
Abstract
The interspecific transmission of pathogens can occur frequently in the environment. Among wild bees, the main spillover cases are caused by pathogens associated with Apis mellifera, whose colonies can act as reservoirs. Due to the limited availability of data in Italy, it is challenging to accurately assess the impact and implications of this phenomenon on the wild bee populations. In this study, a total of 3372 bees were sampled from 11 Italian regions within the BeeNet project, evaluating the prevalence and the abundance of the major honey bee pathogens (DWV, BQCV, ABPV, CBPV, KBV, Nosema ceranae, Ascosphaera apis, Crithidia mellificae, Lotmaria passim, Crithidia bombi). The 68.4% of samples were positive for at least one pathogen. DWV, BQCV, N. ceranae and CBPV showed the highest prevalence and abundance values, confirming them as the most prevalent pathogens spread in the environment. For these pathogens, Andrena, Bombus, Eucera and Seladonia showed the highest mean prevalence and abundance values. Generally, time trends showed a prevalence and abundance decrease from April to July. In order to predict the risk of infection among wild bees, statistical models were developed. A low influence of apiary density on pathogen occurrence was observed, while meteorological conditions and agricultural management showed a greater impact on pathogen persistence in the environment. Social and biological traits of wild bees also contributed to defining a higher risk of infection for bivoltine, communal, mining and oligolectic bees. Out of all the samples tested, 40.5% were co-infected with two or more pathogens. In some cases, individuals were simultaneously infected with up to five different pathogens. It is essential to increase knowledge about the transmission of pathogens among wild bees to understand dynamics, impact and effects on pollinator populations. Implementing concrete plans for the conservation of wild bee species is important to ensure the health of wild and human-managed bees within a One-Health perspective.
Similar content being viewed by others

Introduction
With their pollination service, bees contribute to maintaining the health and function of the ecosystems, ensuring the biodiversity and reproduction of plants and guaranteeing crop production and food security1,2,3,4. Although the demand for pollination services is increasing5,6, many reports are revealing declines in bee diversity worldwide, a reduction of geographic ranges for several species, extinction for some species and a decrease in local abundance3,7,8,9,10,11. Several factors are contributing to this decline, such as pathogens spread, pesticides, climate change and habitat loss3,7,12,13. Regarding pathogens spread, the diffusion and introduction of new diseases (EIDs, Emerging Infectious Diseases) can happen due to host shifts of pathogens between populations3,12,14. The risk of infection increases where human activities occur, like commercial management of bees, mass breeding, transport and trade beyond their original ranges3,15,16. Different correlational evidence hypothesized that the most worldwide managed Apis mellifera can act as a source of pathogens (maintenance host), which can spread into wild bee species (incidental host)17,18,19,20,21,22. However, this hypothesis includes historical reasons, since many pathogens and diseases were originally discovered in honey bees18,21,23. In addition, the route of interspecific transmission is difficult to determine and in most cases is unknown23. Pathogen spread is promoted when infected bees contaminate the same environment, that is populated by other new host species21. This indirect transmission can occur through sharing of food, fecal contact, contact with another infected organism, a vector and predation18,23,24,25,26,27,28,29,30,31,32,33,34,35,36. Sharing the same contaminated resources, such as flowers, pollen, honey and nectar, is a successful route of infection23. Flowers are described as “dirty doorknobs”, as they facilitate the spread of pathogens and can contain infective particles deposited by infected hosts25,26,27,28,29. Since bees are obligate flower visitors, the distribution, diversity, and abundance of floral resources in the environment are important for promoting interspecific interactions and potential pathogen diffusion37. Expansion of agriculture and landscape simplification can affect the floral availability for bees and impact the dynamics and transmission of pathogens38,39. Furthermore, meteorological conditions can alter both lifestyles of the host and pathogen persistence in the environment20; for example, in the case of viruses, high levels of UV can rapidly deactivate virus particles and the same happens when temperatures increase40,41. Interactions between bees and the environment are mediated by the biological traits of bees, affecting disease dynamics, susceptibility to infection and exposure to pathogens37,42. Yet, only a few studies have recently begun to investigate functional trait role in influencing the prevalence of pathogens37,42. Sociality could exacerbate or mitigate the diffusion of disease because living in a colony implies both positive and negative aspects in terms of hygiene37,42,43,44,45. Different nesting locations can influence pathogen transmission and their persistence in the nest42,46,47. Additionally, diet and preference for flower resources can modulate the spread of pathogens, since specialist bees collect pollen on a limited number of plant species, while generalist bees forage on multiple plants29,37,38,42,48. Also, voltinism may influence the diffusion of diseases, through the number of individuals generated and the nest density49. According to a One-Health approach, these traits, along with meteorological and environmental ones, could be crucial to understand pathogens dynamics between wild bees, in order to reduce the spread of diseases and promote concrete and effective conservation projects50,51,52. The purpose of this study was to investigate the occurrence and circulation of honey bee pathogens in the wild bees of Italy. Also, we aimed to use statistical models to assess the relative importance of factors influencing pathogen occurrence and to predict the risk of infection in wild bees. Accordingly, prevalence and abundance data were tested with apiary density, meteorological variables, agricultural management, and biological traits of bees.
Material and methods
Sampling
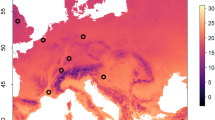
This study was conducted within the project BeeNet, which monitors honey bee colonies and wild bees in Italy in managed agricultural areas on two Corine Land Cover Categories: intensive (category 2.1.1.1) and semi-natural (category 2.4.3)53. The sampling was conducted once a month, from February to September 2022, in 24 sites located in 11 regions of Italy (Fig. 1). In each site, all sampling was carried out excluding fields blooming and focusing on spontaneous vegetation likewise field margins, ditches and meadows21. Table S1 reports the sampling site acronym (used across the text), the agro-environmental characteristics (intensive, semi-natural) and geographic reference (region, closest town, province) of each sampling site.

The geographical location of the sampling agricultural site. Orange spots represent intensive agricultural sites, while the green spots represent semi-natural agricultural sites.
The sampling was conducted on sunny and non-windy days, with an average temperature above 15 °C21,42. Sampling consisted in collecting wild bees foraging on flowers and flying. Honey bees were caught up to a maximum of 5 individuals per sampling, given their abundance. Bees were collected using the sweep net technique for one effective hour and each individual was placed in a sterile single 2-ml microtube or 15-ml tube21. After collection, the tubes were placed in a cooler bag with frozen packs to maintain samples at low temperature. Upon arrival at the CREA laboratories in Bologna, the specimens were all identified to the species level21,42.
Before being identified all collected bees were placed at −80 °C for 30 min. Identification was performed under a stereomicroscope, with the individual placed in a Styrofoam container with dry ice to prevent RNA degradation. Following identification, samples were kept at −80 °C until analysis.
Extraction of nucleic acids
All samples were washed in 95% ethanol before extraction to eliminate any external microbiological contaminations. Each bee was examined individually. The sample was put in a 2-ml microtube with 500 µl of DNA/RNA Shield (Zymo Research, Irvine, CA, USA) and crushed for 3 min at 30 Hz with a TissueLyser II (Qiagen, Hilden, Germany), as previously reported21,54,55. The suspensions were separated into two aliquots from which DNA and RNA were extracted separately. The extraction of the DNA and RNA was performed using a Quick DNA Microprep Plus Kit (Zymo Research) and a Quick RNA Microprep Plus Kit (Zymo Research), respectively, following the modified manufacturer's instructions for solid tissue processing21,35,56. The extracted nucleic acids were eluted in 200 µl of DNAase-RNase-free water and kept at −80 °C until the qPCR analysis.
Quantitative real-time PCR (qPCR) assays
A quantitative Real-Time PCR (qPCR) analysis was performed to determine the abundance of each pathogen in the samples using the extracted DNA and RNA. Ascosphaera apis, Nosema ceranae and trypanosomatids (Lotmaria passim, Crithidia mellificae and C. bombi) were detected using DNA, while viruses (deformed wing virus—DWV; black queen cell virus—BQCV; chronic bee paralysis virus—CBPV; acute bee paralysis virus—ABPV; Kashmir bee virus—KBV) were investigated using RNA. The primers used for the qPCRs are reported in Table S2.
A total reaction volume of 10 µl was produced for each target gene using SYBR™ green assays with forward and reverse primers and nucleic acid extract adding 2 µl of extracted DNA or RNA, as reported in previous studies21,57. The SYBR PowerUp™ SYBR™ Green Master Mix (ThermoFisher, Waltham, MA, USA) and the Power SYBR™ Green Cells-to-CT™ Kit (ThermoFisher Scientific) were used for the DNA and RNA, respectively. The qPCRs were carried out using a QuantStudio™ 3 Real-Time PCR System (ThermoFisher Scientific), according to the protocols for each gene sequence34,58,59,60,61,62,63. DNA and RNA previously extracted from positive honey bees were employed as positive controls for each pathogen investigated; while, as a negative control, sterile water was used. All the analyses were carried out in duplicate.
A standard curve was created for each of the target genes by amplifying serially diluted recombinant plasmids containing the pathogen-specific DNA and RNA fragment from 1 × 101 to 1 × 109 copies in a qPCR assay on QuantStudio TM 3 Real-Time PCR System (ThermoFisher Scientific), as reported in previous studies using the amplification and quantification protocols34,59,60,61,62,63,64.
Statistical analysis
For the statistical analysis, a database (Table S3) was created including all sampled bees (with associated identification code, species name, region, month and sampling site). Besides, regions were associated with a latitude-based macro category (North, Central, or South Italy), while sampling sites were also clustered according to environmental management (semi-natural or intensive). Two meteorological variables were selected for their possible influence on the prevalence and abundance of pathogens: maximum daily temperature (Tmax) and daily mean relative humidity (RH). For each sampling day, the two meteorological data were obtained from the NASA Langley Research Center (LaRC) POWER Project (https://power.larc.nasa.gov/). The honey bee apiary density per kilometre for each province related to the sampling site was also included in the database, collecting data from the National Database of Zootechnical Registry, section Beekeeping (BDN) (https://www.vetinfo.it/j6_statistiche/#/report-pbi/45). The data were employed at the provincial level since it represented the smallest available territorial unit in BDN about apiary density. Five functional traits of bees were selected as explanatory variables: sociality, voltinism, nesting habits, diet specialization and foraging range. These traits have already been hypothesized to influence the prevalence of pathogens42. Data were obtained from the “Wild Bees Functional Traits Database” (S. Roberts, unpublished, pers. comm.), constantly updated by numerous researchers. Each species was clustered in terms of bee group (honey bee, wild bee and cuckoo bees), sociality (highly eusocial, primitively eusocial, communal, solitary or cleptoparasite), nesting habits (managed apiary, mining, renter, carder, mason, and cleptoparasite bees), voltinism (univoltine, bivoltine and multivoltine), diet specialization (lecticism) reduced in only three categories (oligolectic or polylectic) and foraging range (as intertegular distance in mm)65. Cuckoo bees were maintained separated from “wild bees" due to their distinctive anatomy and biology features: they lack of pollen collecting structures, and it is almost impossible to be sure a female cuckoo laid her eggs in the nest of a single host female or at least host females belonging to the same species.
The prevalence of pathogens was calculated as the ration between the number of bees positive for pathogens and the total number of collected individuals. The pathogen abundance was calculated as the log10 transformed average of the results obtained from the two technical replicates. Explorative analyses were carried out through Spearman’s correlation between pathogens’ copy numbers, and the heatmaps creation for prevalence and abundance visualization for each investigated variable.
Time trend was explored through the analysis of variance. The distribution of pathogens’ abundance and prevalence were tested with a Shapiro–Wilk test in order to test their normality. Since normality was not satisfied to carry out parametric analyses, we employed Kruskal–Wallis with Dunn-test as post-doc to investigate differences.
Prevalence and abundance for each pathogen were fit with a generalized linear model (GLM) and linear mixed effects models (LMER). Due to the low number of infections (N < 50), KBV, L. passim, C. bombi, A. apis and C. mellificae were excluded from these analyses. In order to test meteorological, environmental and biological factors on the probability of infection for each pathogen, a logistic regression (GLM) was fit. The risk of infection was tested as a binomial response variable, where 1 specified the infection, and 0 described the absence of infection. Meteorological, environmental and biological factors were selected as predictors variables. Interactions were included in the two models to see their combined effect on pathogens’ prevalence. Attention was paid to the interaction between apiary density and environmental management since the pathogens investigated are closely related to the honey bee and different environmental management could affect pathogens’ prevalence. The interaction between temperature and relative humidity may influence the persistence and viability of the pathogen in the environment. The interaction between temperature and environmental management may affect the pathogens exposition among different environmental management. It was necessary to create two separate models for problems in aliased variables found in biological traits. The first model was tested with all meteorological and environmental variables and the explanatory variable “group of bees” (honey bees, cuckoo bees and wild bees). Model (GLMM1) was fit following this general formula (R syntax):
The second model was tested with the same data, but honey bee and cuckoo bee were excluded (GLMM2), with this general formula (R syntax):
The relationship between pathogens’ abundance and environmental and biological factors was evaluated using linear mixed-effects models (LMER). In these models, sampling sites were chosen as random effects and other meteorological, environmental and biological variables as fixed effects. The same approach to alias data was conducted for these models. Two different models were created. The first model (LMERM1) was fit using meteorological, and environmental variables and the explanatory variable “group of bees”, following this formula (R syntax):
The second model was tested with the same data, but honey bee and cuckoo bee were excluded (LMERM2), with this general formula (R syntax):
Significance for all models was determined by calculating the Type-II analysis of variance, with the sequent test post-hoc Tukey to find significance between factors.
Finally, a chord diagram was created to show co-infection between individuals belonging to the same genus.
The significative threshold was set at 0.05.
All the analyses were conducted in R 4.2.2 (r-project.org). Data manipulation, analysis and graphical representation were carried out with agricolae, car, caret, circlize, corrgram, corrplot, dplyr, ggplot2, rstatix, and sjplot packages66,67,68,69,70,71,72,73,74,75.
Results
A total of 3372 bees were collected and analysed from the11 regions of Italy, involved in the project. A high number of samples was reported for Tuscany (N = 637), Emilia-Romagna (N = 508) and Campania (N = 430), followed by Sicily (N = 383), Piedmont (N = 361), Friuli-Venezia Giulia (N = 272), Apulia (N = 229), Umbria (N = 171), Sardinia (N = 160), Veneto (N = 119) and Abruzzo (N = 102).
The highest number of bees was sampled in July (N = 631), while the lowest in February (N = 45).
Overall, 39 bee genera were identified (Fig. 2). The most frequently sampled genera were Andrena (Fabricius, 1775) (N = 509), Lasioglossum (Curtis, 1833) (N = 417), Halictus (Latreille, 1804) (N = 298), Bombus (Latreille, 1802) (N = 282), Eucera (Scopoli, 1770) (N = 280), Ceratina (Latreille, 1802) (N = 210), Hylaeus (Fabricius, 1793) (N = 176), Seladonia (Robertson, 1918) (N = 175), Megachile spp. (N = 140), Heriades spp. (N = 137), Osmia spp. (N = 84) and Anthophora (Latreille, 1802) (N = 60). Besides, a total of 305 Apis mellifera L. were collected. The number of samples for all other genera was under 50. The number of samples collected per region, sampling site and month are reported in Table S4.

The total number of bees collected. Each genus is represented by different color.
The trypanosomatid C. mellificae was the only pathogen not detected in any samples. A total of 2307 of the individuals sampled were positive for at least one pathogen (68.4%) (Table 1). Overall, DWV, BQCV, N. ceranae, CBPV and ABPV were the five most prevalent pathogens. Lower prevalence was detected for L. passim, KBV, C. bombi and A. apis.
The pathogen with the highest abundance was DWV, followed by BQCV, CBPV, ABPV and N. ceranae (Table 1). All the other pathogens had an abundance below 10 × 102. On average, the pathogen abundance for individuals sampled was found to be 7.05 × 109 ± 2.45 × 1011.
The most prevalent pathogens among almost all genera were DWV, N. ceranae and BQCV (Fig. S1a). A high abundance of DWV was found in Sphecodes spp. (10 × 108) and Pasites spp. (10 × 108), while the abundance was high in Lithurgus spp. (10 × 105) for N. ceranae and in Tetralonia spp. (10 × 105) for BQCV. Regions of Abruzzo and Veneto showed a high prevalence of DWV (21.0% and 24.5%, respectively) (Fig. S1b). Sardinia showed a high abundance of DWV (10 × 107) and BQCV (10 × 105). A high prevalence of N. ceranae was detected for both Sardinia sampling site (SAES with 65.2% and SAAI with 61.5%) and a high abundance of DWV (SAES and SAAI with both 10 × 107) (Fig. S1c.). Multivoltine bees showed a high prevalence and abundance of DWV (48.2% and 10 × 103, respectively) (Fig. S1d). Prevalence and abundance data of multivoltine bees, highly eusocial bees (Fig. S1e) and the managed apiary (Fig. S1f.) corresponded perfectly since these categories were referred to A. mellifera. DWV prevalence and abundance were also high for bivoltine bees (68.3% and 10 × 105, respectively) (Fig. S1d), cleptoparasite bees (60% and 10 × 105, respectively) (Fig. S1e), communal bees (68.2% and 10 × 104, respectively), excavator bees (57.4% and 10 × 104, respectively) (Fig. S1f.) and oligolectic bees (59.6% and 10 × 104, respectively) (Fig. S1g). DWV prevalence was reported as 80.0% in February and 18.8% in March and DWV abundance as 10 × 106 in February and March (Fig. S1h).
Seasonal trend
A different seasonal trend considering pathogen abundance was noted (Fig. 3a). In March, a high abundance of DWV was detected. After a brief decline, DWV increased and reached another peak in June. Then, a drastic decline occurred from June to July and resumed increasing since September. A similar trend was found for BQCV and N. ceranae, but in these cases, the abundance decreased in September. In April a high abundance of ABPV was reported, while for CBPV the peak of abundance was reached in June. Significant differences among months were reported in Table S5. The abundance of C. bombi was significantly higher in April and August, while the abundance of L. passim was significantly higher in April. No significant differences were detected for KBV and A. apis.

Pathogen abundance (a) and prevalence (b) throughout months (March–September). The time trend of each pathogen is highlighted in each graph. Abundance (red lines) is shown as a decimal logarithm and prevalence (blue lines) is shown as a percent.
The analysis of seasonal trend of pathogen prevalence throughout the months (Fig. 3b) started in March since the samples collected in February were only 45. In March, a high prevalence of the pathogens DWV, BQCV, ABPV, CBPV and N. ceranae was found. In April, the prevalence of these pathogens decreased drastically. A slight increase in prevalence was recorded in June. Then, pathogen prevalence again reached a peak in September. Significant differences among months were reported in Table S6. The prevalence of C. bombi was significantly higher in April and August, while the prevalence of L. passim was significantly higher in April. For KBV and A. apis no significant differences were detected.
Pathogens’ correlation
A positive abundance correlation was detected between DWV and ABPV, DWV and CBPV, DWV and BQCV, and DWV and N. ceranae. For CBPV, a positive correlation was reported with BQCV, N. ceranae and A. apis. A positive correlation was found between KBV and C. bombi and between BQCV and N. ceranae. For ABPV, a negative correlation was detected with CBPV and BQCV (Fig. 4). Table S7 reported the p-value correlation between pathogens.

Positive and negative correlation between pathogens. Only significant correlations are shown. High or low values of correlation are shown with ellipsoidal or circular shapes, respectively.
GLM and LMER analysis
GLM are summarized in Table 2 and Figs. S2 and S3, while LMER results in Table 3 and Fig. S4 and S5. Post tests are available in Tables S8 and S9.
DWV models
The occurrence and load of DWV were significantly negatively related to temperature and relative humidity. The GLM models showed that the probability of DWV occurrence was also significantly related to latitude. In particular, there was a high significant probability of DWV occurrence in South Italy. The probability of DWV occurrence and load was significantly higher in cuckoo bees compared to wild bees and honey bees. There was a significant interaction between temperature and relative humidity in relation to DWV infections. Specifically, high temperatures and lower humidity increased the probability of infections. There was another significant interaction between apiary density and environmental management. In particular, in a semi-natural environment, the probability of infections increased with higher density of the apiaries.
In wild bees, the probability of infection and relative load were significantly related to functional traits: voltinism, sociality, nesting and lectism Specifically, bivoltine bees were significantly more susceptible to infection compared to univoltine bees. Primitively eusocial bees were significantly less likely to be infected than solitary bees and communal bees. Excavator bees were significantly more susceptible to infection compared to carder and renter bees. Oligolectic bees were significantly more likely to be infected than polylectic bees. Also, DWV load in LMER models was significantly negatively related to the foraging range.
BQCV models
The occurrence of BQCV was significantly correlated with the apiary density, in particular, the occurrence decreased with the increase in apiary density. Also, the occurrence of BQCV was significantly positively correlated with relative humidity. The probability of infections was significantly lower in the South compared to North or Central Italy, while the infection of BQCV was significantly higher in bees sampled in areas with intensively managed soils than in areas with semi-natural management. The interaction between temperature and relative humidity was significantly correlated with the BQCV occurrence, specifically, there was a high probability of infection with lower temperatures and higher relative humidity. The BQCV load was significantly negatively correlated to temperature. The interaction between temperature and environmental management was significantly correlated with the viral load: the risk to contract a high load of BQCV was higher in low temperatures where bees were sampled in environments with intensive management.
The probability of infection for wild bees was significantly correlated to functional traits: voltinism, sociality, construction of the nest and diet specialization, as well as the BQCV load. The BQCV occurrence and load were significantly higher for bivoltine bees compared to univoltine bees, while the probability of infection was significantly lower for primitively eusocial bees related to solitary bees, especially compared to communal bees. Renter bees were significantly less susceptible to infection than excavator bees, whereas oligolectic bees were significantly more susceptible to infection than polylectic bees, as well as for BQCV load.
CBPV models
The occurrence and load of CBPV were significantly negatively correlated with temperature and significantly positively correlated with relative humidity. The probability of infection and abundance were significantly higher for honey bees compared to wild bees.
The probability of CBPV infection in wild bees was significantly correlated with environmental management, specifically, there was a high probability to contract the infection in environments with intensive management. The occurrence of CBPV in wild bees was significantly correlated with functional traits voltinism and sociality. In particular, bivoltine bees were significantly more susceptible to infection compared to univoltine bees, while communal bees were significantly more susceptible to infection compared to primitively eusocial bees, such as for the CBPV load.
ABPV models
The occurrence of ABPV was significantly positively correlated with the apiary density. The probability of infection and the load were significantly negatively correlated with temperature and relative humidity. The probability to contract infections of ABPV was significantly lower in Central Italy compared to North and South Italy, while this probability was significantly higher for bees sampled in areas with semi-natural management compared to intensive management. There was a significant interaction between temperature and relative humidity related to ABPV occurrence, in particular, the probability of infection increased at lower temperatures and lower relative humidity, as well as for abundance. The occurrence of ABPV was significantly correlated with the interaction between apiary density and environmental management. In detail, the risk of infection increased with the increase of apiary density in semi-natural environments. Another significant interaction between temperature and environmental management showed the increase of ABPV occurrence and load when temperatures decrease in semi-natural environments. Cuckoo bees were significantly highly susceptible to ABPV infections compared to wild bees and honey bees, such as for ABPV abundance.
The probability of infection was significantly correlated to voltinism and sociality. In particular, bivoltine bees were significantly less susceptible to ABPV infection compared to univoltine bees. Also, primitively eusocial bees were significantly less susceptible to ABPV infection compared to solitary bees, the same was found for abundance. In addition, in the LMER model excavator bees were significantly more likely to have a high viral load of ABPV compared to renter bees.
N. ceranae models
The occurrence and load of N. ceranae were significantly negatively correlated to temperature and relative humidity. There was a significant interaction between temperature and relative humidity, in particular, the risk of infection decreased much more at high relative humidity when the temperatures increased. In addition, in the LMER model honey bees were significantly more likely to have a high load of N. ceranae compared to wild bees.
The probability of infection in wild bees was significantly negatively correlated with apiary density. The risk of infection was significantly correlated with functional traits: voltinism, sociality, construction of the nest and diet specialization. Bivoltine bees were significantly more susceptible to N. ceranae compared to univoltine bees, as reported for N. ceranae abundance. The probability of N. ceranae occurrence and load were significantly higher for solitary bees compared to primitively eusocial bees. The probability of infection was significantly lower for renter bees compared to excavator bees, while this probability was significantly higher for oligolectic compared to polylectic bees.
Co-infections
A total of 1365 bees (40.5%) belonging to 173 different species were found to be co-infected with two or more pathogens. Eight infected individuals were detected with five pathogens concurrently. In particular, the co-infection of DWV, BQCV, ABPV, CBPV and N. ceranae was detected in two individuals of Eucera eucnemidea Dours, 1873, one Eucera nigrifacies Lepeletier, 1841, one Chelostoma florisomne (L., 1758) and one Ceratina cucurbitina (Rossi, 1792). One individual of Eucera vulpes (Brullé, 1832) was detected with a co-infection of DWV, BQCV, CPBV, N. ceranae and A. apis. An individual of Andrena impunctata Pérez, 1895 was co-infected with DWV, BQCV, CBPV, N. ceranae and C. bombi. One individual of Andrena humilis Imhoff, 1832 was detected with a co-infection of DWV, BQCV, CBPV, N. ceranae and L. passim. The genera with the highest number of multiple infections were Andrena (Fabricius, 1775), Eucera (Scopoli, 1770), Lasioglossum (Curtis, 1833), Apis mellifera L. and Halictus (Latreille, 1804) (Fig. 5).

A visual schematization of the investigated pathogens that were involved in multiple infections related to the genera implicated. The arrow thickness denotes the number of co-infections observed within the same host genera. In the graph only bees infected with at least two pathogens appear.
Discussion
There is very little information about the detection of pathogens among wild bees in Italy. This is the first study providing large-scale monitoring, involving 11 out of 20 Italian regions. A pilot study was previously conducted in 2021 in the same sites in Emilia-Romagna and Piedmont regions highlighting the infection of 13 pathogens on investigated wild pollinators21. In Italy, other studies have confirmed the presence of honey bee viruses in Vespa velutina Lepeletier, 1836 and V. orientalis L., 1771, probably infected by eating infected honey bees34,35,76,77. In addition, a queen of the hornet V. crabro L., 1761 was found to be symptomatic for DWV, showing short and crippled wings76. The possible effects and symptoms of these pathogens on new hosts are mostly unknown and there are insufficient data to define the real impact of pathogens on wild bee communities28,78,79,80,81.
In this study, 68.4% of the sampled specimens tested positive for at least one pathogen. This result is in line with previous investigations conducted in Italy, France and U.S.A., with 69.3%, 79% and 80.4% of samples scored infected, respectively21,78,82,83.
The highest prevalence was found for DWV, BQCV and N. ceranae (51.5%, 26.6% and 22.7% respectively). DWV was the most prevalent pathogen. This virus is widespread in several arthropod species, indicating its strong adaptability to new hosts18,21,84 and its generalist nature24,78,84,85. BQCV is a less studied but globally distributed virus among honey bees86. In 2016 it was found to be highly prevalent (75%) within Italian honey bee colonies87, while in Croatian bumblebees a prevalence of 58.5% and 88.9% was detected19,88. N. ceranae is also a highly adaptive pathogen and it was found in several arthropod species18,64,89,90,91.
The viruses CBPV and ABPV were also found with a prevalence of 16% and 12.1% respectively. Both viruses were detected in other hymenopterans. In France, 66.7% of investigated samples were infected by ABPV, while CBPV was not detected82. On the other hand, in Croatia, both viruses were detected in 3.7% of the bumblebees88, while in Italy, the prevalence of the two viruses was 9.5% and 5.5%, respectively21. This frequency could be linked to the infection of both viruses in the honey bee colonies in which they have specific seasonal trends and epidemiology87,92,93,94.
Currently, information on the presence of KBV, trypanosomatids and A. apis in Italy is still scarce. In this study, the presence of C. mellificae was not found, while L. passim, C. bombi, KBV and A. apis were also found with a prevalence of 1.2%, 0.6%, 0.7% and 0.2%, respectively. In Italy, the presence of C. mellificae had been detected for the first time in 2014 in one apiary in the Lazio region95, followed by another identification in the Veneto region in 202096. However, in the Veneto region, the presence of L. passim was found to be 48.8% and 62.2% in 2020 and 2021, respectively96, while no presence was detected in the Emilia-Romagna region97. The first Italian identification of KBV was notified in the Lazio region98, and only a few cases were detected in Italy87. Among pollinators, KBV and A. apis prevalence were respectively 0.7% and 0.2%, finding positive bees and syrphids21, in line with the results of this study.
Most pathogens averaged an abundance per individual of less than 10 × 105, which is lower than the threshold generally linked to symptomatic infection in A. mellifera (10 × 106)21,28,99,100,101. DWV was the virus with the highest mean abundance, with an average viral load of 10 × 1010. Besides, a high average abundance was found for BQCV, with an average viral load of 10 × 109. For CBPV the average viral load was found to be 10 × 106. These findings indicated a widespread infection among wild bees that may be related to the high prevalence of all three viruses among honey bee colonies and their spread in the environment80,87,92,93,94,102,103,104,105.
The trends of the five most prevalent pathogens (DWV, BQCV, CBPV, ABPV and N. ceranae) were in line with the same seasonal trends in honey bee colonies21,87,99,106. In honey bees, these pathogens show an initial increase in cases of infection during early spring with a peak in late spring/early summer when the number of workers is very high, and in many cases triggered by Varroa infestation21,87,107,108,109. In this study, it is interesting to note the decrease in cases of infection in the middle of the spring period; this could be related to a possible dilution phenomenon, where an increase in the biodiversity of pollinator species could promote a "thinning" of pathogens among bees43,110. In addition, greater biodiversity and availability in terms of flower species could reduce pathogen transmission among bees30,43,111. The peak of pathogen abundance reached in June may be related both to population dynamics within the honey bee colony, where in this month the colony reaches maximum density and activity, but also to the availability of floral resources87,99,106,111.
The inclusion of meteorological, environmental, and biological factors in the statistical models could be considered as successful predictors to the infection risk among bees. The increase in temperature and relative humidity suggests the risk of infection reduction, probably related to the host ecology and the persistence of the pathogen in the environment20. In fact, UV levels can inactivate viral particles in flowers, and the increase in temperature together with humidity can reduce the permanence of pathogens in the environment20,40,41,112,113. In addition, the high temperatures reduced the flowers anthesis causing a decrease in bees foraging activity114,115, causing also oxidative stress and mortality116,117.
In this study, apiary density was related to an increasing ABPV infection risk, probably linked to the in-hive spread of the virus by Varroa bite23,118,119. On the other hand, apiary density has a negative effect on the risk of BQCV and N. ceranae infection. BQCV is mainly related to honey bee colonies affecting queen brood cells and it can be assumed that it is related to eusociality, as reported also for bumblebees19,88,105,120,121. Besides, N. ceranae spores can remain quiescent in the environment for many years, replicating in the host after ingestion87,122,123.
The probability of infection depended on latitude, probably related to the different meteorological conditions present in Italy. Southern Italy is characterized by a Mediterranean climate, while northern and central Italy ranges from a humid subtropical to a humid continental climate124. Different infections related to latitude may be linked to the presence of different pathogens’ genetic variants and their environmental-adapted circulation in specific geographical areas18,21,103.
The types of agricultural environmental management have a different association with the risk of infection, depending on the pathogen. The increase of BQCV and CPBV infection in intensively managed environments could be linked to the synchronized bloom events of monocultures, leading to a high density of bees in a specific area and consequently exacerbating the spread of pathogens37,39,43. Instead, the risk of ABPV infection in a semi-natural environment could be related to the presence of honey bees, because the massive use of pesticides in intensive agricultural areas could discourage beekeeping activity in them23,37,119.
The biological and social traits of bees were found to play a key role in the risk of infection. DWV and ABPV risks were higher in cuckoo bees. Their cleptoparasitic behavior toward their hosts, such as robbing of food resources and exploitation of parental care, may exacerbate the horizontal transmission23,24. Although in other studies sociality was not found to be relevant in the transmission of pathogens82,125, in this work, primitively eusocial bees presented a lower risk of infection compared to communal bees. In communal bees, multiple reproductive females share the same nest and lay their eggs, this aggregation could promote the horizontal and passive spread of viruses37,126. However, in primitively eusocial bees, specific behavioural or physiological immunity could help to maintain the health of the nest127,128,129. For example, newly-emerged B. terrestris (L. 1758) individuals may improve their resistance against C. bombi by feeding with faeces to obtain bacterial microbiota130, even if there are still little information available for social species, excluding honey bees.
Bivoltine bees and solitary excavator bees were found to be more susceptible to pathogen infection, most likely due to their reproductive cycle and nest location. The generation of two broods per year could intensify vertical transmission37,131. Often in such cases, the same nests are used for the next brood, facilitating the pathogen transmission between generations132, even if the influence of nest characteristics on susceptibility to pathogens remains unclear. However, less sun exposure on the ground and low temperatures may favour the persistence of viral particles in the soil and the resistance of N. ceranae spores42,123,132. In addition, many pesticides used in agriculture remain bound to soil particles from a few months to many years and several studies have shown that these products can affect the immunocompetence of bees132,133.
In this study, a significant correlation was found between N. ceranae and BQCV. Both pathogens could interact together promoting co-infections, and causing a synergistic effect on honey bee mortality134,135,136,137. The association between DWV and N. ceranae could also cause a synergistic effect, accelerating the DWV replication in honey bees138. A significant negative correlation was observed between ABPV and BQCV, hypnotizing a possible competition between these viruses, able to co-infect bees139. The significant and positive correlation between DWV and BQCV confirms the possible interaction of both viruses in the hosts, even if the synergistic effects remain still unclear140.
In this study, a high number of co-infected individuals were detected. There are several researches on the prevalence of pathogens in bees that report multiple infections, and it is often common in the natural environment18,21,102,137,141,142,143,144.
Conclusion
This study extended our knowledge of the epidemiological situation of honey bee pathogens in 11 Italian regions and their circulation among wild bees. The use of models has been fundamental in predicting the risk of infection among bees and in understanding which social and ecological factors influence pathogen interspecific occurrence. Although the investigated pathogens are known as specific to the honey bee for historical reasons, and the most prevalent hypothesis is that this species may act as a reservoir, the directionality of the transfer is difficult to identify and prove. It appears that only the spread of some pathogens is correlated with the presence of the honey bees, while for other pathogens there is already active circulation between different bee species. The share of the same environment and food resources could increase the pathogens' transmission, although further studies are needed to clarify these dynamics.
The most emerging aspect is that some bee species may be more easily susceptible than others, due to their biological and behavioural features. However, the high lack of data for many species makes it difficult to assess the impact of pathogens on these populations. Besides, the effects and symptoms of pathogens infection in wild bees are still lacking, and further studies on fitness, behaviour and development are needed to increase conservation efforts of wild bees.
This study provided evidence of a strong relationship between the health of the environment, pollinators and human activities, as part of a One Health approach, which is essential to protect the features and functionality of ecosystems.
Data availability
All data generated and analysed in this study are included in this published article and in its related supplementary information files.
References
Drossart, M. & Gérard, M. Beyond the decline of wild bees: Optimizing conservation measures and bringing together the actors. Insects 11, 1–23 (2020).
Potts, S. G. et al. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 25, 345–353 (2010).
Potts, S. G. et al. Safeguarding pollinators and their values to human well-being. Nature 540, 220–229 (2016).
Klein, A. M., Boreux, V., Fornoff, F., Mupepele, A. C. & Pufal, G. Relevance of wild and managed bees for human well-being. Curr. Opin. Insect Sci. 26, 82–88 (2018).
Aizen, M. A. & Harder, L. D. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 19, 915–918 (2009).
Panziera, D., Requier, F., Chantawannakul, P., Pirk, C. W. W. & Blacquière, T. The diversity decline in wild and managed honey bee populations urges for an integrated conservation approach. Front. Ecol. Evol. 10, 93 (2022).
Sánchez-Bayo, F. & Wyckhuys, K. A. G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 232, 8–27 (2019).
Zattara, E. E. & Aizen, M. A. Worldwide occurrence records suggest a global decline in bee species richness. One Earth 4, 114–123 (2021).
Carvalheiro, L. G. et al. Species richness declines and biotic homogenisation have slowed down for NW-European pollinators and plants. Ecol. Lett. 16, 870–878 (2013).
Kerr, J. T. et al. Climate change impacts on bumblebees converge across continents. Science (80-) 349, 177–180 (2015).
Bartomeus, I. et al. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Natl. Acad. Sci. USA 110, 4656–4660 (2013).
Cariveau, D. P. & Winfree, R. Causes of variation in wild bee responses to anthropogenic drivers. Curr. Opin. Insect. Sci. 1, 104–109. https://doi.org/10.1016/j.cois.2015.05.004 (2015).
Dicks, L. V. et al. A global-scale expert assessment of drivers and risks associated with pollinator decline. Nat. Ecol. Evol. 5, 1453–1461 (2021).
Goulson, D., Nicholls, E., Botías, C. & Rotheray, E. L. Bee declines driven by combined Stress from parasites, pesticides, and lack of flowers. Science (80-) 347, 1255957 (2015).
Moritz, R. F. A., Härtel, S. & Neumann, P. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Ecoscience 12, 289–301 (2016).
Vanbergen, A. J. et al. Threats to an ecosystem service: pressures on pollinators. Front. Ecol. Environ. 11, 251–259 (2013).
Jones, L. J., Ford, R. P., Schilder, R. J. & López-Uribe, M. M. Honey bee viruses are highly prevalent but at low intensities in wild pollinators of cucurbit agroecosystems. J. Invertebr. Pathol. 185, 107667 (2021).
Nanetti, A., Bortolotti, L. & Cilia, G. Pathogens spillover from honey bees to other arthropods. Pathogens 10, 1044 (2021).
Ocepek, M. P., Toplak, I., Zajc, U. & Bevk, D. The pathogens spillover and incidence correlation in bumblebees and honeybees in Slovenia. Pathogens 10, 884 (2021).
Piot, N. et al. Honey bees and climate explain viral prevalence in wild bee communities on a continental scale. Sci. Rep. 12, 1904 (2022).
Cilia, G. et al. Occurrence of Honey Bee (Apis mellifera L.) Pathogens in Wild Pollinators in Northern Italy. Front. Cell. Infect. Microbiol. 12, 7489 (2022).
Pritchard, Z. A. et al. Do viruses from managed honey bees (Hymenoptera: Apidae) endanger wild bees in native prairies?. Environ. Entomol. 50, 455–466 (2021).
Yañez, O. et al. Bee viruses: Routes of infection in hymenoptera. Front. Microbiol. 11, 943 (2020).
Manley, R., Boots, M. & Wilfert, L. Emerging viral disease risk to pollinating insects: Ecological, evolutionary and anthropogenic factors. J. Appl. Ecol. 52, 331–340 (2015).
Alger, S. A., Burnham, P. A. & Brody, A. K. Flowers as viral hot spots: Honey bees (Apis mellifera) unevenly deposit viruses across plant species. PLoS One 14, e0221800 (2019).
Burnham, P. A. et al. Flowers as dirty doorknobs: Deformed wing virus transmitted between Apis mellifera and Bombus impatiens through shared flowers. J. Appl. Ecol. 58, 2065–2074 (2021).
Graystock, P., Goulson, D. & Hughes, W. O. H. Parasites in bloom: Flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proc. R. Soc. B Biol. Sci. 282, 1371 (2015).
Mazzei, M. et al. Infectivity of DWV associated to flower pollen: Experimental evidence of a horizontal transmission route. PLoS One 9, e113448 (2014).
Piot, N., Smagghe, G. & Meeus, I. Network centrality as an indicator for pollinator parasite transmission via flowers. Insects 11, 872 (2020).
Piot, N. et al. More is less: Mass-flowering fruit tree crops dilute parasite transmission between bees. Int. J. Parasitol. 51, 777–785 (2021).
Yañez, O. et al. Deformed wing virus and drone mating flights in the honey bee (Apis mellifera): Implications for sexual transmission of a major honey bee virus. Apidologie 43, 17–30 (2012).
Gisder, S. & Genersch, E. Viruses of commercialized insect pollinators. J. Invertebr. Pathol. 147, 51–59 (2017).
Gisder, S., Aumeier, P. & Genersch, E. Deformed wing virus: Replication and viral load in mites (Varroa destructor). J. Gen. Virol. 90, 463–467 (2009).
Mazzei, M. et al. First detection of replicative deformed wing virus (DWV) in Vespa velutina nigrithorax. Bull. Insectology 71, 211–216 (2018).
Mazzei, M. et al. Detection of replicative Kashmir Bee Virus and Black Queen Cell Virus in Asian hornet Vespa velutina (Lepelieter 1836) in Italy. Sci. Rep. 9, 10091 (2019).
Forzan, M., Felicioli, A., Sagona, S., Bandecchi, P. & Mazzei, M. Complete genome sequence of deformed wing virus isolated from Vespa crabro in Italy. Genome Announc. 5, 1 (2017).
Proesmans, W. et al. Pathways for novel epidemiology: Plant–pollinator–pathogen networks and global change. Trends Ecol. Evol. 36, 623–636 (2021).
Figueroa, L. L. et al. Landscape simplification shapes pathogen prevalence in plant-pollinator networks. Ecol. Lett. 23, 1212–1222 (2020).
Cohen, H. et al. Mass-flowering monoculture attracts bees, amplifying parasite prevalence. Proc. R. Soc. B 288, 1 (2021).
Lytle, C. D. & Sagripanti, J.-L. Predicted inactivation of viruses of relevance to biodefense by solar radiation. J. Virol. 79, 14244–14252 (2005).
Pascall, D. J., Tinsley, M. C., Clark, B. L., Obbard, D. J. & Wilfert, L. Virus prevalence and genetic diversity across a wild Bumblebee Community. Front. Microbiol. 12, 856 (2021).
Figueroa, L. L., Compton, S., Grab, H. & Mcart, S. H. S. S. H. Functional traits linked to pathogen prevalence in wild bee communities. Sci. Rep. 11, 7529 (2021).
Graystock, P. et al. Dominant bee species and floral abundance drive parasite temporal dynamics in plant-pollinator communities. Nat. Ecol. Evol. 4, 1358–1367 (2020).
Pinilla-Gallego, M. S. et al. Within-colony transmission of microsporidian and trypanosomatid parasites in honey bee and bumble bee colonies. Environ. Entomol. 49, 1393–1401 (2020).
Kappeler, P. M., Cremer, S. & Nunn, C. L. (2015) Sociality and health: Impacts of sociality on disease susceptibility and transmission in animal and human societies. Philos. Trans. R. Soc. B Biol. Sci. 370, 20140116.
Leonard, R. J. & Harmon-Threatt, A. N. Methods for rearing ground-nesting bees under laboratory conditions. Apidologie 50, 689–703 (2019).
Wuellner, C. T. Nest site preference and success in a gregarious, ground-nesting bee Dieunomia triangulifera. Ecol. Entomol. 24, 471–479 (1999).
Ellner, S. P., Hao Ng, W. & Myers, C. R. Specialization and multihost epidemics: Disease spread in plant-pollinator networks. Am. Nat. 195, E118–E131 (2020).
Becker, D. J., Streicker, D. G. & Altizer, S. Using host species traits to understand the consequences of resource provisioning for host–parasite interactions. J. Anim. Ecol. 87, 511–525 (2018).
Wilfert, L., Brown, M. J. F. & Doublet, V. OneHealth implications of infectious diseases of wild and managed bees. J. Invertebr. Pathol. 186, 107506 (2021).
Donkersley, P., Ashton, L., Lamarre, G. P. A. & Segar, S. Global insect decline is the result of wilful political failure: A battle plan for entomology. Ecol. Evol. 12, 9417 (2022).
Gibbs, E. P. J. The evolution of One Health: A decade of progress and challenges for the future. Vet. Rec. 174, 85–91 (2014).
Giovanetti, M. & Bortolotti, L. Report on a project: BeeNet at the start. Bull. Insectology 284, 1 (2021).
Cilia, G. et al. Nosema ceranae infection in honeybee samples from Tuscanian Archipelago (Central Italy) investigated by two qPCR methods. Saudi J. Biol. Sci. 26, 1553–1556 (2019).
Nanetti, A., Ellis, J. D., Cardaio, I. & Cilia, G. Detection of Lotmaria passim, Crithidia mellificae and Replicative Forms of Deformed Wing Virus and Kashmir Bee Virus in the Small Hive Beetle (Aethina tumida). Pathogens 10, 372 (2021).
Nanetti, A. et al. Seed Meals from Brassica nigra and Eruca sativa Control Artificial Nosema ceranae Infections in Apis mellifera. Microorganisms 9, 949 (2021).
Cilia, G., Zavatta, L., Ranalli, R., Nanetti, A. & Bortolotti, L. Replicative Deformed Wing Virus found in the head of adults from symptomatic commercial bumblebee (Bombus terrestris) colonies. Vet. Sci. 8, 117 (2021).
Cilia, G. et al. A novel TaqMan ® assay for Nosema ceranae quantification in honey bee, based on the protein coding gene Hsp70. Eur. J. Protistol. 63, 44–50 (2018).
Xu, G. et al. Triplex real-time PCR for detection of Crithidia mellificae and Lotmaria passim in honey bees. Parasitol. Res. 117, 623–628 (2018).
Chantawannakul, P., Ward, L., Boonham, N. & Brown, M. A scientific note on the detection of honeybee viruses using real-time PCR (TaqMan) in Varroa mites collected from a Thai honeybee (Apis mellifera) apiary. J. Invertebr. Pathol. 91, 69–73 (2006).
James, R. R. & Skinner, J. S. PCR diagnostic methods for Ascosphaera infections in bees. J. Invertebr. Pathol. 90, 98–103 (2005).
Huang, W. F., Skyrm, K., Ruiter, R. & Solter, L. Disease management in commercial bumble bee mass rearing, using production methods, multiplex PCR detection techniques, and regulatory assessment. J. Apic. Res. 54, 516–524 (2015).
Arismendi, N., Bruna, A., Zapata, N. & Vargas, M. PCR-specific detection of recently described Lotmaria passim (Trypanosomatidae) in Chilean apiaries. J. Invertebr. Pathol. 134, 1–5 (2016).
Cilia, G., Cardaio, I., dos Santos, P. E. J., Ellis, J. D. & Nanetti, A. The first detection of Nosema ceranae (Microsporidia) in the small hive beetle, Aethina tumida Murray (Coleoptera: Nitidulidae). Apidologie 49, 619–624 (2018).
Greenleaf, S. S., Williams, N. M., Winfree, R. & Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 153, 589–596 (2007).
Kuhn, M. caret: Classification and Regression Training. (2022).
Fox, J. car: Companion to Applied Regression. (2022).
Wickham, H. dplyr: A Grammar of Data Manipulation. (2023).
Lüdecke, D. sjPlot: Data Visualization for Statistics in Social Science. (2022).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis. ggplot2 (Springer New York, 2009). https://doi.org/10.1007/978-0-387-98141-3.
de Mendiburu, F. agricolae: Statistical Procedures for Agricultural Research. https://cran.r-project.org/web/packages/agricolae/index.html (2021).
Gu, Z., Gu, L., Eils, R., Schlesner, M. & Brors, B. circlize Implements and enhances circular visualization in R. Bioinformatics 30, 2811–2812 (2014).
Wright, K. corrgram: plota a Correlogram. https://CRAN.R-project.org/package=corrgram (2021).
Wei, T. & Simko, V. corrplot: Visualization of a Correlation. https://cran.r-project.org/web/packages/corrplot/citation.html (2021).
Kassambara, A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests [R package rstatix version 0.7.2]. https://cran.r-project.org/web/packages/rstatix/index.html (2023).
Forzan, M., Sagona, S., Mazzei, M. & Felicioli, A. Detection of deformed wing virus in Vespa crabro. Bull. Insectol. 70, 261–265 (2017).
Power, K., Altamura, G., Martano, M. & Maiolino, P. Detection of honeybee viruses in Vespa orientalis. Front. Cell. Infect. Microbiol. 12, 896932 (2022).
Dolezal, A. G. et al. Honey bee viruses in wild bees: Viral prevalence, loads, and experimental inoculation. PLoS One 11, e0166190 (2016).
Zhang, L., Deng, Y., Zhao, H., Zhang, M. & Hou, C. Occurrence and phylogenetic analysis of DWV in Stingless Bee (Apidae sp.) in China: A case report. Front. Insect Sci. 1, 748074 (2021).
Tehel, A., Streicher, T., Tragust, S. & Paxton, R. J. Experimental infection of bumblebees with honeybee-associated viruses: No direct fitness costs but potential future threats to novel wild bee hosts. R. Soc. Open Sci. 7, 20212255 (2020).
Gómez-Moracho, T., Durand, T., Pasquaretta, C., Heeb, P. & Lihoreau, M. Artificial diets modulate infection rates by Nosema ceranae in bumblebees. Microorganisms 9, 158 (2021).
Dalmon, A. et al. Possible spillover of pathogens between bee communities foraging on the same floral resource. Insects 12, 122 (2021).
Levitt, A. L. et al. Cross-species transmission of honey bee viruses in associated arthropods. Virus Res. 176, 232–240 (2013).
Martin, S. J. & Brettell, L. E. Deformed wing virus in honeybees and other insects. Annu. Rev. Virol. 6, 49–69 (2019).
Genersch, E., Yue, C., Fries, I. & de Miranda, J. R. Detection of Deformed wing virus, a honey bee viral pathogen, in bumble bees (Bombus terrestris and Bombus pascuorum) with wing deformities. J. Invertebr. Pathol. 91, 61–63 (2006).
Beaurepaire, A. et al. Diversity and global distribution of viruses of the Western Honey Bee, Apis mellifera. Insects 11, 239 (2020).
Porrini, C. et al. The status of honey bee health in Italy: Results from the nationwide bee monitoring network. PLoS One 11, e0155411 (2016).
Gajger, I. T., Šimenc, L. & Toplak, I. The first detection and genetic characterization of four different honeybee viruses in wild bumblebees from Croatia. Pathogens 10, 808 (2021).
Porrini, M. P. et al. Nosema ceranae in South American native stingless bees and social Wasp. Microb. Ecol. 74, 761–764 (2017).
Purkiss, T. & Lach, L. Pathogen spillover from Apis mellifera to a stingless bee. Proc. R. Soc. B Biol. Sci. 286, 1 (2019).
Martínez-López, V. et al. Detection of microsporidia in pollinator communities of a mediterranean biodiversity hotspot for wild bees. Microb. Ecol. 84, 638–642 (2021).
Budge, G. E. et al. Chronic bee paralysis as a serious emerging threat to honey bees. Nat. Commun. 11, 1–9 (2020).
Bailey, L., Ball, B. V. & Perry, J. N. Honeybee paralysis: Its natural spread and its diminished incidence in England and Wales. J. Apic. Res. 22, 191–195 (1983).
Traynor, K. S. et al. Multiyear survey targeting disease incidence in US honey bees. Apidologie 47, 325–347 (2016).
Cersini, A., Antognetti, V., Conti, R., Velletrani, F. & Formato, G. First PCR isolation of Crithidia mellificae (Euglenozoa: Trypanosomatidae) in Apis mellifera (Hymenoptera: Apidae) in Italy. Fragm. Entomol. 47, 45–49 (2015).
Bordin, F. et al. Presence of Known and Emerging Honey Bee Pathogens in Apiaries of Veneto Region (Northeast of Italy) during Spring 2020 and 2021. Appl. Sci. 12, 2134 (2022).
Cilia, G., Tafi, E., Zavatta, L., Caringi, V. & Nanetti, A. The epidemiological situation of the managed honey bee (Apis mellifera) colonies in the Italian Region Emilia-Romagna. Vet. Sci. 9, 437 (2022).
Cersini, A. et al. First isolation of Kashmir bee virus (KBV) in Italy. J. Apic. Res. 52, 54–55 (2013).
Chen, G. et al. Seasonal variation of viral infections between the eastern honey bee (Apis cerana) and the western honey bee (Apis mellifera). Microbiologyopen 10, e1162 (2021).
Martín-Hernández, R. et al. Comparison of the energetic stress associated with experimental Nosema ceranae and Nosema apis infection of honeybees (Apis mellifera). Parasitol. Res. 109, 605–612 (2011).
Cilia, G., Garrido, C., Bonetto, M., Tesoriero, D. & Nanetti, A. Effect of Api-Bioxal ® and ApiHerb ® Treatments against Nosema ceranae Infection in Apis mellifera Investigated by Two qPCR Methods. Vet. Sci. 7, 125 (2020).
McMahon, D. P. et al. A sting in the spit: Widespread cross-infection of multiple RNA viruses across wild and managed bees. J. Anim. Ecol. 84, 615–624 (2015).
Radzevičiūtė, R. et al. Replication of honey bee-associated RNA viruses across multiple bee species in apple orchards of Georgia, Germany and Kyrgyzstan. J. Invertebr. Pathol. 146, 14–23 (2017).
Murray, E. A. et al. Viral transmission in honey bees and native bees, supported by a global black queen cell virus phylogeny. Environ. Microbiol. 21, 972–983 (2019).
Alger, S. A., Burnham, P. A., Boncristiani, H. F. & Brody, A. K. RNA virus spillover from managed honeybees (Apis mellifera) to wild bumblebees (Bombus spp.). PLoS One 14, e0217822 (2019).
D’Alvise, P., Seeburger, V., Gihring, K., Kieboom, M. & Hasselmann, M. Seasonal dynamics and co-occurrence patterns of honey bee pathogens revealed by high-throughput RT-qPCR analysis. Ecol. Evol. 9, 10241–10252 (2019).
Mondet, F. et al. Transcriptome profiling of the honeybee parasite Varroa destructor provides new biological insights into the mite adult life cycle. BMC Genomics 19, 1–19 (2018).
Smoliński, S., Langowska, A. & Glazaczow, A. Raised seasonal temperatures reinforce autumn Varroa destructor infestation in honey bee colonies. Sci. Rep. 11, 22256 (2021).
Ball, B. V. & Allen, M. F. The prevalence of pathogens in honey bee (Apis mellifera) colonies infested with the parasitic mite Varroa jacobsoni. Ann. Appl. Biol. 113, 237–244 (1988).
Fearon, M. L. & Tibbetts, E. A. Pollinator community species richness dilutes prevalence of multiple viruses within multiple host species. Ecology 102, e03305 (2021).
Cohen, H., Ponisio, L. C., Russell, K. A., Philpott, S. M. & McFrederick, Q. S. Floral resources shape parasite and pathogen dynamics in bees facing urbanization. Mol. Ecol. 31, 2157–2171 (2022).
Figueroa, L. et al. Bee pathogen transmission dynamics: Deposition, persistence and acquisition on flowers. Proc. R. Soc. B 286, 20190603 (2019).
Pinilla-Gallego, M. S., Ng, W. H., Amaral, V. E. & Irwin, R. E. Floral shape predicts bee-parasite transmission potential. Ecology 103, e3730 (2022).
Jagadish, S. V. K. et al. Implications of high temperature and elevated CO2on flowering time in plants. Front. Plant Sci. 7, 913 (2016).
Dudley, L. S., Arroyo, M. T. K. & Fernández-Murillo, M. P. Physiological and fitness response of flowers to temperature and water augmentation in a high Andean geophyte. Environ. Exp. Bot. 150, 1–8 (2018).
Li, X. et al. Tolerance and response of two honeybee species Apis cerana and Apis mellifera to high temperature and relative humidity. PLoS One 14, e0217921 (2019).
Peat, J. & Goulson, D. Effects of experience and weather on foraging rate and pollen versus nectar collection in the bumblebee, Bombus terrestris. Behav. Ecol. Sociobiol. 58, 152–156 (2005).
Locke, B. et al. Adapted tolerance to virus infections in four geographically distinct Varroa destructor-resistant honeybee populations. Sci. Rep. 11, 1–12 (2021).
McMahon, D. P., Wilfert, L., Paxton, R. J. & Brown, M. J. F. Emerging viruses in bees: From molecules to ecology. Adv. Virus Res. 101, 251–291 (2018).
Choi, N. R., Jung, C. & Lee, D.-W.W. Optimization of detection of black queen cell virus from Bombus terrestris via real-time PCR. J. Asia. Pac. Entomol. 18, 9–12 (2015).
Peng, W. et al. Host range expansion of honey bee Black Queen Cell Virus in the bumble bee. Bombus huntii. Apidologie 42, 650–658 (2011).
Higes, M. et al. The stability and effectiveness of fumagillin in controlling Nosema ceranae (Microsporidia) infection in honey bees (Apis mellifera) under laboratory and field conditions. Apidologie 42, 364–377 (2011).
Goblirsch, M. Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie 49, 131–150 (2018).
Fratianni, S. & Acquaotta, F. The Climate of Italy. World Geomorphol. Landscapes 29–38 (2017). https://doi.org/10.1007/978-3-319-26194-2_4/COVER.
Figueroa, L. L., Grincavitch, C. & McArt, S. H. Crithidia bombi can infect two solitary bee species while host survivorship depends on diet. Parasitology 148, 435–442 (2021).
Danforth, B. N., Minckley, R. L., Neff, J. L. & Fawcett, F. The Solitary Bees: biology, evolution and conservation. (Princeton University Press, 2019). https://doi.org/10.1515/9780691189321-004/HTML.
Cremer, S., Armitage, S. & Schmid-Hempel, P. Social immunity. Curr. Biol. 17, R693–R702 (2007).
Meunier, J. Social immunity and the evolution of group living in insects. Philos. Trans. R. Soc. B Biol. Sci. 370, 20140102 (2015).
Zhao, X. & Liu, Y. Current knowledge on bee innate immunity based on genomics and transcriptomics. Int. J. Mol. Sci. 23, 14278 (2022).
Koch, H. & Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. 108, 19288–19292 (2011).
Straub, L. et al. Do pesticide and pathogen interactions drive wild bee declines?. Int. J. Parasitol. Parasites Wildl. 18, 232–243 (2022).
Harmon-Threatt, A. Influence of nesting characteristics on health of wild bee communities. Annu. Rev. Entomol. 65, 39–56 (2020).
Evans, A., Llanos, J., Kunin, W. & Evison, S. Indirect effects of agricultural pesticide use on parasite prevalence in wild pollinators. Agric. Ecosyst. Environ. 258, 40–48 (2018).
Doublet, V., Labarussias, M., de Miranda, J. R., Moritz, R. F. A. & Paxton, R. J. Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 17, 969–983 (2015).
Chagas, D. B. et al. Black queen cell virus and Nosema ceranae coinfection in Africanized honey bees from southern Brazil. Pesqui. Veterinária Bras. 40, 892–897 (2021).
Gajda, A. M., Mazur, E. D., Bober, A. M. & Czopowicz, M. Nosema Ceranae Interactions with Nosema apis and Black Queen Cell Virus. Agriculture 11, 963 (2021).
Ryba, S., Titera, D., Schodelbauerova-Traxmandlova, I. & Kindlmann, P. Prevalence of honeybee viruses in the Czech Republic and coinfections with other honeybee disease. Biologia (Bratisl). 67, 590–595 (2012).
Zheng, H. Q. et al. Evidence of the synergistic interaction of honey bee pathogens Nosema ceranae and Deformed wing virus. Vet. Microbiol. 177, 1–6 (2015).
Benjeddou, M., Leat, N., Allsopp, M. & Davison, S. Detection of acute bee paralysis virus and black queen cell virus from honeybees by reverse transcriptase PCR. Appl. Environ. Microbiol. 67, 2384–2387 (2001).
Zhang, X. et al. New evidence that deformed wing virus and black queen cell virus are multi-host pathogens. J. Invertebr. Pathol. 109, 156–159 (2012).
Meeus, I., de Miranda, J. R., de Graaf, D. C., Wäckers, F. & Smagghe, G. Effect of oral infection with Kashmir bee virus and Israeli acute paralysis virus on bumblebee (Bombus terrestris) reproductive success. J. Invertebr. Pathol. 121, 64–69 (2014).
Manley, R., Temperton, B., Boots, M. & Wilfert, L. Contrasting impacts of a novel specialist vector on multihost viral pathogen epidemiology in wild and managed bees. Mol. Ecol. 29, 380–393 (2020).
Thu, H. T. et al. Prevalence of bee viruses among Apis cerana populations in Vietnam. J. Apic. Res. 55, 379–385 (2016).
Roberts, J. M. K. J., Anderson, D. D. L. & Durr, P. A. P. Absence of deformed wing virus and Varroa destructor in Australia provides unique perspectives on honeybee viral landscapes and colony losses. Sci. Rep. 7, 6925 (2017).
Funding
This study was supported by the project BeeNet (Italian National Fund under FEASR 2014–2020) from the Italian Ministry of Agriculture and Food Sovereignty and Forestry (MASAF).
Author information
Authors and Affiliations
Contributions
The study was designed by G.C. and L.B. Samples were collected by R.T., S.F., L.Z., R.R., M.G., M.B., C.B.B., E.C., R.C., M.D.S., M.P. and G.C. The bee specimens were identified by S.F., and the laboratory analyses were performed by R.T. and G.C. Data were analysed by R.T., L.Z., D.A.G., S.L. and G.C. R.T., L.Z., and G.C. wrote the first version of the manuscript, which was revised by all the other authors (S.F., R.R., M.G., D.A.G., S.L., M.B., C.B.B., E.C., R.C., M.D.S., M.P., M.Q., L.B. and A.N.). L.B. managed the project and the funding acquisition. All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tiritelli, R., Flaminio, S., Zavatta, L. et al. Ecological and social factors influence interspecific pathogens occurrence among bees. Sci Rep 14, 5136 (2024). https://doi.org/10.1038/s41598-024-55718-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-55718-x
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
