution (g).
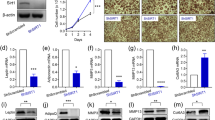
Extended Data Fig. 8 Rosi-HFD eWAT explant-sEVs reverse insulin resistance caused by HFD eWAT explant-sEVs in L6 myotubes in vitro.
a, Glucose uptake in L6 myotubes treated with sEVs isolated from HFD epididymal adipose tissue (eWAT) explants after in vitro treatment with 10 µM Rosi compared to HFD-sEVs or PBS Ctrl liposomes (*P = 0.0378, **P = 0.0028). b,c, Effects of in vitro Rosi treatment on eWAT explants isolated from obese mice on gene expression of Pparg and its target gene Cd36 (P = 0.0190) (b), the ‘M2-like’ macrophage marker gene Mgl1 (P = 0.0303) and miR-690 (c). Statistical data are expressed as mean ± s.e.m., with each data point showing a biologically independent cell replicate, representative of an independent experiment. *P < 0.05, **P < 0.01, ***P < 0.001, with 2-way ANOVA, followed by Tukey′s multiple comparisons tests (a) or an unpaired Mann-Whitney U-test with two-tailed distribution (b,c).
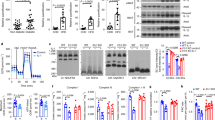
Extended Data Fig. 9 Enrichment of miR-690 and downregulation of Nadk in Rosi-macrophages is dependent on PPARy activation.
a, Relative gene expression of miR-690 in CD11c+CD206low ‘M1-like’ ATMs (P = 0.0499) or CD11c-CD206high ‘M2-like’ (P = 0.0037) Rosi-ATMs compared to HFD-ATMs. b, Effects of Rosi-sEVs isolated from ‘M1-like’ or ‘M2-like’ Rosi-ATMs on glucose uptake in L6 myotubes compared to HFD-ATM-sEVs or PBS Ctrl liposomes (*P = 0.0339, **P = 0.0035, descending ***P = < 0.0001, 0.0003). c,d, Relative gene expression of miR-690 (M1 versus M1+Rosi, P = 0.0070, M1+Rosi versus M1+Rosi PPARy KD, P = 0.0090), its target gene Nadk (P = 0.0039,0.0353), and Pparg (P = 0.0258,0.0106), Cd36 (P = 0.0028,0.0126), as well as M2-macrophage marker genes (Mrc1, P = 0.006, < 0.0001, Mgl1, P = 0.0002,0.0037) (c) and M1-marker genes (Tnf, Il1b, *P = 0.0145, ***P = 0.0005 Infg, P = 0.0161, iNos, *P = 0.0113, **P = 0.0046) (d) in PPARy-deficient Rosi-treated M1 (M1+Rosi PPARy KD) BMDMs compared to Rosi-treated and untreated M1-BMDMs. e, Relative gene expression of miR-690 (descending p = 0.0010,< 0.0001,0.0001,0.0012,< 0.0001,0.0001) and Nadk (*P = 0.0287, descending ***P = 0.0004, < 0.0001) in L6 myotubes treated with M1+Rosi BMDM-sEVs compared to PBS Ctrl liposomes or M0-/M1-/M2-BMDM-sEV-treated L6 cells. f, Plasma miR-690 expression in Rosi-ATM-sEVs- or Rosi-treated mice and liver miR-690 (descending P = 0.0002, 0.0010, <0.0001, 0.0005) or Nadk (P = 0.0476,0.0488) expression in Rosi-ATM-sEVs-, HFD-ATM-sEVs-, PBS Ctrl liposomes-, or Rosi-treated mice. g, Relative gene expression of miR-690 (P < 0.0001) and Nadk (P = 0.0257,0.0481) in HFD islets treated with Rosi-ATM-sEVs compared to PBS Ctrl liposomes with and without miR-690 mimic, co-treatment of Rosi-ATM-sEVs and miR-690 antagomir, or Rosi-Dicer-KD-ATM-sEV treatment alone. Statistical data are expressed as mean ± s.e.m., with each data point representing a biologically independent cell replicate from one cohort (a,f), or representative of one (b–d) or two (g) independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, with an unpaired Mann-Whitney U-test with two-tailed distribution (a), with 2-way ANOVA, followed by Tukey′s multiple comparisons tests (b) or one-way ANOVA, followed by Tukey′s multiple comparisons tests (c-g).
Extended Data Fig. 10 miR-690 antagomir treatment does not induce changes in body weight or haematocrit levels but enhances AKT phosphorylation and regulates miR-690, Nadk expression in insulin target tissues.
Obese mice were fed with HFD for 10 weeks and were then treated for 4 weeks with 3 mg kg-1 d-1 rosiglitazone (Rosi) and with miR-690 antagomir compared to Rosi, HFD, and Chow control mice: a, Effects of miR-690 antagomir treatment on insulin-stimulated AKT phosphorylation in muscle and liver. b, Relative gene expression of miR-690 (Chow versus Rosi+miR-690 antagomir, P = 0.0107, Rosi versus Rosi+miR-690 antagomir, P = 0.0445) and its target Nadk (**P = 0.0055, *P = 0.0164) and NAD+ levels (P = 0.0408,0.0184) in the liver. c, miR-690 and Nadk expression in muscle (miR-690, P = 0.0388, Nadk, descending P = 0.0399,0.0181,0.0367,0.0174) and pancreas (miR-690, *P = 0.0432, **P = 0.0052, Nadk, P = 0.0391). d, Body weight (**P = 0.0039, ***P < 0.0001), haematocrit (descending P = 0.0019, 0.0016), and total cholesterol levels (values detected as < 100 are shown as 100, *P < 0.0001, ***P = < 0.0001, 0.0009) in the plasma. Statistical data are expressed as mean ± s.e.m., with each data point representing a biologically independent mouse from one cohort. Data is representative of one cohort. *P < 0.05, **P < 0.01, ***P < 0.001, with one-way ANOVA, followed by Tukey′s multiple comparisons tests.
Supplementary information
Supplementary Information
Supplementary Figs. 1 and 2 and Tables 1–3.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 3
Unprocessed western blots.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 4
Unprocessed western blots.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 5
Unprocessed western blots.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Statistical source data.
Source Data Fig. 8
Statistical source data.
Source Data Fig. 8
Unprocessed western blots.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 6
Unprocessed western blots.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 7
Unprocessed western blots.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 10
Statistical source data.
Source Data Extended Data Fig. 10
Unprocessed western blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Rohm, T.V., Castellani Gomes Dos Reis, F., Isaac, R. et al. Adipose tissue macrophages secrete small extracellular vesicles that mediate rosiglitazone-induced insulin sensitization. Nat Metab (2024). https://doi.org/10.1038/s42255-024-01023-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42255-024-01023-w
This article is cited by
-
Macrophage vesicles in antidiabetic drug action
Nature Metabolism (2024)
