
Abstract
Dazomet is a dry powder formulation that releases toxic gas containing methyl isothiocyanate, which controls soil-borne pests and weeds, improving crop yields when applied to moist soils. To explore the efficacy of dazomet fumigation in the cultivation of the perennial herb Codonopsis pilosula, four typical cultivars (G1, G2, W1 and TCK) in Gansu Province were selected for seedling cultivation after soil fumigation (F) by dazomet, and non-fumigated soil was used as a control (CK). The experiments took 2 years to complete. The functional diversity of the soil enzymes and microorganisms, seedling emergence and physiological characteristics, and the quality and yield of Codonopsis seedlings and Radix were assessed. The results showed that the seed emergence rate, seedling re-green rate and several antioxidant enzymatic activities improved in the treatments involving soil fumigation with dazomet, and membrane lipid peroxidation in the seedlings decreased. On average, compared with those of the respective controls, the root viability and yield of the seedlings of the tested cultivars also increased by 34.87% and 42.4%, respectively, and the incidence of root rot in the seedlings was reduced by 83.9%, compared with their respective controls. After harvest, the yield increased by 23.9%, the incidence of root rot decreased by 61.3%, increase in yield and a 61.3% reduction in incidence, and the medicinal materials were determined to be safe and residue-free. The effects of fumigation were cultivar-specific and were especially prominent in G2. Therefore, soil fumigation with dazomet could improve the quality and productivity of Codonopsis pilosula seedlings. Taken together, these findings suggest that when herbs are bred by seedling transplantation, especially cultivars of good quality but poor resistance or species with rare germplasm resources, soil fumigation provides a way to improve cultivation effectiveness and, more importantly, ensures the probability of successfully breeding the species.
Similar content being viewed by others


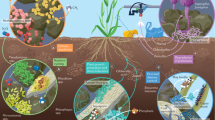
Introduction
Codonopsis pilosula is a traditional herbal plant that is widely used in Asian countries, mainly planted in China, Japan, and Korea1,2, and its dry roots are used as medicine (Codonopsis Radix, also Dangshen)3. The use of this herb has been extensively recorded in Traditional Chinese medicine (CHM) texts, such as “Ben Cao Cong Xin”, “Supplements to Compendium of Materia Medica” and “Ben Cao Qiu Zhen”4. Furthermore, more than 110 CHM preparations containing Codonopsis Radix or its extracts are listed in the Chinese Pharmacopoeia3. Codonopsis Radix is also widely used as a food additive in wine, soup, porridge, etc.5,6. Moreover, these agents have a wide range of pharmacological effects, such as strengthening the spleen, benefiting the lungs, nourishing the blood, and promoting the production of body fluids3,5,7,8. People in countries such as China, Japan, North Korea, South Korea, and the United States also use it as a food ingredient9,10. Owing to these health benefits, the demand for Codonopsis Radix has increased globally, stimulating the large-scale cultivation of C. pilosula, especially in Gansu Province11,12. Gansu Province has more than 90% of the C. pilosula plantation area13 in China and was recognized as “The Hometown of C. pilosula in China” by the Chinese Special Products Township Committee in 2001. Because the medicinal quality of these plants is closely related to their origin, the plantation area of C. pilosula in Gansu Province is limited. Continuous planting has caused a decrease in plant quality and yield. Thus, studies are urgently needed to overcome these continuous cropping obstacles and improve medicinal quality and productivity.
It has been reported that exogenous soil, intercropping14, crop rotation15, the application of organic materials16, and soil fumigation17 can reduce soil-borne diseases, pests, and weeds. Soil fumigation is the most effective and stable method for protecting crops from soil-borne diseases, nematodes, and weeds during continuous cropping18,19. Dazomet (DZ) is a highly effective microbicide for controlling fungi, weeds, and subterranean pests20,21,22; it induces a carbonylation reaction at nucleophilic sites (such as amino, hydroxyl, and thiol groups). Most studies of DZ have been performed on strawberry, tomato, flower, ginger, cucumber, and other high-value crops23,24,25,26. It can also be applied in integrated pest management programs for ginger to control ginger bacterial wilt in China24, and it has successfully replaced methyl bromide, which damages the ozone layer27.
Codonopsis pilosula is a perennial herbaceous plant in the Campanulaceae family that mainly propagates by seed reproduction28. The yield and quality of Codonopsis Radix depend on the C. pilosula seed and seedling quality. High-quality Codonopsis seedlings are fundamental for improving the Codonopsis Radix yield and quality29,30. However, several problems have limited the quality and production performance of C. pilosula during cultivation. First, the seeds of C. pilosula are tiny and the weight of 1000 seeds is only approximately 0.30 g, which limits the emergence rate. Second, the occurrence of various weeds strongly affects the seedling growth and development. Third, in the native producing areas, growers have implemented continuous cropping instead of crop rotation to pursue high profits, which can lead to soil-borne diseases, pests, and weeds, strongly affecting Codonopsis seedling quality. Although many studies have been performed on evaluating the effects of sowing time, sowing density, seed treatment, cover materials and sowing methods31, these were aimed at increasing production and thus failed to solve problems related to quality. Therefore, the present study was designed to improve Codonopsis seedling quality through soil improvement. Briefly, four typical cultivars of C. pilosula in Gansu Province were selected for seedling cultivation after soil fumigation with DZ. The seed emergence rate, root viability, antioxidant enzymatic activity, transplant re-green rate, and other indicators of the plant seedlings in the fumigated and non-fumigated plots were recorded. Additionally, the quality and yield of the seedlings and Radix were measured, after which the safety of the medicine was evaluated. The present study was aimed at providing systematic findings to promote the sustainable and efficient cultivation of C. pilosula while also protecting the environment.
Materials and methods
Experimental field and test cultivars
The seedling cultivation experiment was conducted in Lichuan (E 104° 01ʹ, N 33° 46 ʹ) in northern Tanchang County, Longnan city, Gansu Province, China, which is the main production area of C. pilosula and has a mild and humid climate. The previous crop in the experimental field was Codonopsis pilosula (continuous cropping). To eliminate test errors, four cultivars, “Gandang No. 1” (G1) and “Gandang No. 2” (G2), bred by the Eco-cultivation and Breeding Group of Professors Yuan Chen and Fengxia Guo at Gansu Agricultural University, the local representative cultivar “Weidang No. 1” (W1) and the Tanchang traditional cultivar (TCK), were used synchronously in this experiment. After Codonopsis seedlings were harvested, transplanted in a Scutellaria baicalensis stubble field without any soil treatment in Pangjiacha (E 107° 87ʹ, N 44° 44ʹ) in Fuxing, Longxi County, Gansu Province.
Experimental design
A two-factor (soil pretreatment and cultivar) scheme was used in the seedling cultivation experiment, in which the soil pretreatment factor included two levels of soil fumigation (F) by 98% dazomet and non-fumigation (CK). Dazomet particles (98%) were obtained from Jiangsu Nantong Chemical Co., Ltd.. Based on preliminary results from our laboratory pot experiments and the findings of other researchers with respect to other crops20,25,32, the applied concentration was set to 45 g m−2, and the duration of fumigation was 25 days. A paired design for soil fumigation and non-fumigation treatments was used for the four cultivars in this experiment, including eight experimental plots of 30 m2 (6 m × 5 m) in each block. To facilitate soil fumigation and field management, the experimental field was divided into two adjacent parallel long blocks from north to south, separated by ridges 0.80 m wide and 0.50 m high. According to the NY/T 3129-2017 technical specification for soil fumigation with dazomet33, the soil was fumigated in the northern long block with 45.0 g m−2 dazomet on Mar. 15, 2019, and the non-fumigation one in the southern region was used as the control. At the end of fumigation, soil fertility was measured. The results were consistent (nonsignificant difference according to t-test, P > 0.05) between the two blocks (local production fields Table 1). Then, each long block was divided into four plots separated by a 0.5 m ridge, in which the four cultivars were randomly sown to cultivate seedlings. Other than the different treatments applied, the field management and operation procedures used were the same as those used for local production fields.
Seed sowing
Seeds of the four cultivars of C. pilosula were sown in the above plots of the F and CK blocks on 7 May 2019. The seeds were uniformly spread and were distributed by a single person, with a seed density of 3.0 g m−2 during sowing. After sowing, the block was covered with a black shading net (60% shading rate). The seedlings were ordinally sampled at three locations in each plot at three stages of seedling cultivation (18 Aug., 22 Sep., and 20 Oct.) and were subsequently transported to the laboratory. The sampled seedlings were washed briefly in running water to remove soil residues. Surface moisture was removed with absorbent paper, and the samples were subsequently stored at -80 °C for determination of physiological indicators.
Determination of soil indicators
The soils were sampled by a multipoint method with a soil auger before sowing and at the middle and late stages of seedling growth and were mixed as composite soil samples of the 0–10 cm and 10–20 cm layers in each block. The samples were subsequently placed in a tagged aseptic self-sealing bag and transported to the laboratory to be stored in a refrigerator at − 80 °C for subsequent determination of soil microorganisms, functional diversity of the soil communities and enzymatic activity.
Soil enzymatic activity was determined according to methods by Guan et al.34. The quantity of culturable soil microbes was determined using a dilution plate coating method. Soil samples were accurately weighed to 10.0 g and dissolved in 90 mL of dd-water with shaking and mixing; after the samples were left undisturbed, the supernatant was aspirated and diluted 10–3, 10–4, 10–5 times. The samples were subsequently coated with a premixed fungal, actinomycetes, and bacterial media, sealed with a sealing film and cultured in an incubator at 28 °C. Microbial colonies were counted when they appeared on the medium until the colonies grew into blurred patches. The microorganisms were expressed as clone numbers per gram of sample.
where M is the microbial colony, D is the dilution times, m is the weight of the fresh soil sample (g) and R is the soil relative water content (%).
The functional diversity of the soil microbial communities was determined according to Zak et al.35. After incubation, the data on a Biolog microplate reader were read every 24 h (1 day) for 8 readings (8 days). The metabolic capacity of soil microorganisms to metabolize 31 carbon sources was reflected by the average rate of colour change (AWCD) on Biolog microtiter plates.
where Ci is the optical density at 590 nm for each reaction hole, R is the optical density values of ecological board control holes (dd-water) and holes with a Ci-R less than 0 are all noted as 0 in the calculations.
The culture samples obtained after 5 days of vigorous microbial growth and metabolism were selected for functional diversity analysis of microbial communities. The numbers of coloured holes were used to express the intensity of carbon source utilization, and the Shannon–Wiener diversity index (Hʹ), Evenness index (E), Simpson diversity index (D), and McIntosh index (U) were calculated36,37. The distributions of carbon sources in the Biolog EcoPlate are shown in Table S1.
where Pi is the relative absorbance value of the i-th hole/sum of all absorbances and S is the absorbance value for 31 carbon sources that can be utilized by the microbes.
Emergence rate of Codonopsis seedlings
Three 400 cm2 (20 cm × 20 cm) sample quadrants in the centre of each plot were designated for the investigation of the number of plants that emerged per m2 (Qs). Finally, the emergence rate (Er) was calculated according to the seeds sown per m2 (m2, g) and 1000-seed weight (m1) using the following equation:
Root viability of Codonopsis seedlings
The root viability of Codonopsis seedlings was determined according to a slightly modified 2,3,5-triphenyl tetrazolium chloride (TTC) reduction method from Monika Dalal and Renu Khanna-Chopra38. Briefly, 0.05 g of detached roots were incubated in 3 mL of 0.08% TTC in phosphate buffer (pH 7.5) for 24 h in the dark at 18 °C. Then, the samples were collected and washed with distilled water 3 times. The reduced TTC in fresh roots was extracted with 5 mL of 95% ethanol at 60 °C in the dark for 10 min, and the reduction in TTC, which represents root viability, was determined by determining the absorbance at 485 nm (UV 2450/Vis spectrometer).
Assays of lipid peroxidation
Lipid peroxidation was measured following treatment and after 5 h by using malondialdehyde (MDA) equivalents39. The automatic rate (AR) was calculated as the average increase in the MDA concentration per hour40.
Assays of the antioxidant enzymatic activity
The antioxidant enzymatic activity was measured at 4 °C. The samples were homogenized in liquid nitrogen with 5 mL of extraction buffer containing 0.2 mM ethylenediaminetetraacetic (EDTA) and 2% polyvinylpyrrolidone (PVP) in 25 mM PBS (pH 7.8). The homogenate was centrifuged at 15,000 r min−1 for 20 min. The supernatant was subjected to antioxidant enzyme analysis41,42,43.
The activity of SOD was expressed as U g−1, and the CAT and POD activities were expressed as U g−1 min−1 according to Niu et al.43.
Re-green rate of transplanted C. pilosula
After transplanting, the dynamics of the seedlings that re-greened in each plot were periodically observed and recorded, and the re-green rate was calculated as follows:
Determination of the quality and yield of Codonopsis seedlings and Codonopsis Radix
After harvesting on 15 Mar. 2020, the yield and incidence of disease in Codonopsis seedlings in each plot were measured. When 2-year-old C. pilosula was harvested on 10 Nov. 2020, the yield and incidence of root rot in each plot were measured. A total of 20 plants were randomly sampled from each plot to determine the quality of the roots, such as length, taproot length, taproot diameter, fresh weight per root, and lateral root number. After drying in the shade, the dry weight of a single root was measured, and the drying rate was calculated based on the fresh and dried weight.
Root length and taproot length were measured with a measuring tape (cm, 1/10); the taproot diameter was measured with a Vernier caliper (mm, 1/100); and the fresh and dry weight of a single roots was measured with an electronic scale (g, 1/100).
Subsequently, the water content, total ash content, acid-insoluble ash content, and extract content were measured according to the methods of Chinese Pharmacopoeia3. The polysaccharide content was measured via the phenol sulfuric acid method in SN/T 4260-201544. Lobetyolin was measured using the assay of Yang et al.45 and Gao et al.46.
Evaluation of Codonopsis Radix safety
Residues of methyl isothiocyanate, a metabolite of DZ, were analysed by GC‒MS47. Briefly, 10.0 g of a 2-year-old C. pilosula root sample was accurately weighed and mixed with 10 mL of ethyl acetate in a 50 mL centrifuge tube. After vortexing for 2 min, 4.0 g of NaCl was added, and the mixture was shaken well. After centrifugation at 4000 r min−1 for 5 min, 1 mL of the supernatant was taken and passed to a 0.22 μm microporous membrane for GC‒MS analysis with an OB-1701 MS capillary column (60 m × 250 μm × 0.25 μm). The analysis conditions consisted of a He carrier gas, 1 mL min−1 column flow, 1 μL injection volume, no diversion, an injection port temperature of 150 °C, and a gas mass interface temperature at of 180 °C. When the column temperature was increased, the initial temperature was set to 50 °C for 4 min and increased to 150 °C at 20 °C min−1 over 2 min. The retention time of the peak for the target compound was 8.9 min.
Statistical analysis
Analyses of variance (ANOVAs) and principal component analysis (PCA) were conducted using SPSS 22.0 software (USA), and the least significant difference (LSD) method at P = 0.05 (LSD 0.05) was used for multiple comparisons.
A comprehensive evaluation was conducted according to the methods of Jin et al.48. First, based on factor analysis, the principal component values of each indicator whose initial characteristic root was greater than 1 were extracted, and then, the weight (Wj) and positive and negative membership function values were calculated according to the relevant properties. Finally, the comprehensive index (CI) was estimated.
where Cl,j represents the l-th principal component of the j-th indicator, VPl is the percentage of the variance of the first principal component, Wj is the weight value of the j-th indicator, i is different treatment, j is the measurement indicator, R(Xij) is the membership function value of indicator j under i treatment, RR(Xij) represents the value of anti membership function of i processing j indicator, Xij is the average observation value of indicator j of i treatment, Xjmin is the minimum value of j indicator in all treatments, Xjmax is the maximum value of j indicator in all treatments, CIj is the cumulative composite indicator of the i-th indicator. CIj is the cumulative composite indicator of the i-th processed j indicator.
Results
Effect of DZ soil fumigation on soil enzymatic activity
Soil enzymes are involved in plant growth activities and play a prominent role in soil nutrient cycling and plant nutrient supply. Soil microorganisms and plant root secretions are significant sources of soil enzymes. As shown in Fig. 1, soil fumigation significantly decreased soil urease, protease, and invertase activity but not catalase activity before sowing. With the growth of Codonopsis seedlings, soil urease, protease, and cellulase activity began to increase, and there were no significant changes compared with those in the CK treatment. However, after soil fumigation, the soil catalase activity was greater than that in the CK treatment.

Effect of DZ soil fumigation on soil enzymatic activity. F means soil fumigation, and CK means soil non-fumigation. The data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05.
Effect of DZ fumigation on the number of culturable soil microorganisms
Culturable soil microorganisms play a crucial role in the decomposition, metabolism, and other processes of substances in soils. Throughout the reproductive period of Codonopsis seedlings, the number of colonies of culturable microbes in the soil increased in the order bacteria > actinomycetes > fungi, and actinomycetes were most affected by soil fumigation but recovered more easily than fungi and bacteria (Fig. 2). The number of culturable soil microorganisms in the 10–20 cm soil layer was lower than that in the 0–10 cm layer. Compared with that in the CK treatment, the number of fungi in the fumigated block decreased by 95.72% on 7 May, 83.53% on 12 Jul. and 70.19% on 22 Sep.; the number of bacteria decreased by 89.17% on 7 May, 68.55% on 12 Jul. and 51.38% on 22 Sep.; and the number of actinomycetes decreased by 92.35% on 7 May, 67.20% on 12 Jul., and 64.32% on 22 Sep..

Effect of DZ fumigation on the culturable soil microbial quantity in Codonopsis seedlings fields. F means soil fumigation, and CK means soil non-fumigation. The data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05.
Effect of DZ soil fumigation on the metabolic functional diversity of soil microbial communities
AWCD is a crucial indicator of the metabolism of soil microbial carbon sources and can reflect the ability of soil microbial communities to utilize carbon sources. The faster the AWCD value increases, the more the microbial community metabolizes the carbon source and vice versa. The AWCD values of each soil layer rapidly increased from 1 to 5 days, and after 5 days, the increase in AWCD values slowed. This indicates that the microbial community strongly metabolized carbon sources from 1 to 5 days, while after 5 days, the microbial community metabolism of carbon sources gradually weakened. Overall, soil microbial activity was weakest in the fumigation plot before sowing, where soil microbial activity was more strongly affected in the 0–10 cm soil layer than in the 10–20 cm layer (Fig. 3).

AWCD of soil microbial community under different treatments. F means soil fumigation, and CK means soil non-fumigation. The data in the figure is the X ± SD.
As shown in Fig. 4, soil fumigation reduced the microbial diversity index and Evenness index of each soil layer before sowing, which increased with the growth of Codonopsis seedlings. Among the indices, the McIntosh indices were all above 1, which also indicated that there were only a few microbial species in the communities. The Evenness index varied between 0 and 1.0, and the variation in the Evenness index was consistent with that of the Shannon–Wiener index.

Soil microbial diversity index of Codonopsis seedlings field with different treatments. F means soil fumigation, and CK means soil non-fumigation. The data in the figure is the X ± SD.
To visualize the effect of dazomet fumigation on the metabolic characteristics of soil microbial communities, principal component analysis was carried out in this study using AWCD values from 5 days of incubation (Fig. 5). Soil fumigation affected soil microbial carbon source utilization in the early and middle stages of Codonopsis seedlings development, the effects gradually disappeared in the later stages, and the differences were not significant.

PCA of carbon utilization of soil microbes under different dates of different treatments. F means soil fumigation, CK means soil non-fumigation, M—May, J—July, and S—September.
Effect of DZ soil fumigation on the emergence rate of Codonopsis seedlings
Soil fumigation had significant effects on the emergence rate of Codonopsis seedlings (F = 27.49, P < 0.05; Fig. 6) relative to non-fumigated soil. The average emergence rates of the four cultivars of C. pilosula in the fumigation plot (F) and the non-fumigated plot (CK) were 65.75% and 31.38%, respectively. The emergence rate of the F plot was 34.38% greater than that of the CK plot. The seedling emergence rate in W1 increased by 322.3% compared with that in the corresponding control, showing the most significant increase, and that in G1 increased by 22.2%, the smallest increase.

The emergence rate of Codonopsis seedlings. Data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05; *is a significant correlation at P < 0.05.
Effect of DZ soil fumigation on physiological characteristics of Codonopsis seedlings
As shown in Fig. 7, soil fumigation had a significant effect on the physiological characteristics of the roots of Codonopsis seedlings (P < 0.05). Soil fumigation had a significant effect on the viability of the roots of Codonopsis seedlings of each cultivar, and the effect increased gradually with the growing period. Root viability of the seedlings under soil fumigation was 34.87% greater than that under CK treatment during the three growing periods. The degree of membrane lipid peroxidation is indicated by the content of MDA. The MDA and AR contents in Codonopsis seedling roots decreased gradually with the growing period, and the MDA content in the F plot was significantly lower than that in the CK plot (P < 0.05). Compared with that in CK, the average MDA content in the F plot decreased by 19.07%, and the AR decreased by 18.28%. In August, the effects of fumigation on the MDA content in G1, G2, and W1 were significant, but those on the TCK content were not significant. In September and October, W1 was significantly affected. The results showed that soil fumigation could reduce the content of MDA in Codonopsis seedlings and thus reduce cell membrane lipid damage.

The physiological characteristics of Codonopsis seedlings. Data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05.
Moreover, the soil fumigation had a significant effect on the POD activity of Codonopsis seedlings. Compared with that in the CK plot, the POD activity increased by 336.83% in August (P < 0.05). Soil fumigation also had a significant effect on the SOD activity of Codonopsis seedlings (P < 0.05) in August. However, there was no significant difference in CAT activity in the different growth stages (P > 0.05).
Effect of DZ soil fumigation on the yield and root rot incidence rate of Codonopsis seedlings
As shown in Fig. 8, there was a significant difference in the yield of Codonopsis seedlings among the cultivars. The yield of the F plot was greater than that of CK. The yield of G2 in the F plot was the highest, at approximately 6182.00 kg hm−2. The yield of W1 was the lowest, at approximately 4554.53 kg hm−2. The yield of G1 in the CK plot was the highest, at approximately 429.90 kg hm−2. The yield of the WCK treatment was the lowest, at approximately 2750.80 kg hm−2. The yield in the F plot was 1543.23 kg hm−2 greater than that in the CK plot (42.4%), and the difference between them was significant (P < 0.05). Soil fumigation had the most significant effect on G2, with a yield increase of 2362.07 kg hm−2 (increase of 61.8%), while soil fumigation had the weakest effect on G1, with a yield increase of 646.60 kg hm−2 (increase of 15.4%). Moreover, soil fumigation significantly affected the incidence of root rot in Codonopsis seedlings. Compared with that in the CK treatment, the incidence of root rot in the F treatment decreased by 83.9%.

The yield and incidence rate of Codonopsis seedlings. Data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05; “*” is a significant correlation at P < 0.05.
Re-green rate and survival rate of C. pilosula after transplantation
After Codonopsis seedlings were transplanted and turned green, the individual phenological distributions of the different cultivars of C. pilosula changed to varying degrees (Fig. 9). Compared with that in the CK treatment, the re-green rate in the F treatment increased by 10.09%, and the survival rate increased by 1.62%. Among them, the re-green rate in G2 was significantly greater than that in W1 and TCK (P < 0.05).

The re-green rate and survival rate of C. pilosula. Data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05; “*” is a significant correlation at P < 0.05.
Effects of seedlings on the yield and quality of Codonopsis Radix
When different treatment Codonopsis seedlings were cultivated, the yield of Codonopsis Radix increased (P < 0.05), and root rot incidence of Codonopsis Radix decreased (Fig. 10). Compared with that of CK, the yield of Codonopsis Radix increased by 23.94%, and the root rot incidence decreased by 61.32%. There were also differences among the different cultivars of C. pilosula. For instance, the yields of G1, G2, W1, and TCK increased by 26.51%, 20.99%, 22.57%, and 26.68%, respectively, and the root rot incidence decreased by 91.06%, 67.43%, 35.44%, and 51.18%, respectively.

The yield and incidence of Codonopsis Radix. Data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05; “*” is a significant correlation at P < 0.05.
The differences in the external quality (Fig. 11) and internal component content (Fig. 12) of Codonopsis Radix were also determined. The transplanted Codonopsis seedlings cultivated after soil fumigation exhibited improvements in terms of root length, taproot root length, taproot root diameter, and single dry weight of Codonopsis Radix to varying degrees, and the most significant effect was observed in G2 (P < 0.05). However, this treatment had no significant effect on the drying rate or the number of lateral roots of Codonopsis Radix. Moreover, there was no significant effect on the internal component content of Codonopsis Radix, although the extract composition of the different cultivars was different.

The external quality of Codonopsis Radix. Data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05.

The internal component content of Codonopsis Radix. Data in the figure is the X ± SD. Different small letters mean a significant difference at P < 0.05.
Comprehensive factor analysis of different C. pilosula cultivars under different treatments
Based on principal component analysis of thirty-eight indicators for four cultivars of Codonopsis seedlings and Codonopsis Radix under the two treatments, the characteristic roots of the first six principal components were greater than 1. The contribution rates were 49.83%, 15.56%, 11.63%, 9.56%, 7.39%, and 3.47%, respectively. The cumulative contribution rate was 97.44%. Therefore, the characteristic roots and contribution rates of the first six principal components (Table S2) were extracted to calculate the weight values of each indicator (Table S3). According to the test indicator membership degree, weight value, and addition and multiplication of all the indicators of Codonopsis seedlings and Codonopsis Radix, the comprehensive evaluation indices were F > CK (Table S4), and in the CK treatment, the order was G1 > G2 > W1 > TCK, in F was G2 > G1 > W1 > TCK.
Safety evaluation of Codonopsis Radix
Codonopsis Radix is a traditional Chinese medicine used for medicine and food. Currently, agencies in China have set standards for the maximum residual content of methyl isothiocyanate allowable in food, but the threshold for the residual content of methyl isothiocyanate in food specified by the EU is 0.02 mg kg−1. In this study, the content of methyl isothiocyanate residues in Codonopsis Radix was determined. Methyl isothiocyanate was not detected in Codonopsis Radix (< 0.02 mg kg−1).
Discussion
Codonopsis seedling emergence rate and root viability can be enhanced by dazomet soil fumigation
Root systems are essential functional organs in plants that absorb nutrients and water, participate in the process of biosynthesis and transformation, and have a vital impact on the growth and yield of crops49,50. Soils are essential for material and energy exchange, and a soil’s microecological environment directly affects the growth and development of plants. Numerous studies have shown that the application of soil amendments is an effective way to improve the soil microecological environment and increase crop quality and yield51,52,53. In our study, we found that, during the pre-seedling period, DZ soil fumigation affected soil enzymes, the number of culturable soil microorganisms, and the functional diversity of the microbial community; however, all the microbial communities recovered to varying degrees during the post-seedling period. Thus, due to the altered soil environment, fumigation also had an impact on the emergence and root viability of Codonopsis seedlings. The emergence rate of Codonopsis seedlings increased by 34.38% and root viability increased by 38.42% compared with CK, among which the root viability of TCK was most responsive and G2 was least responsive to the environment. This difference may be due to the strong adaptability of TCK, a local cultivar, to continuous cropping, while G2 was more environmentally adaptable. Liu et al.19 studied the effect of dazomet on the growth of Pingyi sweet tea seedlings in pots and reported that dazomet increased the root viability of Pingyi sweet tea by 48.8%, which was similar to the findings of the present study.
Soil fumigation with DZ improved the environmental adaptability of Codonopsis seedlings
MDA is the final product of membrane lipid peroxidation. A high concentration of MDA has a toxic effect on plants, but a low concentration of MDA can induce enhanced stress tolerance41,42. The rate of tissue automatic oxidation (AR) is the rate of MDA production per unit of time and can directly reflect the degree of membrane lipid peroxidation41,42,54. Under continuous cropping conditions, the abundance of weeds55 and harmful bacteria56 in the soil increases, which might lead to stress during plant growth. Under adverse conditions, large quantities of reactive oxygen species such as hydrogen peroxide, superoxide anions, and hydroxyl radicals are produced in plants, leading to an increase in MDA content, which in turn affects plant growth57. To scavenge reactive oxygen species, plants activate an antioxidant defence enzyme system consisting of enzymes such as SOD, CAT, and POD58,59 and regulate dysregulated reactive oxygen species metabolism via protective enzyme activity in plants. Soil fumigation with dazomet can significantly reduce the number of weeds55 and the number of pathogenic bacteria60 in the soil. Although soil enzymatic activity and the functional diversity of the soil microbial community were reduced for a short duration, the microbial communities recovered to varying degrees during the later stages, and the Codonopsis seedlings were in a better soil environment with a lower MDA content and greater enzymatic activity. The high re-green rate and survival rate of Codonopsis seedlings further explained this difference. Therefore, it is evident that changes in the soil environment significantly improve the environmental adaptability of Codonopsis seedlings, allow for rapid regreening, and increase field productivity.
Soil fumigation with DZ improved the quality and yield of Codonopsis seedlings
The appearance of Codonopsis seedlings is directly related to the quality of the medicine and its clinical efficacy. The quality of Codonopsis seedlings is determined by the genetic composition of the C. pilosula seeds, the growth environment and cultivation measures; therefore, the quality of Codonopsis seedlings can be improved under suitable soil environments and cultivation conditions61. Studies have shown that the application of soil amendments could improve the soil environment and increase crop yield and quality62,63. In this study, when soil moisture conditions and the amount, duration, and mulching material of fumigants were strictly regulated when dazomet was applied, soil fumigation increased the yield of Codonopsis seedlings by 42.4% and reduced the morbidity rate by 84.48%. Similarly, compared with the CK treatment, the yield of Codonopsis Radix increased by 23.94%, and the root rot incidence decreased by 61.32%. The effect of dazomet fumigation on the quality and yield components of Codonopsis seedlings was mainly reflected as the indicators of root length, root thickness, single root weight, and the number of lateral roots. This was further corroborated by the appearance of the Codonopsis Radix, showing that the quality of the seedlings was directly related to the growth and development of the plants after transplanting and the yield and quality of the herbs harvested in the same year64. Considering the seedling traits, root rot incidence, and yield, fumigation had a significant effect on the quality and yield of all the C. pilosula cultivars. The most significant effect was seen in G2, with a 61.8% increase in yield, and the weakest effect was observed in G1, with a 15.4% increase in yield. In addition, soil amendments had little effect on the intrinsic quality of Codonopsis Radix, which is generally determined by genetic factors, and the intrinsic quality varies significantly among cultivars. In this study, there were no significant differences in the ash, leachate content, polysaccharide, or lobetyolin content of Codonopsis Radix between the treatments; however, the polysaccharide and lobetyolin content of Codonopsis Radix varied among cultivars. Moreover, we did not find methyl isothiocyanate residues in Codonopsis Radix, indicating that the safety of Codonopsis Radix was high after fumigation of the seedbed by dazomet.
Conclusions
Soil fumigation has been proven to be a critical technology for ensuring stable crop yields. The results of this study revealed that the application of dazomet could effectively improve the seeds’ emergence rate and seedling antioxidant enzymatic activity, and reduce membrane lipid peroxidation and the incidence of root rot. After transplant, the seedling re-green rate was increased, and the yield and appearance quality of Codonopsis Radix improved. These findings were closely related to those for C. pilosula, especially because of its high yield but poor resistance. Therefore, when herbs are bred that require seedling transplants, especially cultivars with good quality but poor resistance or species with rare germplasm resources, soil fumigation can be carried out before seedling, which not only improves yield but also, more importantly, ensures the probability of successful cultivation of the species.
Data availability
The data used to support the findings of this study are available from the corresponding author upon request.
References
Chen, L. & Huang, G. L. The antiviral activity of polysaccharides and their derivatives. Int. J. Biol. Macromol. 115, 77–82 (2018).
Liu, W., Lv, X., Huang, W. H., Yao, W. B. & Gao, X. D. Characterization and hypoglycemic effect of a neutral polysaccharide extracted from the residue of Codonopsis Pilosula. Carbohyd. Polym. 197, 215–226 (2018).
CP Committee. Edition of Chinese Pharmacopoeia 293–294 (Chemical Industry Press, 2020).
Chai, R. J. Textual research on the herbal history of Codonopsis pilosula. Chin. J. Integr. Med. Cardio-Cerebro. Dis. 6, 43–45 (1991).
Gao, S. M. et al. Traditional uses, phytochemistry, pharmacology and toxicology of codonopsis: A review. J. Ethnopharmacol. 20, 25 (2018).
Zou, Y. F. et al. Structural features and complement fixing activity of polysaccharides from Codonopsis pilosula nannf. Var. modesta L. T. Shen roots. Carbohyd. Polym. 113, 420–429 (2014).
He, J. Y., Zhu, S., Komatsu, K., Goda, Y. & Cai, S. Q. Genetic polymorphism of medicinally-used Codonopsis species in an internal transcribed spacer sequence of nuclear ribosomal DNA and its application to authenticate Codonopsis Radix. J. Nat. Mde-Tokyo. 68, 112–124 (2014).
Sun, Q. L. et al. Structural characterization of three polysaccharides from the roots of Codonopsis pilosula and their immunomodulatory effects on RAW264.7 macrophages. Int. J. Biol. Macromol. 130, 556–563 (2019).
Moon, K. G. et al. Effect of organic fertilizer application on growth characteristics and saponin content in Codonopsis lanceolata. Hortic. Environ. Biotech. 59, 125–130 (2018).
Boo, H. O. et al. Codonopsis lanceolata and Nelumbo nucifera Gaertn root extracts for functional food: Metabolic profiling by MS, FTIR and fluorescence and evaluation of cytotoxicity and anti-obesity properties on 3T3-L1 cell line. Eur. Food. Res. Technol. 243, 689–700 (2017).
Liu, D. J. et al. EDEM simulation study on the performance of a mechanized ditching device for Codonopsis planting. MDPI Agr. 12, 1238–1253 (2022).
Guo, H. B., Lu, B. R., Wu, Q. H., Chen, J. K. & Zhou, T. S. Abundant genetic diversity in cultivated codonopsis pilosula populations revealed by RAPD polymorphisms. Genet. Resour. Crop Evol. 54, 917–924 (2007).
Yang, J. et al. Establishment of in vitro culture system for Codonopsis pilosula transgenic hairy roots. 3 Biotech 10, 137–145 (2010).
Zhou, X. G., Yu, G. B. & Wu, F. Z. Effects of intercropping cucumber with onion or garlic on soil enzyme activities, microbial communities and cucumber yield. Eur. J. Soil Bio. 47, 279–287 (2011).
Sainju, U. M., Lenssen, A. W., Allen, B. L., Stevens, W. B. & Jabro, J. D. Soil total carbon and nitrogen and crop yields after eight years of tillage, crop rotation, and cultural practice. Heliyon 3, e00481–e00503 (2017).
Ling, N., Zhang, W. W., Tan, S. Y., Huang, Q. W. & Shen, Q. R. Effect of the nursery application of bioorganic fertilizer on spatial distribution of Fusarium oxysporum f. sp. niveum and its antagonistic bacterium in the rhizosphere of watermelon. Appl. Soil Ecol. 59, 13–19 (2012).
Yao, S. R., Merwin, I. A., Abawi, G. S. & Thies, J. E. Soil fumigation and compost amendment alter soil microbial community composition but do not improve tree growth or yield in an apple replant site. Soil Biol. Biochem. 38, 587–599 (2006).
Shin, J., Kathuria, A. & Lee, Y. S. Effect of hydrophilic and hydrophobic cyclodextrins on the release of encapsulated allyl isothiocyanate (AITC) and their potential application for plastic film extrusion. J. Appl. Poly. Sci. 136, 48137–48146 (2019).
Liu, E. T., Li, Y. Y., Hu, Y. L., Sun, C. X. & Mao, L. Q. Effects of dazomet on edaphon and growth of Malus hupehensis rehd under continuous apple cropping. Acta Ecol. Sin. 34, 847–852 (2014).
Cha, S. et al. Pre-Plant fumigation of soils for nematode control affects the seedling production and morphological properties of pine and larch seedlings. MDPI For. 10, 843–854 (2019).
Fang, W. S. et al. Responses of nitrogen-cycling microorganisms to dazomet fumigation. Front. Microbiol. 9, 2529–2542 (2018).
Prider, J. & Williams, A. Using dazomet to reduce broomrape seed banks in soils with low moisture content. Crop Prot. 59, 43–50 (2014).
Mao, L. G. et al. Evaluation of the combination of dimethyl disulfide and dazomet as an efficient methyl bromide alternative for cucumber production in China. J. Agric. Food Chem. 62, 4864–4869 (2014).
Mao, L. G., Jiang, H. Y., Wang, Q. X., Yan, D. D. & Cao, A. C. Efficacy of soil fumigation with dazomet for controlling ginger bacterial wilt (Ralstonia solanacearum) in China. Crop Prot. 100, 111–116 (2017).
Harris, D. C. A comparison of dazomet, chloropicrin and methyl bromide as soil disinfestants for strawberries. J. Hortic. Sci. 66, 51–58 (1991).
Gilreath, J. P. & Santos, B. M. Efficacy of methyl bromide alternatives on purple nutsedge (Cyperus rotundus) control in tomato and pepper. Weed Technol. 18, 341–345 (2004).
Eo, J. & Park, K. C. Effects of dazomet on soil organisms and recolonisation of fumigated soil. Pedobiologia 57, 147–154 (2014).
He, J. Y. et al. The genus Codonopsis (Campanulaceae): A review of phytochemistry, bioactivity and quality control. J. Nat. Med. 69, 1–21 (2014).
Liu, Y., Ren, X. X. & Jeong, B. R. Manipulating the difference between the day and night temperatures can enhance the quality of Astragalus membranaceus and Codonopsis lanceolata plug seedlings. MDPI Agr. 9, 654–668 (2019).
Xiao, S. X. et al. Effects of different seedling grades on yield and commercial quality of Codonopsis pilosula. Agric. Biotech. 9, 18–21 (2020).
Bai, B. et al. Summarization of research on seedling raising technology of Codonopsis pilosula. Gansu Agric. Sci. Technol. 9, 69–73 (2018).
Jin, X. S. et al. Control effect and safety of dazomet 98% MG against weeds in Ginseng field. Agrochemistry 57, 682–686 (2018).
People’s Republic of China. Technical code of practice for dazomet soil disinfestation. NY/T 3129-2017.
Guan, S. Y. Soil Enzymes and Their Research Methods (Agricultural Press, 1986).
Zak, J. C., Willig, M. R., Moorhead, D. L. & Wildman, H. G. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem. 26, 1101–1108 (1994).
Garland, J. L. & Mills, A. L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 57, 2351–2359 (1991).
Garland, J. L. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiol. Ecol. 24, 289–300 (1997).
Dalal, M. & Khanna-Chopra, R. Lipid peroxidation is an early event in necrosis of wheat hybrid. Biochem. Bioph. Res. Co. 262, 109–112 (1999).
Dhindsa, R. S. & Wandekayi, M. Drought tolerance in two mosses: Correlated with enzymatic defence against lipid peroxidation. J. Exp. Bot. 1, 79–91 (1981).
Li, B. L. & Mei, H. S. Relationship between oat leaf senescence and activated oxygen metabolism. Acta Phytol. Sin. 15, 6–12 (1989).
Guo, F. X. et al. Relation of several antioxidant enzymes to rapid freezing resistance in suspension cultured cells from alpine Chorispora bungeana. Cryobiology 52, 241–250 (2006).
Guo, F. X. et al. Initiation of early bolting by pre-enhancing anthocyanin and catalase activity in Angelica sinensis tender leaf during medicine formation cultivation year. Russ. J. Plant Physl. 68, 763–773 (2021).
Niu, J. et al. The compensation effects of physiology and yield in cotton after drought stress. J. Plant Physl. S0176–1617(18), 30039–30047 (2018).
Inspection and Quarantine of the People's Republic of China. Industry standard for entry-exit inspection and quarantine of the People's Republic of China: Determination of crude polysaccharides in vegetable foods for export—Phenol-sulfuric acid method: SNT 4260–2015.
Yang, D. D. et al. Comparative analysis of chemical composition, antioxidant and antimicrobial activities of leaves, leaf tea and root from Codonopsis pilosula. Ind. Crop. Prod. 142, 111844–111852 (2019).
Gao, S. M. et al. Quantitative and HPLC fingerprint analysis combined with chemometrics for quality evaluation of Codonopsis Radix processed with different methods. Chin. Herb. Med. 2, 160–168 (2019).
Feng, Y. Z., Liu, Y. & Liu, W. Analytical method of dazomet and its metabolite methyl-isothiocyanate residue in tomato and soil. Agrochemicals 54, 588–590 (2015).
Jin, Y. B. et al. Effect of various crop residues on growth and disease resistance of Angelica sinesis seedlings in Min County. Acta Pratacul. Sin. 27, 69–78 (2018).
Qiu, X. Q. et al. Temporal and spatial distribution of root morphology of winter wheat. Sci. Agric. Sin. 46, 2211–2219 (2013).
Benlloch-Gonzalez, M., Berger, J., Bramley, H., Rebetzke, G. & Palta, J. A. The plasticity of the growth and proliferation of wheat root system under elevated CO2. Plant Soil 374, 963–976 (2014).
Wang, Z. H. et al. Effects of biochar on the microenvironment of saline-sodic soil and maize growth. MDPI Agric. 12, 2859–2876 (2022).
Sun, J. L. et al. Biochar and nitrogen fertilizer promote rice yield by altering soil enzyme activity and microbial community structure. GCB Bioenergy 14, 1266–1280 (2022).
Wang, G., Ren, Y., Bai, X. J., Su, Y. Y. & Han, J. P. Contributions of beneficial microorganisms in soil remediation and quality improvement of medicinal plants. MDPI Plants 11, 3200–3232 (2022).
Bai, G., Guo, F. X., Chen, Y., Yuan, H. C. & Xiao, W. J. Differences in physiological resistance traits of Angelica sinessis seedlings from uncultivated and cultivated fields in Min County. Acta Pratacul. Sin. 28, 86–95 (2019).
Wang, H. Y. et al. Effects of dazomet soil fumigation on characteristics of weed community in Codonopsis pilosula seedling cultivated fields. Chin. J. Eco-Agric. 29, 1827–1837 (2021).
Nicola, L. et al. Fumigation with dazomet modifies soil microbiota in apple orchards affected by replant disease. Appl. Soil Ecol. 113, 71–79 (2017).
Srivastava, R. K., Pandey, P., Rajpoot, R., Rani, A. & Dubey, R. S. Cadmium and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings. Protoplasma 251, 1047–1065 (2014).
Kanazawa, S., Sano, S., Koshiba, T. & Ushimaru, T. Changes in antioxidative enzymes in cucumber cotyledons during natural senescence: Comparison with those during dark-induced senescence. Physiol. Plant. 109, 211–216 (2000).
Zhu, J. K. Abiotic stress signaling and responses in plants. Cell 167, 313–324 (2016).
Jiang, W. T. et al. Effects of dazomet fumigation on growth, biological characteristics of Malus hupehensis seedlings and soil environment. Chin. J. Appl. Ecol. 31, 3085–3092 (2020).
Xu, B. Q., Chen, Y., Guo, F. X., Bai, D. T. & Zhou, D. X. Effect of transplanting densities on growth, development, yield and quality of Astragalus membranaceus var. mongholicus. Chin. J. Exp. Tra. Med. Formul. 26, 135–143 (2020).
Wu, H. M. & Lin, W. X. A commentary and development perspective on the consecutive monoculture problems of medicinal plants. Chin. J. Eco-Agric. 28, 775–793 (2020).
Fang, W. S. et al. Comparative analysis of the effects of five soil fumigants on the abundance of denitrifying microbes and changes in bacterial community composition. Ecotox. Environ. Saf. 187, 109850–109860 (2020).
Cai, Z. P., Wang, H. X., Wang, G. X., Mi, Y. W. & Wu, W. G. Effects of different seedling raising modes on the quality of Codonopsis pilosula seedlings in Weiyuan. Gansu. J. Chin. Med. Mater. 40, 779–781 (2017).
Acknowledgements
We gratefully acknowledge the help of the farmers Guo Z J, Guo T M in field cultivation and management. The authors are also very grateful to Jiao X.-S, Liu L.-L, Ge X, Zhang B.-Q for their diligent scientific work.
Funding
This study was supported by the “Star of Innovation” project for graduate students in Gansu Province in 2023 (NO. 2023CXZX-649); The Ministry of Agriculture and Rural Affairs of the People's Republic of China Foundation for Traditional Chinese Medicinal Materials Advantaged Characteristic Industry Cluster Construction of Improved Variety Breeding (NO. 2022GSCYJQ07-33); National Key R & D Project of China (NO. 2018YFC1706301); A Grant for a Chief Expert of Traditional Chinese Medicinal Industry in Modern Agricultural Industry System & Safety (NO. GARS-ZYC-1); The Breeding and Base Construction of New Varieties of Chinese Medicinal Materials (NO. 7010722087); Lanzhou youth science and technology talent innovation project (No. 2023-QN-162).
Author information
Authors and Affiliations
Contributions
Y.C. conceived and designed the study, F.X.G. improved the design and analyzed results. H.Y.W. performed the experiments. H.Y.W. analyzed the data and wrote the manuscript. F.X.G. and Y.C. revised and edited the manuscript. W.L., P.B.D. and J.L.C. participated in the field index determination. All authors read and approved the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, H., Chen, Y., Guo, F. et al. Improvement in the quality and productivity of Codonopsis pilosula seedlings by dazomet soil fumigation. Sci Rep 14, 5407 (2024). https://doi.org/10.1038/s41598-024-56093-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-56093-3
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
