
Abstract
Stress-related overeating can lead to excessive weight gain, increasing the risk of metabolic and cardiovascular disease. Mindfulness meditation has been demonstrated to reduce stress and increase interoceptive awareness and could, therefore, be an effective intervention for stress-related overeating behavior. To investigate the effects of mindfulness meditation on stress-eating behavior, meditation-naïve individuals with a tendency to stress-eat (N = 66) participated in either a 31-day, web-based mindfulness meditation training or a health training condition. Behavioral and resting-state fMRI data were acquired before and after the intervention. Mindfulness meditation training, in comparison to health training, was found to significantly increase mindfulness while simultaneously reducing stress- and emotional-eating tendencies as well as food cravings. These behavioral results were accompanied by functional connectivity changes between the hypothalamus, reward regions, and several areas of the default mode network in addition to changes observed between the insula and somatosensory areas. Additional changes between seed regions (i.e., hypothalamus and insula) and brain areas attributed to emotion regulation, awareness, attention, and sensory integration were observed. Notably, these changes in functional connectivity correlated with behavioral changes, thereby providing insight into the underlying neural mechanisms of the effects of mindfulness on stress-eating.
Clinical trial on the ISRCTN registry: trial ID ISRCTN12901054
Similar content being viewed by others

Introduction
Chronic stress has been demonstrated to have negative implications on many health-related domains, including eating behavior. This is due, in part, to the physiological stress response, which includes the sympathetic and parasympathetic nervous systems in addition to the hypothalamic-pituitary-adrenal (HPA) axis1,2. While these systems work together to allow the body to effectively respond to acute stress, chronic stress can lead to the dysregulation of the HPA axis, which can result in the aberrant production of stress hormones1,3. This can, in turn, affect the production of leptin, ghrelin, and neuropeptide Y, which play an integral role in regulating perceived hunger and satiety1,2,4,5,6,7. Chronic stress can, therefore, increase feelings of hunger and increase caloric intake8,9,10,11,12.
Furthermore, chronic stress has been shown to implicate executive functioning including emotion regulation and cognitive control13. This can have a significant effect on decision-making abilities as well as influence impulsive snacking and emotional eating behaviors13,14,15. This is particularly noteworthy as a large portion of the global population lives in an obesogenic environment, i.e., individuals are persistently being exposed to high-calorie food items16,17,18. Given that the mere presence of high-calorie food and its odors can elicit appetite-enhancing effects 19, stressed individuals may not only be more prone to experiencing an increase in appetite through the dysregulation of their HPA axis but may also be less able to utilize cognitive control (i.e., restraint) when encountering high-calorie foods.
Ultimately, the long-term effects of chronic stress and its influence on eating behavior can have negative implications on metabolic and cardiovascular health. For example, frequently engaging in stress-related overeating behavior (and the subsequent weight gain) can lead to type 2 diabetes and hypertensionor other cardiovascular diseases12,20,21,22. Considering the profound effect stress has on eating behavior, its long-term health consequences, in addition to the high global prevalence of cardiovascular and metabolic disease23,24,25, it is crucial for the scientific community to establish and investigate stress-related overeating behavior interventions.
To this end, mindfulness meditation training (MMT) has been demonstrated to be a reliable stress-reduction mechanism that can also improve overall well-being26,27,28,29,30,31,32. MMT involves the intentional focus of attention as well as open awareness of the present moment without judgment or distraction33. Through its ability to bring attention to thoughts, feelings, and bodily sensations, MMT can support individuals in developing greater interoceptive awareness, emotion regulation, and reduce stress sensitivity27,34,35,36,37,38,39. By becoming more aware of thoughts and emotions, individuals can observe stressors with greater objectivity and compassion, thereby reducing the physiological stress response in addition to the susceptibility to chronic stress40,41,42. Given the role MMT plays in reducing perceived stress as well as the physiological stress response, it is hypothesized that individuals suffering from stress-related overeating behavior would benefit from a food-related MMT.
As both stress and mindfulness have an effect on the neural level29,43,44,45,46,47, this study seeks to investigate the neural mechanisms through which MMT influences stress-eating behavior via seed-based resting-state functional connectivity (FC) changes. As the hypothalamus and insula are the fundamental regions in the mediation of the neural processes of hunger and satiety cues, these brain areas were selected as a-priori seed regions of interest.
The hypothalamus and its individual nuclei, through its involvement in the HPA axis, initiates change in appetite and food intake2,4,5,6,7. While the lateral hypothalamus, through the release of hormones such as neuropeptide Y and agouti-related peptides, regulates perceived hunger46,48,49,50, the medial hypothalamus regulates perceived satiety through the release of hormones such as melanin-concentrating-hormone (MCH)50.
Furthermore, a review published by Syan, McIntyre-Wood46 could demonstrate an increase in FC between the medial hypothalamus and areas of the reward system, suggesting an increased interdependence between perceived reward and satiety in individuals with obesity while additionally demonstrating that, in comparison to controls, individuals with obesity exhibited an increase in FC between the lateral hypothalamus and somatosensory areas. These findings suggest that differences in eating behavior are accompanied by specific FC changes in the brain. Therefore, given its central role in the complex interplay between stress, appetite, and eating behavior, the hypothalamus was selected as a region to observe possible neuronal changes elicited by food-related MMT.
The insula is a brain area that contributes to cognitive processes underlying both MMT and eating behavior. For example, the insula has been demonstrated to be involved in interoception and attentional control, which are both fundamental aspects of MMT as well as eating behavior51. Moreover, the insula and its nuclei play a role in the subjective experience of flavor, texture, and smell52,53,54, which contribute to the hedonic evaluation of food. For example, MMT and its involvement in fostering awareness has been associated with the anterior insula, an area involved in emotion regulation and interoceptive awareness55,56,57, which are two factors relevant to eating behavior, hunger, and satiety cues, thereby allowing for or less emotion-dependent food selections51,58. The posterior insula, on the other hand, is associated with sensory processing and the integration of taste-related stimuli, which ultimately influence food choice59. The insula is, therefore, not only associated with regulating perceived hunger and satiety cues but also plays a fundamental role in mediating cognitive, emotional, and sensory processes related to both mindfulness and stress-related eating behavior.
This research investigating the FC changes between the hypothalamus, insula, and the whole brain will not only provide insight into the effects of MMT on stress-related eating behavior but also into the neural mechanisms underlying eating behavior and stress reduction. These results could, therefore, provide critical evidence of an intervention for stress-eaters to cultivate a more mindful relationship with food.
Methods
Participants
Addressing individuals with a tendency to overeat when stressed, participants were recruited via the university hospital’s mailing list and online advertisements. Participants were required to report moderate to high levels of stress, as assessed by the Perceived Stress Scale (PSS)60 while expressing a tendency to engage in stress-eating and needed to fulfill the following criteria: (1) between the ages of 18 and 45 (2) general MRI suitability (i.e., no metal implants and not prone to claustrophobia), (3) body-mass-index (BMI) between 18 and 30, (4) no dietary restrictions (including vegetarianism or veganism)61, (5) no use of oral contraceptives or intrauterine devices, (6) no known, untreated, thyroid dysfunction, (7) no chronic respiratory diseases. All participants provided written, informed, consent and were given monetary compensation for their participation. This study is listed as a clinical trial on the ISRCTN registry with trial ID ISRCTN12901054 registered on 19/05/2023, was performed in accordance with the Declaration of Helsinki, and was approved by the Ethics Committee of Klinikum Rechts der Isar, Technical University Munich.
Procedure
This study was designed as a pseudo-randomized, actively controled trial to investigate the effects of MMT on stress-eating behavior and its neural correlates. Participants were single-blindedly (subject only) allocated to either the MMT condition or the active control, health training (HT) condition. All participants underwent magnetic resonance imaging (MRI), and psychometric testing evaluating perceived mindfulness, stress, emotional eating, food cravings, dietary restraint, and the assessment of body weight prior to and after completing the intervention. All measures were acquired after a subjectively stressful day and participants were instructed to abstain from eating five hours prior to their scheduled measure. The training programs for both conditions were accessible via an online platform and consisted of 31 daily 15 min sessions. To promote training adherence, daily reminders were sent to participants by email.
The MMT was developed in close cooperation with author BKH who is a certified MBSR instructor. In the MMT, participants were provided with a detailed introduction to the theoretical framework of MMT while additionally guiding participants through the daily meditation exercises via video or audio clips. Written instructions emphasized the relationship between MMT and eating behavior, encouraging participants to engage more mindfully with food, for example, by consciously attending to the flavor and texture of the food they consume or to any emotions that arise in the context of eating. The HT condition provided participants with informative, health-related video and audio clip excerpts from popular science broadcasting networks in Germany. Designed to imitate the MMT, the HT was delivered via the same online platform and followed the same order of video and audio material. However, topics in the HT did not include any information pertaining to mindfulness, eating behavior, or nutrition. For a detailed description of the training content, please see Table S1. Participants were required to complete at least 27 training sessions to be included in the final analysis.
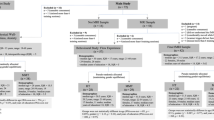
295 participants were assessed for eligibility, and 112 participants fulfilled the inclusion criteria. 87 participants completed the first MRI measure, and 74 participants were also available for the second MRI measure. After preprocessing, data from 66 participants (33 female) were included in the final analysis. Data acquisition took place between June 2019 and June 2021 and is depicted in Fig. 1.

Data acquisition process (June 2019—June 2021). MMT Mindfulness Meditation Training, HT Health Training.
To verify the pseudo-randomization process, final samples were compared for demographic characteristics, measuring intervals, and an average number of sessions completed using t-tests for independent samples or chi-square tests, respectively.
Behavioral data
To investigate various behavioral measures pertaining to the effects of MMT on eating behavior, participants completed the German version of the Mindful Attention and Awareness Scale (MAAS)62, Food Cravings Questionnaire-Trait (FCQ-T), Salzburg Stress Eating Scale (SSES), Salzburg Emotional Eating Scale (SEES), as well the Restraint scale.
The statistical analyses of behavioral data were performed using SPSS v29.0. Independent t-tests were conducted on data obtained during the first measure to identify and exclude potential baseline differences. To assess training effects, psychometric data was entered into a 2 × 2 mixed effects analysis of variance (ANOVA), where group was the between-subject factor and time was the within-subject factor. Results were thresholded at p < 0.05.
MRI data acquisition
MRI data were acquired on a 3 T Philips MRI scanner with a 32-channel head coil at Klinikum Rechts der Isar’s department of neuroradiology in Munich, Germany.
T2*-weighted resting-state functional images were acquired using echo planar imaging (EPI) with the following scanning parameters: Multiband (MB) factor 2, repetition time (TR) 2.3 s, echo time (TE) 30 ms and flip angle 75°. The field of view (FOV) was set to (192 × 192 × 136) mm3, corresponding to a matrix size of 96 × 96 with 62 axial slices of 2 × 2 × 2 mm3 large isotropic voxels. 250 volumes were obtained over the course of approximately 10 min. Subjects were instructed to keep their eyes closed, to refrain from engaging in any trains of thought as much as possible, and to not fall asleep.
Additional high-resolution T1-weighted anatomical images were acquired using an MPRAGE sequence with the following scanning parameters: TR 11 ms, TE 5.2 ms and flip angle 8°. 230 axial AC-PC slices encompassing a 384 × 384 matrix of 0.7 × 0.7 × 0.7 mm3 large isotropic voxels were obtained. All anatomical images underwent clinical inspection by a neuroradiological specialist to detect possible structural pathologies.
Preprocessing
Preprocessing was conducted using SPM 12 (The Wellcome Centre for Human Neuroimaging; http://www.fil.ion.ucl.ac.uk/spm). The preprocessing pipeline was created using RestPLUS63 during which the anatomical image was coregistered to the mean functional image and segmented into tissue probability maps used to create a group-specific DARTEL template64. Using these templates, the realigned functional time-series were normalized to MNI space and smoothed using a 4 × 4 × 4 mm3 full width at half maximum (FWHM) Gaussian Kernel. Additional preprocessing steps were performed using the CONN Toolbox v21.a65 and included denoising by regressing out white matter and cerebrospinal fluid using CompCor66, filtering time courses with a bandpass filter of 0.01 to 0.1 Hz, as well as de-trending and de-spiking.
Seed-based functional connectivity
Prior evidence demonstrated the differential involvement of hypothalamic and insular subnuclei. Therefore, bihemispheric medial and lateral hypothalamus or the anterior and posterior insula seeds were utilized. Canonical parcellations of the hypothalamus were extracted from the WFU PickAtlas 67, while parcellations of the insula were extracted from the Hammers Atlas68. Atlas selection was determined by both the availability and anatomic quality of seeds corresponding to our nuclei of interest. It is important to note that since no single atlas provided seed masks for all regions of interest (ROI), seeds from different atlas sources were selected according to their level of establishment and anatomic correspondence.
The seed-based FC analysis was conducted using the CONN Toolbox. First-level connectivity maps using Pearson correlation coefficients between the average time course of voxels within each ROI and every voxel (whole brain) were computed and transformed to z-scores. For the second-level analysis, the resulting FC maps were entered into a 2 × 2 mixed ANOVA, where training group was the between-subject factor and time was the within-subject factor. Results were simultaneously contrasted at MMT > HT and Post > Pre and thresholded at p < 0.05, FDR-corrected for multiple comparisons. To evaluate the directionality of the results, both positive and negative contrasts were applied and the p- value was adjusted to p < (0.05/2 =) 0.025, according to the Bonferroni method.
Correlations between changes in FC and behavioral measures for both MMT and HT groups were assessed. For every subject and time point, a mean signal from each significant cluster was extracted from the first-level maps using DPABI v6.069. The cluster-specific mean signal from the first time point was subtracted from the mean signal of the second time point, resulting in one value per subject which represented the change in cluster-specific FC after the intervention. Changes in behavioral measures were computed by calculating the difference scores (post–pre-intervention). Both values were then entered into a linear regression in SPSS. To avoid overcorrection, we incorporated the number of anatomical regions for multiple comparison corrections, resulting in an adjusted p-value of < (0.05/2 =) 0.025.
Results
Sample characteristics
Groups did not significantly differ in age, gender, years of education, or BMI (please see Table 1). Participants received their second MRI scan no later than 5 days upon completing the training condition.
Behavioral data
Groups did not differ significantly in behavioral measures obtained at baseline (see Table S3). Upon completing the training conditions, both groups displayed a slight decline in body weight (MMT: MPre = 69.8 kg, SD = 2.0; MPost = 69.4 kg, SD = 1.9; HT: MPre = 75.8 kg, SD = 2.5; MPost = 75.5 kg, SD = 2.6). This change, however, was not statistically significant for either group.
In line with our hypotheses, the MMT was found to effectively increase levels of perceived mindfulness demonstrated via an increase in MAAS (MPre = 53.9, SD = 10.6; MPost = 57.9, SD = 8.7). No increase in perceived mindfulness was observed in participants of the HT (MPre = 55.5, SD = 12.6; MPost = 54.9, SD = 11.6). An ANOVA was conducted and determined a significant group-by-time interaction (F (1, 64) = 7.74, p = 0.007, partial η2 = 0.108), including a significant main effect of time (F (1, 64) = 4.41, p = 0.040, partial η2 = 0.065).
Following the MMT, participants also reported lower FCQ-T scores indicating an overall reduction of food cravings (MPre = 84.0, SD = 30.3; MPost = 55.6, SD = 27.9). In the HT condition, FCQ-T scores demonstrated no significant difference upon completing the training (MPre = 79.4, SD = 34.6; MPost = 78.0, SD = 36.0). An ANOVA was conducted and resulted in a significant group-by-time interaction (F (1, 64) = 20.60, p < 0.001, partial η2 = 0.243) and a significant main effect of time (F(1, 64) = 24.90, p < 0.001, partial η2 = 0.280).
Furthermore, the MMT resulted in a significant reduction of stress- and emotional-eating as assessed by the SSES (MPre = 33.7, SD = 8.1; MPost = 31.8, SD = 6.7) and SEES (MPre = 64.3, SD = 7.1; MPost = 58.4, SD = 9.2). Participants of the HT, however, did not demonstrate any significant changes in stress-eating (MPre = 32.4, SD = 9.9; MPost = 31.8, SD = 10.5) or emotional-eating behavior (MPre = 61.8, SD = 10.0; MPost = 60.2, SD = 10.7). An ANOVA was conducted and determined a significant group-by-time interaction for both the SSES (F (1, 64) = 4.06, p = 0.048, partial η2 = 0.06) and SEES (F (1, 64) = 4.94, p = 0.030, partial η2 = 0.072) questionnaires as well as a significant main effect of time for both scales (SSES: F (1, 64) = 7.92, p = 0.006, partial η2 = 0.110; SEES: F (1, 64) = 15.28, p < 0.001, partial η2 = 0.193). It is important to note that a Levene’s test revealed that the homogeneity of error variances was not given for data pertaining to SSESPost and SEESPre. We therefore repeated the analysis by comparing individual difference scores with t-tests for independent samples between groups. Results for the SSES (t (64) = 2.02, two-sided p = 0.048) and the SEES (t (64) = 2.22, two-sided p = 0.030) remained consistent, thereby confirming the results of the prior analysis.
No significant group effects (F (1, 64) = 3.17, p = 0.080, partial η2 = 0.047) or main effects of time (F (1, 64) = 1.26, p = 0.267, partial η2 = 0.019) of the MMT on dietary restraint as measured by the Restraint scale were observed.
An overview of the behavioral results can be viewed in Fig. 2.

Visualization of the change in various self-report measures between T1 (pre-intervention) and T2 (post-intervention). Experimental group = MMT; Control group = HT.
Seed-based functional connectivity
The interaction results of the 2 × 2 ANOVA from the whole brain, FDR-corrected analyses revealed significant changes in hypothalamic and insular FC in participants of the MMT group whereas no changes in FC were observed in the HT group, although not all of these changes survived multiple comparisons correction.
The analyses using the left medial hypothalamus as a seed region demonstrated FC increases with two clusters within the right precuneus and an additional cluster within the right angular gyrus. FC analyses using the left medial hypothalamus demonstrated decreases in FC with a cluster extending across the left dorsal striatum and thalamus. The analyses conducted with the right lateral hypothalamus exhibited increased FC with a cluster within the left vPCC, whereas the analyses conducted with the left lateral hypothalamus yielded decreased FC with the left pre-supplementary motor area.
Additional FC increases were observed between the left anterior insula and bilateral clusters within the postcentral gyrus as well as a cluster within the right occipital gyrus. Increased FC was also observed between the left posterior insula and the left postcentral gyrus in addition to observing increases in FC between the right posterior insula and the right inferior parietal lobule.
An overview of the FC results can be seen in Table 2 and Fig. 3 and S2.

Overview of the FC changes observed between the hypothalamus (A), the insula (B), and the whole brain. HT Hypothalamus, BG Basal ganglia, PreSMA Pre supplementary motor area. PCC Precuneus, AG Angular gyrus, INS Insula, PoG Postcentral gyrus, IPL Inferior parietal lobe.
Interestingly, changes in hypothalamic connectivity patterns in the MMT group (but not the HT group) were accompanied by a multitude of changes on the behavioral level (see Fig. 4 and Table S4). For example, greater decoupling of FC between the left medial hypothalamus and the left dorsal striatum and thalamus significantly correlated with a greater reduction of FCQ-T scores. Additionally, SSES scores were inversely correlated with an increased FC between the right lateral hypothalamus and the left vPCC. These results indicate that the greater the decline in stress eating behavior, the greater the increase in FC between the lateral hypothalamus and the vPCC was observed. Furthermore, our results demonstrated that increases in MAAS score exhibited a trending towards significant with increased FC between the left medial hypothalamus and the right precuneus, whereas a negative correlation between MAAS score and an increase in FC between the left medial hypothalamus and the right angular gyrus was observed.

Scatter plots of significant and trending results of the linear regression between the change in functional connectivity and the change in behavioral measures. Dashed lines represent the 95% confidence intervals. FC Functional connectivity, R Right hemispheric, L Left hemispheric, lat. lateral, med. medial, AG Angular gyrus, vPCC Ventral posterior cingulate cortex, FCQT Food Cravings Questionnaire—trait, MAAS Mindful Attention and Awareness Scale, SSES Salzburg Stress Eating Scale).
It is important to note that FC changes of the insula did not correlate with any of the changes of behavioral measures, nor were any significant correlations observed between FC changes and behavioral measures of the HT group.
Discussion
This study could successfully demonstrate that MMT reduces stress-eating tendencies while also increasing perceived mindfulness. In fact, the observed behavioral changes significantly correlated with FC alterations demonstrating increased and decreased coupling of brain areas relevant to eating behavior, self-referential thinking, mind-wandering, reward perception, and the processing of sensory stimuli.
Self-Report Measures
Our results demonstrate a significant reduction in perceived stress-eating, emotional eating, and food cravings as well as an increase in perceived mindfulness in daily life. Our findings are in line with prior observations supporting the positive effects MMT has on stress, emotion regulation, interoceptive awareness, and perceived mindfulness70,71,72,73,74,75.
Functional connectivity measures
Hypothalamus
The results of this study demonstrate an increase in FC between subregions of the hypothalamus and multiple clusters within the precuneus, vPCC, and angular gyrus in the MMT group. Interestingly, these clusters pertain to hub regions of the DMN. The DMN is associated with self-referential thinking and mind-wandering76 and has been implicated in prior mindfulness research. For example, in comparison to meditation-naïve individuals, studies could demonstrate that experienced meditators yielded less connectivity between hubs of the DMN, which indicates an overall reduction of network activation44. In addition, a growing body of evidence suggests that MMT increases connectivity between the DMN and other networks, especially the salience network (SN)43,77, whose hub regions process emotional and sensory information76. These connectivity increases could be interpreted as an increase in awareness as a result of MMT43. Similarly, increased connectivity between hub regions of the DMN and the hypothalamus could indicate that MMT facilitates the perception of hunger and satiety, which could reflect increased interoceptive awareness, a cognitive process fundamental in the regulation of eating behavior.
Remarkably, these findings were corroborated by correlations observed between alterations in FC and behavioral changes. For example, increased FC between the medial hypothalamus and the precuneus positively correlated with an increase in MAAS scores. Therefore, when individuals perceive themselves to be more mindful, satiety cues may be more strongly linked with self-referential processing. An additional negative correlation between an increase in FC between the lateral hypothalamus and the vPCC with the reduction in SSES scores was observed. This suggests that the greater the increase in FC between these brain areas, the less participants perceived their tendency to succumb to stress-eating. Taken together, our findings indicate that MMT strengthens the interaction between hub regions of the DMN and the hypothalamus which goes along with increased levels of perceived mindfulness and a reduction in self-reported stress-eating behavior.
We could also observe MMT’s influence on the reward system through a reduction in FC between the left medial hypothalamus and the left dorsal striatum. The dorsal striatum has been demonstrated to elicit cravings and reward-seeking behaviors78. Therefore, a reduction in FC between the medial hypothalamus and the dorsal striatum could indicate a decoupling of brain areas responsible for processing feelings of satiety and reward. Notably, a significant, positive, correlation between the observed reduction in FC between these two brain areas and the decrease in FCQ-T scores was observed. This indicates that a greater reduction in FC between the lateral hypothalamus and the dorsal striatum was associated with a greater reduction in food cravings in participants of the MMT group. These results imply that feelings of satiety may be perceived as less rewarding, which could consequently reduce the tendency to develop food cravings.
Moreover, we could observe FC increases between the medial hypothalamus and the angular gyrus (AG) which is said to play a role in the integration of sensory information relevant to the processing of the smell and taste of food79. Interestingly, these connectivity increments negatively correlated with the reduction in perceived food cravings (via the FCQ-T) following the MMT. This indicates that MMT strengthens connectivity between areas involved in the processing of perceived satiety and integrating food-related sensory perception which goes along with a reduced tendency of displaying food cravings.
In addition, our results demonstrate alterations in FC between the left lateral hypothalamus and the left PreSMA, a brain region known to regulate automatic movement and reward behavior80,81. Our results may indicate that MMT reduces automatic, action-oriented behaviors when hunger is perceived. This could be attributed to more reflective decision-making about perceived hunger elicited via MMT prior to engaging in goal-directed behavior to find or eat food.
Overall, the findings pertaining to the hypothalamus support MMT’s role in cultivating a more mindful relationship with food. A recent systematic review conducted by Syan, McIntyre-Wood46 aimed to characterize hypothalamic resting state FC patterns in individuals with obesity in comparison to controls. Authors could identify patterns of aberrant DMN connectivity (i.e., hypoconnectivity), as well as increases in FC between the hypothalamus and regions attributed with limbic regions associated with reward processing. Interestingly, the MMT utilized in the present study induced opposing changes, i.e., increased hypothalamic connectivity with hubs of the DMN and a decrease of FC with limbic regions. This suggests that MMT might be able to counteract obesity-related neuronal activation patterns. In addition to our results demonstrating sensory integration and automatic eating behavior, these findings indicate that MMT might be able to beneficially influence neural correlates of stress-related eating behavior.
Insula
Our results demonstrated an increase in FC between both the anterior and the posterior division of the insula and bilateral clusters within the postcentral gyrus in the MMT-group. The postcentral gyrus, or primary somatosensory cortex, receives sensory information from the periphery; sensory input is organized topographically with tactile information originating from facial and oral sensations processed in the lateral postcentral gyrus which is where the clusters were observed82.
Given the emphasis MMT puts on the conscious perception of bodily, especially oral sensations, it seems plausible that areas responsible for perceiving somatosensory sensations in addition to areas that integrate the sensations into awareness are simultaneously activated. As it is known that regions and networks involved in sensory processing are stably active (also during rest), participants of the food-related MMT could demonstrate an increase in activation and connectivity within and between sensory areas pertaining to eating behavior at rest. In addition, the insula has been identified as part of the primary gustatory cortex, therefore responsible for the perception of taste83. An increase in the interaction between areas of the primary gustatory cortex and the postcentral gyrus suggest an increased integration of the sensory components, i.e., texture and taste, involved in the sensation of eating. Additional connectivity increases between the posterior insula and the supramarginal gyrus, an area associated with the somatosensory cortex, provide further evidence for the MMT-elicited integration of components involved in the processing of sensations. These results indicate that a food-related MMT can increase the efficiency of sensory integration all while facilitating increased awareness.
Given that the FC changes observed together with the insula involved brain regions essential in sensory processing and integration, a plausible explanation as to why no significant correlations between FC and self-report measures were observed may be because our self-report measures did not account for changes in sensory perception. Nevertheless, these results provide insight into MMT’s ability to alter FC pertaining specifically to the processing of sensory stimuli, which can consequently alter eating behavior.
Overall, the MMT-associated FC changes provide evidence of MMT’s ability to alter resting-state FC pertaining to regions of the DMN, reward perception, and the processing of sensory stimuli, in addition to providing evidence of MMT’s ability to reduce stress- and emotional-eating tendencies. Corresponding to our initial hypotheses, these results suggest that MMT might be an effective intervention strategy for potentially harmful eating behaviors, but more research is needed to confirm this. Moreover, following high methodological standards, this study was able to demonstrate that these changes are accompanied by specific alterations in brain function, thereby contributing to our understanding of the beneficial effects of mindfulness meditation.
Limitations
The results of this study should be interpreted under the consideration of methodological limitations. First, given our hypothesis-driven, a-priori selection of seed region, approach, our results may overlook relevant interactions. Nevertheless, the existing literature suggests that the hypothalamus and insula provide a relevant perspective on both mindfulness and eating behavior. Therefore, we believe that the results generated from the present study allow for a better understanding of the mechanisms underlying the intersection of mindfulness and eating behavior. While we acknowledge the complex nature of both eating behavior and MMT, and therefore do not rule out the possibility of further relevant influences, data-driven methods, e.g., independent component analyses should be utilized to further investigate its effects.
Future studies should also consider the influence of other potentially relevant variables, such as BMI, on susceptibility to MMT or similar interventions, e.g., by stratifying the sample by these variables. This could yield useful information for adjusting these interventions to individual requirements and, thereby, potentially improve their effectiveness.
Moreover, it is important to note that not all the results withstood multiple comparison corrections. While it would be a considerable alternative to make the thresholding more liberal, the recurrent patterns across ROIs, along with the statistical interdependence with behavioral measures, give sufficient grounds to assume the plausibility of these results.
Finally, stress-eating was measured using self-report, i.e., indirect, behavioral measures. While we acknowledge that these measures are not an immediate representation of behavior, we believe that the fact that we observed consistent changes across various scales demonstrates the ability of MMT to beneficially influence stress-related overeating behavior. A more immediate estimate of behavior could be better observed through fMRI with task-based paradigms. However, the functional organization of the brain during rest provides the foundation for behavior, i.e., through the formation of continuously activated or inactivated neuronal interaction patterns. Resting-state fMRI, therefore, not only makes it possible to investigate the neural foundation of stress-eating behavior but also allows for a broader perspective beyond a task-dependent state. Hence, we believe, that, despite their individual limitation, the synthesis of our behavioral and resting-state fMRI results conveys valuable information on how MMT influences the functional architecture of the brain and, thereby, cultivates a healthier relationship with food.
Data availability
Data have been made publicly available via the Open Science Framework at https://osf.io/pf3gv/.
References
McEwen, B. S. Physiology and neurobiology of stress and adaptation: central role of the brain. Physiol. Rev. 87(3), 873–904 (2007).
McEwen, B. S. & Stellar, E. Stress and the individual. Mechanisms leading to disease. Arch. Intern. Med. 153(18), 2093–2101 (1993).
McEwen, B. S. Central effects of stress hormones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. Eur. J. Pharmacol. 583(2–3), 174–185 (2008).
Bose, M., Oliván, B. & Laferrère, B. Stress and obesity: The role of the hypothalamic–pituitary–adrenal axis in metabolic disease. Curr. Opin. Endocrinol. Diabetes Obes. 16(5), 340–346 (2009).
Holmes, M. E., Ekkekakis, P. & Eisenmann, J. C. The physical activity, stress and metabolic syndrome triangle: A guide to unfamiliar territory for the obesity researcher. Obes. Rev. 11(7), 492–507 (2010).
Kuo, L. E. et al. Neuropeptide Y acts directly in the periphery on fat tissue and mediates stress-induced obesity and metabolic syndrome. Nat. Med. 13(7), 803–811 (2007).
Sojcher, R., Fogerite, S. G. & Perlman, A. Evidence and potential mechanisms for mindfulness practices and energy psychology for obesity and binge-eating disorder. Explore 8(5), 271–276 (2012).
Adam, T. C. & Epel, E. S. Stress, eating and the reward system. Physiol. Behav. 91(4), 449–458 (2007).
Epel, E. et al. Stress may add bite to appetite in women: A laboratory study of stress-induced cortisol and eating behavior. Psychoneuroendocrinology 26(1), 37–49 (2001).
Groesz, L. M. et al. What is eating you? Stress and the drive to eat. Appetite 58(2), 717–721 (2012).
Oliver, G., Wardle, J. & Gibson, E. L. Stress and food choice: A laboratory study. Psychosom. Med. 62(6), 853–865 (2000).
Torres, S. J. & Nowson, C. A. Relationship between stress, eating behavior, and obesity. Nutrition 23(11–12), 887–894 (2007).
Mathews, A. & MacLeod, C. Cognitive vulnerability to emotional disorders. Annu. Rev. Clin. Psychol. 1, 167–195 (2005).
Stansbury, K. & Gunnar, M. R. Adrenocortical activity and emotion regulation. Monogr. Soc. Res. Child Dev. 59(2–3), 108–134 (1994).
Wang, Y. & Beydoun, M. A. The obesity epidemic in the United States–gender, age, socioeconomic, racial/ethnic, and geographic characteristics: A systematic review and meta-regression analysis. Epidemiol. Rev. 29, 6–28 (2007).
Hawkes, C. Uneven dietary development: linking the policies and processes of globalization with the nutrition transition, obesity and diet-related chronic diseases. Global. Health 2(1), 4 (2006).
Lake, A. & Townshend, T. Obesogenic environments: Exploring the built and food environments. J. R. Soc. Promot. Health 126(6), 262–267 (2006).
Swinburn, B. A. et al. The global obesity pandemic: shaped by global drivers and local environments. Lancet 378(9793), 804–814 (2011).
Yeomans, M. R. Olfactory influences on appetite and satiety in humans. Physiol. Behav. 87(4), 800–804 (2006).
Bray, G. A. Medical consequences of obesity. J. Clin. Endocrinol. Metab. 89(6), 2583–2589 (2004).
Epel, E. S. et al. Stress and body shape: Stress-induced cortisol secretion is consistently greater among women with central fat. Psychosom. Med. 62(5), 623–632 (2000).
Gami, A. S. et al. Metabolic syndrome and risk of incident cardiovascular events and death: A systematic review and meta-analysis of longitudinal studies. J. Am. Coll. Cardiol. 49(4), 403–414 (2007).
Benjamin, E. J. et al. Heart disease and stroke statistics-2019 update: A report from the American heart association. Circulation 139(10), e56–e528 (2019).
Hales, C. M. et al. Prevalence of obesity and severe obesity among adults: United States, 2017–2018. NCHS Data Brief. 360, 1–8 (2020).
Virani, S. S. et al. Heart Disease and stroke statistics-2020 update: a report from the American heart association. Circulation 141(9), e139–e596 (2020).
Creswell, J. et al. Brief mindfulness meditation training alters psychological and neuroendocrine responses to social evaluative stress. Psychoneuroendocrinology 44(1), 12 (2014).
Goyal, et al. Meditation programs for psychological stress and well-being: a systematic review and meta-analysis. JAMA Intern. Med. 174(3), 357–368 (2014).
Hariprasad, V. R. et al. Yoga increases the volume of the hippocampus in elderly subjects. Indian J. Psychiatry 55(Suppl 3), S394–S396 (2013).
Hölzel, B. K. et al. Mindfulness practice leads to increases in regional brain gray matter density. Psychiatry Res. Neuroimaging 191(1), 36–43 (2011).
Luders, E. et al. Global and regional alterations of hippocampal anatomy in long-term meditation practitioners. Hum. Brain Mapp. 34(12), 3369–3375 (2013).
Pickut, B. A. et al. Mindfulness based intervention in Parkinson’s disease leads to structural brain changes on MRI: A randomized controlled longitudinal trial. Clin. Neurol. Neurosurg. 115(12), 2419–2425 (2013).
Sharma, M. & Rush, S. E. Mindfulness-based stress reduction as a stress management intervention for healthy individuals. J. Evid.-Based Complement. Altern. Med. 19(4), 271–286 (2014).
Kabat-Zinn, J. Mindfulness-based interventions in context: past, present, and future. Clin. Psychol. Sci. Pract. https://doi.org/10.1093/clipsy/bpg016 (2003).
Butzer, B. et al. School-based yoga programs in the United States: A survey. Adv. Mind Body Med. 29(4), 18–26 (2015).
Chong, C. S. et al. Effects of yoga on stress management in healthy adults: A systematic review. Altern. Ther. Health Med. 17(1), 32–38 (2011).
D’Silva, S. et al. Mind-body medicine therapies for a range of depression severity: A systematic review. Psychosomatics 53(5), 407–423 (2012).
Gard, T. et al. Potential self-regulatory mechanisms of yoga for psychological health. Front. Hum. Neurosci. 8, 770 (2014).
Lemay, V., Hoolahan, J. & Buchanan, A. Impact of a yoga and meditation intervention on students’ stress and anxiety levels. Am. J. Pharm. Educ. 83(5), 7001 (2019).
Wahbeh, H., Elsas, S. M. & Oken, B. S. Mind-body interventions: Applications in neurology. Neurology 70(24), 2321–2328 (2008).
Creswell, J. & Lindsay, E. How does mindfulness training affect health? A mindfulness stress buffering account. Curr. Dir. Psychol. Sci. 23, 401–407 (2014).
Marcus, M. T. et al. Change in stress levels following mindfulness-based stress reduction in a therapeutic community. Addict. Disord. Their Treat. 2(3), 63–68 (2003).
Miller, R. L. et al. Effects of a mindfulness-induction on subjective and physiological stress response in adolescents at-risk for adult obesity. Eat. Behav. 40, 101467 (2021).
Bremer, B. et al. Mindfulness meditation increases default mode, salience, and central executive network connectivity. Sci. Rep. https://doi.org/10.1038/s41598-022-17325-6 (2022).
Brewer, J. A. et al. Meditation experience is associated with differences in default mode network activity and connectivity. Proc. Natl. Acad. Sci. 108(50), 20254–20259 (2011).
Kilpatrick, L. A. et al. Impact of mindfulness-based stress reduction training on intrinsic brain connectivity. NeuroImage 56(1), 290–298 (2011).
Syan, S. K. et al. Dysregulated resting state functional connectivity and obesity: A systematic review. Neurosci. Biobehav. Rev. 131, 270–292 (2021).
Tang, Y.-Y., Hölzel, B. K. & Posner, M. I. The neuroscience of mindfulness meditation. Nat. Rev. Neurosci. 16(4), 213–225 (2015).
Aston-Jones, G. et al. Role of lateral hypothalamic orexin neurons in reward processing and addiction. Neuropharmacology 56(Suppl 1), 112–121 (2009).
Bernardis, L. L. & Bellinger, L. L. The lateral hypothalamic area revisited: ingestive behavior. Neurosci. Biobehav. Rev. 20(2), 189–287 (1996).
Saper, C. B., Chou, T. C. & Elmquist, J. K. The need to feed: homeostatic and hedonic control of eating. Neuron 36(2), 199–211 (2002).
Craig, A. D. How do you feel—now? The anterior insula and human awareness. Nat. Rev. Neurosci. 10(1), 59–70 (2009).
Rolls, E. T. & McCabe, C. Enhanced affective brain representations of chocolate in cravers vs. non-cravers. Eur. J. Neurosci. 26(4), 1067–1076 (2007).
Small, D. M. Taste representation in the human insula. Brain Struct. Funct. 214(5–6), 551–561 (2010).
Torske, A. et al. Localizing the human brain response to olfactory stimulation: A meta-analytic approach. Neurosci. Biobehav. Rev. 134, 104512 (2022).
Hölzel, B. K. et al. Investigation of mindfulness meditation practitioners with voxel-based morphometry. Soc. Cogn. Affect. Neurosci. 3(1), 55–61 (2007).
Laneri, D. et al. Mindfulness meditation regulates anterior insula activity during empathy for social pain. Hum. Brain Mapp. 38(8), 4034–4046 (2017).
Farb, N. A. S., Segal, Z. V. & Anderson, A. K. Mindfulness meditation training alters cortical representations of interoceptive attention. Soc. Cogn. Affect. Neurosci. 8(1), 15–26 (2012).
Critchley, H. D. et al. Neural systems supporting interoceptive awareness. Nat. Neurosci. 7(2), 189–195 (2004).
Rolls, E. T. Brain mechanisms underlying flavour and appetite. Philos. Trans. R. Soc. Lond B Biol. Sci. 361(1471), 1123–1136 (2006).
Cohen, S., Kamarck, T. & Mermelstein, R. A global measure of perceived stress. J. Health Soc. Behav. https://doi.org/10.2307/2136404 (1983).
Bontempi, C., Jacquot, L. & Brand, G. Diet and odor hedonic ratings: comparative study between vegetarians flexitarians, and omnivores. Nutr. Neurosci. https://doi.org/10.1080/1028415X.2022.2145425 (2022).
Brown, K. W. & Ryan, R. M. The benefits of being present: mindfulness and its role in psychological well-being. J. Personal. Soc. Psychol. 84(4), 822 (2003).
Xi-Ze, J. et al. RESTplus: an improved toolkit for resting-state functional magnetic resonance imaging data processing. Sci. Bull. 64, 953–954 (2019).
Ashburner, J. A fast diffeomorphic image registration algorithm. NeuroImage 38(1), 95–113 (2007).
Whitfield-Gabrieli, S. & Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2(3), 125–141 (2012).
Behzadi, Y. et al. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 37(1), 90–101 (2007).
Maldjian, J. A. et al. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. Neuroimage 19(3), 1233–1239 (2003).
Hammers, A. et al. Three-dimensional maximum probability atlas of the human brain, with particular reference to the temporal lobe. Hum. Brain Mapp. 19(4), 224–247 (2003).
Yan, C.-G. et al. DPABI: Data processing & analysis for (Resting-State) brain imaging. Neuroinformatics 14(3), 339–351 (2016).
Chiesa, A. & Serretti, A. Mindfulness-based stress reduction for stress management in healthy people: A review and meta-analysis. J. Altern. Complement. Med. 15(5), 593–600 (2009).
Kabat-Zinn, J. Mindfulness-based stress reduction (MBSR). Construct. Hum. Sci. 8(2), 73 (2003).
Khoury, B. et al. Mindfulness-based stress reduction for healthy individuals: A meta-analysis. J. Psychosom. Res. 78(6), 519–528 (2015).
Roemer, L., Williston, S. K. & Rollins, L. G. Mindfulness and emotion regulation. Curr. Opin. Psychol. 3, 52–57 (2015).
Teper, R., Segal, Z. V. & Inzlicht, M. Inside the mindful mind: How mindfulness enhances emotion regulation through improvements in executive control. Curr. Dir. Psychol. Sci. 22(6), 449–454 (2013).
Fissler, M. et al. An investigation of the effects of brief mindfulness training on self-reported interoceptive awareness, the ability to decenter, and their role in the reduction of depressive symptoms. Mindfulness 7(5), 1170–1181 (2016).
Menon, V. Large-scale brain networks and psychopathology: A unifying triple network model. Trends Cogn. Sci. 15(10), 483–506 (2011).
Rahrig, H. et al. Meta-analytic evidence that mindfulness training alters resting state default mode network connectivity. Sci. Rep. https://doi.org/10.1038/s41598-022-15195-6 (2022).
Grall-Bronnec, M. & Sauvaget, A. The use of repetitive transcranial magnetic stimulation for modulating craving and addictive behaviours: A critical literature review of efficacy, technical and methodological considerations. Neurosci. Biobehav. Rev. 47, 592–613 (2014).
Contreras-Rodriguez, O. et al. Neural-based valuation of functional foods among lean and obese individuals. Nutr. Res. 78, 27–35 (2020).
Nachev, P. et al. The role of the pre-supplementary motor area in the control of action [Supplement 2]. NeuroImage 36(Suppl 2), T155–T163 (2007).
Tabu, H. et al. Functional relevance of pre-supplementary motor areas for the choice to stop during Stop signal task. Neurosci. Res. 70(3), 277–284 (2011).
Grabenhorst, F. & Rolls, E. T. The representation of oral fat texture in the human somatosensory cortex. Hum. Brain Mapp. 35(6), 2521–2530 (2014).
Iannilli, E. et al. Spatio-temporal correlates of taste processing in the human primary gustatory cortex. Neuroscience 273, 92–99 (2014).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Study conception and design: AT, BKH, KK; data collection: AT, AM; analysis and interpretation of results: AT, BB; draft manuscript preparation: AT, BB. All authors reviewed the results and approved the current version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
This study was supported by the German Ministry for Education and Research (BMBF) grant to author KK (BMBF 01EA1807H), and the preparation of this paper was supported by the enable Cluster (http://enable-cluster.de) in addition to a private donation made by Maria Tussi Kluge. Author AT was additionally funded by the Studienstiftung des Deutschen Volkes. Author BKH is the author of commercially available books and online mindfulness-based interventions. The remaining authors declare that they have no conflict of interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Torske, A., Bremer, B., Hölzel, B.K. et al. Mindfulness meditation modulates stress-eating and its neural correlates. Sci Rep 14, 7294 (2024). https://doi.org/10.1038/s41598-024-57687-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-57687-7
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
