
Abstract
Obesity is associated with chronic low-grade white adipose tissue (WAT) inflammation that can contribute to the development of insulin resistance in mammals. Previous studies have identified interleukin (IL)-12 as a critical upstream regulator of WAT inflammation and metabolic dysfunction during obesity. However, the cell types and mechanisms that initiate WAT IL-12 production remain unclear. Here we show that conventional type 1 dendritic cells (cDC1s) are the cellular source of WAT IL-12 during obesity through analysis of mouse and human WAT single-cell transcriptomic datasets, IL-12 reporter mice and IL-12p70 protein levels by enzyme-linked immunosorbent assay. We demonstrate that cDC1s contribute to obesity-associated inflammation by increasing group 1 innate lymphocyte interferon-γ production and inflammatory macrophage accumulation. Inducible depletion of cDC1s increased WAT insulin sensitivity and systemic glucose tolerance during diet-induced obesity. Mechanistically, endocytosis of apoptotic bodies containing self-DNA by WAT cDC1s drives stimulator of interferon genes (STING)-dependent IL-12 production. Together, these results suggest that WAT cDC1s act as critical regulators of adipose tissue inflammation and metabolic dysfunction during obesity.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

Data availability
The source data and single-cell RNA sequencing datasets are accessible from their respective publications19,26,27, and can be found under the accession numbers: GSE155960 and GSE156110 (ref. 19), GSE182233 (ref. 26), and through the Single Cell Portal (study no. SCP1376)27. Mouse assembly mm10 (GRCm39) was sourced from GenBank: GCA_000001635.9. The bulk RNA sequencing datasets from this work are accessible from GEO with accession number GSE236110. Sequencing data used to generate Figs. 1 and 5 and Extended Data Fig. 1, as well as unprocessed blot images for immunoblots in Fig. 4, can be found within the supplementary information included with this manuscript. Source data for additional figures were not included as the individual points presented in each figure clearly present the underlying data, regardless of phenotype. Because of this, the authors do not believe inclusion of the source data will significantly enhance the conclusions drawn. However, for re-analysis purposes, all source data will be made available upon request from the corresponding author. Source data are provided with this paper.
Code availability
This paper does not report original code. All codes used in this paper are available from the lead contact upon request.
References
Ward, Z. J. et al. Projected U.S. state-level prevalence of adult obesity and severe obesity. N. Engl. J. Med. 381, 2440–2450 (2019).
Khaodhiar, L., McCowen, K. C. & Blackburn, G. L. Obesity and its comorbid conditions. Clin. Cornerstone 2, 17–31 (1999).
Guh, D. P. et al. The incidence of co-morbidities related to obesity and overweight: a systematic review and meta-analysis. BMC Public Health 9, 88 (2009).
Kahn, S. E., Hull, R. L. & Utzschneider, K. M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 444, 840–846 (2006).
Després, J.-P. & Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 444, 881–887 (2006).
de Luca, C. & Olefsky, J. M. Inflammation and insulin resistance. FEBS Lett. 582, 97–105 (2009).
Shoelson, S. E. Inflammation and insulin resistance. J. Clin. Invest. 116, 1793–1801 (2006).
Lee, Y. S., Wollam, J. & Olefsky, J. M. An integrated view of immunometabolism. Cell 172, 22–40 (2018).
Lackey, D. E. & Olefsky, J. M. Regulation of metabolism by the innate immune system. Nat. Rev. Endocrinol. 12, 15–28 (2016).
Lumeng, C. N., DeYoung, S. M., Bodzin, J. L. & Saltiel, A. R. Increased inflammatory properties of adipose tissue macrophages recruited during diet-induced obesity. Diabetes 56, 16–23 (2007).
Ying, W. et al. Adipose tissue macrophage-derived exosomal miRNAs can modulate in vivo and in vitro insulin sensitivity. Cell 171, 372–384.e12 (2017).
Hotamisligil, G. S. Inflammation, metaflammation and immunometabolic disorders. Nature 542, 177–185 (2017).
Patsouris, D. et al. Ablation of CD11c-positive cells normalizes insulin sensitivity in obese insulin resistant animals. Cell Metab. 8, 301–309 (2008).
O’Sullivan, T. E. et al. Adipose-resident group 1 innate lymphoid cells promote obesity-associated insulin resistance. Immunity 45, 428–441 (2016).
Wensveen, F. M. et al. NK cells link obesity-induced adipose stress to inflammation and insulin resistance. Nat. Immunol. 16, 376–385 (2015).
Lee, B.-C. et al. Adipose natural killer cells regulate adipose tissue macrophages to promote insulin resistance in obesity. Cell Metab. 23, 685–698 (2016).
Wang, H. et al. Adipose group 1 innate lymphoid cells promote adipose tissue fibrosis and diabetes in obesity. Nat. Commun. 10, 3254 (2019).
O’Rourke, R. W. et al. Systemic inflammation and insulin sensitivity in obese IFN-γ knockout mice. Metabolism 61, 1152–1161 (2012).
Hildreth, A. D. et al. Single-cell sequencing of human white adipose tissue identifies new cell states in health and obesity. Nat. Immunol. 22, 639–653 (2021).
Bradley, D. et al. Interferon gamma mediates the reduction of adipose tissue regulatory T cells in human obesity. Nat. Commun. 13, 5606 (2022).
Pacifico, L. et al. Increased T-helper interferon-γ-secreting cells in obese children. Eur. J. Endocrinol. 154, 691–697 (2006).
Dobrian, A. D. et al. STAT4 deficiency reduces obesity-induced insulin resistance and adipose tissue inflammation. Diabetes 62, 4109–4121 (2013).
Nam, H., Ferguson, B. S., Stephens, J. M. & Morrison, R. F. Impact of obesity on IL-12 family gene expression in insulin responsive tissues. Biochim. Biophys. Acta Mol. Basis Dis. 1832, 11–19 (2013).
Schmidt, F. M. et al. Inflammatory cytokines in general and central obesity and modulating effects of physical activity. PLoS ONE 10, e0121971 (2015).
Suárez-Álvarez, K. et al. Serum IL-12 is increased in Mexican obese subjects and associated with low-grade inflammation and obesity-related parameters. Mediators Inflamm. 2013, 967067 (2013).
Cottam, M. A., Caslin, H. L., Winn, N. C. & Hasty, A. H. Multiomics reveals persistence of obesity-associated immune cell phenotypes in adipose tissue during weight loss and weight regain in mice. Nat. Commun. 13, 2950 (2022).
Emont, M. P. et al. A single-cell atlas of human and mouse white adipose tissue. Nature 603, 926–933 (2022).
Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 3, 133–146 (2003).
Reinhardt, R. L., Hong, S., Kang, S.-J., Wang, Z. & Locksley, R. M. Visualization of IL-12/23p40 in vivo reveals immunostimulatory dendritic cell migrants that promote Th1 differentiation. J. Immunol. 177, 1618–1627 (2006).
Cabeza-Cabrerizo, M., Cardoso, A., Minutti, C. M., Pereira da Costa, M. & Reis e Sousa, C. Dendritic cells revisited. Annu. Rev. Immunol. 39, 131–166 (2021).
Vignali, D. A. A. & Kuchroo, V. K. IL-12 family cytokines: immunological playmakers. Nat. Immunol. 13, 722–728 (2012).
Bertola, A. et al. Identification of adipose tissue dendritic cells correlated with obesity-associated insulin-resistance and inducing Th17 responses in mice and patients. Diabetes 61, 2238–2247 (2012).
Cho, K. W. et al. Adipose tissue dendritic cells are independent contributors to obesity-induced inflammation and insulin resistance. J. Immunol. 197, 3650–3661 (2016).
Stefanovic-Racic, M. et al. Dendritic cells promote macrophage infiltration and comprise a substantial proportion of obesity-associated increases in CD11c+ cells in adipose tissue and liver. Diabetes 61, 2330–2339 (2012).
Böttcher, J. P. et al. NK cells stimulate recruitment of cDC1 into the tumor microenvironment promoting cancer immune control. Cell 172, 1022–1037.e14 (2018).
Ghilas, S. et al. Natural killer cells and dendritic epidermal γδ T cells orchestrate type 1 conventional DC spatiotemporal repositioning toward CD8+ T cells. iScience 24, 103059 (2021).
Efremova, M., Vento-Tormo, M., Teichmann, S. A. & Vento-Tormo, R. CellPhoneDB: inferring cell–cell communication from combined expression of multi-subunit ligand–receptor complexes. Nat. Protoc. 15, 1484–1506 (2020).
Weizman, O.-E. et al. ILC1 confer early host protection at initial sites of viral infection. Cell 171, 795–808.e12 (2017).
Riggan, L. et al. CRISPR-Cas9 ribonucleoprotein-mediated genomic editing in mature primary innate immune cells. Cell Rep. 31, 107651 (2020).
Yamazaki, C. et al. Critical roles of a dendritic cell subset expressing a chemokine receptor, XCR1. J. Immunol. 190, 6071–6082 (2013).
Nishimura, S. et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 15, 914–920 (2009).
Macdougall, C. E. et al. Visceral adipose tissue immune homeostasis is regulated by the crosstalk between adipocytes and dendritic cell subsets. Cell Metab. 27, 588–601.e4 (2018).
Hernández-García, E. et al. Conventional type 1 dendritic cells protect against age-related adipose tissue dysfunction and obesity. Cell. Mol. Immunol. 19, 260–275 (2022).
Hamade, H. et al. BATF3 protects against metabolic syndrome and maintains intestinal epithelial homeostasis. Front. Immunol. 13, 841065 (2022).
Hopfner, K.-P. & Hornung, V. Molecular mechanisms and cellular functions of cGAS–STING signalling. Nat. Rev. Mol. Cell Biol. 21, 501–521 (2020).
Montecino-Rodriguez, E. & Dorshkind, K. Use of busulfan to condition mice for bone marrow transplantation. STAR Protoc. 1, 100159 (2020).
Deng, L. et al. STING-dependent cytosolic DNA sensing promotes radiation-induced type I interferon-dependent antitumor immunity in immunogenic tumors. Immunity 41, 843–852 (2014).
Wang, H. et al. cGAS is essential for the antitumor effect of immune checkpoint blockade. Proc. Natl Acad. Sci. USA 114, 1637–1642 (2017).
Carozza, J. A. et al. Extracellular cGAMP is a cancer-cell-produced immunotransmitter involved in radiation-induced anticancer immunity. Nat. Cancer 1, 184–196 (2020).
Marcus, A. et al. Tumor-derived cGAMP triggers a STING-mediated interferon response in non-tumor cells to activate the NK cell response. Immunity 49, 754–763.e4 (2018).
Ritchie, C., Cordova, A. F., Hess, G. T., Bassik, M. C. & Li, L. SLC19A1 is an importer of the immunotransmitter cGAMP. Mol. Cell 75, 372–381.e5 (2019).
Klarquist, J. et al. STING-mediated DNA sensing promotes antitumor and autoimmune responses to dying cells. J. Immunol. 193, 6124–6134 (2014).
Woo, S.-R. et al. STING-dependent cytosolic DNA sensing mediates innate immune recognition of immunogenic tumors. Immunity 41, 830–842 (2014).
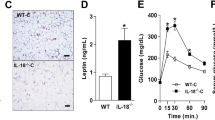
Nishimoto, S. et al. Obesity-induced DNA released from adipocytes stimulates chronic adipose tissue inflammation and insulin resistance. Sci. Adv. 2, e1501332 (2016).
Merrick, D. et al. Identification of a mesenchymal progenitor cell hierarchy in adipose tissue. Science 364, eaav2501 (2019).
Kumari, M., Heeren, J. & Scheja, L. Regulation of immunometabolism in adipose tissue. Semin. Immunopathol. 40, 189–202 (2018).
Kakarla, R., Hur, J., Kim, Y. J., Kim, J. & Chwae, Y.-J. Apoptotic cell-derived exosomes: messages from dying cells. Exp. Mol. Med. 52, 1–6 (2020).
Atkin-Smith, G. K. & Poon, I. K. H. Disassembly of the dying: mechanisms and functions. Trends Cell Biol. 27, 151–162 (2017).
Essandoh, K. et al. Blockade of exosome generation with GW4869 dampens the sepsis-induced inflammation and cardiac dysfunction. Biochim. Biophys. Acta Mol. Basis Dis. 1852, 2362–2371 (2015).
de Mingo Pulido, Á. et al. The inhibitory receptor TIM-3 limits activation of the cGAS-STING pathway in intra-tumoral dendritic cells by suppressing extracellular DNA uptake. Immunity 54, 1154–1167.e7 (2021).
Iyoda, T. et al. The CD8ϩ dendritic cell subset selectively endocytoses dying cells in culture and in vivo. J. Exp. Med. 195, 1289–1302 (2002).
Platt, C. D. et al. Mature dendritic cells use endocytic receptors to capture and present antigens. Proc. Natl Acad. Sci. USA 107, 4287–4292 (2010).
Zelenay, S. et al. The dendritic cell receptor DNGR-1 controls endocytic handling of necrotic cell antigens to favor cross-priming of CTLs in virus-infected mice. J. Clin. Invest. 122, 1615–1627 (2012).
Pamir, N. et al. Granulocyte/macrophage colony-stimulating factor-dependent dendritic cells restrain lean adipose tissue expansion. J. Biol. Chem. 290, 14656–14667 (2015).
Harly, C. et al. The transcription factor TCF-1 enforces commitment to the innate lymphoid cell lineage. Nat. Immunol. 20, 1150–1160 (2019).
Ataide, M. A. et al. BATF3 programs CD8+ T cell memory. Nat. Immunol. 21, 1397–1407 (2020).
Lee, W., Kim, H. S., Hwang, S. S. & Lee, G. R. The transcription factor Batf3 inhibits the differentiation of regulatory T cells in the periphery. Exp. Mol. Med. 49, e393 (2017).
Kim, D.-H. et al. The role of GM-CSF in adipose tissue inflammation. Am. J. Physiol. Endocrinol. Metab. 295, E1038–E1046 (2008).
Darrasse-Jèze, G. et al. Feedback control of regulatory T cell homeostasis by dendritic cells in vivo. J. Exp. Med. 206, 1853–1862 (2009).
McKenna, H. J. et al. Mice lacking flt3 ligand have deficient hematopoiesis affecting hematopoietic progenitor cells, dendritic cells, and natural killer cells. Blood 95, 3489–3497 (2000).
Guendel, F. et al. Group 3 innate lymphoid cells program a distinct subset of IL-22BP-producing dendritic cells demarcating solitary intestinal lymphoid tissues. Immunity 53, 1015–1032.e8 (2020).
Sano, T. et al. Ccr7 null mice are protected against diet-induced obesity via Ucp1 upregulation and enhanced energy expenditure. Nutr. Metab. (Lond.) 16, 43 (2019).
Mao, Y. et al. STING–IRF3 triggers endothelial inflammation in response to free fatty acid-induced mitochondrial damage in diet-induced obesity. Arterioscler. Thromb. Vasc. Biol. 37, 920–929 (2017).
Stutte, S. et al. High-fat diet rapidly modifies trafficking, phenotype, and function of plasmacytoid dendritic cells in adipose tissue. J. Immunol. 208, 1445–1455 (2022).
Ohta, T. et al. Crucial roles of XCR1-expressing dendritic cells and the XCR1-XCL1 chemokine axis in intestinal immune homeostasis. Sci. Rep. 6, 23505 (2016).
Silva-Sanchez, A. et al. Activation of regulatory dendritic cells by Mertk coincides with a temporal wave of apoptosis in neonatal lungs. Sci. Immunol. 8, eadc9081 (2023).
Hildreth, A. D., Padilla, E. T., Tafti, R. Y., Legala, A. R. & O’Sullivan, T. E. Sterile liver injury induces a protective tissue-resident cDC1-ILC1 circuit through cDC1-intrinsic cGAS-STING-dependent IL-12 production. Cell Rep. 42, 112141 (2023).
Deczkowska, A. et al. XCR1+ type 1 conventional dendritic cells drive liver pathology in non-alcoholic steatohepatitis. Nat. Med. 27, 1043–1054 (2021).
Maric, I. et al. Sex and species differences in the development of diet-induced obesity and metabolic disturbances in rodents. Front. Nutr. 9, 828522 (2022).
Pettersson, U. S., Waldén, T. B., Carlsson, P.-O., Jansson, L. & Phillipson, M. Female mice are protected against high-fat diet induced metabolic syndrome and increase the regulatory T cell population in adipose tissue. PLoS ONE 7, e46057 (2012).
Hildreth, A. D., Riggan, L. & O’Sullivan, T. E. CRISPR-Cas9 ribonucleoprotein-mediated genomic editing in primary innate immune cells. STAR Protoc. https://doi.org/10.1016/j.xpro.2020.100113 (2020).
Riggan, L. et al. The transcription factor Fli1 restricts the formation of memory precursor NK cells during viral infection. Nat. Immunol. 23, 556–567 (2022).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e29 (2021).
Raudvere, U. et al. g:Profiler: a web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 47, W191–W198 (2019).
Acknowledgements
We thank members of the O’Sullivan, Su, Dalod, Covarrubias, Pellegrini, Vallim and Villanueva laboratories for discussions. A.D.H. was supported by the NIH National Research Service Award F31, NIDDK (grant no. F31DK130585). T.E.O. was supported by the NIH (grant no. AI145997).
Author information
Authors and Affiliations
Contributions
A.D.H. and T.E.O. designed the study. A.D.H., E.T.P., M.G., Y.Y.W., R.S. and A.R.L. performed the experiments. E.T.P. performed bioinformatics analyses. T.E.O. and A.D.H. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
T.E.O. is an advisory board member for Modulus Therapeutics and Xyphos Inc. The other authors declare no competing interests.
Peer review
Peer review information
Nature Metabolism thanks Frederic Geissmann, Philippe Pierre and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Yanina-Yasmin Pesch, Isabella Samuelson, in collaboration with the Nature Metabolism team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Lineage-associated gene signatures of myeloid cells from obese mouse and human WAT scRNAseq datasets.
a, Dot plots showing selected top differentially expressed marker genes for each human myeloid cluster, supporting assignment of clusters to compartments shown in Fig. 1a. Color saturation indicates the strength of expression in positive cells, while dot size reflects the percentage of each cell cluster expressing the gene. b, Dot plots showing selected top differentially expressed marker genes for each mouse myeloid cluster, supporting assignment of clusters to compartments shown in Fig. 1b. Color saturation indicates the strength of expression in positive cells, while dot size reflects the percentage of each cell cluster expressing the gene.
Extended Data Fig. 2 IL-12 production is increased in WAT cDC1s during obesity, independent of diet.
a-h, Il12bYFP mice were fed a LFD or HFD for 4 weeks. a, Representative gating strategy of unbiased analysis of YFP+ cells in the WAT from LFD (left) or HFD (right) Il12bYFP mice. b, Representative gating strategy for WAT dendritic cell and macrophage populations. Dendritic cells: CD45+Lineage-(CD3−TCRβ-NK1.1-CD19−Ly6G−)CD88−MHCII+CD11c+: Conventional type 2 Dendritic cells (cDC2): CD11b+XCR1−, Conventional type 1 Dendritic cells (cDC1): CD11b−XCR1+. Macrophages: CD45+Lineage−(CD3−TCRβ-NK1.1-CD19−Ly6G−)CD88+: Perivascular Macrophages (PVM): Tim-4+CD11c−, Inflammatory Macrophages (IM): Tim-4-CD11c+, Lipid Associated Macrophages (LAM): Tim-4-CD11c−CD9+. c, Representative flow plots of YFP+ cell percentage in WAT IM, PVM, cDC2, and cDC1 populations. d, YFP MFI of the indicated populations isolated from the visceral and subcutaneous adipose tissues (VAT, SAT). (LFD n = 3, HFD n = 5). The percentage (e) and MFI (f) of YFP+ cDC1s isolated from the spleen. The percentage (g) and MFI (h) of YFP+ cDC1 isolated from the liver. i–k, Il12bYFP mice were fed a LFD (n = 5), 60% HFD (n = 5), or Western diet (n = 4) for 4 weeks. The percentage (i), MFI (j), and Absolute density (k) of VAT YFP+ cDC1 isolated from the indicated cohorts. l, m, Dot plots showing obese human IL23A (l) or obese mouse Il23a (m) expression in the WAT populations depicted from each individual dataset. Color saturation indicates the strength of expression in positive cells, while dot size reflects the percentage of each cell cluster expressing the gene. (d–k) Data are representative of two independent experiments and samples were compared using two-tailed Student’s t test with Welch’s correction, assuming unequal SD. Data are presented as individual points with the mean ± SEM. NS: not significant. p < 0.05 was considered significant.
Extended Data Fig. 3 NK cells are the major source of Xcl1 and NKCre+Eomesfl/fl mice can be used to interrogate NK cell function in vivo.
a,b, Xcl1mTfp1-fl or WT mice were fed a LFD (n = 4) or HFD (n = 8) for 4 weeks before assessment of WAT TFP expression. a, Representative flow plots of TFP expression in visceral adipose tissue (VAT) NK cells from HFD-fed WT (left), LFD-fed Xcl1mTfp1-fl (middle), or HFD-fed Xcl1mTfp1-fl (right) mice. b, Percent of total TFP+ cells derived from the indicated populations, isolated from HFD-fed mice (n = 8). c–f, NKCre−Eomesfl/fl and NKCre+Eomesfl/fl mice were placed on a LFD (NKCre−Eomesfl/fl n = 6, NKCre+Eomesfl/fl n = 3) or 60% HFD (NKCre−Eomesfl/fl n = 6, NKCre+Eomesfl/fl n = 7) for 12 weeks. Average weight gained (c) and average food intake (d) for each cohort, measured weekly. Total VAT mass (e) and VAT mass as a percentage of body mass (f) after week 12. g, Representative flow plots showing VAT NK populations in NKCre−Eomesfl/fl mice (left), NKCre+Eomesfl/fl mice (middle), and NKCre+Eomesfl/fl mice after adoptive transfer of isolated splenic NK cells (right). h, Representative flow plots showing the congenics of VAT NK cells in CD45.2 NKCre−Eomesfl/fl mice (left) and CD45.2 NKCre+Eomesfl/fl mice after adoptive transfer of CD45.1 isolated splenic NK cells (right). The total numbers (i) and frequencies (j) of NK1.1+CD49b+ NK cells from NKCre−Eomesfl/fl mice, NKCre+Eomesfl/fl mice, and NKCre+Eomesfl/fl mice after adoptive transfer of isolated splenic NK cells (n = 6 for all). Data are representative of two independent experiments and samples were compared using two-tailed Student’s t test with Welch’s correction, assuming unequal SD. Data are presented as individual points with the mean ± SEM. NS: not significant. p < 0.05 was considered significant.
Extended Data Fig. 4 Depletion of cDC1s does not impact cDC2 populations, type 1 lymphoid cell accumulation, or ILC1 proliferation in the WAT.
a,b, Xcr1DTR mice were injected with 20 ng/g DT (n = 6) or PBS (n = 6) i.p. before assessment of visceral adipose tissue (VAT) dendritic cell populations 24 hours later. The frequencies of cDC2s from total VAT CD45+ cells (a) and the total numbers of VAT cDC2s (b) from the indicated cohorts (n = 6 for all). c-i, Xcr1DTR mice were fed a LFD or HFD for 4 weeks, receiving either 4 ng/g DT or 1x PBS injections i.p. every 3 days. cDC1 and lymphocyte populations were analyzed at week 4 unless indicated otherwise. c, The numbers of VAT cDC1s isolated from each indicated cohort after 4 weeks (n = 7 for all). d, Representative gating strategy for WAT type 1 immune cell populations. Natural Killer (NK) cells: CD45+CD3−TCRβ-NK1.1+CD200r−CD49b+, Type 1 Innate Lymphoid cells (ILC1): CD45+CD3−TCRβ-NK1.1+CD49b−CD200r+, NKT cells: CD45+CD3+TCRβ+NK1.1+, CD8+ T cells: CD45+CD3+TCRβ+NK1.1-TCRγδ- CD4−CD8+. e, Absolute densities of the indicated VAT lymphocyte populations (LFD + PBS n = 4, LFD + DT n = 5, HFD + PBS n = 7, HFD + DT n = 6). The percentages (f) and numbers (g) of VAT IFN-γ+ NKT cells and CD8+ T cells from each cohort (LFD + PBS n = 4, LFD + DT n = 5, HFD + PBS n = 7, HFD + DT n = 6). h, Representative flow plots of VAT Group 1 ILC Ki-67 staining. i, Frequencies of VAT Ki-67+ ILC1s or NK cells from the indicated cohorts (LFD + PBS n = 4, LFD + DT n = 5, HFD + PBS n = 7, HFD + DT n = 6). Data are representative of two independent experiments and samples were compared using two-tailed Student’s t test with Welch’s correction, assuming unequal SD. Data are presented as individual points with the mean ± SEM. NS: not significant. p < 0.05 was considered significant.
Extended Data Fig. 5 Neither DT treatment of WT mice nor depletion of cDC1s in adult mice impacts weight gain, consumption, or tissue mass accumulation.
a,b,e–i, Xcr1DTR mice were fed either a LFD or HFD for 6 weeks. Starting week 2, two cohorts of mice received 20 ng/g DT injections to deplete endogenous cDC1 populations before continuous treatment with either 4 ng/g DT or 1x PBS every 3 days for 4 weeks. The third cohort was left untreated. a, The proportion of cDC1s reconstituted within the visceral WAT of PBS-treated Xcr1DTR mice on the indicated days post initial 20 ng/g DT injection, compared to untreated cDC1 levels (n = 3 for all). b, The numbers of VAT cDC1s isolated from each indicated cohort after 4 weeks (n = 7 for all). c,d, WT mice were placed on a HFD for 4 weeks, receiving a 20 ng/g DT injection at week 0, followed by treatment with either 4 ng/g DT or 1x PBS every 3 days for 4 weeks. The average weight gained (c) and average food intake (d) for each cohort, measured weekly (n = 6 for all). e, Average food intake for each cohort, measured weekly (LFD n = 7, HFD n = 12). Total VAT mass (f) and VAT mass as a percentage of body mass (g) (LFD n = 7, HFD n = 12, HFD Untreated n = 11). Total liver mass (h) and liver mass as a percentage of body mass (i) after week 6 (LFD n = 7, HFD n = 12, HFD Untreated n = 10). Data is representative of 3 (e–i) and 2 (a–d) independent experiments. Samples were compared using two-tailed Student’s t test with Welch’s correction, assuming unequal SD. Data are presented as individual points with the mean ± SEM. NS: not significant. p < 0.05 was considered significant.
Extended Data Fig. 6 STING deficiency does not affect weight gain, food consumption, or cDC1 IL-12 production in the liver and spleen during obesity.
a–d, Tmem173gt x Il12bYFP (Il12bYFPxStingGt) mice or Il12bYFP littermate controls were placed on LFD (n = 6) or HFD (n = 8) for 4 weeks. Average weight gained (a) and average food intake (b) for each cohort. Total visceral adipose tissue (VAT) mass (c) and VAT mass as a percentage of body mass (d) at week 4. e–h, Busulfan-treated WT hosts were reconstituted with a 50:50 mix of Il12bYFP and Il12bYFPxStingGt bone marrow (e–g) or WT and cGAS-/- hosts were reconstituted with Il12bYFP bone marrow (h). 6 weeks later, mice were placed on a HFD and cDC1 IL-12 production was assessed after 4 weeks. e, The YFP MFI of VAT cDC1s from the indicated donor bone marrow paired within the same mouse (n = 5). The frequency (f) and MFI (g) of splenic IL-12+ cDC1s from the indicated donor bone marrow paired within the same mouse (n = 5). h, The YFP MFI of VAT cDC1s isolated from the indicated cohorts (WT: Il12bYFP n = 7, cGAS-/-: Il12bYFP n = 9). i, Representative flow plots showing VAT cDC1 populations in Xcr1DTR mice 24 hours after PBS (left), DT (middle), or DT treatment plus adoptive transfer of bone marrow derived cDC1s (right) (n = 6 per cohort). j, Representative flow plots showing the congenics of VAT cDC1s in PBS-treated CD45.2 Xcr1DTR mice (top) and DT-treated CD45.2 Xcr1DTR mice after adoptive transfer of CD45.1 bone marrow-derived cDC1s (bottom). The frequencies of cDC1s from total VAT CD45+ cells (k) and total numbers of cDC1s (l) from PBS, DT, or DT-treated plus adoptively transferred ( + cDC1) Xcr1DTR mice 24 h after treatment (n = 6 per cohort). Data is representative 2 independent experiments. Samples were compared using two-tailed Student’s t test with Welch’s correction, assuming unequal SD. Data are presented as individual points with the mean ± SEM. NS: not significant. p < 0.05 was considered significant.
Extended Data Fig. 7 Adipocyte lineage cells incorporate EdU in vivo, and EdU-labeling of 3T3-L1 preadipocytes allows for interrogation of DNA uptake mechanisms of cDC1 in vitro.
a, Representative gating strategy for WAT non-immune cell populations (CD45−CD31−PDGFRα+): Adipocyte precursor cells: ICAM-1-DPP4+, Interstitial progenitor cells: ICAM-1-DPP4−, Committed preadipocytes: DPP4−ICAM-1+. b-d, Xcr1DTR mice were fed a LFD or HFD for 4 weeks, receiving either 100 μg EdU or vehicle (1x PBS) injections intraperitoneally every 3 days. After 4 weeks, EdU treatment was terminated, and mice were DT treated to deplete host cDC1 populations. 1x107 congenically-distinct bone marrow-derived cDC1s were then adoptively transferred and assessment of EdU+ cells was performed 24 hours later. b, Representative flow plots of EdU-labeled visceral adipose tissue (VAT) adipocyte lineage cell populations from LFD- or HFD-fed EdU-treated mice. c, The frequencies of EdU+ adipocyte precursor cells (APC), preadipocytes (pAD) and interstitial progenitor cells (Int. Progen.) isolated from the VAT of each cohort (HFD + EdU: n = 4, all others: n = 3). (d) Representative flow plots of EdU-labeled adoptively transferred visceral VAT cDC1s isolated from LFD- or HFD-fed EdU-treated mice. e,f, 3T3-L1 preadipocytes were differentiated in media with or without 10 μM EdU before assessment of EdU-labeling and maturation via BODIPY staining. e, Representative flow plots of EdU-labeling in 3T3-L1 cultured without (left) or with (right) EdU. f, BODIPY 493/503 staining of undifferentiated vs differentiated EdU-/+ 3T3-L1s. g, Apoptotic bodies (ABs) were generated from EdU-labeled 3T3-L1 adipocytes (see Methods) and then cultured with bone marrow-derived cDC1s for 2 hours before assessment of EdU+ DNA uptake by cDC1s. Representative flow plots of cDC1 EdU+ DNA uptake from culture with EdU+ ABs or with electroporated EdU+ 3T3-L1 ABs treated with 750 U/mL DNase1 (n = 6 for all). Data is representative 2 independent experiments. Samples were compared using two-tailed Student’s t test with Welch’s correction, assuming unequal SD. Data are presented as individual points with the mean ± SEM. p < 0.05 was considered significant.
Extended Data Fig. 8 Proposed model: cDC1 contribute to obesity-associated inflammation and metabolic dysfunction via endocytosis of dead cell-derived DNA and cGAS-STING-mediated IL-12 production.
During obesity, adipocyte hypertrophy and hyperplasia leads to the initiation of stress responses and cell death, releasing DNA-containing apoptotic bodies into the WAT microenvironment. WAT cDC1s endocytose these apoptotic bodies, leading to the activation of cytoplasmic cGAS-STING signaling and production of IL-12. Recruitment of cDC1s via NK cell-derived Xcl1 contributes to the accumulation of cDC1s in the WAT and increases the density of IL-12 producing cDC1s. IL-12 activates tissue-resident ILC1s and WAT-infiltrating NK cells to produce IFN-γ, leading to the polarization of inflammatory macrophages and ultimately contributing to the development of obesity-associated inflammation and insulin resistance in the WAT.
Supplementary information
Supplementary Table
Combined Excel workbook with multiple tabs for Supplementary Tables 1–5.
Source data
Source Data Fig. 4
Source western blots from Fig. 4h.
Source Data Fig. 4
Source western blots from Fig. 4j.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hildreth, A.D., Padilla, E.T., Gupta, M. et al. Adipose cDC1s contribute to obesity-associated inflammation through STING-dependent IL-12 production. Nat Metab 5, 2237–2252 (2023). https://doi.org/10.1038/s42255-023-00934-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-023-00934-4
