
Abstract
Adipose tissue lipolysis is mediated by cAMP–protein kinase A (PKA)-dependent intracellular signalling. Here, we show that PKA targets p21-activated kinase 4 (PAK4), leading to its protein degradation. Adipose tissue-specific overexpression of PAK4 in mice attenuates lipolysis and exacerbates diet-induced obesity. Conversely, adipose tissue-specific knockout of Pak4 or the administration of a PAK4 inhibitor in mice ameliorates diet-induced obesity and insulin resistance while enhancing lipolysis. Pak4 knockout also increases energy expenditure and adipose tissue browning activity. Mechanistically, PAK4 directly phosphorylates fatty acid-binding protein 4 (FABP4) at T126 and hormone-sensitive lipase (HSL) at S565, impairing their interaction and thereby inhibiting lipolysis. Levels of PAK4 and the phosphorylation of FABP4-T126 and HSL-S565 are enhanced in the visceral fat of individuals with obesity compared to their lean counterparts. In summary, we have uncovered an important role for FABP4 phosphorylation in regulating adipose tissue lipolysis, and PAK4 inhibition may offer a therapeutic strategy for the treatment of obesity.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
The source data for all bars and line graphs as well as the original uncropped western blots can be found in the online source data file. Raw and processed RNA-seq datasets were deposited into the NCBI GEO database under the accession number GSE214769. Source data are provided with this paper.
Code availability
No custom code was used for this study.
References
Grabner, G. F. et al. Lipolysis: cellular mechanisms for lipid mobilization from fat stores. Nat. Metab. 3, 1445–1465 (2021).
Blondin, D. P. et al. Human brown adipocyte thermogenesis is driven by β2-AR stimulation. Cell Metab. 32, 287–300 (2020).
Collins, S. β-adrenergic receptors and adipose tissue metabolism: evolution of an old story. Annu Rev. Physiol. 84, 1–16 (2022).
Zimmermann, R. et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science 306, 1383–1386 (2004).
Miyoshi, H. et al. Control of adipose triglyceride lipase action by serine 517 of perilipin A globally regulates protein kinase A-stimulated lipolysis in adipocytes. J. Biol. Chem. 282, 996–1002 (2007).
Watt, M. J. et al. Regulation of HSL serine phosphorylation in skeletal muscle and adipose tissue. Am. J. Physiol. Endocrinol. Metab. 290, E500–E508. (2006).
Garton, A. J. et al. Phosphorylation of bovine hormone-sensitive lipase by the AMP-activated protein kinase. A possible antilipolytic mechanism. Eur. J. Biochem. 179, 249–254 (1989).
Greenberg, A. S. et al. Stimulation of lipolysis and hormone-sensitive lipase via the extracellular signal-regulated kinase pathway. J. Biol. Chem. 276, 45456–45461 (2001).
Furuhashi, M. Fatty acid-binding protein 4 in cardiovascular and metabolic diseases. J. Atheroscler. Thromb. 26, 216–232 (2019).
Hotamisligil, G. S. & Bernlohr, D. A. Metabolic functions of FABPs–mechanisms and therapeutic implications. Nat. Rev. Endocrinol. 11, 592–605 (2015).
Xu, A. et al. Circulating adipocyte-fatty acid binding protein levels predict the development of the metabolic syndrome: a 5-year prospective study. Circulation 115, 1537–1543 (2007).
Yeung, D. C. et al. Serum adipocyte fatty acid-binding protein levels were independently associated with carotid atherosclerosis. Arterioscler Thromb. Vasc. Biol. 27, 1796–1802 (2007).
Smith, A. J., Sanders, M. A., Juhlmann, B. E., Hertzel, A. V. & Bernlohr, D. A. Mapping of the hormone-sensitive lipase binding site on the adipocyte fatty acid-binding protein (AFABP). Identification of the charge quartet on the AFABP/aP2 helix-turn-helix domain. J. Biol. Chem. 283, 33536–33543 (2008).
Shen, W. J., Sridhar, K., Bernlohr, D. A. & Kraemer, F. B. Interaction of rat hormone-sensitive lipase with adipocyte lipid-binding protein. Proc. Natl Acad. Sci. USA 96, 5528–5532 (1999).
Mita, T. et al. FABP4 is secreted from adipocytes by adenyl cyclase-PKA- and guanylyl cyclase-PKG-dependent lipolytic mechanisms. Obesity 23, 359–367 (2015).
Won, S. Y., Park, J. J., Shin, E. Y. & Kim, E. G. PAK4 signaling in health and disease: defining the PAK4-CREB axis. Exp. Mol. Med. 51, 1–9 (2019).
Park, M. H. et al. p21-activated kinase 4 promotes prostate cancer progression through CREB. Oncogene 32, 2475–2482 (2013).
Costa, T. D. F. et al. PAK4 suppresses RELB to prevent senescence-like growth arrest in breast cancer. Nat. Commun. 10, 3589 (2019).
Siu, M. K. et al. p21-activated kinase 4 regulates ovarian cancer cell proliferation, migration, and invasion and contributes to poor prognosis in patients. Proc. Natl Acad. Sci. USA 107, 18622–18627 (2010).
Liu, Y. et al. The pak4 protein kinase plays a key role in cell survival and tumorigenesis in athymic mice. Mol. Cancer Res. 6, 1215–1224 (2008).
Mao, Y. et al. p21-activated kinase 4 inhibition protects against liver ischemia/reperfusion injury: role of nuclear factor erythroid 2-related factor 2 phosphorylation. Hepatology 76, 345–356 (2022).
Mao, Y. et al. p21-activated kinase 4 phosphorylates peroxisome proliferator-activated receptor γ and suppresses skeletal muscle regeneration. J. Cachexia Sarcopenia Muscle 12, 1776–1788 (2021).
Shi, M. Y. et al. p21-activated kinase 4 suppresses fatty acid β-oxidation and ketogenesis by phosphorylating NCoR1. Nat. Commun. 14, 4987 (2023).
Stralfors, P., Bjorgell, P. & Belfrage, P. Hormonal regulation of hormone-sensitive lipase in intact adipocytes: identification of phosphorylated sites and effects on the phosphorylation by lipolytic hormones and insulin. Proc. Natl Acad. Sci. USA 81, 3317–3321 (1984).
Schlottmann, I., Ehrhart-Bornstein, M., Wabitsch, M., Bornstein, S. R. & Lamounier-Zepter, V. Calcium-dependent release of adipocyte fatty acid binding protein from human adipocytes. Int J. Obes. 38, 1221–1227 (2014).
Ertunc, M. E. et al. Secretion of fatty acid binding protein aP2 from adipocytes through a nonclassical pathway in response to adipocyte lipase activity. J. Lipid Res. 56, 423–434 (2015).
Cao, H. et al. Adipocyte lipid chaperone AP2 is a secreted adipokine regulating hepatic glucose production. Cell Metab. 17, 768–778 (2013).
Wu, L. E. et al. Identification of fatty acid binding protein 4 as an adipokine that regulates insulin secretion during obesity. Mol. Metab. 3, 465–473 (2014).
Garton, A. J. & Yeaman, S. J. Identification and role of the basal phosphorylation site on hormone-sensitive lipase. Eur. J. Biochem. 191, 245–250 (1990).
Li, Y. et al. Nucleo-cytoplasmic shuttling of PAK4 modulates β-catenin intracellular translocation and signaling. Biochim. Biophys. Acta 1823, 465–475 (2012).
Shen, W. J. et al. Characterization of the functional interaction of adipocyte lipid-binding protein with hormone-sensitive lipase. J. Biol. Chem. 276, 49443–49448 (2001).
Hofer, P. et al. Fatty acid-binding proteins interact with comparative gene identification-58 linking lipolysis with lipid ligand shuttling. J. Biol. Chem. 290, 18438–18453 (2015).
Coe, N. R., Simpson, M. A. & Bernlohr, D. A. Targeted disruption of the adipocyte lipid-binding protein (aP2 protein) gene impairs fat cell lipolysis and increases cellular fatty acid levels. J. Lipid Res. 40, 967–972 (1999).
Scheja, L. et al. Altered insulin secretion associated with reduced lipolytic efficiency in aP2−/− mice. Diabetes 48, 1987–1994 (1999).
London, E., Bloyd, M. & Stratakis, C. A. PKA functions in metabolism and resistance to obesity: lessons from mouse and human studies. J. Endocrinol. 246, R51–R64 (2020).
Iankova, I. et al. Regulator of G protein signaling-4 controls fatty acid and glucose homeostasis. Endocrinology 149, 5706–5712 (2008).
Sohn, J. H. et al. Perilipin 1 (Plin1) deficiency promotes inflammatory responses in lean adipose tissue through lipid dysregulation. J. Biol. Chem. 293, 13974–13988 (2018).
Han, C. Y. et al. Differential effect of saturated and unsaturated free fatty acids on the generation of monocyte adhesion and chemotactic factors by adipocytes: dissociation of adipocyte hypertrophy from inflammation. Diabetes 59, 386–396 (2010).
Girousse, A. et al. Partial inhibition of adipose tissue lipolysis improves glucose metabolism and insulin sensitivity without alteration of fat mass. PLoS Biol. 11, e1001485 (2013).
Schoiswohl, G. et al. Impact of reduced ATGL-mediated adipocyte lipolysis on obesity-associated insulin resistance and inflammation in male mice. Endocrinology 156, 3610–3624 (2015).
Haemmerle, G. et al. Defective lipolysis and altered energy metabolism in mice lacking adipose triglyceride lipase. Science 312, 734–737 (2006).
Ahmadian, M. et al. Desnutrin/ATGL is regulated by AMPK and is required for a brown adipose phenotype. Cell Metab. 13, 739–748 (2011).
Shin, H. et al. Lipolysis in brown adipocytes is not essential for cold-induced thermogenesis in mice. Cell Metab. 26, 764–777 (2017).
Haemmerle, G. et al. ATGL-mediated fat catabolism regulates cardiac mitochondrial function via PPAR-α and PGC-1. Nat. Med. 17, 1076–1085 (2011).
Mottillo, E. P., Bloch, A. E., Leff, T. & Granneman, J. G. Lipolytic products activate peroxisome proliferator-activated receptor (PPAR) α and δ in brown adipocytes to match fatty acid oxidation with supply. J. Biol. Chem. 287, 25038–25048 (2012).
Markussen, L. K. et al. Lipolysis regulates major transcriptional programs in brown adipocytes. Nat. Commun. 13, 3956 (2022).
Buelt, M. K., Shekels, L. L., Jarvis, B. W. & Bernlohr, D. A. In vitro phosphorylation of the adipocyte lipid-binding protein (p15) by the insulin receptor. Effects of fatty acid on receptor kinase and substrate phosphorylation. J. Biol. Chem. 266, 12266–12271 (1991).
Hresko, R. C., Hoffman, R. D., Flores-Riveros, J. R. & Lane, M. D. Insulin receptor tyrosine kinase-catalyzed phosphorylation of 422(aP2) protein. Substrate activation by long-chain fatty acid. J. Biol. Chem. 265, 21075–21085 (1990).
Prentice, K. J., Saksi, J. & Hotamisligil, G. S. Adipokine FABP4 integrates energy stores and counterregulatory metabolic responses. J. Lipid Res. 60, 734–740 (2019).
Huang, C. L. et al. Serum adipocyte fatty acid-binding protein levels in patients with critical illness are associated with insulin resistance and predict mortality. Crit. Care 17, R22 (2013).
Nowak, C. et al. Protein biomarkers for insulin resistance and type 2 diabetes risk in two large community cohorts. Diabetes 65, 276–284 (2016).
Baar, R. A. et al. Investigation of in vivo fatty acid metabolism in AFABP/aP2−/− mice. Am. J. Physiol. Endocrinol. Metab. 288, E187–E193 (2005).
Anthony, N. M., Gaidhu, M. P. & Ceddia, R. B. Regulation of visceral and subcutaneous adipocyte lipolysis by acute AICAR-induced AMPK activation. Obesity 17, 1312–1317 (2009).
Large, V. et al. Decreased expression and function of adipocyte hormone-sensitive lipase in subcutaneous fat cells of obese subjects. J. Lipid Res. 40, 2059–2066 (1999).
Martin, L. F. et al. Alterations in adipocyte adenylate cyclase activity in morbidly obese and formerly morbidly obese humans. Surgery 108, 228–234 (1990).
Virtue, S. & Vidal-Puig, A. GTTs and ITTs in mice: simple tests, complex answers. Nat. Metab. 3, 883–886 (2021).
Jenkins, A. E., Hockenberry, J. A., Nguyen, T. & Bernlohr, D. A. Testing of the portal hypothesis: analysis of a V32G, F57G, K58G mutant of the fatty acid binding protein of the murine adipocyte. Biochemistry 41, 2022–2027 (2002).
Jang, H. Y. et al. Sirtuin 6 is a negative regulator of FcepsilonRI signaling and anaphylactic responses. J. Allergy Clin. Immunol. 149, 156–167 (2022).
Shi, M. Y. et al. Statin suppresses sirtuin 6 through miR-495, increasing FoxO1-dependent hepatic gluconeogenesis. Theranostics 10, 11416–11427 (2020).
Acknowledgements
This work was supported by grants from the Medical Research Center Program (grant no. 2017R1A5A2015061), Bio & Medical Technology Development Program (grant no. 2022M3E5F2017607), Korea Drug Development Fund (grant no. HN21C0447) and Basic Science Research Program (grant no. 2023R1A2C3002389), all of which were funded by the Korean government. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
H. Chan Yu, Y.G.J. and C.Y.H. performed the in vitro and in vivo experiments. M.R.L., J.D.Y. and H. Chul Yu performed human tissue analysis. J.B.S. and N.D.K. synthesized the PAK4 inhibitor. A.Y.N. and S.L. performed the LC–MS/MS and phosphoproteomic analyses. J.B.K. interpreted the data. E.J.B. and B.-H.P. designed the experiments, interpreted the data and wrote the manuscript. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Metabolism thanks Masato Furuhashi, Dominique Langin and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Yanina-Yasmin Pesch, in collaboration with the Nature Metabolism team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
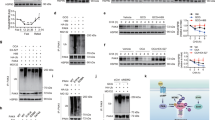
Extended Data Fig. 1 Proteasomal degradation of PAK4 by PKA activation.
a. 3T3-L1 adipocytes were transfected with catalytic subunit of PKA (His-PKA Cα) in combination with PAK4 on day 4 post-MDI treatment. Seven days after MDI treatment, PKA binding with PAK4 in the presence of 3 μM MG132 was analysed by Co-IP assay. b. Recombinant PAK4 was incubated with recombinant PKA and [32P]ATP for 30 min, and the proteins in the mixture were resolved by SDS-PAGE. The band was visualized by autoradiography of 32P-labelled protein. Protein loading was confirmed by Coomassie blue staining. c. 3T3-L1 adipocytes transfected with PKA and PAK4 were treated with isoproterenol (1 μM) for 1 h. Phosphorylation of PAK4 was determined in PAK4 immunoprecipitates using LC-MS/MS analysis after in-gel digestion. Representative spectra are shown. d. 3T3-L1 adipocytes were transfected with siRNA targeting various E3 ubiquitin ligases, followed by treatment with isoproterenol (1 μM) for 1 h. Protein levels of PAK4 were analyzed by Western blotting (left), while mRNA levels were assessed using qPCR (n = 7). Data are representative of at least three independent experiments. Values are mean ± SD. Unpaired two-tailed t-test between two groups was conducted for statistical analyses (d). Source data are provided as a Source Data file. NTC, no template control.
Extended Data Fig. 2 Increase of adaptive thermogenesis in male adipocyte Pak4 KO mice after cold exposure.
a. Eight-week-old male adipocyte Pak4 KO mice and their wild-type (WT) littermates were housed at 30 °C for 1 day and then exposed to cold (6 °C). The rectal temperature of mice was measured at indicated time points after cold exposure (n = 5). b. Representative infrared image showing Pak4 KO and WT mice (upper panel) and gross morphology of EAT, IAT and BAT (lower panel) housed at 6 °C for three days. c. Weights of EAT, IAT and BAT are presented as percentages of body weight (BW) (n = 4). d. IAT and BAT were H&E stained or immunostained with an antibody against UCP1. Bars=50 μm. e. Protein levels of thermogenic markers and phosphorylation levels of lipolysis-related proteins were determined by Western blotting. f. Representative transmission electron micrographs of IAT after 3-day cold exposure. Numbers (n = 6) and sizes (n = 21) of mitochondria were determined from images. g. Western blot analysis of OxPhos complex in IAT (n = 6). Data are representative of at least three independent experiments. Values are mean ± SD. Unpaired two-tailed t-test between two groups (c, f, g) and one-way repeated measure ANOVA followed by Bonferroni’s post hoc analysis (a) were conducted for statistical analyses. Source data are provided as a Source Data file. TH1, tyrosine hydroxylase 1.
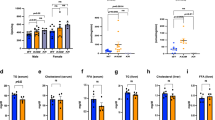
Extended Data Fig. 3 Metabolic characteristics of adipocyte Pak4 KO mice after HFD feeding.
All experimental procedures were the same as those described in Fig. 3’s legend. a. Body weight changes in six-week-old male WT and Pak4 KO mice over 16 weeks of NCD or HFD feeding (n = 5). *p < 0.05 versus NCD WT; #p < 0.05 and ##p < 0.01 versus HFD WT. b, c. Food intake and locomoter activity were measured by indirect calorimetric assay on a daily basis (n = 5). d-i. Plasma levels of leptin, adiponectin, triacylglycerol (TG), total cholesterol (TC), LDL cholesterol (LDL-C), and fatty acid binding protein 4 (FABP4) were measured using a specific assay kit (n = 5). j. Stromal vascular cells (SVCs) were isolated from EAT and stained with F4/80-FITC, CD11b-APC, and CD11c-PE. After dead cells were excluded, the remaining cells were analyzed by flow cytometry. The subpopulation of M1-like macrophages (F4/80+CD11b+CD11c+) and M2-like macrophages (F4/80+CD11b+CD11c−) are expressed as a percentage of SVCs (n = 5). k. Expression of macrophage infiltration- and macrophage subtype-related genes was determined by qPCR (n = 6). l. Plasma levels of CCL2, IL-6, IL-1β, IL-4 and IL-10 were assayed using ELISA (n = 6). m. Homeostasis Model Assessment-insulin resistance (HOMA-IR) was calculated using the following formula: HOMA-IR = fasting glucose (mg/dl) × insulin (μU/ml)/405 (n = 5). n. Adipose tissue- and liver-specific insulin sensitivity was measured by assessing the level of insulin-stimulated AKT phosphorylation. The intensities of the total- and phospho-AKT bands were quantified (n = 9). Data are representative of at least three independent experiments. Values are mean ± SD. Unpaired two-tailed t-test between two groups (b-n) and two-way repeated-measures ANOVA followed by Bonferroni’s post hoc analysis (a) were conducted for statistical analyses. Source data are provided as a Source Data file.
Extended Data Fig. 4 Oxygen consumption rate (OCR) and energy expenditure.
All experimental procedures were the same as those described in Fig. 3’s legend. a-c. Indirect calorimetry was performed using an 8-chamber Oxymax system. The mice were acclimatized to cages for 24 h, and data were collected for an additional 48 h to determine the 24-hour O2 consumption rates (VO2), CO2 production rates (VCO2), and average energy expenditure (EE) in mice (n = 5 for NCD WT, NCD KO and HFD WT, n = 6 for HFD KO). d. Oxygen consumption rate was measured using a Seahorse XF analyser in isolated adipocytes. Basal respiration, respiration related to ATP production (uncoupled, difference between OCR before and after 1 µM oligomycin injection), and maximal respiration (difference between OCR after 1 µM FCCP and 0.5 µM rotenone/antimycin A injection) were determined (n = 12). *p < 0.05 versus WT. Data are representative of at least three independent experiments. Values are mean ± SD. ANCOVA analysis (a-c) and unpaired two-tailed t-test between two groups (d) were conducted for statistical analyses. Source data are provided as a Source Data file. MR, metabolic rate.
Extended Data Fig. 5 Abrogation of weight loss effects of PAK4 deficiency under thermoneutral conditions.
Eight-week-old male Pak4 KO mice and their littermates (WT) were fed either an NCD or a 60% HFD for 16 weeks at 30 °C. a. Representative photographs of Pak4 KO and WT mice. b. Body weight changes in WT and Pak4 KO mice over 16 weeks of NCD or HFD feeding (n = 5). c. Gross examination of adipose tissues and weights of EAT, IAT, and BAT presented as percentages of body weight (BW) (n = 7). d. EAT sections were H&E stained or immunostained with antibodies against F4/80 and perilipin (Plin1). Bars=50 μm. Adipocyte size in H&E sections was quantified (n = 3).Values are expressed as mean ± SD. Unpaired two-tailed t-test between two groups (c) and two-way repeated-measures ANOVA followed by Bonferroni’s post hoc analysis (b, d) were conducted for statistical analyses. Source data are provided as a Source Data file.
Extended Data Fig. 6 Effects of PAK4 overexpression on TG lipolysis in high fat diet-fed mice under fasting condition.
a. Schematic illustration of the preparation of PAK4- or PAK4S474A-loaded in vivo-jetPEI. Six-week-old male WT and Pak4 KO mice were injected with 20 μg of PAK4 or PAK4S474A and then fed an HFD for 9 weeks. b. Protein levels of Flag were examined in various organs at 9 weeks of HFD feeding in WT mice. c. PAK4 protein levels at indicated time points were compared in WT mice. d, h. Body weight change was monitored over time (n = 7 for d, n = 6 for h). *p < 0.05 versus mock. e-g, i-l. After 24-hour fasting, wet weights of EAT, plasma levels of FFA and glycerol and activation of lipolytic pathway in EAT were compared (n = 10 for e-g, n = 6 for i-k). Data are representative of at least three independent experiments. Values are mean ± SD. One-way ANOVA followed by Bonferroni’s post hoc analysis (e-g, i-k) and one-way repeated-measures ANOVA followed by Bonferroni’s post hoc analysis (d, h) were conducted for statistical analyses. Source data are provided as a Source Data file.
Extended Data Fig. 7 Identification of HSL phosphorylation site by PAK4.
a. After transfecting 3T3-L1 adipocytes as indicated, co-IP was performed to determine PAK4′s interaction with and phosphorylation of ATGL. b. 3T3-L1 cells were transfected with PAK4 and HSL. Phosphorylation of HSL was determined using LC-MS/MS analysis after in-gel digestion. A representative spectrum is shown. c. The primary sequence of murine HSL is shown with highlighted sequence coverage (78%; letters in bold) and phosphorylation of HSL (S565, letter in red) revealed by LC-MS/MS. d. 3T3-L1 adipocytes were transfected as indicated and then treated with 1 μM isoproterenol (Iso) for 1 h. Phosphorylation of HSL, glycogen synthase kinase-3 beta (GSK3β), and AMP activated kinase (AMPK) was determined by Western blotting. Data are representative of at least three independent experiments. Source data are provided as a Source Data file.
Extended Data Fig. 8 Phosphorylation of FABP4 on T126 by PAK4.
a. Recombinant FABP4 was incubated with recombinant PAK4 and [32P]ATP for 30 min, and proteins in the mixture were resolved by SDS-PAGE. The band was visualized by autoradiography of 32P-labeled protein. Protein loading was confirmed by Coomassie blue staining. b. 3T3-L1 adipocytes were transfected with PAK4 and then treated with 1 μM isoproterenol for 1 h. FABP4 binding with CGI-58 was analyzed by PLA (n = 26). c. 3T3-L1 adipocytes were transfected as indicated. PAK4 phosphorylation of FABP4 was analyzed by Western blotting. d. A predicted model of the interaction between PAK4 and FABP4, represented by green and yellow, respectively, in their ternary structure. The green ball-and-stick model depicts ATP bound to PAK4, while the yellow stick model represents palmitate binding to FABP4. Additionally, the sphere located in the ATP binding pocket of PAK4 indicates the presence of Mg2+ ion. Notably, the model suggests that the phosphorylation of FABP4 on the T126 residue by PAK4 is structurally feasible. e. 3T3-L1 adipocytes were transfected as indicated and then treated with 1 μM isoproterenol for 1 h. The size of lipid droplets was then analyzed (n = 52). Data are representative of at least three independent experiments. Values are mean ± SD. Two-way ANOVA followed by Bonferroni’s post hoc analysis (b, e) was conducted for statistical analyses. Source data are provided as a Source Data file.
Extended Data Fig. 9 Relationship between PAK4 expression in visceral fat and obesity parameters in non-diabetic humans.
a, b. Scatter plot of adipose PAK4 protein and mRNA levels and metabolic parameters. The coefficient of determination in two sided distributions was used to compare the association of PAK4 expression with metabolic parameters (n = 88). c. Western blot analysis of PAK4, p-HSL-S565, and p-FABP4-T126 in primary visceral adipocytes isolated from lean (BMI < 25), overweight (25 ≤ BMI < 30), and obese (BMI ≥ 30) cohorts (n = 10). d. After PAK4 silencing or overexpression in human adipocytes, the proteins involved in mitochondrial dynamics were analyzed by Western blotting. Data are representative of at least three independent experiments. Values are mean ± SD. One-way ANOVA followed by Bonferroni’s post hoc analysis (c) was conducted for statistical analyses. Source data are provided as a Source Data file. PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha; NRF1, nuclear respiratory factor 1; DRP1, dynamin-related protein 1; Fis1, fission 1.
Supplementary information
Supplementary Information
Supplementary Figs. 1–12 and Tables 1–7.
Supplementary Data
PAK4 expression in mice and 3T3-L1 adipocytes. a, PAK4 expression was examined in brain, lung, liver, bone marrow (BM), heart, skeletal muscle (SM), BAT, IAT and EAT by western blotting (n = 9). b, The band intensity from Fig. 1a was quantified by densitometry (n = 10). c, The band intensity from Fig. 1b was quantified by densitometry (n = 9), and Pak4 mRNA levels were quantified by qPCR (n = 11). d, PAK4 protein levels in 3T3-L1 preadipocytes and mature adipocytes were determined (n = 10). e, The band intensity from Fig. 1d was quantified by densitometry (n = 9), and Pak4 mRNA levels were quantified by qPCR (n = 8). f, 3T3-L1 adipocytes were treated with 1 μM norepinephrine (NE), 1 μM isoproterenol (Iso), 1 μM forskolin (Fsk), 100 nM CL-316243 (CL), 1 μM 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), 1 μM H89, 1 μM SR59230A (SR) or 20 nM insulin (Ins) for 1 h, and PAK4 protein levels were analysed by western blotting (n = 6). g, 3T3-L1 adipocytes were treated with 1 μM isoproterenol for 1 h with or without 10 μM Rp-cAMP, and PAK4 protein levels were analysed. The red Ponceau S-stained gel was used as the loading control for the PKA substrate. h, 3T3-L1 adipocytes were treated with 1 μM isoproterenol with or without either 3 μM MG132 or 1 μM chloroquine for 1 h and PAK4 protein levels were analysed (n = 7). Data are representative of at least three independent experiments. Values are mean ± s.d. Unpaired two-tailed t-test between two groups (b,d) and one-way ANOVA followed by Bonferroni’s post hoc analysis (c,e,f,h) were conducted for statistical analyses. Source data are provided as a source data file. Veh, vehicle. Generation of adipocyte-specific Pak4 KO mice. a, Schematic diagram illustrating the adipocyte-specific ablation of Pak4 using Adipoq-Cre mice. b, PCR analyses of genotyping from tail biopsies of Adipoq-Cre;Pak4flox/flox (KO) mice and their littermates (Pak4flox/flox, WT). c, Protein levels of PAK4 in the brain, lung, liver, bone marrow (BM), heart, skeletal muscle (SM), BAT, IAT and EAT of WT and Pak4 KO mice were analysed by western blotting. d, Protein levels of PAK family members in EATs were analysed by western blotting and their protein density was quantified (n = 7). Data are representative of at least three independent experiments. Values are mean ± s.d. Unpaired two-tailed t-test between two groups was conducted for statistical analyses (d). Source data are provided as a source data file. Increase in lipolysis in female adipocyte Pak4 KO mice under the fasting condition. a, Body weight changes in 6-week-old female WT and Pak4 KO mice fed chow diet ad libitum, 24 h fasted or 6 h refed (n = 7). b. Wet weight of gonadal WAT (GAT), IAT and BAT was measured (n = 6). c, H&E staining of GATs from 6-week-old female WT and Pak4 KO mice in fed and fasted states. Bars, 50 μm. Adipocyte sizes in the H&E sections were quantified (n = 98). d. Protein levels of β-adrenergic receptors and lipolysis-related proteins in GATs were analysed by western blotting. Protein density was quantified (n = 9). e–g, Plasma levels of FFA, glycerol and FABP4 were analysed (n = 6). Data are representative of at least three independent experiments. Values are mean ± s.d. Unpaired two-tailed t-test between two groups (a–d) and two-way ANOVA followed by Bonferroni’s post hoc analysis (e–g) were conducted for statistical analyses. Source data are provided as a source data file. Non-classical FABP4 release from adipocytes in WT and Pak4 KO mice. a,b, Primary adipocytes isolated from Pak4 KO mice and their littermates (WT) were treated with 1 μM isoproterenol for 1 h in the presence or absence of H89 (1 μM), chloroquine (1 μM) and brefeldin A (0.5 μM). FABP4 concentration in the culture medium (CM) and cAMP (n = 7) and free calcium (Ca2+) concentrations (n = 7) in the cytosol were analysed. Immunoblot analyses of FABP4 were performed either on culture medium or whole cell lysates (WCL). c, Primary adipocytes isolated from WT and Pak4 KO mice were treated with 1 μM isoproterenol for 0.5 h and washed out with basal culture media. Afterwards, a 2 h conditioned medium was harvested from the cell culture and added to primary hepatocytes for a 6 h incubation period. Glucose production was analysed following the 6 h incubation with the glucose production buffer (n = 4). Glucagon (3 μM) was used as a positive control. Values are mean ± s.d. NS, not significant. Unpaired two-tailed t-test between two groups (b) and two-way ANOVA followed by Bonferroni’s post hoc analysis (c) were conducted for statistical analyses. Source data are provided as a source data file. Increase of adaptive thermogenesis in female adipocyte Pak4 KO mice after cold exposure. a, Eight-week-old female adipocyte Pak4 KO mice and their WT littermates were housed at 30 °C for 1 day and then exposed to cold (6 °C). The rectal temperature of mice was measured at indicated time points after cold exposure (n = 5). *P < 0.05. b, Representative infrared image showing Pak4 and WT mice (upper panel) and gross morphology of GAT, IAT and BAT (lower panel) housed at 6 °C for 3 days. c. Weights of GAT, IAT and BAT are presented as percentages of body weight (n = 4). d, IAT and BAT were H&E stained or immunostained with an antibody against UCP1. Bars, 50 μm. e,f, Protein levels of thermogenic markers and OxPhos complex (n = 6) were determined by western blotting. Data are representative of at least three independent experiments. Values are mean ± s.d. Unpaired two-tailed t-test between two groups (c,f) and one-way repeated measure ANOVA followed by Bonferroni’s post hoc analysis (a) were conducted for statistical analyses. Source data are provided as a source data file. d, IAT and BAT were H&E stained or immunostained with an antibody against UCP1. Scale bars, 50 μm. e,f, Protein levels of thermogenic markers and OxPhos complex (n = 6) were determined by western blotting. Data are representative of at least three independent experiments. Values are mean ± s.d. *P < 0.05. Unpaired two-tailed t-test between two groups (c,f) and one-way repeated measure ANOVA followed by Bonferroni’s post hoc analysis (a) were conducted for statistical analyses. Source data are provided as a source data file. Gating strategy for flow cytometry and western blotting analysis of lipolysis-related proteins. All experimental procedures were the same as those described in Fig. 3’s legend. a, Gating strategy for flow cytometry analysis for Extended Data Fig. 3j. After exclusion of doublets and debris, a gating strategy was used to identify M1 macrophages (F4/80+CD11b+CD11c+) and M2 macrophages (F4/80+CD11b+CD11c−). b, Total and phosphorylated protein levels of β-adrenergic receptors and lipolysis-related proteins were analysed by western blotting (n = 9). Data are representative of at least three independent experiments. Values are mean ± s.d. Unpaired two-tailed t-test between two groups was conducted for statistical analyses. Source data are provided as a source data file. Resistance to weight gain in female adipocyte Pak4 KO mice. Eight-week-old female Pak4 KO mice and their littermates (WT) were fed either an NCD or a 60% HFD for 16 weeks. a, Representative photographs of female Pak4 KO mice and their littermates. b, GAT sections were H&E stained or immunostained with antibodies against F4/80 and perilipin (Plin1). Bars, 50 μm. c, Adipocyte size in H&E sections was quantified (n = 3). d, Body weight change was monitored over time (n = 5). e–g. Plasma levels of adiponectin, TG and TC were assayed using specific kits (n = 5). h, Total and phosphorylated protein levels of lipolysis-related proteins were analysed by western blotting (n = 8). Values are mean ± s.d. *P < 0.05 and **P < 0.01 versus NCD WT; #P < 0.05 and ##P < 0.01 versus HFD WT (for c,d). Unpaired two-tailed t-test between two groups (e–h) and two-way repeated-measures ANOVA followed by Bonferroni’s post hoc analysis (c,d) were conducted for statistical analyses. Source data are provided as a source data file. Genotype tissue expression (GTEx) analysis of human adipose tissues. a,b, Biological processes of gene ontology categories (a) and KEGG pathway (b) were analysed for 178 genes that negatively correlated with PAK4 mRNA (Pearson coefficient < −0.3) in human adipose tissues (n = 797). RNA-seq analysis of EATs. a, Total RNA from WT and Pak4 KO EATs was used for RNA-seq analysis. DEGs are shown in a hierarchically clustered heatmap. For hierarchical clustering, the distance metric between rows was calculated according to the log2 of FPKM (fragments per kilobase of transcript per million mapped reads) values and then normalized by Z score. The lowest Z score values are shown in dark blue and the highest values are shown in bright yellow. b, A volcano plot is shown, depicting the log2 fold-difference in mRNA expression in EATs of WT and Pak4 KO mice in two-sided distributions, assessed by StringTie analysis of RNA-seq data. Red and blue dots represent upregulated and downregulated DEGs with more than twofold change and P < 0.05, respectively. Each sample was analysed in triplicate. c, Biological process analysis was performed to analyse the DEGs. d, KEGG pathway analysis was performed to analyse the DEGs. Phosphoproteomic analysis of EAT. a, A representative phosphoimage of two-dimensional polyacrylamide gel electrophoresis. EATs from 24 h fasted-Pak4 KO and WT mice were subjected to two-dimensional gel electrophoresis. The gels from WT and KO mice were stained with Cy3 (green) and Cy5 (red), respectively. The phosphorylated gels were stained with a ProQ Diamond phosphorylation detection dye and then scanned. The overlay images were generated using Typhoon 9400 software. The molecular mass is indicated on the left and the pH range is shown at the bottom. The proteins that showed a visible difference in phosphorylation levels are indicated by circles and labelled with serial numbers. Spot no. 65 (shown as red circle) corresponds to FABP4. Phosphoproteins with a significantly altered abundance, which have been identified by mass spectrometry, are listed in Supplementary Table 3. b, Whole cell lysates were subjected to LC–MS/MS phospho-tryptic peptide identification and quantification. The top ranked phosphoproteins are presented. c. Whole cell lysates were subjected to co-IP to determine PAK4’s interaction with and phosphorylation of SPA3K. a, ANS displacement assay of palmitate (PA) and BMS309403 (BMS) for various forms of FABP4. 3T3-L1 adipocytes were transfected with GFP-FABPWT, GFP-FABP4T126A or GFP-FABP4T126D at day 4 post-MDI treatment. The fluorescent probe ANS (4 mM) was then added in the presence of indicated concentrations of palmitate or BMS, a cell-permeable, potent inhibitor of FABP4 that targets the fatty acid-binding pocket. Fluorescent intensity was calculated as a percentage of it in the absence of palmitate or BMS (n = 4). Each fitting curve was acquired from the mean of three separate experiments using Prism v.9.5 software. b. 3T3-L1 adipocytes were transfected with GFP-FABPWT, GFP-FABP4T126A or GFP-FABP4T126D at day 4 post-MDI treatment. Seven days after MDI treatment, adipocytes were incubated with 3 μM NBD-PA for 30 min and then treated with isoproterenol (0.3 μM) for 1 h in the presence or absence of BMS. FABP4 binding with palmitate was analysed by PLA. Data are representative of at least three independent experiments. Values are mean ± s.d. Source data are provided as a source data file. ANS, 1-anilinonaphthalene-8-sulfonic acid; NBD, 7-nitrobenzenz-2-oxa-1,3-diazol-4-yl palmitic acid. Indirect calorimetric study and seahorse analysis. All experimental procedures were the same as those described in Fig. 7’s legend. Mice were acclimatized to cages for 24 h, and data was collected for an additional 48 h. a–d. Food intake, locomotor activity, 24 h O2 consumption rates (VO2) and 48 h average energy expenditure (EE) were determined (n = 4 for Veh, n = 5 for ND25, ND50). e, Oxygen consumption rate was measured using the Seahorse XF analyser in isolated adipocytes (n = 15). *P < 0.05 and **P < 0.01 versus Veh. f, Western blotting of OxPhos complex in primary adipocytes. Data are representative of at least three independent experiments. Values are mean ± s.d. One-way ANOVA followed by Bonferroni’s post hoc analysis (a,b), analysis of covariance (c,d) and one-way repeated-measures ANOVA followed by Bonferroni’s post hoc analysis (e) were conducted for statistical analyses. Source data are provided as a source data file. ND25, ND201651 25 mg kg−1; ND50, ND201651 50 mg kg−1; ND30, ND201651 30 nM; ND100, ND201651 100 nM; MR, metabolic rate.
Source data
Source Data Fig. 1
Unprocessed western blots.
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Unprocessed western blots.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Unprocessed western blots.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Unprocessed western blots.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Unprocessed western blots.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Unprocessed western blots.
Source Data Fig. 7
Statistical source data.
Source Data Fig. 8
Unprocessed western blots.
Source Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 1
Unprocessed western blots.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Unprocessed western blots.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Unprocessed western blots.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Unprocessed western blots.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Unprocessed western blots.
Source Data Extended Data Fig. 8
Unprocessed western blots.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Unprocessed western blots.
Source Data Extended Data Fig. 9
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yu, H.C., Jeon, Y.G., Na, AY. et al. p21-activated kinase 4 counteracts PKA-dependent lipolysis by phosphorylating FABP4 and HSL. Nat Metab 6, 94–112 (2024). https://doi.org/10.1038/s42255-023-00957-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-023-00957-x
