
Abstract
Tuberculosis (TB), caused by Mycobacterium tuberculosis, ranks among the top causes of global human mortality, as reported by the World Health Organization’s 2022 TB report. The prevalence of M. tuberculosis strains that are multiple and extensive-drug resistant represents a significant barrier to TB eradication. Fortunately, having many completely sequenced M. tuberculosis genomes available has made it possible to investigate the species pangenome, conduct a pan-phylogenetic investigation, and find potential new drug targets. The 442 complete genome dataset was used to estimate the pangenome of M. tuberculosis. This study involved phylogenomic classification and in-depth analyses. Sequential filters were applied to the conserved core genome containing 2754 proteins. These filters assessed non-human homology, virulence, essentiality, physiochemical properties, and pathway analysis. Through these intensive filtering approaches, promising broad-spectrum therapeutic targets were identified. These targets were docked with FDA-approved compounds readily available on the ZINC database. Selected highly ranked ligands with inhibitory potential include dihydroergotamine and abiraterone acetate. The effectiveness of the ligands has been supported by molecular dynamics simulation of the ligand–protein complexes, instilling optimism that the identified lead compounds may serve as a robust basis for the development of safe and efficient drugs for TB treatment, subject to further lead optimization and subsequent experimental validation.
Similar content being viewed by others

Introduction
Tuberculosis (TB) proved a challenging global challenge, claiming 1.6 million lives, including 187,000 individuals grappling with TB and HIV. TB was ranked the world’s second-most severe infectious disease after COVID-19 and ultimately the thirteenth most prevalent cause of mortality. Significantly, throughout the year, there were 1.4 million deaths from TB among individuals without HIV and 187,000 deaths among those with HIV, adding to the total fatalities. This figure represents a rise compared to previous years, in line with the levels observed in 2017, underscoring the ongoing urgency of addressing TB within the broader context of global health1.
TB primarily spreads through airborne transmission, with M. tuberculosis infiltrating the human body and subverting host immune defenses. Within the host, the bacteria adeptly invade macrophages, establishing a prolonged presence that leads to chronic infection, a manifestation of compromised host immunity2. M. tuberculosis strains are increasingly becoming multi and extensively drug resistant (MDR & XDR) as the genes of the bacteria naturally evolve, making them more challenging against the medicines typically employed for tuberculosis treatment. Consequently, these mutated strains emerge as dominant players in the population, outcompeting their more susceptible counterparts. This phenomenon underscores the critical role of genetic adaptation in the evolution of drug resistance, presenting a substantial challenge in managing and controlling TB. Consequently, the efficacy of conventional anti-TB medications such as isoniazid and rifampicin has been compromised3.
Advancements in genomics and other fields have made it possible to more fully comprehend the functional importance of many distinct proteins encoded within the M. tuberculosis genome since the first genome sequencing of M. tuberculosis in 19984,5. The pangenome concept can now be thoroughly investigated, especially to identify the core genome, denoting the set of genes consistently identified across all strains within the dataset6. Utilizing these genes presents an opportunity to develop comprehensive therapeutic interventions targeting pathogenic species across a broad spectrum2.
Bioinformatics tools are essential in systematically analyzing pathogen data for potential drug targets in drug development. Similarly, advances in cheminformatics, like those shown by resources like DrugBank, allow for efficient virtual screening of ligand-target binding affinities. Computer-aided drug discovery (CADD) accelerates precision in identifying therapeutic candidates within the complex landscape of pathogenic biology7,8. Scientists have made significant efforts to find promising compounds for TB by utilizing these invaluable resources8,9. Hence, for the efficient prioritization of potential drug targets in the M. tuberculosis genome, an integrative genomics approach was adopted in this study that included pangenome analyses, subtractive proteomics, and CADD. Moreover, using molecular docking, the structural interactions between these potential targets and FDA-approved drugs were investigated. Employing this method led to the identification of a group of promising lead molecules, marking notable progress in the effort to develop drugs against TB.
The traditional drug development method requires at least ten years of in-depth study and significant funds. However, integrating computer-aided analyses during the initial phases can significantly reduce this endeavor’s time and financial investments10,11. In drug discovery, in silico drug screening is a valuable technique, allowing for the focused selection of compounds with the most significant relevance for subsequent experimental investigations. This approach streamlines the research process by eliminating biomolecules that do not meet the necessary criteria, contributing to the refinement and enhancement of drug discovery efforts as a whole12. All the identified high-ranking potential targets were analyzed rather than focusing exclusively on a single drug target. This study can potentially establish a solid basis for developing innovative therapies for the challenging bacterial pathogen M. tuberculosis.
Materials and methods
Collection of genomic data
The genome sequences of all strains of M. tuberculosis currently in existence, along with their corresponding proteomes, were obtained from the PATRIC database (accessed on May 26, 2023; accessible at https://www.bv-brc.org/)13. These strains were collected from different parts of the world. Pan-genome analysis was made possible by the subsequent integration of these genomic data.
Pangenome analysis of M. tuberculosis strains
The bacterial pan-genome analysis (BPGA) tool was used to perform pangenome analysis on retrieved genomes to find highly conserved proteins from M. tuberculosis. The default 50% identity threshold and the USEARCH algorithm were used to produce orthologous protein clusters. BPGA computed the pan and core proteome sizes by taking into account 20 permutations and gradually calculating median values as each genome was added. A ratio of the total number of unique and common gene families to the total number of genomes was used to establish the core and pan-genome curves graphically. The results also gave insights into how each genome’s inclusion led to the emergence of new genes14. To identify potential therapeutic targets, the analysis was further focused on protein sequences associated with the core genome.
Identification of non-host homologous, essential, and virulence-associated proteins
PanRV was used to identify new therapeutic targets within the core proteome. This strategy includes several screening stages, such as filters for non-homology, virulence, and essentiality15. Ensuring the safety of drug therapy in humans and minimizing potential adverse effects necessitates identifying drug targets distinct from human proteins. The core genome was compared to the human genome using the NCBI BLASTp tool (with an e-value 1e−5)16. Essential genes were determined from the subset of non-host homologous conserved proteins by conducting BLASTp searches against the database of essential genes (DEG) with a focus on stringent criteria (e-value < 0.0001 and a bit score > 100)17. DEG compiles data from eukaryotes, bacteria, and archaea that have been experimentally verified. This information contains essential genomic components required for a cell’s survival. The selected proteins were then subjected to BLASTp searches within the virulence factor database (VFDB) to find potential proteins linked to virulence18. The VFDB is a substantial online database that details bacterial pathogens’ virulence characteristics. Virulence factors are gene products that enhance a bacterium’s ability to cause disease19.
Identification of putative M. tuberculosis drug targets
Physiochemical properties were used to refine filtered proteins further and find potential drug targets. The molecular weight, isoelectric point, GRAVY (grand average of hydropathicity) value, instability index, and aliphatic index were considered using the ProtParam tool (found at https://web.expasy.org/protparam/)20. Proteins with low molecular weight are particularly favorable as drug targets, given their susceptibility to drug interactions21. A higher aliphatic index value indicates enhanced thermostability, whereas a lower GRAVY value signifies that proteins are hydrophilic22. After being physiochemically characterized, a comparative pathway analysis was used to analyze the filtered proteins further. The KEGG Automatic Annotation Server (KAAS) version 2.1 was used to identify proteins linked to pathogen-specific pathways. This strategic method improves the possibility of precisely targeting the pathogen23.
Finally, the selected proteins druggability was evaluated using BLASTp with default parameters against the Drug Bank database (https://go.drugbank.com/)24. The draggability of possible targets is another vital screening stage that assesses the likelihood that a drug will regulate selected targets25. Moreover, all the FDA-approved ligands from the ZINC database (accessed at https://zinc.docking.org/ on 23, August 2023) were retrieved in SDF format and checked against identified proteins which were found druggable earlier in druggability analysis26.
Molecular docking of putative drug targets with drugs
The sequences of the identified drug targets were examined for the presence of corresponding crystal structures in the RCSB Protein Data Bank (accessible at https://www.rcsb.org/)27. The relevant PDB files were obtained from the RCSB protein databank, and any attached ligands were taken out of the crystal structures if they were there. The structural configurations of the drug targets were molecularly docked with the drugs previously identified in the draggability analysis using PyRx’s AutoDock Vina tool28,29. The grid box was modified during the molecular docking process to enclose the protein space entirely, and blind docking was conducted, selecting a value of 32 for exhaustiveness. The targets and the corresponding ligands were prepared in the PDBQT format for molecular docking analysis. The visualization of docked complexes and the interactions formed was performed in Discovery Studio Visualizer. To evaluate the effectiveness of the drug, both the binding affinity \(E\) (in kcal/mol) from AutoDock Vina and the fit quality score are used30, where the fit score is defined as follows:
In these equations, \(HA\) is the number of heavy (non-hydrogen) atoms, \(LE\) is the ligand efficiency, \(LE\_Scale\) is the scaling factor, and \(FQ\) is the fit quality score. An \(FQ\) of near or larger than 1 indicates that the ligand binding is optimal, while a small \(FQ\) represents a sub-optimal binding. A comparative study of the docked complexes with the lowest binding energies was performed to identify ligands strongly linked to the drug targets. As a result, these preferred ligands were chosen for later molecular dynamic simulations.
Molecular dynamics simulation
Molecular dynamics (MD) simulation is a powerful computational technique used to study the behavior and properties of molecules and materials at the atomic level. Desmond software package was used for MD simulations of biological systems, including proteins, nucleic acids, and membranes.
The Desmond module of Schrodinger was exploited to conduct the MD simulation studies. The dynamic behavior and stability of the protein–ligand complex were investigated using its docked poses. The protein–ligand complex was preprocessed using the Protein Preparation Wizard of Maestro, which included complex optimization and minimization. All of the systems were prepared using the System Builder tool. The solvation of the complexes was performed with the simple point-charge (SPC) water model with an orthorhombic box, along with a 10-Å distance from the edge of the box, and the system was neutralized with Na+/Cl− ions. To mimic the physiological conditions, 0.15 M sodium chloride (NaCl) was added. The potential energy of the protein complex was minimized by employing the NPT ensemble. The molecular dynamics simulations were performed at 300 K temperature and 1 atm pressure for 100 ns, and NPT production ran under the OPLS4 force field. The models were relaxed before the simulation. The short-range electrostatic interactions were calculated using the particle mesh Ewald method31. The cutoff radius in the Coulomb interactions was 9.0 Å. The water molecules were explicitly described using the simple point charge model32. The Martyna–Tuckerman–Klein chain coupling scheme with a coupling constant of 2.0 ps was used for the pressure control, and the Nosé–Hoover chain coupling scheme33 was used for the temperature control. The trajectories were saved for examination after every 100 ps, and the simulation’s stability was verified by comparing the root mean square deviation (RMSD) of the protein complex over time. The projected changes in their conformation from the initial structure over the entire simulation period were expressed as root mean square deviation (RMSD) and root mean square fluctuation (RMSF) for MD simulations. A detailed description of the methodology can also be found elsewhere34,35,36,37. The radius of gyration (Rg) is calculated using GROMACS38. The Desmond trajectory is first converted to XTC (GROMACS) format via MDTraj39, and the calculation is conducted using the gmx gyrate command.
Principal component analysis (PCA)
Finding coordinated motions in a group of conformational structures obtained through experimental or molecular simulation data has been shown to be a useful application of principal component analysis (PCA). This method has been extensively employed to investigate the impact of conformational modifications on receptor function. We created the covariance matrix for PCA using the C-alpha atomic coordinates that we had collected from the molecular dynamic simulations. After that, a set of eigenvalues and eigenvectors (principal components) is obtained by diagonalizing this matrix. The coordinates of atoms are mapped to the principal components. GROMACS38 is utilized for this purpose using the XTC format of trajectory. The generation of covariance matrices and their diagonalization is achieved by the gmx covar command. The projection of the atomic coordinates to the principal components is done by the gmx anaeig command with the -proj parameter.
Free energy landscape (FEL)
Free energy landscape (FEL) is used to measure the change in energy during MD simulation of the protein as a function of principal components generated in PCA. This allows us to identify the metastable conformations of the protein in different systems and their relative energies. The energy function used is the Gibbs free energy. In this work, we used GROMACS38 for the analysis of FEL. The Gibbs free energy as a function of the first two principal components, PC1 and PC2, is evaluated using the gmx sham command. Locally estimated scatterplot smoothing (LOESS) regression is employed to smooth the FEL plots. The FEL plots are also verified by kernel density estimation (KDE) plots generated from the PCA results.
Dynamic cross-correlation matrix (DCCM)
Dynamic cross-correlation matrix (DCCM) can capture the correlation in motions of different residues. The motions between two residues can be correlated or anticorrelated, so a network of cross-correlation among residues can be formed40. We used the motions of the C-alpha atoms to represent those of the residues. The correlation between two residues \(i\) and \(j\) is measured by the cross-correlation coefficient \(C\left(i,j\right)\):
where \({\varvec{\Delta}}{{\varvec{r}}}_{{\varvec{i}}}\) is the displacement vector of the C-alpha atom of the \(i\)th residue, and the angle brackets represent the average values of the quantities inside them. \(C\left(i,j\right)>0\), \(C\left(i,j\right)<0\) and \(C\left(i,j\right)=0\) are the indication of positive correlation, negative correlation (anti-correlated) and no correlation (uncorrelated), respectively.
In this work, we converted the Desmond trajectory to DCD format with MDTraj39 and we used the dccm function in the Bio3D package41,42 in R language to calculate DCCM.
MM/GBSA binding free energy of complex
Molecular mechanics/generalized Born surface area (MM/GBSA) is for the calculation of free energy of binding (\(\Delta {G}_{bind}\)) for the docked postures. It is defined as follows:
where the energy estimates of the optimized complex (complex), optimized free ligand (ligand), and optimized free receptor (receptor) are represented by the \({G}_{complex}\), \({G}_{ligand}\), and \({G}_{receptor}\), respectively.
Since MM/GBSA is more accurate than most molecular docking scoring systems, it is without a doubt one of the most used techniques for predicting binding free energy. Since MM/GBSA requires less computing power than other chemical free energy scoring methods, it is frequently employed in bimolecular studies involving protein folding, protein–ligand binding, and protein–protein interaction, among other topics. The accurate computation of the free energy, which powers molecular motions, is one of the most important issues in bimolecular studies. Using the MM/GBSA approach, it is possible to calculate the binding free energy by utilizing the solvation free energy differential upon binding and the interaction energy between the ligand and receptor complexes.
In this work, we used Uni-GBSA43 for the calculation of MM/GBSA binding energy. Uni-GBSA uses gmx_MMPBSA44 and MMPBSA.py in Amber45. After removing water molecules and ions, the frames in Desmond trajectory are split into separate PDB files. The pdbsplit function in Bio3D41 is used to separate the protein and the ligand into two PDB files. The ligand PDB files are converted to MOL2 format using Open Babel46. The protein PDB and the ligand MOL2 files are used as input for Uni-GBSA. The default configuration file (default.ini) is used except that the input pose process method is set as “input”, which means that the original input pose is used without energy minimization.
Results and discussion
Pangenome and pan-phylogeny analysis of M. tuberculosis genomes
A total of 442 complete M. tuberculosis genomes and their associated proteomes were obtained from the PATRIC database. Supplementary Table S1 provides comprehensive data, including accession numbers, strain names, genome statistics, isolation sources, the countries of isolation, and other information. A comprehensive pangenome analysis of these 442 complete M. tuberculosis genomes determined 5809 genes within the pangenome. Among these, 2754 proteins were identified as a core genome, representing genes consistently present across all studied genomes. The calculated pangenome size to core genome size ratio was 0.474, indicating that the core genome constitutes 47.4% of the entire pangenome. This observation underscores the limited genetic diversity observed among M. tuberculosis strains. This trend is graphically illustrated by the pan-core genome plot (Fig. 1), which also suggests that the global gene repertoire of this species is likely to remain relatively stable and resistant to significant changes in the future.

The pan-core plot, constructed using 442 M. tuberculosis genomes, demonstrates how the pangenome size increases while the core genome size decreases as genomes are added. It is essential to highlight that the pangenome curve reaches a level (in brown), signaling that significant alterations in the overall gene pool of this species are improbable in the future, suggesting that the pangenome is nearly closed, with limited potential for substantial genetic variations in M. tuberculosis.
The power-law regression model’s “b” value of 0.121 shows that the pangenome is open. However, this value also suggests it may gradually transition towards a closed pangenome soon. Each genome, on average, contained approximately 3727 protein-encoding genes. The core genome, consisting of 2754 genes, accounted for a significant percent of the average genome size, 73.9%. Strain M0018684.2 had the lowest number of protein-encoding genes (3680), while strain MYC004 had the highest number (4462). Pan-matrix data was used to create the pan phylogeny, and the associated pangenome tree was produced using the neighbor-joining method with a default iteration value of 20. Figure 2 includes the pan-phylogeny-based phylogenetic tree and geographic source data to provide more information about the connections between these genomes.

The pan-phylogeny tree, showcasing 442 M. tuberculosis genomes, is color-coded to represent the continents of origin for these strains. Interestingly, strains from different global regions tend to cluster into distinct branches within the tree. This tree is based on the presence or absence of accessory genes. It offers insights into the genetic relationships and evolutionary history of these M. tuberculosis strains, emphasizing the influence of geography on their genomic characteristics.
Subtractive proteomics revealed putative M. tuberculosis drug targets
Finding potential therapeutic targets started with subtractive proteomics performed on M. tuberculosis core proteins. This analysis involved assigning Rv numbers (gene identifiers specific to M. tuberculosis) and gene annotations to these core proteins. Supplementary Table S2 contains the dataset associated with the core genomes. A careful screening process was implemented to ensure the exclusion of proteins from human homologs, as human homologs could potentially interfere with host metabolism. Among the identified proteins, 2386 were classified as non-human homologous. These non-human homologs were subjected to further scrutiny, specifically assessing their essentiality. Among this subset, 685 proteins emerged as essential components for the pathogen’s survival. When essential proteins also exhibit functional characteristics associated with virulence, they become particularly noteworthy. Such proteins are pivotal in enabling bacteria to manipulate or break down host defense mechanisms, potentially contributing to pathogenesis47.
Consequently, the focus shifted to identifying genes linked to pathogenicity within the pool of 685 essential proteins. The VFDB has identified 113 virulence-related proteins necessary for M. tuberculosis pathogenicity (Supplementary File S1). Surprisingly, all 113 proteins are essential components linked to virulence and lack human homologs. This dataset has much potential and can be investigated for projects like creating a TB vaccine and novel therapeutic therapies. To specifically meet the objectives of this study, the size of the dataset was further reduced to pinpoint a few potential drug targets. This procedure makes designing drugs specifically suited to combat M. tuberculosis easier.
After extensive physiochemical analyses that included criteria like low molecular weights, a high aliphatic index, and a negative GRAVY score, 38 proteins were identified (Supplementary File S2). This group’s only six proteins were pinpointed through a comparative pathway analysis as part of distinct bacterial metabolic pathways. These six proteins were then subject to further scrutiny. Interestingly, among these proteins, those that played roles in multiple pathways (specifically, more than three pathways) were deliberately selected as potential drug targets for M. tuberculosis. This selection approach was taken to minimize the risk of inadvertently targeting any human pathways during drug therapy, as detailed in Table 1.
In the final stage of our analysis, the top 2 proteins (isocitrate lyase and pantothenate synthetase) were focused. The similarity of these targets with drugs in the DrugBank database was evaluated. This step aimed to determine whether these proteins are potentially druggable. Our investigation revealed that the selected targets were druggable with experimental drugs available in the drug bank database but not with FDA-approved drugs. This finding suggests that these proteins could serve as promising novel drug targets. All FDA-approved drugs (n = 1613) were retrieved from the ZINC database, and docking experiments were conducted with the selected proteins to explore their druggability further. The ligands with the lowest binding scores were then determined, and their fit quality scores were also evaluated (Table 2). The fit quality scores are near or exceed 1, which is indicative of optimal binding.
On these two proteins from Table 2, literature studies were done to determine their applicability as drug targets. Isocitrate lyase is a vital enzyme in the metabolism of M. tuberculosis. Importantly, they are absent in mammals, making them valuable targets for TB treatment48. Isocitrate lyase is pivotal in the glyoxylate cycle, a metabolic pathway bypassing several Krebs cycle steps. It converts isocitrate into succinic acid and glyoxylate, which generate energy49,50. Additionally, pyruvate is converted to alanine by the upregulated alanine dehydrogenase, while glycine dehydrogenase converts glyoxylate into glycine while oxidizing NADH to NAD51. Glyoxylate and pyruvate are produced by the glyoxylate, and the methyl citrate cycles, respectively, which are necessary building blocks for these reactions. These cycles are catalyzed by two isomers of isocitrate lyase, ICL1 and ICL2, encoded by the genes icl and aceA, respectively52. Notably, when either ICL or aceA genes are deleted, M. tuberculosis exhibits reduced virulence and impaired survival in activated macrophages53. M. tuberculosis cannot survive in the latent TB model without isocitrate lyase. Due to the host’s absence of these metabolic pathways, M. tuberculosis isocitrate lyase is a desirable drug target for treating latent bacteria. Consequently, isocitrate lyase has gained significant attention as a potential target for future research and drug development54,55,56.
Pantothenate synthetase in M. tuberculosis is a crucial enzyme in pantothenate biosynthesis. Its crystal structure reveals that AMP and β-alanine are involved in its active site, with β-alanine binding after pantoyl adenylate formation57,58. Pantothenate synthetase is crucial for M. tuberculosis survival in the host, and recent findings show that pyrazinamide, a common antitubercular drug, also inhibits pantothenate biosynthesis. Confirmation of this mechanism of action is provided by mutations in the panD gene identified in pyrazinamide-resistant MTB strains. This finding highlights the pantothenate biosynthesis pathway as a potentially effective target for developing new TB drugs59,60,61,62. These enzymes may be suitable for designing or selecting inhibitors because they are not found in mammals. Given these insights, Pantothenate synthetase emerges as a promising antimycobacterial target, potentially effective against non-replicating persistent forms of MTB63,64.
Docking analyses of drug targets revealed potential lead compounds for drug discovery
The results of the AutoDock Vina docking of ligands with drug targets are shown in Table 2. Dihydroergotamine (ZINC000003978005) is the highest-ranked ligand for isocitrate lyase. The binding energy of this substance to isocitrate lyase was − 10.3 kcal/mol (Fig. 3a). ZINC000164528615 ligand shows the second highest binding affinity (− 10 kcal/mol) with isocitrate lyase, followed by Ergotamine (ZINC000052955754), which shows − 9.9 kcal/mol binding affinity.

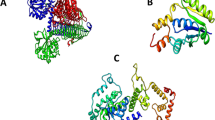
The docked complex of dihydroergotamine and isocitrate lyase.
Dihydroergotamine (ZINC3978005) exhibited the highest binding affinity (− 10.3) for isocitrate lyase. Moreover, the docking interactions that formed for this complex demonstrated various types of bonds, such as conventional hydrogen bonds, carbon hydrogen bonds, pi-sigma, pi-pi T-shaped, amide-Pi stacked, alkyl, and pi-alkyl bonds. The aminoacids in contact with these bonds were leucine, tyrosine, alanine, glycine, and arginine (Fig. 3b). These variety of interactions suggests a strong and stable binding between dihydroergotamine and isocitrate lyase65. The presence of multiple types of bonds indicates a complex and intricate molecular interaction, potentially leading to a more efficient inhibition of the enzyme’s activity66. Overall, the detailed analysis of the docking interactions provides valuable insights into the mechanism of action of dihydroergotamine as a potential inhibitor of isocitrate lyase. By targeting this enzyme, dihydroergotamine may offer a promising approach for developing new antibiotics to address bacterial infections67.
Dihydroergotamine has the highest binding affinity with isocitrate lyase and is an FDA-Approved drug mainly used to treat cluster and migraine headaches. The drug belongs to the subclass of ergot alkaloids. Dihydroergotamine treats migraine and cluster headaches by constricting blood vessels in the brain to relieve pain68. DHE can be administered via several routes, including intravenous, intramuscular, subcutaneous, intranasal, and oral. Studies have shown that dihydroergotamine can effectively relieve migraine pain and associated symptoms like nausea and sensitivity to light and sound69,70. It is particularly useful in cases of severe or refractory migraines. Dihydroergotamine is contraindicated in patients with uncontrolled hypertension, coronary artery disease, peripheral vascular disease, and hepatic or renal impairment. It should not be used concomitantly with other ergot alkaloids or potent CYP3A4 inhibitors. Due to its vasoconstrictive properties, DHE should be used cautiously in patients with cardiovascular risk factors. Regular monitoring of blood pressure and cardiac function may be necessary during treatment. DHE can interact with several medications, including certain antidepressants, antifungals, and protease inhibitors71. These interactions can affect the metabolism and effectiveness of dihydroergotamine, necessitating dose adjustments or alternative treatments.
Analysis of drug-ligand interactions showed that Abiraterone acetate/Zytiga (ZINC000003797541) had a higher binding energy (− 10.4 kcal/mol) to pantothenate synthetase (Fig. 4a). Another ligand, Methotrexate (ZINC000001529323), shows the second highest binding affinity (− 10.2 kcal/mol), followed by ZINC000000607986, showing − 10.1 kcal/mol binding affinity.

The docked complex of abiraterone acetate and pantothenate synthetase.
Abiraterone acetate—pantothenate synthetase docked complex showed interactions of pi-pi-T-shaped, alkyl and pi-alkyl bonds in contacts with different amino acids of the receptor protein, such as lysine, histidine, leucine, methionine, phenylananine and proline (Fig. 4b). This suggests that Abiraterone acetate/Zytiga could be a promising lead compound for drug discovery targeting pantothenate synthetase. The many amino acids interacting indicates that the drug-ligand complex has the potential for strong and specific binding, which is crucial for effective drug design72,73. Further research and development on this interaction could lead to the development of new and improved treatments for diseases or conditions related to pantothenate synthetase. The diverse nature of the interactions also suggests that Abiraterone acetate/Zytiga may have a multi-targeted approach, making it a versatile candidate for therapeutic applications74. In addition, Alkyl bonds suggest that Abiraterone acetate/Zytiga has the potential to be stable and long-lasting within the body, which is essential for consistent and effective treatment75. In a similar study Rajan et al. 2023 proved the inhibition of pantothenate synthetase with seaweed metabolites showing potential inhibitory properties, however the best binding energies obtained were − 5.547 kcal/mol and − 2.73 kcal/mol, which were not as strong as the interactions observed with Abiraterone acetate/Zytiga. This further highlights the potential of Abiraterone acetate/Zytiga as a promising candidate for therapeutic use in the treatment of certain conditions. The stability, multi-targeted approach, and strong interactions observed with this compound make it a stand-out option for further research and development in the medical field.
Abiraterone acetate, which has the highest binding affinity, is an FDA-approved drug used to treat advanced prostate cancer. Abiraterone acetate is a steroidal cytochrome P450 17α-hydroxylase-17,20-lyase (CYP17) inhibitor. It inhibits androgen biosynthesis, specifically targeting androgen synthesis within the testes, adrenal glands, and prostate tumor tissue. This inhibition ultimately suppresses androgen-sensitive prostate cancer growth. Abiraterone acetate is indicated in combination with prednisone for the treatment of metastatic castration-resistant prostate cancer (mCRPC) in patients who have received prior chemotherapy containing docetaxel and for metastatic castration-sensitive prostate cancer (mCSPC). It is available in oral tablet form. Common adverse effects include hypertension, hypokalemia, fluid retention, and liver function test abnormalities. Abiraterone acetate is metabolized primarily by hepatic CYP3A4 enzymes76,77,78.
MD simulations
Molecular dynamics simulations were performed on the top hits having high binding energies. Over the simulation period, the projected conformational changes from the initial structure were presented in terms of root mean square deviation (RMSD). Moreover, structural stability, atomic mobility, and residue flexibility at times of interaction of protein-hit were expressed with root mean square fluctuation (RMSF) values. We also analyzed the change in secondary structures of the proteins during MD simulations. Principal component analysis (PCA) and free energy landscape (FEL) are conducted for the investigation of conformational changes in the proteins. Dynamic cross-correlation matrix (DCCM) is calculated for identification of the patterns of correlation of motions of residues. Finally, molecular mechanics/generalized Born surface area (MM/GBSA) is utilized to study the binding energy of the protein–ligand complexes in the course of MD simulation.
Isocitrate lyase with dihydroergotamine
Molecular dynamics (MD) simulation is performed for isocitrate lyase for three systems, the apoenzyme, the control system and the isocitrate lyase-dihydroergotamine complex. The control system is the isocitrate lyase-ZINC2111081 complex, where ZINC2111081 was previously shown to be an effective drug that inhibits isocitrate lyase79. The root mean square deviation (RMSD) plot, in which only C-alpha atoms are considered, shows that all three systems attain equilibrium after 50 ns (Fig. 5). The apoenzyme system has the highest value, followed by the protein-dihydroergotamine complex system, and the control system has the lowest value (after 50 ns, apoenzyme: mean: 8.23 Å, SD: 0.275 Å; protein-dihydroergotamine: mean: 5.94 Å, SD: 0.358 Å; control: mean: 4.61 Å, SD: 0.461 Å). The difference in RMSD between the two ligand-bound protein systems is less significant than that between the apoenzyme and the ligand-bound protein systems. This indicates that the fluctuation of the protein system is the highest for the apoenzyme, and the binding of drugs lowers the degree of fluctuation of the enzyme probably due to the interactions between the protein and the drugs that hinder the motions of the residues, which in turn might affect the normal function of the enzyme.

Root mean square deviation (RMSD) of isocitrate lyase in three systems, namely Apoenzyme (apoenzyme system), Control (isocitrate lyase-ZINC2111081 complex) and Dihydroergotamine (isocitrate lyase-dihydroergotamine complex). Only the C-alpha atoms are considered.
The degree of fluctuation of various residues in the three systems is shown in the RMSF plot (Fig. 6). The regions near the C-terminus fluctuate the most for all systems. The overall RMSF is highest for the apoenzyme system and is lower for the two ligand-bound systems, the difference in RMSF between which is less significant, agreeing with the results of RMSD. For the control system, the ZINC2111081 ligand interacts with the isocitrate lyase via Pro21–Asp25, Trp57–Glu64, Ala276–Asp280, Lys302–Met309 and Lys334–Lys342. For the protein-dihydroergotamine complex system, the dihydroergotamine ligand interacts with the enzyme via Leu69–Ala83, Ala349–Ser357 and Glu380–His393. It should be noted that the decrease in the degree of fluctuation is not localized at the residues of interactions but is contributed by almost every part of the protein, which indicates that the interactions of the residues with the drugs have effects on other segments of the enzyme, probably by hindering the motions the residues of interactions, which in turn affects the movements of residues that are further away via the peptide chain or hydrogen bonds.

Root mean square fluctuation (RMSF) of isocitrate lyase in three systems, namely Apoenzyme (apoenzyme system), Control (isocitrate lyase-ZINC2111081 complex) and Dihydroergotamine (isocitrate lyase-dihydroergotamine complex). Only the C-alpha atoms are considered. The red and blue vertical lines represent the residues that interact with the ligand in the Control and Dihydroergotamine systems, respectively. Only the C-alpha atoms are considered.
We also analyzed the radius of gyration (Rg) of the simulation of the three systems, which shows the degree of compactness of the protein (Fig. 7). The apoenzyme has the lowest Rg among the three systems, which suggests that it folds the fastest during the simulation and is more compact. For the two ligand-bound protein systems, the control one has higher Rg then the dihydroergotamine one (after 50 ns, apoenzyme: mean: 22.5 Å, SD: 0.137 Å; protein-dihydroergotamine: mean: 23.0 Å, SD: 0.129 Å; control: mean: 24.5 Å, SD: 0.217 Å). This indicates that the binding of drug molecules lowers the compactness of isocitrate lyase, which could be due to the formation of hydrogen bonds between the drug and the residues which expand the protein. The lower compactness of the enzyme-drug complex might suggest a destabilization effect.

The radius of gyration (Rg) of isocitrate lyase in three systems, namely Apoenzyme (apoenzyme system), Control (isocitrate lyase-ZINC2111081 complex) and Dihydroergotamine (isocitrate lyase-dihydroergotamine complex). Only the protein backbone is considered.
The % secondary structure plot offers a comprehensive overview of the temporal evolution of secondary structure elements in isocitrate lyase in the three systems of the apoenzyme, the control system and the protein-dihydroergotamine complex, supported by aligned structural snapshots at 0, 50, and 100 ns (Fig. 8). All simulations showcase prevalent alpha helices, followed by beta strands, in line with typical protein structural patterns. Notably, isocitrate lyase with ligand demonstrates the highest total secondary structure elements at 50.47%, dominated by alpha helices at 40.53% and slightly lower beta strands at 9.94%, suggesting a potential stabilizing effect of the ligand on helical elements. In comparison, the apoenzyme displays slightly lower percentages of both alpha helices (39.67%) and beta strands (9.63%). This could be a reason for the higher level of fluctuation of the apoenzyme system according to the RMSD results. The control simulation exhibits a marginally lower percentage of alpha helices (39.45%) and slightly higher beta strands (10.26%). The timeline plot visually illustrates the stability of the isocitrate lyase with ligand complex, while apoenzyme and control simulations show comparable trends, emphasizing shared characteristics in the absence of the ligand.

%age secondary structure timeline of isocitrate lyase in three systems, namely Apoenzyme (apoenzyme system), Control (isocitrate lyase-ZINC2111081 complex) and Dihydroergotamine (isocitrate lyase-dihydroergotamine complex).
Principal component analysis (PCA) is conducted for analysis of the trajectories of the three systems. The C-alpha coordinates of all protein systems are projected to the principal components generated with the isocitrate lyase-dihydroergotamine system. From the PCA plots, there are substantial differences in their trajectories and conformational changes for the three systems during simulation (Fig. 9). Considering the apoenzyme system, it occupies two regions in the PCA plot. There is some overlapping between the apoenzyme and control systems and between the apoenzyme and protein-dihydroergotamine ones. However, there are regions that are reachable by the apoenzyme system but not by the control and protein-dihydroergotamine ones, which indicates that the drugs hinder the motion and conformational changes of the protein. The overlapping between the control and protein-dihydroergotamine systems is small, suggesting that the dihydroergotamine has a different inhibition mechanism when compared to ZINC2111081.

Principal component analysis (PCA) of isocitrate lyase in three systems, namely Apoenzyme (apoenzyme system, a), Control (isocitrate lyase-ZINC2111081 complex, b) and Dihydroergotamine (isocitrate lyase-dihydroergotamine complex, c), and (d) the overlaid PCA plot of three systems. Only the C-alpha atoms are considered.
Free energy landscape (FEL) is plotted using the results of PCA. It is for the analysis of the metastable conformations of the isocitrate lyase in the three systems, and it is verified by the kernel density estimation (KDE) plots (Fig. 10). In the apoenzyme system, two energy minima can be observed, indicating the existence of two metastable conformers (conformers A and B). The control system shows two energy minima which correspond to two conformers (conformers A1 and A2). The conformers A1 and A2 are closer to conformer A than B in the apoenzyme system in terms of position, which suggests that A1 and A2 should be more similar to A. The protein-dihydroergotamine system shows one energy minimum, and there is one metastable conformer (conformer B’). The position of B’ is closer to that of B than A in the apoprotein system, which suggests that B’ is more similar to B. From these results, the functional apoenzyme has two conformers, and it can be deduced that the binding of drugs inhibits the protein by locking it in one specific conformer. It can be also shown that the inhibition mechanisms of ZINC2111081 and dihydroergotamine are different because they lock the protein at two different conformers (conformer A for ZINC2111081; conformer B for dihydroergotamine).


Free energy landscape (FEL) of isocitrate lyase in three systems, namely Apoenzyme (apoenzyme system, a), Control (isocitrate lyase-ZINC2111081 complex, b) and Dihydroergotamine (isocitrate lyase-dihydroergotamine complex, c). PCA results colored by Gibbs free energy of Apoenzyme (d), Control (e) and Dihydroergotamine (f). Kernel density estimation (KDE) plots of PCA results of Apoenzyme (g), Control (h) and Dihydroergotamine (i).
To analyze the correlation of motions of residues in the three protein systems, dynamic cross-correlation matrix (DCCM) is calculated. The RMSD results show that the three systems attain equilibrium after 50 ns, so we extracted those frames for DCCM analysis. It can be observed that the drug-bound protein systems show very different patterns when compared with the apoenzyme (Fig. 11). The binding of ZINC2111081 to isocitrate lyase changes its correlation pattern, but the number of correlated motions is still substantial. In contrast, the binding of dihydroergotamine to the enzyme ruins most of the correlated motions, possibly indicating that dihydroergotamine is a more effective drug. This might also indirectly suggest that the mechanisms of inhibition are not the same for both drugs, which supports the deduction from the analysis of PCA and FEL.

Dynamic cross-correlation matrix (DCCM) of isocitrate lyase in three systems, namely (a) Apoenzyme (apoenzyme system), (b) Control (isocitrate lyase-ZINC2111081 complex), and (c) Dihydroergotamine (isocitrate lyase-dihydroergotamine complex).
Molecular mechanics/generalized Born surface area (MM/GBSA) is employed to evaluate the binding energy of isocitrate lyase and the two drugs (ZINC2111081 and dihydroergotamine). The MM/GBSA results (Fig. 12) show that the isocitrate lyase-dihydroergotamine complex is lower than the isocitrate lyase-ZINC2111081 complex in terms of binding energy during simulation (after 50 ns, isocitrate lyase-dihydroergotamine complex: mean: − 55.4 kcal/mol, SD: 3.59 kcal/mol; isocitrate lyase-ZINC2111081 complex: mean: − 39.8 kcal/mol, SD: 4.21 kcal/mol), suggesting that the protein-dihydroergotamine complex is more stable. This also suggests that dihydroergotamine might be a more effective drug when compared to ZINC2111081, which is supported by the results of the DCCM analysis.

MM/GBSA energy of isocitrate lyase in the two ligand-bound systems, namely Control (isocitrate lyase-ZINC2111081 complex) and Dihydroergotamine (isocitrate lyase-dihydroergotamine complex).
Pantothenate synthetase with abiraterone acetate
MD simulation is performed for pantothenate synthetase for three systems, the apoenzyme, the control system and the pantothenate synthetase-abiraterone acetate complex. The control system is the pantothenate synthetase-(2 s)-2-Amino-4-({4-[(5-methyl-1,2-oxazol-3-yl)sulfamoyl]phenyl}carbamoyl)butanoic acid (pantothenate synthetase-C15H18N4O6S) complex, where C15H18N4O6S is an effective drug targeting pantothenate synthetase according to previous research80. The RMSD plot, in which only C-alpha atoms are considered, shows that all three systems attain equilibrium after 80 ns (Fig. 13). The protein-abiraterone acetate system has the highest value, followed by the apoenzyme and control systems (after 80 ns, protein-abiraterone acetate: mean: 4.57 Å, SD: 0.495 Å; apoenzyme: mean: 3.32 Å, SD: 0.338 Å; control: mean: 2.91 Å, SD: 0.206 Å), which means that the protein-abiraterone acetate complex fluctuates the most, and the binding the abiraterone acetate might destabilize the enzyme. The difference in RMSD between the apoenzyme and the control systems is small and not very significant. This indicates that the binding of abiraterone acetate decreases the degree of fluctuation of the protein, while that of C15H18N4O6S does not affect the protein significantly in terms of RMSD, which might suggest that abiraterone acetate is a better drug when compared to C15H18N4O6S.

Root mean square deviation (RMSD) of pantothenate synthetase in three systems, namely Apoenzyme (apoenzyme system), Control (pantothenate synthetase-C15H18N4O6S complex) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex). Only the C-alpha atoms are considered.
The degree of fluctuation of various residues in the three systems is shown in the RMSF plot (Fig. 14). The overall RMSF is highest for the protein-abiraterone acetate system. The aforementioned higher fluctuation protein-abiraterone acetate system is mainly contributed by the residues near the C-terminal. For the control system, the C15H18N4O6S ligand interacts with pantothenate synthetase mainly via Pro38–Leu50, Ser65–Tyr82, Arg132–Val142, Phe156–Gln164, Thr186–Val187 and Met195–Arg198. For the protein-abiraterone acetate complex system, the abiraterone acetate ligand interacts with the enzyme mainly via Met40–Leu50, Gln72–Ala81, Glu128–Val139, Phe157–Gln164, Val184–Val187 and Met195–Arg198. It should be noted that the regions near the residues that have interactions with the drugs do not have a huge change in fluctuation.

Root mean square fluctuation (RMSF) of pantothenate synthetase in three systems, namely Apoenzyme (apoenzyme system), Control (pantothenate synthetase-C15H18N4O6S complex) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex). Only the C-alpha atoms are considered. The red and blue vertical lines represent the residues that interact with the ligand in the Control and Abiraterone Acetate systems, respectively. Only the C-alpha atoms are considered.
The Rg, which is an indicator of compactness, of the three systems are also analyzed. Figure 15 shows that the apoenzyme has the lowest Rg among the three systems, which suggests that it is the most compact system. The control system has similar Rg values when compared to the apoenzyme one, while the protein-abiraterone acetone system has overall higher Rg values (after 80 ns, apoenzyme: mean: 20.4 Å, SD: 0.0821 Å; control: mean: 20.2 Å, SD: 0.128 Å; protein-abiraterone acetone: mean: 20.8 Å, SD: 0.148 Å). This indicates that the binding of abiraterone acetone makes the pantothenate synthetase more expanded, which could be due to the hydrogen bonds formed between the protein and the ligand. The lower compactness might destabilize the enzyme. It is noteworthy that the binding of C15H18N4O6S does not lower the compactness much because of the small difference in Rg values between the protein-C15H18N4O6S complex and the apoenzyme. This might again suggest that abiraterone acetone is a more effective drug, which is supported by the RMSD results.

The radius of gyration (Rg) of pantothenate synthetase in three systems, namely Apoenzyme (apoenzyme system), Control (pantothenate synthetase-C15H18N4O6S complex) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex). Only the protein backbone is considered.
The % secondary structure plot shows the distribution of secondary structures inside the pantothenate synthetase simulations in the apoenzyme, control and protein-abiraterone acetate systems (Fig. 16). In the pantothenate synthetase-abiraterone acetate simulation, the protein has 31.82% alpha-helix, 19.12% beta-strand, and a total secondary shape element (SSE) content of 50.94%. Similarly, the apoenzyme simulation demonstrates 31.08% helix, 18.38% strand, and a total SSE of 49.46%, whilst the control simulation indicates 31.88% helix, 18.70% strand, and a complete SSE of 50.57%. The aligned timeline plot depicts the development of these simulations through the simulation of 100 ns. Additionally, the frames of the three systems were extracted at 1, 50, and 100 ns and then superimposed on each other. There was no major difference observed which reinforces the stability of protein structures.

%age secondary structure timeline of of pantothenate synthetase in three systems, namely Apoenzyme (apoenzyme system), Control (pantothenate synthetase-C15H18N4O6S complex) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex).
PCA is conducted for analysis of the trajectories. The C-alpha coordinates of all protein systems are projected to the principal components generated with the pantothenate synthetase-abiraterone acetate system. Substantial differences in their trajectories and conformational changes for the three systems are observed in the PCA plots (Fig. 17), and there are some degrees of overlapping among them. The control system overlaps with the apoenzyme one considerably although there are some points of the control system that do not lie in the apoenzyme regions. In contrast, the overlapping between the pantothenate synthetase-abiraterone acetate system, which occupies roughly two regions, and the apoenzyme one is extremely small, or they almost do not overlap. This might indicate that the binding of abiraterone acetate can better distort the structure of the protein so that it is less similar to its native structure when compared to C15H18N4O6S, which in turn suggests that abiraterone acetate is a better inhibitor of pantothenate synthetase. This agrees with the results of the RMSD and Rg analyses. It should be noted that some degree of overlap between the control and protein-abiraterone acetate regions exists, indicating that both drugs inhibit the protein by guiding it to a similar distorted conformation.

Principal component analysis (PCA) of pantothenate synthetase in three systems, namely Apoenzyme (apoenzyme system, a), Control (pantothenate synthetase-C15H18N4O6S complex, b) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex, c), and (d) the overlaid PCA plot of three systems. Only the C-alpha atoms are considered.
Using the result of PCA, FEL is plotted for the analysis of the metastable conformations, and it is verified by the KDE plots (Fig. 18). In the apoenzyme system, only one energy minimum or metastable conformer (conformer P) is observed. In the control system, two metastable conformers, one with lower energy (conformer P’) and another with higher energy (conformer Q’), exist. The protein-abiraterone acetate system has two metastable conformers (conformers Q and R). In terms of the positions of the conformers, P is similar to P’ and Q is similar to Q’. Due to the similarity between P’ and P, the conformation adopted by the control system is close to that adopted by the apoenzyme because P’ has lower energy than Q’, which means that the control system stays at P’ for a longer period. The conformations adopted by the protein-abiraterone acetate system are very different when compared to the apoenzyme because Q and R do not lie close to P. These results indicate that the binding to ligands can change the conformation of the protein. When C15H18N4O6S is bound, the major conformation changes from P to P’, while it changes from P to Q and R when abiraterone acetate is bound, from which we can also deduce that abiraterone acetate can make a more significant modification to the protein conformations. We should also note the similarity between Q in the protein-abiraterone acetate system and Q’ in the control system, which indicates that both drugs change the conformation towards the Q or Q’ direction, but C15H18N4O6S does so less effectively because the energy of Q’ is high when compared to the dominant conformer P’ of the control system.


Free energy landscape (FEL) of pantothenate synthetase in three systems, namely Apoenzyme (apoenzyme system, a), Control (pantothenate synthetase-C15H18N4O6S complex, b) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex, c). PCA results colored by Gibbs free energy of Apoenzyme (d), Control (e) and Abiraterone Acetate (f). Kernel density estimation (KDE) plots of PCA results of Apoenzyme (g), Control (h) and Abiraterone Acetate (i).
DCCM is used to analyze the correlation of motions of residues in the three protein systems. The RMSD results show that the three systems attain equilibrium after 80 ns, so we extracted those frames for DCCM analysis. The three systems show very different patterns of DCCM (Fig. 19). For the apoenzyme, the major correlation comes from the residues 1–200. The binding of C15H18N4O6S changes the correlation patterns such that the contribution is mainly from residues 200–300. The protein-abiraterone acetate system exhibits correlations in motions between residues 295–309 and all other parts of the protein. Also, the reduction of correlations caused by binding of drugs is higher for abiraterone acetate than C15H18N4O6S, this might further support the previous deduction that abiraterone acetate is a more effective drug.

Dynamic cross-correlation matrix (DCCM) of pantothenate synthetase in three systems, namely Apoenzyme (apoenzyme system, a), Control (pantothenate synthetase-C15H18N4O6S complex, b) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex, c).
MM/GBSA is employed to evaluate the binding energy of pantothenate synthetase and the two drugs, namely C15H18N4O6S and abiraterone acetate. The MM/GBSA results show overall higher energy of the binding of abiraterone acetate (Fig. 20). Despite this, after the system has equilibrated after 80 ns according to the RMSD results, there is, in fact, no significant difference in binding energy between the two systems (after 80 ns, pantothenate synthetase-C15H18N4O6S complex: mean: − 47.2 kcal/mol, SD: 3.32 kcal/mol; pantothenate synthetase-abiraterone acetate complex: mean: − 44.0 kcal/mol, SD: 2.78 kcal/mol). While the difference in binding energy is small, the binding of abiraterone acetate can induce a higher degree of conformational change of the protein according to the results of PCA and FEL. This shows that the binding energy is not disadvantageous for the drug effectiveness of abiraterone acetate.

MM/GBSA energy of pantothenate synthetase in the two ligand-bound systems, namely Control (pantothenate synthetase-C15H18N4O6S complex, b) and Abiraterone Acetate (pantothenate synthetase-abiraterone acetate complex, c).
Conclusion
Exploring new ways to fight TB is essential to reducing the need for extensive treatment and drug resistance. The study identifies the enzymes isocitrate lyase and pantothenate synthetase as promising targets for treating M. tuberculosis strains, whether drug-sensitive or drug-resistant. In addition, this research has uncovered the best ligands (dihydroergotamine, abiraterone acetate) with high binding affinity for these drug targets, providing crucial direction for upcoming M. tuberculosis drug development efforts. These findings suggest efficient TB therapeutics that may even replace currently available medications. However, it is necessary to emphasize that experimental validation is essential to confirm these results’ authenticity.
Gaps and limitations
While this study identified promising drug candidates, further testing is necessary. In vitro and in vivo experiments will be crucial to confirm the efficacy and safety of these drugs for treating Mycobacterium tuberculosis infection.
Data availability
The datasets used and analyzed during the current study have already been reported here. Any further information needed will be made available by the correspondence author.
References
Bagcchi, S. WHO’s global tuberculosis report 2022. Lancet Microbe 4, e20 (2023).
Dar, H. A. et al. Pangenome analysis of Mycobacterium tuberculosis reveals core-drug targets and screening of promising lead compounds for drug discovery. Antibiotics 9, 819 (2020).
Bainomugisa, A. et al. Evolution and spread of a highly drug resistant strain of Mycobacterium tuberculosis in Papua New Guinea. BMC Infect. Dis. 22, 437 (2022).
Banerjee, U., Sankar, S., Singh, A. & Chandra, N. A multi-pronged computational pipeline for prioritizing drug target strategies for latent tuberculosis. Front. Chem. 8, 593497 (2020).
Cole, S. et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 396, 190–190 (1998).
Vallenet, D. et al. MicroScope: An integrated platform for the annotation and exploration of microbial gene functions through genomic, pangenomic and metabolic comparative analysis. Nucleic Acids Res. 48, D579–D589 (2020).
Pedelacq, J. D., Nguyen, M. C., Terwilliger, T. C. & Mourey, L. A comprehensive review on mycobacterium tuberculosis targets and drug development from a structural perspective. In Structural Biology in Drug Discovery: Methods, Techniques, and Practices 545–566. https://doi.org/10.1002/9781118681121.ch23 (2020).
Ejalonibu, M. A. et al. Drug discovery for Mycobacterium tuberculosis using structure-based computer-aided drug design approach. Int. J. Mol. Sci. 22, 13259 (2021).
Ayu Eka Pitaloka, D. et al. Bioinformatics analysis to uncover the potential drug targets responsible for Mycobacterium tuberculosis peptidoglycan and lysine biosynthesis. Bioinform. Biol. Insights 17, 11779322231171774 (2023).
Timo, G. O. et al. Predictive power of in silico approach to evaluate chemicals against M. tuberculosis: A systematic review. Pharmaceuticals 12, 135 (2019).
Schaller, D. et al. Next generation 3D pharmacophore modeling. Wiley Interdiscip. Rev.: Comput. Mol. Sci. 10, e1468 (2020).
Al-Sanea, M. M. et al. Identification of novel potential VEGFR-2 inhibitors using a combination of computational methods for drug discovery. Life 11, 1070 (2021).
Wattam, A. R. et al. PATRIC, the bacterial bioinformatics database and analysis resource. Nucleic Acids Res. 42, D581–D591 (2014).
Chaudhari, N. M., Gupta, V. K. & Dutta, C. BPGA-an ultra-fast pan-genome analysis pipeline. Sci. Rep. 6, 24373 (2016).
Naz, K. et al. PanRV: Pangenome-reverse vaccinology approach for identifications of potential vaccine candidates in microbial pangenome. BMC Bioinform. 20, 1–10 (2019).
Nazir, Z., Afridi, S. G., Shah, M., Shams, S. & Khan, A. Reverse vaccinology and subtractive genomics-based putative vaccine targets identification for Burkholderia pseudomallei Bp1651. Microb. Pathog. 125, 219–229 (2018).
Luo, H. et al. DEG 15, an update of the database of essential genes that includes built-in analysis tools. Nucleic Acids Res. 49, D677–D686 (2021).
Liu, B., Zheng, D., Zhou, S., Chen, L. & Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 50, D912–D917 (2022).
Peterson, J. W. Bacterial pathogenesis. In Medical Microbiology 4th edn (1996).
Gasteiger, E. et al. Protein Identification and Analysis Tools on the ExPASy Server (Springer, 2005).
Naorem, R. S. et al. Identification of putative vaccine and drug targets against the methicillin-resistant Staphylococcus aureus by reverse vaccinology and subtractive genomics approaches. Molecules 27, 2083 (2022).
Aguttu, C., Okech, B. A., Mukisa, A. & Lubega, G. W. Screening and characterization of hypothetical proteins of Plasmodium falciparum as novel vaccine candidates in the fight against malaria using reverse vaccinology. J. Genet. Eng. Biotechnol. 19, 1–13 (2021).
Moriya, Y., Itoh, M., Okuda, S., Yoshizawa, A. C. & Kanehisa, M. KAAS: an automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 35, W182–W185 (2007).
Wishart, D. S. et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 46, D1074–D1082 (2018).
Mondal, S. I., Ferdous, S., Jewel, N. A., Akter, A., Mahmud, Z., Islam, M. M., Afrin, T. & Karim, N. Identification of potential drug targets by subtractive genome analysis of Escherichia coli O157: H7: An in silico approach. In Advances and Applications in Bioinformatics and Chemistry 49–63. https://doi.org/10.2147/AABC.S88522 (2015).
Irwin, J. J. & Shoichet, B. K. ZINC: A free database of commercially available compounds for virtual screening. J. Chem. Inf. Model. 45, 177–182 (2005).
Burley, S. K. et al. RCSB protein data bank (RCSB. org): Delivery of experimentally-determined PDB structures alongside one million computed structure models of proteins from artificial intelligence/machine learning. Nucleic Acids Res. 51, D488–D508 (2023).
Dallakyan, S. & Olson, A. J. Small-molecule library screening by docking with PyRx. In Chemical Biology: Methods and Protocols 243–250. https://doi.org/10.1007/978-1-4939-2269-7_19 (2015).
Trott, O. & Olson, A. J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 31, 455–461 (2010).
Reynolds, C. H., Bembenek, S. D. & Tounge, B. A. The role of molecular size in ligand efficiency. Bioorg. Med. Chem. Lett. 17, 4258–4261 (2007).
Toukmaji, A. Y. & Board, J. A. Jr. Ewald summation techniques in perspective: A survey. Comput. Phys. Commun. 95, 73–92 (1996).
Zielkiewicz, J. Structural properties of water: Comparison of the SPC, SPCE, TIP4P, and TIP5P models of water. J. Chem. Phys. 123, 104501 (2005).
Martyna, G. J., Klein, M. L. & Tuckerman, M. Nosé-Hoover chains: The canonical ensemble via continuous dynamics. J. Chem. Phys. 97, 2635–2643 (1992).
Rehman, H. M. et al. Identification of RdRp inhibitors against SARS-CoV-2 through E-pharmacophore-based virtual screening, molecular docking and MD simulations approaches. Int. J. Biol. Macromol. 237, 124169 (2023).
Gul, R. et al. Molecular cloning, expression, sequence characterization and structural insight of Bubalus bubalis growth hormone-receptor. Mol. Biotechnol. 65, 1062–1075 (2023).
Rehman, H. M. et al. Identification of NS2B-NS3 protease inhibitors for therapeutic application in ZIKV infection: A pharmacophore-based high-throughput virtual screening and MD simulations approaches. Vaccines 11, 131 (2023).
Shinwari, K. et al. Novel disease-associated missense single-nucleotide polymorphisms variants predication by algorithms tools and molecular dynamics simulation of human TCIRG1 gene causing congenital neutropenia and osteopetrosis. Front. Mol. Biosci. 9, 879875 (2022).
Pronk, S. et al. GROMACS 4.5: A high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 29, 845–854 (2013).
McGibbon, R. T. et al. MDTraj: A modern open library for the analysis of molecular dynamics trajectories. Biophys. J. 109, 1528–1532 (2015).
Fenwick, R. B., Orellana, L., Esteban-Martín, S., Orozco, M. & Salvatella, X. Correlated motions are a fundamental property of β-sheets. Nat. Commun. 5, 4070 (2014).
Grant, B. J., Rodrigues, A. P., ElSawy, K. M., McCammon, J. A. & Caves, L. S. Bio3d: An R package for the comparative analysis of protein structures. Bioinformatics 22, 2695–2696 (2006).
Skjærven, L., Yao, X.-Q., Scarabelli, G. & Grant, B. J. Integrating protein structural dynamics and evolutionary analysis with Bio3D. BMC Bioinform. 15, 1–11 (2014).
Yang, M. et al. Uni-GBSA: An open-source and web-based automatic workflow to perform MM/GB (PB) SA calculations for virtual screening. Brief. Bioinform. 24, bbad218 (2023).
Valdés-Tresanco, M. S., Valdés-Tresanco, M. E., Valiente, P. A. & Moreno, E. gmx_MMPBSA: A new tool to perform end-state free energy calculations with GROMACS. J. Chem. Theory Comput. 17, 6281–6291 (2021).
Miller, B. R. III. et al. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 8, 3314–3321 (2012).
O’Boyle, N. M. et al. Open Babel: An open chemical toolbox. J. Cheminform. 3, 1–14 (2011).
Qin, S. et al. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Therapy 7, 199 (2022).
Pham, T. V. et al. Mechanism-based inactivator of isocitrate lyases 1 and 2 from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. 114, 7617–7622 (2017).
Vanni, P., Giachetti, E., Pinzauti, G. & McFadden, B. A. Comparative structure, function and regulation of isocitrate lyase, an important assimilatory enzyme. Comp. Biochem. Physiol. Part B Comp. Biochem. 95, 431–458 (1990).
Giffin, M. M., Modesti, L., Raab, R. W., Wayne, L. G. & Sohaskey, C. D. ald of Mycobacterium tuberculosis encodes both the alanine dehydrogenase and the putative glycine dehydrogenase. J. Bacteriol. 194, 1045–1054 (2012).
Höner Zu Bentrup, K., Miczak, A., Swenson, D. L. & Russell, D. G. Characterization of activity and expression of isocitrate lyase in Mycobacterium avium and Mycobacterium tuberculosis. J. Bacteriol. 181, 7161–7167 (1999).
Duan, C. et al. Discovery of a novel inhibitor structure of Mycobacterium tuberculosis isocitrate lyase. Molecules 27, 2447 (2022).
Bhusal, R. P., Bashiri, G., Kwai, B. X., Sperry, J. & Leung, I. K. Targeting isocitrate lyase for the treatment of latent tuberculosis. Drug Discov. Today 22, 1008–1016 (2017).
Campaniço, A., Harjivan, S. G., Warner, D. F., Moreira, R. & Lopes, F. Addressing latent tuberculosis: New advances in mimicking the disease, discovering key targets, and designing hit compounds. Int. J. Mol. Sci. 21, 8854 (2020).
Nandakumar, M., Nathan, C. & Rhee, K. Y. Isocitrate lyase mediates broad antibiotic tolerance in Mycobacterium tuberculosis. Nat. Commun. 5, 4306 (2014).
Sharma, R., Das, O., Damle, S. G. & Sharma, A. K. Isocitrate lyase: A potential target for anti-tubercular drugs. Recent Patents Inflamm. Allergy Drug Discov. 7, 114–123 (2013).
Suresh, A. et al. Inhibitors of pantothenate synthetase of Mycobacterium tuberculosis: A medicinal chemist perspective. RSC Adv. 10, 37098–37115 (2020).
Wang, S. & Eisenberg, D. Crystal structure of the pantothenate synthetase from Mycobacterium tuberculosis, snapshots of the enzyme in action. Biochemistry 45, 1554–1561 (2006).
Pandey, B. et al. Alanine mutation of the catalytic sites of pantothenate synthetase causes distinct conformational changes in the ATP binding region. Sci. Rep. 8, 903 (2018).
Abrahams, G. L. et al. Pathway-selective sensitization of Mycobacterium tuberculosis for target-based whole-cell screening. Chem. Biol. 19, 844–854 (2012).
Kumar, A. et al. A high-throughput screen against pantothenate synthetase (PanC) identifies 3-biphenyl-4-cyanopyrrole-2-carboxylic acids as a new class of inhibitor with activity against Mycobacterium tuberculosis. PloS One 8, e72786 (2013).
Patil, K., Bagade, S., Bonde, S., Sharma, S. & Saraogi, G. Recent therapeutic approaches for the management of tuberculosis: Challenges and opportunities. Biomed. Pharmacother. 99, 735–745 (2018).
Wellington, S. & Hung, D. T. The expanding diversity of Mycobacterium tuberculosis drug targets. ACS Infect. Dis. 4, 696–714 (2018).
Rakesh, K., Shantharam, C., Sridhara, M., Manukumar, H. & Qin, H.-L. Benzisoxazole: A privileged scaffold for medicinal chemistry. MedChemComm 8, 2023–2039 (2017).
Abu-Dief, A. M. et al. Synthesis and intensive characterization for novel Zn (II), Pd (II), Cr (III) and VO (II)-Schiff base complexes; DNA-interaction, DFT, drug-likeness and molecular docking studies. J. Mol. Struct. 1242, 130693 (2021).
Alterio, V., Di Fiore, A., D’Ambrosio, K., Supuran, C. T. & De Simone, G. Multiple binding modes of inhibitors to carbonic anhydrases: How to design specific drugs targeting 15 different isoforms?. Chem. Rev. 112, 4421–4468 (2012).
Dunn, M., Ramirez-Trujillo, J. & Hernández-Lucas, I. Major roles of isocitrate lyase and malate synthase in bacterial and fungal pathogenesis. Microbiology 155, 3166–3175 (2009).
Saper, J. R. & Silberstein, S. Pharmacology of dihydroergotamine and evidence for efficacy and safety in migraine. Headache J. Head Face Pain 46, S171–S181 (2006).
Reddy, D. S. The pathophysiological and pharmacological basis of current drug treatment of migraine headache. Expert Rev. Clin. Pharmacol. 6, 271–288 (2013).
Ong, J. J. Y. & De Felice, M. Migraine treatment: Current acute medications and their potential mechanisms of action. Neurotherapeutics 15, 274–290 (2018).
Srisuma, S., Lavonas, E. & Wananukul, W. Ergotism and factitious hypotension associated with interaction of ergotamine with CYP3A4 inhibitors. Clin. Toxicol. 52, 674–677 (2014).
Thafar, M., Raies, A. B., Albaradei, S., Essack, M. & Bajic, V. B. Comparison study of computational prediction tools for drug-target binding affinities. Front. Chem. 7, 782 (2019).
Narkhede, R. R., Cheke, R. S., Ambhore, J. P. & Shinde, S. D. The molecular docking study of potential drug candidates showing anti-COVID-19 activity by exploring of therapeutic targets of SARS-CoV-2. Eurasian J. Med. Oncol. 4, 185–195 (2020).
Ramsay, R. R., Popovic-Nikolic, M. R., Nikolic, K., Uliassi, E. & Bolognesi, M. L. A perspective on multi-target drug discovery and design for complex diseases. Clin. Transl. Med. 7, 1–14 (2018).
Irfandi, R. et al. Design anticancer potential of Zn (II) isoleucinedithiocarbamate complex on MCF-7 cell lines: Synthesis, characterization, molecular docking, molecular dynamic, ADMET, and in-vitro studies. Mol. Divers. https://doi.org/10.1007/s11030-023-10747-y (2023).
Logothetis, C. J., Efstathiou, E., Manuguid, F. & Kirkpatrick, P. Abiraterone acetate. Nat. Rev. Drug Discov. 10, 573 (2011).
Fizazi, K. et al. Abiraterone acetate for treatment of metastatic castration-resistant prostate cancer: final overall survival analysis of the COU-AA-301 randomised, double-blind, placebo-controlled phase 3 study. Lancet Oncol. 13, 983–992 (2012).
Ryan, C. J. et al. Abiraterone acetate plus prednisone versus placebo plus prednisone in chemotherapy-naive men with metastatic castration-resistant prostate cancer (COU-AA-302): Final overall survival analysis of a randomised, double-blind, placebo-controlled phase 3 study. Lancet Oncol. 16, 152–160 (2015).
Shukla, R., Shukla, H., Sonkar, A., Pandey, T. & Tripathi, T. Structure-based screening and molecular dynamics simulations offer novel natural compounds as potential inhibitors of Mycobacterium tuberculosis isocitrate lyase. J. Biomol. Struct. Dyn. 36, 2045–2057 (2018).
Pradhan, S. & Sinha, C. High throughput screening against pantothenate synthetase identifies amide inhibitors against Mycobacterium tuberculosis and Staphylococcus aureus. In Silico Pharmacol. 6, 1–24 (2018).
Funding
This study is funded by UKRI MRC (MRC IAA2129, MR/R026297/1, and MR/X005895/1) and EPSRC (EP/Y018842/1) grants.
Author information
Authors and Affiliations
Contributions
M.F.K., A.J. and A.A. conceived and directed the project. M.F.K. and M.W. performed bioinformatic and statistical analyses under the supervision of A.J. and A.A. M.F.K. and A.J. and A.A. interpreted the results. M.F.K., H.M.R., and Y.W. performed MD simulation. M.F.K. wrote the first draft of the manuscript. T.G.C. reviewed the manuscript. All authors commented and edited on various versions of the draft manuscript and approved the final manuscript. M.F.K., A.J. and T.G.C. compiled the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Khan, M.F., Ali, A., Rehman, H.M. et al. Exploring optimal drug targets through subtractive proteomics analysis and pangenomic insights for tailored drug design in tuberculosis. Sci Rep 14, 10904 (2024). https://doi.org/10.1038/s41598-024-61752-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-61752-6
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
