
Abstract
Tumour cells utilize multiple strategies to evade the immune system, but the underlying metabolic mechanisms remain poorly understood. The pyruvate dehydrogenase (PDH) complex converts pyruvate to acetyl-coenzyme A in mitochondria, thereby linking glycolysis to the ricarboxylic acid cycle. Here we show that the PDH complex E1 subunit α (PDHE1α) is also located in the cytosol. Cytosolic PDHE1α interacts with IKKβ and protein phosphatase 1B, thereby facilitating the inhibition of the NF-κB pathway. Cytosolic PDHE1α can be phosphorylated at S327 by ERK2 and translocated into mitochondria. Decreased cytosolic PDHE1α levels restore NF-κB signalling, whereas increased mitochondrial PDHE1α levels drive α-ketoglutarate production and promote reactive oxygen species detoxification. Synergistic activation of NF-κB and reactive oxygen species detoxification promotes tumour cell survival and enhances resistance to cytotoxic lymphocytes. Consistently, low levels of PDHE1α phosphorylation are associated with poor prognosis of patients with lung cancer. Our findings show a mechanism through which phosphorylation-dependent subcellular translocation of PDHE1α promotes tumour immune evasion.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Data availability
Source data are provided with this paper. The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Fang, Y. et al. MAL2 drives immune evasion in breast cancer by suppressing tumor antigen presentation. J. Clin. Invest. https://doi.org/10.1172/JCI140837 (2021).
Yamamoto, K. et al. Autophagy promotes immune evasion of pancreatic cancer by degrading MHC-I. Nature 581, 100–105 (2020).
Beatty, G. L. & Gladney, W. L. Immune escape mechanisms as a guide for cancer immunotherapy. Clin. Cancer Res. 21, 687–692 (2015).
Pinzon-Charry, A., Maxwell, T. & Lopez, J. A. Dendritic cell dysfunction in cancer: a mechanism for immunosuppression. Immunol. Cell Biol. 83, 451–461 (2005).
Kearney, C. J. et al. PD-L1 and IAPs co-operate to protect tumors from cytotoxic lymphocyte-derived TNF. Cell Death Differ. 24, 1705–1716 (2017).
Voskoboinik, I., Whisstock, J. C. & Trapani, J. A. Perforin and granzymes: function, dysfunction and human pathology. Nat. Rev. Immunol. 15, 388–400 (2015).
Kearney, C. J. et al. Tumor immune evasion arises through loss of TNF sensitivity. Sci. Immunol. https://doi.org/10.1126/sciimmunol.aar3451 (2018).
Hanahan, D. & Weinberg, R. A. Hallmarks of cancer: the next generation. Cell 144, 646–674 (2011).
Patel, M. S. & Korotchkina, L. G. Regulation of the pyruvate dehydrogenase complex. Biochem. Soc. Trans. 34, 217–222 (2006).
Anderson, N. M., Mucka, P., Kern, J. G. & Feng, H. The emerging role and targetability of the TCA cycle in cancer metabolism. Protein Cell 9, 216–237 (2018).
Byron, O. & Lindsay, J. G. The pyruvate dehydrogenase complex and related assemblies in health and disease. Subcell. Biochem 83, 523–550 (2017).
Vander Heiden, M. G., Cantley, L. C. & Thompson, C. B. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324, 1029–1033 (2009).
Denko, N. C. et al. Investigating hypoxic tumor physiology through gene expression patterns. Oncogene 22, 5907–5914 (2003).
Kim, J. W., Tchernyshyov, I., Semenza, G. L. & Dang, C. V. HIF-1-mediated expression of pyruvate dehydrogenase kinase: a metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 3, 177–185 (2006).
Dang, C. V. The interplay between MYC and HIF in the Warburg effect. In Oncogenes Meet Metabolism. Ernst Schering Foundation Symposium Proceedings Vol. 2007/4 (Eds Kroemer, G. et al.) 35–53 (Springer, 2007); https://doi.org/10.1007/2789_2008_088
Zhang, Y. et al. Estrogen-related receptors stimulate pyruvate dehydrogenase kinase isoform 4 gene expression. J. Biol. Chem. 281, 39897–39906 (2006).
Contractor, T. & Harris, C. R. p53 negatively regulates transcription of the pyruvate dehydrogenase kinase Pdk2. Cancer Res. 72, 560–567 (2012).
Cai, Z. et al. Phosphorylation of PDHA by AMPK Drives TCA Cycle to Promote Cancer Metastasis. Mol. Cell 80, 263–278 e267 (2020).
Sutendra, G. et al. A nuclear pyruvate dehydrogenase complex is important for the generation of acetyl-CoA and histone acetylation. Cell 158, 84–97 (2014).
Swann, J. B. & Smyth, M. J. Immune surveillance of tumors. J. Clin. Invest. 117, 1137–1146 (2007).
Thibaut, R. et al. Bystander IFN-gamma activity promotes widespread and sustained cytokine signaling altering the tumor microenvironment. Nat. Cancer 1, 302–314 (2020).
Inamura, K., Ninomiya, H., Ishikawa, Y. & Matsubara, O. Is the epidermal growth factor receptor status in lung cancers reflected in clinicopathologic features? Arch. Pathol. Lab. Med. 134, 66–72 (2010).
Sui, H. et al. Anti-PD-1/PD-L1 therapy for non-small-cell lung cancer: toward personalized medicine and combination strategies. J. Immunol. Res. 2018, 6984948 (2018).
Wu, Y. & Zhou, B. P. TNF-alpha/NF-kappaB/Snail pathway in cancer cell migration and invasion. Br. J. Cancer 102, 639–644 (2010).
Aggarwal, B. B. Signalling pathways of the TNF superfamily: a double-edged sword. Nat. Rev. Immunol. 3, 745–756 (2003).
Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 9, 361–371 (2009).
Van Antwerp, D. J., Martin, S. J., Verma, I. M. & Green, D. R. Inhibition of TNF-induced apoptosis by NF-kappa B. Trends Cell Biol. 8, 107–111 (1998).
Liu, S., He, L. & Yao, K. The antioxidative function of alpha-ketoglutarate and its applications. BioMed. Res. Int. 2018, 3408467 (2018).
Mailloux, R. J. et al. The tricarboxylic acid cycle, an ancient metabolic network with a novel twist. PLoS ONE 2, e690 (2007).
Sun, W. et al. PPM1A and PPM1B act as IKKbeta phosphatases to terminate TNFalpha-induced IKKbeta-NF-kappaB activation. Cell. Signal. 21, 95–102 (2009).
Pardoll, D. & Allison, J. Cancer immunotherapy: breaking the barriers to harvest the crop. Nat. Med. 10, 887–892 (2004).
Drake, C. G., Jaffee, E. & Pardoll, D. M. Mechanisms of immune evasion by tumors. Adv. Immunol. 90, 51–81 (2006).
Pietrocola, F., Galluzzi, L., Bravo-San Pedro, J. M., Madeo, F. & Kroemer, G. Acetyl coenzyme A: a central metabolite and second messenger. Cell Metab. 21, 805–821 (2015).
Zhang, Y. et al. Macrophage-associated PGK1 phosphorylation promotes aerobic glycolysis and tumorigenesis. Mol. Cell 71, 201–215 e207 (2018).
Wang, X. et al. UDP-glucose accelerates SNAI1 mRNA decay and impairs lung cancer metastasis. Nature 571, 127–131 (2019).
Wu, Y. B. et al. Concurrent quantification of proteome and phosphoproteome to reveal system-wide association of protein phosphorylation and gene expression. Mol. Cell. Proteom.: MCP 8, 2809–2826 (2009).
Zhang, Y. et al. Cul4A-DDB1-mediated monoubiquitination of phosphoglycerate dehydrogenase promotes colorectal cancer metastasis via increased S-adenosylmethionine. J. Clin. Invest. https://doi.org/10.1172/JCI146187 (2021).
Acknowledgements
This work was supported by the National Key Research and Development Programme of China (grant no. 2019YFA0802000) to W.Y.; the National Natural Science Foundation of China (grant nos 91857120 and 32025013) to W.Y.; the CAS Project for Young Scientists in Basic Research (grant no. YSBR-014) to W.Y.; the Chinese Academy of Sciences Interdisciplinary Innovation Team (grant no. JCTD-2018-14) to W.Y.; the Programme of Shanghai Academic/Technology Research Leader (grant no. 20XD1424400) to W.Y.; the CAS Facility-based Open Research Programme to W.Y.; the National Natural Science Foundation of China (grant no. 81902807) to Y. Zhang; the National Postdoctoral Programme for Innovative Talents (grant no. BX20190349) to Y. Zhang; the Shanghai Postdoctoral Excellence Programme (grant no. 2018244) to Y. Zhang; the Youth Innovation Promotion Association of the Chinese Academy of Sciences (grant no. 2022265) to Y. Zhang; and the China Postdoctoral Science Foundation (grant no. 2020M671258) to Y. Zhang. We gratefully acknowledge the support of the SA-SIBS Scholarship Programme and the Heye Scholarship Programme. We thank X. Liu for providing OVA-specific cytotoxic CD8 + T cells. We also thank the Cell Analysis Technology Platform and the Chemical Biology Technology Platform of SIBCB for technical support.
Author information
Authors and Affiliations
Contributions
This study was conceived by W.Y. W.Y. and Y. Zhang designed the study. Y. Zhang performed most of the experiments and also contributed to data analyses and figure editing. M.Z. constructed plasmids, purified proteins and provided experimental assistance. H.G. assisted in reviewing the manuscript. G.Y. and F.Y. provided reagents and pathological assistance. Y. Zhao provided reagents and constructive suggestions. W.Y. wrote the manuscript with comments from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Metabolism thanks David Barbie, Jiyeon Kim and Kathryn O’Donnell for their contribution to the peer review of this work. Primary Handling Editor: Alfredo Giménez-Cassina
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
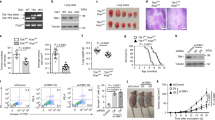
Extended Data Fig. 1 Cytosolic PDHE1α enhances TNFα-induced apoptosis.
a, H1299 cells stably expressing shNT or shPDHE1α were subcutaneously implanted into BALB/c nude mice. IF analyses were performed with an anti-PDHE1α antibody in dissected tumour tissues. b, H1299 or A549 cells were infected with the lentivirus expressing shNT or shPDHE1α. IF analysis with anti-PDHE1α and anti-COX4 antibodies were performed. c, H1299 or A549 cells were harvested and subcellular fractionation assay was performed. d,e, HEK293T cells were infected with the lentivirus expressing PDHE1α D1-30-Flag. Subcellular fractionation assay (d) and IF staining (e) were performed. f-h, H1299, A549 or HEK293T cells were infected with the lentivirus stably expressing EV or PDHE1α-Flag WT or D1-30. PDHE1α-Flag expression was examined by immunoblotting analyses (f). Cells were treated with or without 50 ng/ml TNFα for 24 h. Cells were harvested and stained with FITC-annexin V, followed by flow cytometry analysis (g). Cell proliferation was also examined (h). i-k, H1299 or A549 cells were infected with the lentivirus stably expressing EV or PDHE1α-Flag D1-30. Cells were treated with or without 50 ng/ml IFNγ for 24 h, followed by apoptosis analysis (i). Cells were treated with or without 50 ng/ml TNFα and 50 ng/ml IFNγ for 24 h, followed by apoptosis analysis (j). Cells were treated with or without 20 μM cisplatin for 24 h, followed by apoptosis analysis (k). Rel, relative. Immunoblots are representative of three independent experiments. (g,h) 1-way ANOVA with Tukey’s multiple comparison test. (i-k) Unpaired, two-tailed t-test.
Extended Data Fig. 2 Cytosolic PDHE1α impairs lung cancer development.
a-e, LLC cells stably expressing luciferase were infected with the lentivirus stably expressing EV, mPDHE1α-Flag WT or D1-30. mPDHE1α-Flag expression was examined by immunoblotting analyses (a). Subcellular fractionation assay was performed (b). Cells were subcutaneously implanted into C57BL/6J mice (6 mice per group), tumor growth over time was measured (c). Tumor weight was measured at day 11 (d). TUNEL assay of dissected tumors was performed and the percentage of apoptotic tumor cells was shown (e). Data represent the means ± SD of six mice. f-h, Cells in (a) were injected into lung of randomized C57BL/6J mice (6 mice per group). 14 days after inoculation, mice were euthanized. IHC staining of dissected tumors using anti-F4/80, anti-CD8 or anti-TNFα antibody was performed. Representative images (top) and semiquantitative analysis (bottom) of IHC staining were shown. Data represent the means ± SD of six mice. Immunoblots are representative of three independent experiments. (c-h) 1-way ANOVA with Tukey’s multiple comparison test.
Extended Data Fig. 3 ERK2 phosphorylates PDHE1α to promote its translocation.
a, A549 cells were treated with or without 20 ng/ml EGF for 30 min. Subcellular fractionation assay was performed. b, H1299 cells were treated with or without no glucose DMEM or 200 μM CoCl2 for 24 h. Subcellular fractionation assay was performed. c, H1299 or A549 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT, S327A, S327D, S331A or S331D. d, A549 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT, S327A, S327D, S331A or S331D. Cells were treated with or without 20 ng/ml EGF for 30 min and subcellular fractionation assay was performed. e, Mass spectrometry analysis of PDHE1α D1-30-associated proteins were performed in H1299 cells. The top 10 protein kinase that showed strong interactions with PDHE1α D1-30 were listed. f, H1299 cells were infected with the lentivirus stably expressing shNT or shERK2. g, H1299 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α-Flag WT or S327A. PDHE1α-Flag proteins were immunoprecipitated using anti-Flag agarose beads. IP, immunoprecipitation. h, H1299 or A549 cells stably expressing PDHE1α-Flag were infected with the lentivirus stably expressing shNT or shERK2. i, A549 cells stably expressing PDHE1α-Flag were infected with the lentivirus stably expressing shNT or shERK2. PDHE1α-Flag proteins were immunoprecipitated using anti-Flag agarose beads. Immunoblots are representative of three independent experiments.
Extended Data Fig. 4 Oncogenic signaling induces PDHE1α pS327 and promotes its translocation.
a, EGFR WT (HTB182, H1299, H292 and A549) or EGFR mutant/active (PC9, H3255, HCC827 and H1650) lung cancer cells were collected. Cell fraction assay was performed. b, EGFR and KRAS mutation status were shown. c-e, H1299 cells were transfected with EV, KRAS G12V (c), MET-H1094R (d) or ERBB2 G776VC (e) with or without PDHE1α-HA. PDHE1α-HA proteins were immunoprecipitated using anti-HA agarose beads (left panel). Cell fraction assay was performed (right panel). Immunoblots are representative of three independent experiments.
Extended Data Fig. 5 PDHE1α pS327 promotes tumor cell resistance to CTLs’ killing.
a, A549 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT, S327A or S327D. Cells were treated with or without 50 ng/ml TNFα for 24 h. Cells were harvested for apoptosis analysis. b-d, H1299 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT, S327A or S327D. Cells were treated with or without 50 ng/ml TNFα and 50 ng/ml IFNγ for 24 h and then cells were harvested for apoptosis analysis (b). Proliferation was examined with these cells (c). Cells were harvested for PDH activity assay (d). e, LLC cells stably expressing ovalbumin (OVA) were depleted of endogenous mPDHE1α and rescued with rmPDHE1α WT, S327A or S327D. f-i, LLC cells stably expressing luciferase were depleted of endogenous mPDHE1α and rescued with rmPDHE1α WT, S327A or S327D. PDHE1α expression was examined by immunoblotting analyses (f). Cells were injected into lung of randomized C57BL/6J mice (6 mice per group). 14 days after inoculation, mice were euthanized. IF analysis with anti-Flag and COX4 antibodies were performed in lung sections from these mice. The levels of cytosolic PDHE1α in tumors were compared (g). IHC staining of dissected tumors using anti-F4/80 or anti-CD8 antibodies was performed. Semiquantitative analysis of IHC staining were shown (h,i). Data represent the means ± SD of six mice. (a-d, g-i) 1-way ANOVA with Tukey’s multiple comparison test.
Extended Data Fig. 6 Inhibiting PDHE1α pS327 improves the efficacy of ICB therapy.
a, LLC cells stably expressing luciferase were depleted of endogenous mPDHE1α and rescued with rmPDHE1α WT, S327A or S327D. Cells were injected into lung of randomized C57BL/6J mice (6 mice per group). These mice were treated with control IgG2a or anti-PD-1 antibody by intraperitoneal injection. 14 days after inoculation, mice were euthanized. IHC staining of dissected tumors using anti-CD8 antibody were performed. Representative images of IHC staining were presented (left). Semi-quantitative analyses were performed (right). Data represent the means ± SD of six mice. b,c, LLC cells stably expressing luciferase were depleted of endogenous mPDHE1α and rescued with rmPDHE1α WT, S327A or S327D. Cells were subcutaneously implanted into C57BL/6J mice (6 mice per group). These mice were treated with control IgG2a or anti-PD-1 antibody by intraperitoneal injection. Tumor growth over time was measured (b). Tumor weight was measured at day 11 (c). Data represent the means ± SD of six mice. (a-c) Unpaired, two-tailed t-test.
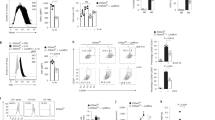
Extended Data Fig. 7 PDHE1α pS327 activates NF-κB and sustains ROS homeostasis.
a, H1299 cells were infected with the lentivirus expressing EV or PDHE1α D1-30-Flag. Cells were treated with 20 ng/ml TNFα for indicated time. b, A549 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT, S327A or S327D. Cells were treated with or without 20 ng/ml TNFα for 10 min. c, DNA sequencing of genomic DNA from H1299 cells with or without PDHE1α S327A knockin. d, HEK293T cells were co-transfected with EV or PDHE1α D1-30-HA and NF-κB reporter. Cells were treated with 10 ng/ml TNFα for 3 h. The relative levels of luciferase activity of Firefly were normalized to the levels of luciferase activity of control Renilla. e, H1299 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT or S327A. Cells were then transfected with EV or HA-p65. f,g, H1299 cells stably expressing EV or PDHE1α D1-30-Flag were transfected with EV or HA-p65. Immunoblotting analysis (f) and apoptosis analysis (g) were performed. h, LLC cells stably expressing luciferase were depleted of endogenous mPDHE1α and rescued with rmPDHE1α WT or S327A. Cells were infected with the lentivirus expressing EV or HA-mp65. i, H1299 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT or S327A. Cells were treated with or without 50 ng/ml TNFα for 24 h. Cells were harvested and subjected to real-time qPCR. j, H1299 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT or S327A. Cells were then transfected with EV or HA-BCL2A1. k, H1299 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT, S327A or S327D. Cells were treated with 20 ng/ml TNFα for 1 h and mitochondria ROS was measured using MitoSOX. Immunoblots are representative of three independent experiments. (d,g) Unpaired, two-tailed t-test. (k) 1-way ANOVA with Tukey’s multiple comparison test.
Extended Data Fig. 8 NF-κB and α-KG are required for PDHE1α pS327-inhibited apoptosis.
a-f, H1299 cells were depleted of endogenous PDHE1α with another shRNA (shPDHE1α-2) and rescued with rPDHE1α WT, S327A or S327D (a). Cells were treated with or without 20 ng/ml TNFα for 10 min and immunoblotting analysis was performed (b). Cells were transfected with EV or HA-p65 and treated with 50 ng/ml TNFα for 24 h. Cell apoptosis was measured (c,d). Cells were harvested for the measurement of α-KG levels (e). Cells were treated with or without 0.5 mM octyl-α-KG and 50 ng/ml TNFα for 24 h. Then cells were harvested for apoptosis analysis (f). (d-f) Unpaired, two-tailed t-test.
Extended Data Fig. 9 Cytosolic PDHE1α facilitates IKKβ dephosphorylation via PPM1B.
a, H1299 cells were co-transfected with EV or SFB-IKKβ, HA-PPM1B and PDHE1α-Flag. Cells were treated with 10 ng/ml TNFα for 10 min. b, H1299 or A549 cells depleted of endogenous PDHE1α and rescued with rPDHE1α WT or S327A were infected with the lentivirus expressing shNT or shPPM1B. c, H1299 or A549 cells stably expressing EV or PDHE1α D1-30-Flag were infected with the lentivirus expressing shNT or shPPM1B. d, Cells in (c) were treated with 20 ng/ml TNFα for 10 min and immunoblotting analysis was performed. e, H1299 cells depleted of endogenous PDHE1α and rescued with rPDHE1α WT or S327A were infected with the lentivirus expressing shNT or shPPM1B. Cells were treated with 20 ng/ml TNFα for 10 min. Cell fraction assay was performed. f, A549 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT or S327A. Cells were then infected with the lentivirus expressing shNT or shPPM1B. Cells were treated with 50 ng/ml TNFα for 24 h. Cells were harvested for apoptosis analysis. g, H1299 or A549 cells stably expressing EV or PDHE1α D1-30-Flag were infected with the lentivirus expressing shNT or shPPM1B. Cells were treated with 50 ng/ml TNFα for 24 h. Cells were harvested for apoptosis analysis. Immunoblots are representative of three independent experiments. (f,g) Unpaired, two-tailed t-test.
Extended Data Fig. 10 PPM1B is required for cytosolic PDHE1α-inhibited tumor growth.
a-c, LLC cells stably expressing luciferase were infected with the lentivirus expressing EV or mPDHE1α D1-30-Flag and shNT or shmPPM1B (a). Cells were subcutaneously implanted into C57BL/6J mice (6 mice per group), tumor growth over time was measured (b). Tumor weight was measured at day 11 (c). Data represent the means ± SD of six mice. d, H1299 cells were depleted of endogenous PDHE1α and rescued with rPDHE1α WT, S327A or S327D. Cells were co-transfected with EV or SFB-IKKβ and HA-PPM1B. Cells were treated with 10 ng/ml TNFα for 10 min. Cell fraction assay was performed and SFB-IKKβ proteins were pulldown using streptavidin agarose beads. Immunoblots are representative of three independent experiments. (b,c) Unpaired, two-tailed t-test.
Supplementary information
Supplementary Information
Supplementary Figs. 1–3.
Supplementary Data 1
PDHE1α-Flag was immunoprecipitated from H1299 cells treated with or without 20 ng ml−1 EGF for 30 min. The S327 and S331 residues in PDHE1α were identified to be phosphorylated after EGF treatment by mass spectrometry analysis.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 1
Unprocessed western blots.
Source Data Fig. 2
Unprocessed western blots.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 4
Unprocessed western blots.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 5
Unprocessed western blots.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 1
Unprocessed western blots.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 2
Unprocessed western blots.
Source Data Extended Data Fig. 3
Unprocessed western blots.
Source Data Extended Data Fig. 4
Unprocessed western blots.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 5
Unprocessed western blots.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 7
Unprocessed western blots.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 8
Unprocessed western blots.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 9
Unprocessed western blots.
Source Data Extended Data Fig. 10
Statistical source data.
Source Data Extended Data Fig. 10
Unprocessed western blots.
Rights and permissions
About this article

Cite this article
Zhang, Y., Zhao, M., Gao, H. et al. MAPK signalling-induced phosphorylation and subcellular translocation of PDHE1α promotes tumour immune evasion. Nat Metab 4, 374–388 (2022). https://doi.org/10.1038/s42255-022-00543-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-022-00543-7
This article is cited by
-
A paradigm shift in cancer research based on integrative multi-omics approaches: glutaminase serves as a pioneering cuproptosis-related gene in pan-cancer
BMC Women's Health (2024)
-
Disparate macrophage responses are linked to infection outcome of Hantan virus in humans or rodents
Nature Communications (2024)
-
Comprehensive Analysis of the SUMO-related Signature: Implication for Diagnosis, Prognosis, and Immune Therapeutic Approaches in Cervical Cancer
Biochemical Genetics (2024)
-
Roles of protein post-translational modifications in glucose and lipid metabolism: mechanisms and perspectives
Molecular Medicine (2023)
-
ZNF582 overexpression restrains the progression of clear cell renal cell carcinoma by enhancing the binding of TJP2 and ERK2 and inhibiting ERK2 phosphorylation
Cell Death & Disease (2023)
