
Abstract
This study aimed to investigate structural synaptic plasticity in the medial prefrontal cortex of rats under treadmill exercise pretreatment or naive conditions in a vascular dementia model, followed by recognition memory performance in a novel object recognition task. In this study, 24 Sprague–Dawley rats were obtained and randomly assigned into 4 groups as follows: control group (Con group, n = 6), vascular dementia (VD group, n = 6), exercise and vascular dementia group (Exe + VD group, n = 6), and exercise group (Exe group, n = 6). Initially, 4 weeks of treadmill exercise intervention was administered to the rats in the Exe + VD and Exe groups. Then, to establish the vascular dementia model, the rats both in the VD and Exe + VD groups were subjected to bilateral common carotids arteries surgery. One week later, open-field task and novel recognition memory task were adopted to evaluate anxiety-like behavior and recognition memory in each group. Then, immunofluorescence and Golgi staining were used to evaluate neuronal number and spine density in the rat medial prefrontal cortex. Transmission electron microscopy was used to observe the synaptic ultrastructure. Finally, microdialysis coupled with high-performance liquid chromatography was used to assess the levels of 5-HT and dopamine in the medial prefrontal cortex. The behavior results showed that 4 weeks of treadmill exercise pretreatment significantly alleviated recognition memory impairment and anxiety-like behavior in VD rats (P < 0.01), while the rats in VD group exhibited impaired recognition memory and anxiety-like behavior when compared with the Con group (P < 0.001). Additionally, NeuN immunostaining results revealed a significant decrease of NeuN-marked neuron in the VD group compared to Con group (P < 0.01), but a significantly increase in this molecular marker was found in the Exe + VD group compared to the Con group (P < 0.01). Golgi staining results showed that the medial prefrontal cortex neurons in the VD group displayed fewer dendritic spines than those in the Con group (P < 0.01), and there were more spines on the dendrites of medial prefrontal cortex cells in Exe + VD rats than in VD rats (P < 0.01). Transmission electron microscopy further revealed that there was a significant reduction of synapses intensity in the medial prefrontal cortex of rats in the VD group when compared with the Con group(P < 0.01), but physical exercise was found to significantly increased synapses intensity in the VD model (P < 0.01). Lastly, the levels of dopamine and 5-HT in the medial prefrontal cortex of rats in the VD group was significantly lower compared to the Con group (P < 0.01), and treadmill exercise was shown to significantly increased the levels of dopamine and 5-HT in the VD rats (P < 0.05). Treadmill exercise pretreatment ameliorated structural synaptic plasticity impairments of medial prefrontal cortex in VD rat and improved recognition memory.
Similar content being viewed by others

Introduction
VD involve the remarkable vascular cognitive impairment cause by multiple vascular factors. In the early and developmental stages of VD, it most often characterized by various degrees cognitive decline1. Neurobiologists have made several attempts to elucidate the mechanism underlying the VD-induced memory impairment2,3. Accumulating evidence aimed to elucidate the link between VD and memory function has focused on synaptic plasticity, which could also be a potential target for treatment interventions. Synaptic plasticity, or the change in connections between neurons caused by experience, underpins learning and memory in the brain4. VD can induce a deficit of synaptic plasticity in the hippocampus5,6, striatum7 and corpus callosum8 regions, such as the decrease of dendritic spine density and synaptic proteins. It has been known for a few years that acupuncture9, social interaction10 and exercise11 have a positive influence in patients with VD. It is worth mentioning that exercise can not only ameliorate VD symptoms, but also prevent the development at an early stage of VD12,13. Furthermore, there is evidence supporting the conclusion that the neuroprotective effect of exercise in a VD model14. However, it remains to be clarified whether exercise has a positive effect in medial prefrontal cortex regions.
Recent research advances in humans15 and animals16 have demonstrated that physical exercise has a significant effect on learning and memory. There is growing evidence that exercise can enhance hippocampal neurogenesis in normal17 or diseases states, such as APP/PS1 transgenic18, stress19, and VD models20. To date, an increasing number of researchers have further explored the mechanism underlying exercise-induced memory improvement. Structural synaptic plasticity, which is necessary for memory formation during brain development, provides a possible explanation21. Exercise is a non-drug therapy that has proved to be beneficial for increasing structural synaptic plasticity in different regions of the brain22,23,24. As an important region in the brain, the medial prefrontal cortex is closely related to learning memory and execution function25. It has a high neuronal density and metabolic rate, marking it susceptible to ischemia and hypoxia injury26,27. Prefrontal cortex injury caused by prolonged bilateral common carotid artery ligation can significantly affect higher brain functions, such as learning and memory functions26,28. However, no previous research has investigated the effect of exercise on VD via structural synaptic plasticity in the prefrontal cortex region. Therefore, the current study sought to determine whether treadmill exercise pretreatment can alleviate recognition memory impairment by regulating structural synaptic plasticity parameters of the prefrontal cortex in a VD rat model.
Materials and methods
Experimental animals and grouping
Sprague–Dawley rats (males 200–225 g; Shanghai SLAC Laboratory Animal Co., Ltd) were housed (three per cage; cage size:460 × 300 × 160 mm) socially in a controlled room (Temperature: 22 ± 3 °C; Humidity: 40 to 70%) on a reversed 12 h light–dark cycle (light off at 6 AM). Water and food were made available all the time. All the animal procedures and methods conducted in this study were approved by the Ethics Committee of Experimental Animals of Capital University of Physical Education and Sport (Approval number 2020A76). We confirm that all methods were performed in accordance with the relevant guidelines and regulations and ARRIVE guidelines。
Rats in this experiment were given a one-week period to adapt to their environment. Then, the rats (n = 24) were randomly divided into 4 groups as follows: control group (Con group, n = 6), VD group (n = 6), exercise and vascular dementia group (Exe- VD group, n = 6), and exercise group (Exe group, n = 6). The treadmill exercise interventions and the establishment of VD model are in accordance with Fig. 1. Regarding the behavioral task, all the animals were subjected to an open-field task and a novel recognition memory task. Subsequently, all the animals were sacrificed for further assays. NeuN immunostaining was used to evaluate the quantity of positive cells in the medial prefrontal cortex. Golgi staining was used to analyze morphological alterations in neuronal dendritic spines. Transmission electron microscopy was used for in vivo synaptic imaging. Microdialysis coupled with high-performance liquid chromatography was used to analyze the level of 5-HT and dopamine in the medial prefrontal cortex.

Schematic timeline of study design.
Treadmill exercise pretreatment
In this study, treadmill exercise was applied for 4 weeks. Thus, the rats in the Exe + VD and Exe groups were forced to run on a treadmill device (BHW-PT/5 s, Anhui, China) for 4 weeks. All the rats involved in the intervention groups were conditioned for treadmill exercise for 30 min on 3 consecutive days (at 8 m/min on the first day and 12 m/min on the second and third days). Subsequently, the rats were subjected to a treadmill exercise pretreatment on a 0° slope, starting with a speed of 12 m/min for 10 min. Then, this was scaled up to 15 m/min for 50 min. In sum, all the rats in the Exe + VD and Exe groups were trained for an hour per day (6 PM to 10 PM), 5 days per week, for a total of 4 weeks. According to previous study, the intensity of treadmill exercise in this experimentwas categorized as low intensity exercise29. The running speed suitable for treadmill exercise was determined based on our previous studies that showed protective effects against AD and other brain diseases29,30.
VD model
The VD model was established using the two vessel occlusion method according to previous studies31,32. First, the animals in the VD and Exe + VD groups were anesthetized with an intraperitoneal (IP) injection of 1% pentobarbital sodium (60 mg/kg). Regarding the two-vessel occlusion procedure, the skin of the neck was prepped with alcohol. Then, a ventral midline cervical incision was made, through which we identified and isolated the bilateral common carotid arteries (BCCAs) from the vagus. Subsequently, BCCAs were gently isolated and ligated with 10–0 silk sutures. The wound area was treated locally with penicillina after the incision was sutured. Afterwards, the general condition and body weight of each rat were monitored for the next 3 days. The mortality rate after BCCA occlusion in this study is 14.29%. After 1 week, the “Zea-Longa” 5-point scale was used to confirm whether the animal model had been established successfully. The success rate of establishing VD rat model is 92.31%.
Open-field test
The open-field test (OFT) measures spontaneous locomotor activity as well as anxiety-like behaviors33. This test was conducted with apparatus being placed on a square arena (Black; 100 cm × 100 cm × 50 cm), and the bottom of the arena was divided into two zones: an outside and a center zone. At the beginning of testing, the animal was placed in a particular corner of the area, and its behavior was recorded over 10 min. Then, 70% ethanol was used to clean the test apparatus, followed by drying with paper towels, and the rat were returned to their home cage. In this test, total distance, time traveled in every zone, number of entries into every zone and were recorded.
Novel object recognition task
The recognition memory and the behavior to explore a new environment were evaluated via the novel object recognition task34. The rats were allowed to freely explore two identical objects for 10 min on the first day. The test phase began after a delay of 5 min (short-term recognition memory) or 24 h (long-term recognition memory), during which one of the objects was replaced by a novel one (similar size texture and color, but different in shape), and the rats were allowed to explore it for 10 min. Under normal states, rats often tend to spend more time examining unfamiliar objects. As a result, the time spent by a rat in exploring the novel object was used as a measure of memory. Thus, the novel object discrimination index was determined by dividing the time interacting with the novel object by the total time explored both objects (i.e., [Time Novel/(Time Novel + Time Familiar)] × 100%).
Immunofluorescence
Regarding NeuN immunostaining, the number of positive cells in the medial medial prefrontal cortex region was counted. After perfusion with saline and 4% paraformaldehyde, the whole brain tissues were quickly removed, and the medial prefrontal cortex block were fixed in 4% paraformaldehyde overnight at 4 °C and cryoprotected in sucrose. Coronal sections were cut at 30 μm thickness using a cryostat. Then, the coronal sections were subjected to fluorescence immunostaining using primary antibodies (rabbit monoclonal to NeuN—Neuronal marker, 1:200, ab177487, Abcam) at 4 °C overnight, followed by secondary antibody staining (Alexa Fluor 488-conjugated goat anti-Rabbit IgG, 1:1000, ab150077, Abcam) to verify neuron expression. Images were acquired with a confocal fluorescence microscope (Olympus) under 400 × magnification. All the measurements for immunofluorescent observation were performed in 3 rats from each group.
Golgi staining
The FD Rapid Golgi Stain™ Kit (FD Neurotechnologies, Inc) was used to perform Golgi staining. This can evaluate minimal morphological changes in neuronal dendrites and dendritic spines35. Briefly, isolated tissue blocks (prefrontal cortex) were rinsed quickly with PBS and immersed in the fixative for 48 h. Then, tissue blocks were added to the Golgi-cox staining solution and incubated for 14 days (new staining solution was changed after 48 h, and then every 3 days). After treatment with 80% glacial acetic acid and 30% sucrose, the tissue blocks were sliced into sections of 100 microns using an oscillating microtome and pasted on a gelatin slide. Then, the tissue blocks were hardened and fixed with acid for 15 min, distilled water was used to wash for 3 min,
and the sections were dried and sealed using glycerin gelatin. Finally, a digital slice scanner (Nikon DS-U3) was used to acquire panoramic images of brain tissue, which was analyzed using ImageJ (Fiji) software.
Transmission electron microscopy
For the quantification of medial prefrontal cortex synaptic changes, transmission electron microscopy was used to analyze the number of synapses36. To analyze synaptosomes, the medial prefrontal cortex block (no more than 1 mm3) was washed with PBS and immediately placed into an EP tube containing TEM fixative at 4 °C. Then, the tissue was fixed using 1% OsO4 in 0.1 M PB (pH 7.4) at room temperature for 7 h, and rinsed 3 times in 0.1 M PB (pH 7.4). Dehydration was performed at room temperature using 30–100% ethanol and pure acetone for 1 h. Subsequently, the tissue was subjected to resin penetration and embedding as well as stored overnight in an oven at 37 °C. The tissue embedding model was transferred to a hotter oven (65 °C) and allowed to polymerize for > 48 h. Then, the tissue was sectioned into 60–80 nm slices using an ultra-microtome (Leica UC7) and collected on copper grids, followed by examination with transmission electron microscopy (Hitachi). The number of synapse was counted under the 200 × magnification and calculated by ImageJ.
High-performance liquid chromatography for DA tissue content
After behavior test, animals were placed in a stereotaxic apparatus to ensure the skull is at the same level. Then, the bregma is regarded as origin of coordinates, microdialysis (MD) probe was stereotaxically implanted in the rat medial prefrontal cortex (A/P, + 3.2 mm; M/L, ± 0.6 mm and D/V, − 1.5 mm from dura), and extracellular fluid was collected in the EP tubes. When obtained extracellular fluid of each rats, high-performance liquid chromatography with electrochemical detection was used to determine the levels of 5-HT and DA in extracellular fluid. Before the analysis, the samples were diluted with ice-cold mobile phase that was deoxygenated with argon in a 1:10 ratio, followed by direct injection into the high-performance liquid chromatography device37. The mobile phase was flowing at a rate of 0.5 mL/min.
Data analysis
Data in this experiment were summarized as mean ± SEM. All statistical analyses were performed by GraphPad Prism software (GraphPad Software, La Jolla, CA, USA). One-way analysis of variance was used to compare means among multiple groups, followed by Tukey’s post hoc test. P < 0.05 was chosen as the level of statistical significance.
Results
Treadmill exercise pretreatment ameliorated anxiety behavior in VD rat
Results were obtained for all behavioral parameters in the OFT (Fig. 2). There was no significant difference in total distance traveled among the four groups (P > 0.001; Fig. 2A). In terms of the time spent in the central zone, the rats in the VD group spent significantly lesser time than those in the Con group (P < 0.001; Fig. 2B). Moreover, the rats in the Exe + VD group spent significantly more time in the central zone than those in the VD group (P < 0.001; Fig. 2B).

Treadmill exercise prevented anxiety- like behavior after VD in open field test. The figure shows total distance traveled (A), time traveled in each area (B), number of entries into each area (C). N = 6/group. #P < 0.05 and ##P < 0.01 vs. Con group; *P < 0.05 and **P < 0.01 vs. VD group.
In addition, rats in the VD group had fewer number of times to enter the central area than those in the Con group (P < 0.01; Fig. 2C). After treadmill exercise pretreatment, there was a significantly increased number of times to enter the central area in the Exe + VD rats when compared with VD group (P < 0.01, P < 0.001; Fig. 2C).
Treadmill exercise pretreatment ameliorated recognition memory impairment in VD rat
To assess novelty discrimination, a novel object recognition task was adopted in each group. In this test, the discrimination index of rats was significantly lower in the VD group as compared to the Con group (P < 0.001; Fig. 3), which indicating impaired object discrimination of VD rats. Conversely, the discrimination index of rats in the Exe + VD group was significantly greater than that of rats in the VD group (P < 0.01; Fig. 3), which suggests the VD rats have a positive trend of object recognition after exercise interventions. Thus, these results suggest that treadmill exercise pretreatment can improve VD-induced recognition memory for objects that had been previously encountered.

Discrimination index in test phase of novel object recognition task. N = 6/group. ###P < 0.001vs. Con group; **P < 0.01 and ***P < 0.001 vs. VD group.
Treadmill exercise pretreatment enhanced neuron density of medial prefrontal cortex in VD rat
To investigate whether there is an association between recognition memory restoration via treadmill exercise pretreatment restored in recognition memory and structural synaptic plasticity, we first counted the number of neurons in the cortex necessary for the transmission of information linked to memory and learning. NeuN immunostaining was conducted to examine the expression of NeuN-marked neurons in rat medial prefrontal cortex from the four groups listed in Fig. 4A. The results showed a significant decrease in NeuN-marked intensity in Con group when compared to VD group (P < 0.01; Fig. 4B). Regarding the effects of treadmill exercise on NeuN expression in the VD model, we found a significant increase in this molecular marker in rats that received a systemic treadmill exercise compared to the Con group (P < 0.01; Fig. 4B). Thus, we concluded that treadmill exercise pretreatment can ameliorate VD-induced the decrese of neuron intensity in the medial prefrontal cortex.

Quantification analysis of single immunofluorescent staining with mouse anti-NeuN primary.. N = 3/group. ##P < 0.01 vs. Con group; **P < 0.01 vs. VD group.
Treadmill exercise pretreatment enhanced the spine density of the medial prefrontal cortex in VD rat
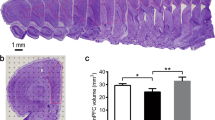
Golgi staining was performed to investigate whether treadmill exercise pretreatment alters the manifestation of dendritic spine in VD brain tissue, which are presented in Fig. 5A. We found that the medial prefrontal cortex neurons in the VD group revealed a significantly lower quantity of dendritic spines than those in the Con group (P < 0.01; Fig. 5B). Conversely, significantly differences were observed of the dendritic spine number in the Exe + VD group when compared with VD group. (P < 0.01; Fig. 5B). Thus, treadmill exercise pretreatment can inhibited VD-induced impairment of dendritic spines in the medial prefrontal cortex.

(A) Golgi-stained basal dendrites of medial prefrontal cortex pyramidal neurons in each group. (B) Quantification of mature spine density of the dendritic segments. N = 3/group. ##P < 0.01 vs. Con group; **P < 0.01 vs. VD group; &P < 0.01 vs. Con group.
Treadmill exercise pretreatment inhibited ultrastructure impairment of the medial prefrontal cortex in VD rat
We quantified the number of synapses in the medial prefrontal cortex, which are necessary for the establishment of recognition memory (Fig. 6A). The results of transmission electron microscopy revealed a significant reduction in the synapse density in the medial prefrontal cortex of rats in the VD group compared to the Con group (P < 0.05; Fig. 6B). Conversely, there was a significant increase in the synapse density in the medial prefrontal cortex of VD rats following the treadmill exercise pretreatment (P < 0.05; Fig. 6B). These data supported the hypothesis that treadmill exercise pretreatment can increase the synapse density in the medial prefrontal cortex of VD rats.

Representative images of ultrastructure from the medial prefrontal cortex neurons (A) and quantitative analysis of the density of synapse (B) are shown. The synapses are marked by the blue arrowheads. N = 3/group. #P < 0.05 vs. Con group; *P < 0.05 vs. VD group.
Treadmill exercise pretreatment inhibited the decrease of medial prefrontal cortex dopamine and 5-HT in VD rat
We observed dynamic changes in medial prefrontal cortex dopamine and 5-HT in the four groups. Our results revealed that the level of medial prefrontal cortex dopamine and 5-HT in the VD group was significantly lower when compared to the Con group (P < 0.01; Fig. 7). On the contrary, the level of medial prefrontal cortex dopamine and 5-HT in the Exe + VD group was significantly higher compared to the VD group, (P < 0.05; Fig. 7). The results indicated that treadmill exercise pretreatment can increase the level of medial prefrontal cortex dopamine and 5-HT in VD rats.

Dynamic changes of medial prefrontal cortex dopamine and 5-HT in four group. A: Dopamine B:5-HT. N = 3/group. ##P < 0.01 vs. Con group; *P < 0.05 and **P < 0.01 vs. VD group.
Discussion
Our recent study demonstrated that VD rats exhibited impaired recognition memory, while treadmill exercise pretreatment was found to prevent recognition memory impairment by enhancing structural synaptic plasticity in a VD model. Moreover, we demonstrated that in a VD rat model established by prolonged two-vessel occlusion, treadmill exercise pretreatment ameliorates recognition memory impairment and anxiety-like behavior in a novel object recognition task and an open-field test. Following the treadmill exercise pretreatment, there was a resultant increase in neurons, dendritic spines density, density of synapses, and repair of structural synaptic plasticity of the medial prefrontal cortex in VD rats. Therefore, a possible mechanism through which treadmill exercise ameliorates recognition memory impairment and anxiety-like behavior could be by strengthening structural synaptic plasticity of the medial prefrontal cortex.
In the behavior test, after subjecting the rats in each group to behavioral tests (novel object recognition tasks and open-field), we found that the discrimination index of recognition memory was significantly decreased in the VD group compared to the Con group. Recognition memory is often an essential part of cognitive function. As mentioned above, the two-vessel occlusion-induced VD rat model is characterized by damage to the hippocampus and medial prefrontal cortex, caused by prolonged cerebral hypoperfusion, which results in cognitive deficits. As the VD disease progresses, the patients gradually experience recognition memory impairment leading to logical memory impairments and declarative metamemory disorders. Moreover, VD patients were found to exhibited anxiety-like behavior38, and this phenomenon was also found in our study and others animal studies6,39. Thus, we suggested that significant recognition memory impairment and anxiety-like behavior were present in the VD model. We also found that the discrimination index of rats in the Exe + VD group was significantly greater than that of rats in the VD group. This indicates that the rats in the Exe + VD group could recognize the presence of a novel object and exhibited recall the memory of an object previously observed. This result suggested that 4-week treadmill exercise pretreatment can ameliorate VD-induced recognition memory impairment and anxiety-like behavior in novel recognition memory and open-field tasks. Thus, improvement of recognition memory is a potential target for interventions against VD. There is currently a high interest in research about the role of exercise in reducing cognitive impairment and anxiety-like conditions. Previous studies demonstrated that exercise reverses recognition memory impairment and anxiety-like behavior in VD rats model40,41. Thus, our results showed that 4 weeks of treadmill exercise pretreatment ameliorates recognition memory impairment and anxiety-like behavior caused by two-vessel occlusion to some extent in VD rats. Although we have demonstrated that treadmill exercise pretreatment is involved in alleviating recognition memory impairment and anxiety-like behaviors, the potential mechanisms of the positive effects of treadmill exercise pretreatment on recognition memory in VD rat models needs to be further investigated.
Next, the potential underlying mechanism through which treadmill exercise induced an improvement of recognition memory in the VD rat model was examined. The morphological basis of memory is structural synaptic plasticity, which also plays important roles in learning and memory formation42,43. Learning and memory requires neural adaptations and dendritic spine participation, which are thought to be mediated by activity-dependent structural synaptic plasticity44. Therefore, the neuronal impairment will likely cause changes in structural synaptic plasticity. Previous studies demonstrated via Nissl staining that VD rats displayed significantly neuron structural changes in the hippocampal45. The immunostaining results in our study demonstrated that treadmill exercise pretreatment can ameliorate VD-induced the decrease of neuron density in the medial prefrontal cortex region of VD rats, as determined by NeuN immunostaining. In addition to NeuN (which is considered a nuclear marker), we also detected fluorescence levels higher than the threshold in the neurons from the rat brains46. Thus, we concluded that there was a significant decrease in medial prefrontal cortex neurons in the VD rats, which were restored by treadmill exercise pretreatment. the results by golgi staining also demonstrated that 4 weeks of treadmill exercise pretreatment can ameliorate VD-induced impairment of dendritic spines in the medial prefrontal cortex. As small protrusions that form on dendrites, structural changes in dendritic spines of neurons are crucial in the implementation of synaptic plasticity, and for memory storage and neuronal connectivity21. Synaptic plasticity is influenced by the spatial pattern of synaptic inputs to the dendritic spines47. In vivo studies have shown that dendritic spines of the cortex in the VD mice were also impaired48. Consistent with the previously mentioned study, we also demonstrated that there was a significant reduction in the density of dendritic spines in the medial prefrontal cortex in the VD group when compared to the Con group. Enright et al.49 illustrated our result that hypoxic ischemic induced dendritic impaired in the brain is associated with cognitive impairment in VD rat model. Our study further demonstrated that the decrease in medial prefrontal cortex dendritic spines in the VD rats was ameliorated by treadmill exercise pretreatment. This phenomenon can be explained by the fact that exercise can exert neuroprotective effect by increasing cellular proliferation, dendritic complexity, and spine density in normal states50, and even to counter VD-induced neuron damage and dendritic injury51. In conclusion, treadmill exercise pretreatment can ameliorate VD-induced impairment of neuron and dendritic spines in the medial prefrontal cortex.
In our study, the results of transmission electron microscopy further demonstrated that there was a significant reduction in the density of synapses in the medial prefrontal cortex of VD rats, while treadmill exercise pretreatment increased the density of synapses in the medial prefrontal cortex of VD rats. In vivo imaging further demonstrated that VD rats showed remarkable damage of the synapse ultrastructure, such as the synaptic active zone, PSD thickness, and synaptic vesicles were significantly decreased52,53. The presynaptic processes of structural synaptic plasticity is reflected by the synaptic active zone, while PSD thickness reflects the postsynaptic neuronal mechanisms of structural synaptic plasticity54,55. In addition, synaptic vesicles mainly participated in information transmission and memory formation by releasing neurotransmitters56. Yang et al.57 demonstrated a significant reduction in the number of synaptic vesicles in VD rats. We further found that after VD, the dopamine and 5-HT levels in the medial prefrontal cortex are significantly decreased. However, these alterations were reversed by treadmill exercise pretreatment. Several body functions including reward, learning and executive functions are mediated by dopamine. Similarly, 5-HT is regarded as a modulator in various central nervous system functions, such as emotion, behavior, and cognition58,59. It has been practically proven that moderate exercise can increase neurotransmitters in both human60 and animal studies37,61, which are closely associated with learning memory and cognition. The studies performed by Li et al.62 indicated that voluntary wheel running induced switching from acetylcholine to gamma-aminobutyric acid expression in neurons in the caudal pedunculopontine nucleus. The beneficial effect of voluntary wheel running on motor skill learning could be blocked if viral vectors are used to override transmitter switching. Others studies have shown that cholinergic deficits and altered glutamatergic signaling that occur in VD patients may worsen cognitive impairment63,64. Our results supported this hypothesis as treadmill exercise pretreatment was found to alleviate VD-induced reduction in the number of synapses, and improve dopamine and 5-HT levels in the medial prefrontal cortex.
In summary, impaired structural synaptic plasticity in the rat medial prefrontal cortex is accompanied by recognition memory impairment and anxiety-like behavior under VD conditions. Such deficits can be ameliorated by treadmill exercise pretreatment. But this study still has some limitations, such as the behavioral results should be replicated using larger samples.
Conclusion
Treadmill exercise pretreatment can ameliorate structural synaptic plasticity impairments of medial prefrontal cortex in VD rat and improved recognition memory.
Data availability
The datasets are available from the corresponding author on reasonable request.
References
Graff-Radford, J. Vascular cognitive impairment. Continuum (Minneap Minn) 25(1), 147–164 (2019).
Van Loenhoud, A. C. et al. High occurrence of transportation and logistics occupations among vascular dementia patients: An observational study. Alzheimers Res. Ther. 11(1), 112 (2019).
Lecordier, S., Manrique-Castano, D., El Moghrabi, Y. & ElAli, A. Neurovascular alterations in vascular dementia: Emphasis on risk factors. Front. Aging Neurosci. 13, 727590 (2021).
Yasuda, R., Hayashi, Y. & Hell, J. W. CaMKII: A central molecular organizer of synaptic plasticity, learning and memory. Nat. Rev. Neurosci. 23(11), 666–682 (2022).
Deng, Y. et al. Higher circulating trimethylamine N-oxide aggravates cognitive impairment probably via downregulating hippocampal SIRT1 in vascular dementia rats. Cells 11(22), 3650 (2022).
Impellizzeri, D. et al. Açai berry mitigates vascular dementia-induced neuropathological alterations modulating Nrf-2/Beclin1 pathways. Cells 11(16), 2616 (2022).
Wang, J. et al. Bone marrow mononuclear cell transplantation promotes therapeutic angiogenesis via upregulation of the VEGF-VEGFR2 signaling pathway in a rat model of vascular dementia. Behav. Brain Res. 15(265), 171–180 (2014).
Culmone, L. et al. Treatment with an angiopoietin-1 mimetic peptide improves cognitive outcome in rats with vascular dementia. Front. Cell. Neurosci. 6(16), 869710 (2022).
Li, H. et al. Acupuncture reversed hippocampal mitochondrial dysfunction in vascular dementia rats. Neurochem. Int. 92, 35–42 (2016).
Korczyn, A. D., Vakhapova, V. & Grinberg, L. T. Vascular dementia. J. Neurol. Sci. 322(1–2), 2–10 (2012).
Yoo, J. Y. et al. Effect of yoga exercise on blood pressure, physical fitness, and blood variables in elderly with vascular dementia. Korean J. Growth Dev. 18(3), 217–226 (2010).
Protecting against vascular dementia with exercise. Johns Hopkins Med Lett Health After 50. 25(1), 8 (2013).
Aarsland, D., Sardahaee, F. S., Anderssen, S. & Ballard, C. Alzheimer’s Society Systematic Review group. Is physical activity a potential preventive factor for vascular dementia? A systematic review. Aging Ment. Health 14(4), 386–395 (2010).
Gallaway, P. J. et al. Physical activity: A viable way to reduce the risks of mild cognitive impairment, Alzheimer’s disease, and vascular dementia in older adults. Brain Sci. 7(2), 22 (2017).
Kuhne, L. A. et al. The effects of acute cardiovascular exercise on memory and its associations with exercise-induced increases in neurotrophic factors. Front. Aging Neurosci. 8(13), 750401 (2021).
Okamoto, M. et al. High-intensity intermittent training enhances spatial memory and hippocampal neurogenesis associated with BDNF signaling in rats. Cerebral Cortex 31(9), 4386–4397. https://doi.org/10.1093/cercor/bhab093 (2021).
Diederich, K. et al. Effects of different exercise strategies and intensities on memory performance and neurogenesis. Front. Behav. Neurosci. 16(11), 47 (2017).
Yu, H., Zhang, C., Xia, J. & Xu, B. Treadmill exercise ameliorates adult hippocampal neurogenesis possibly by adjusting the APP proteolytic pathway in APP/PS1 transgenic mice. Int. J. Mol. Sci. 22(17), 9570 (2021).
Wang, P. et al. Potential involvement of adiponectin signaling in regulating physical exercise-elicited hippocampal neurogenesis and dendritic morphology in stressed mice. Front. Cell. Neurosci. 3(14), 189 (2020).
Choi, D. H., Lee, K. H. & Lee, J. Effect of exercise-induced neurogenesis on cognitive function deficit in a rat model of vascular dementia. Mol. Med. Rep. 13(4), 2981–2990 (2016).
Ji, B. & Skup, M. Roles of palmitoylation in structural long-term synaptic plasticity. Mol. Brain 14(1), 8. https://doi.org/10.1186/s13041-020-00717-y (2021).
Patten, A. R. et al. Benefits of exercise on structural and functional plasticity in the rodent hippocampus of different disease models. Brain Plast. 1(1), 97–127. https://doi.org/10.3233/BPL-150016 (2015).
Guo, L., Yang, X., Zhang, Y., Xu, X. & Li, Y. Effect of exercise on cognitive function and synaptic plasticity in Alzheimer’s disease models: A systematic review and meta-analysis. Front. Aging Neurosci. 14, 1077732 (2023).
Ivy, A. S. et al. A unique mouse model of early life exercise enables hippocampal memory and synaptic plasticity. Sci. Rep. 10(1), 9174 (2020).
Sheynikhovich, D., Otani, S., Bai, J. & Arleo, A. Long-term memory, synaptic plasticity and dopamine in rodent medial prefrontal cortex: Role in executive functions. Front. Behav. Neurosci. 16, 1068271. https://doi.org/10.3389/fnbeh.2022.1068271 (2023).
Keller, D., Erö, C. & Markram, H. Cell densities in the mouse brain: A systematic review. Front. Neuroanat. 12, 83 (2018).
Zhang, H., Zou, Y. & Lei, H. Regional metabolic differences in rat prefrontal cortex measured with in vivo 1H-MRS correlate with regional histochemical differences. NMR Biomed. 32(1), e4024 (2019).
Liu, K. et al. Dynamic regional changes of extracellular ascorbic acid during global cerebral ischemia: Studied with in vivo microdialysis coupled with on-line electrochemical detection. Brain Res. 1253, 161–168 (2009).
Wang, R. et al. Impacts of exercise intervention on various diseases in rats. J. Sport Health Sci. 9(3), 211–227 (2020).
Zhang, L. et al. Treadmill exercise improve recognition memory by TREM2 pathway to inhibit hippocampal microglial activation and neuroinflammation in Alzheimer’s disease model. Physiol. Behav. 251, 113820 (2022).
Lu, Y. et al. Molar loss further exacerbates 2-VO-induced cognitive impairment associated with the activation of p38MAPK/NFκB pathway. Front. Aging Neurosci. 3(14), 930016 (2022).
Zhang, Y. M., Wu, W., Ma, W., Wang, F. & Yuan, J. Effect of glycosides of Cistanche on the expression of mitochondrial precursor protein and keratin type II cytoskeletal 6A in a rat model of vascular dementia. Neural Regen. Res. 12(7), 1152–1158 (2017).
Knight, P. et al. Sex differences in the elevated plus-maze test and large open field test in adult Wistar rats. Pharmacol. Biochem. Behav. 204, 173168 (2021).
Balu, D. et al. Behavioral and physiological characterization of PKC-dependent phosphorylation in the Grin2a∆PKC mouse. Brain Res. 1(1646), 315–326 (2016).
Kim, S. J. et al. Retinoic acid-induced protein 14 controls dendritic spine dynamics associated with depressive-like behaviors. Elife 25(11), e77755 (2022).
Apóstolo, N. et al. Synapse type-specific proteomic dissection identifies IgSF8 as a hippocampal CA3 microcircuit organizer. Nat. Commun. 11(1), 5171 (2020).
Bastioli, G. et al. Voluntary exercise boosts striatal dopamine release: Evidence for the necessary and sufficient role of BDNF. J. Neurosci. 42(23), 4725–4736 (2022).
Schwertner, E. et al. Behavioral and psychological symptoms of dementia in different dementia disorders: A large-scale study of 10,000 individuals. J. Alzheimers Dis. 87(3), 1307–1318 (2022).
Liu, J. et al. Neuroprotective effects of clostridium butyricum against vascular dementia in mice via metabolic butyrate. Biomed. Res. Int. 2015, 412946 (2015).
Fan, Y., Kong, X., Liu, K. & Wu, H. Exercise on striatal dopamine level and anxiety-like behavior in male rats after 2-VO cerebral ischemia. Behav. Neurol. 13(2022), 2243717 (2022).
Ohtomo, R. et al. Treadmill exercise during cerebral hypoperfusion has only limited effects on cognitive function in middle-aged subcortical ischemic vascular dementia mice. Front. Aging Neurosci. https://doi.org/10.3389/fnagi.2021.756537 (2021).
Basu, S. & Lamprecht, R. The role of actin cytoskeleton in dendritic spines in the maintenance of long-term memory. Front. Mol. Neurosci. 11, 143 (2018).
Segal, M. Dendritic spines and long-term plasticity. Nat. Rev. Neurosci. 6(4), 277–284 (2005).
Milstein, A. D. et al. Bidirectional synaptic plasticity rapidly modifies hippocampal representations. Elife 9(10), e73046 (2021).
Wei, C., Xu, X., Zhu, H., Zhang, X. & Gao, Z. Promotive role of microRNA-150 in hippocampal neurons apoptosis in vascular dementia model rats. Mol. Med. Rep. 23(4), 257 (2021).
Hahn, Y. K., Masvekar, R. R., Xu, R., Hauser, K. F. & Knapp, P. E. Chronic HIV-1 Tat and HIV reduce Rbfox3/NeuN: Evidence for sex-related effects. Curr HIV Res. 13(1), 10–20 (2015).
Dorman, D. B. & Blackwell, K. T. Synaptic plasticity is predicted by spatiotemporal firing rate patterns and robust to in vivo-like variability. Biomolecules 12(10), 1402 (2022).
Ma, X. et al. Exosomal miR-132-3p from mesenchymal stromal cells improves synaptic dysfunction and cognitive decline in vascular dementia. Stem Cell Res. Ther. 13(1), 315 (2022).
Enright, L. E., Zhang, S. & Murphy, T. H. Fine mapping of the spatial relationship between acute ischemia and dendritic structure indicates selective vulnerability of layer V neuron dendritic tufts within single neurons in vivo. J. Cereb. Blood Flow Metab. 27(6), 1185–1200 (2007).
Eadie, B. D., Redila, V. A. & Christie, B. R. Voluntary exercise alters the cytoarchitecture of the adult dentate gyrus by increasing cellular proliferation, dendritic complexity, and spine density. J. Comp. Neurol. 486(1), 39–47 (2005).
Biose, I. J. et al. The effects of physical activity on experimental models of vascular dementia: A systematic review and meta-analysis. Ann. Neurosci. https://doi.org/10.1177/09727531231192759 (2024).
Bailey, C. H., Kandel, E. R. & Harris, K. M. Structural components of synaptic plasticity and memory consolidation. Cold Spring Harb. Perspect. Biol. 7(7), a021758 (2015).
Huang, Y. et al. Effects of imperatorin on apoptosis and synaptic plasticity in vascular dementia rats. Sci. Rep. 11(1), 8590 (2021).
Mu, L. et al. Treadmill exercise prevents decline in spatial learning and memory in 3×Tg-AD mice through enhancement of structural synaptic plasticity of the hippocampus and prefrontal cortex. Cells 11(2), 244 (2022).
Briz, V. et al. The non-coding RNA BC1 regulates experience-dependent structural plasticity and learning. Nat. Commun. 8(1), 293 (2017).
Eshra, A., Schmidt, H., Eilers, J. & Hallermann, S. Calcium dependence of neurotransmitter release at a high fidelity synapse. Elife. 6(10), e70408 (2021).
Yang, H. Y. et al. Effects of repetitive transcranial magnetic stimulation on synaptic plasticity and apoptosis in vascular dementia rats. Behav. Brain Res. 281, 149–155 (2014).
De Jong, J. W. et al. A neural circuit mechanism for encoding aversive stimuli in the mesolimbic dopamine system. Neuron 101(1), 133-151.e7 (2019).
Carhart-Harris, R. L. & Nutt, D. J. Serotonin and brain function: A tale of two receptors. J. Psychopharmacol. 31(9), 1091–1120 (2017).
LIFE Study Group et al. Dopamine-related genotypes and physical activity change during an intervention: The lifestyle interventions and independence for elders study. J. Am. Geriatr. Soc. 66(6), 1172–1179 (2018).
Tai, F. et al. Treadmill exercise ameliorates chronic REM sleep deprivation-induced anxiety-like behavior and cognitive impairment in C57BL/6 J mice. Brain Res. Bull. 164(Suppl 3), 198 (2020).
Li, H. Q. & Spitzer, N. C. Exercise enhances motor skill learning by neurotransmitter switching in the adult midbrain. Nat. Commun. 11(1), 2195 (2020).
Zhang, X. Q., Li, L., Huo, J. T., Cheng, M. & Li, L. H. Effects of repetitive transcranial magnetic stimulation on cognitive function and cholinergic activity in the rat hippocampus after vascular dementia. Neural Regen Res. 13(8), 1384–1389 (2018).
Dimatelis, J. J. et al. Impaired energy metabolism and disturbed dopamine and glutamate signalling in the striatum and prefrontal cortex of the spontaneously hypertensive rat model of attention-deficit hyperactivity disorder. J. Mol. Neurosci. 56, 696–707 (2015).
Author information
Authors and Affiliations
Contributions
Conceptualization: L.Z. and Y.C.; Methodology: Y.F.; Formal analysis: L.Z.; Investigation: Y.C.; Resources: Y.F.; Data Curation: L.S.; Writing—Original Draft: L.Z.; Writing—Review & Editing: L.S.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, L., Chen, Y., Fan, Y. et al. Treadmill exercise pretreatment ameliorated structural synaptic plasticity impairments of medial prefrontal cortex in vascular dementia rat and improved recognition memory. Sci Rep 14, 7116 (2024). https://doi.org/10.1038/s41598-024-57080-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-57080-4
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
