
Abstract
Considerable evidence suggests that musical education induces structural and functional neuroplasticity in the brain. This study aimed to explore the potential impact of such changes on word-reading proficiency. We investigated whether musical training promotes the development of uncharted orthographic regions in the right hemisphere leading to better reading abilities. A total of 60 healthy, right-handed culturally matched professional musicians and controls took part in this research. They were categorised as normo-typical readers based on their reading speed (syl/sec) and subdivided into two groups of relatively good and poor readers. High density EEG/ERPs were recorded while participants engaged in a note or letter detection task. Musicians were more fluent in word, non-word and text reading tests, and faster in detecting both notes and words. They also exhibited greater N170 and P300 responses, and target-non target differences for words than controls. Similarly, good readers showed larger N170 and P300 responses than poor readers. Increased reading skills were associated to a bilateral activation of the occipito/temporal cortex, during music and word reading. Source reconstruction also showed a reduced activation of the left fusiform gyrus, and of areas devoted to attentional/ocular shifting in poor vs. good readers, and in controls vs. musicians. Data suggest that music literacy acquired early in time can shape reading circuits by promoting the specialization of a right-sided reading area, whose activity was here associated with enhanced reading proficiency. In conclusion, music literacy induces measurable neuroplastic changes in the left and right OT cortex responsible for improved word reading ability.
Similar content being viewed by others


Introduction
Professional musicians' brains constitute a model of experience-dependent brain plasticity that has attracted increasing interest in neuroscience1,2,3,4,5,6. Several brain areas, including the anterior corpus callosum, cerebellum, primary motor area and auditory cortices, show differences in structure and size in musicians compared to non-musicians3. Higher demands for bimanual coordination and rapid information exchange appear to promote neural fiber growth7, or at least prevent neural tissue loss during synaptic pruning8. In particular, the volume of the cerebellum is larger in musicians than in non-musicians2,9,10 because of its role in regulating coordination, precise timing and accuracy of motor commands. Other structural and functional changes due to musical experience have been observed in somatosensory and motor cortex11,12,13,14, and along the auditory pathway: in the brainstem15, primary auditory cortex16, and higher-order auditory areas17,18.
These neuroplastic changes are associated with the multiple benefits of music education in terms of improvements in attentional, executive, coordination, motor, cognitive, and motivational skills (e.g.,19,20,21). Specifically for reading, the benefit would come from refined auditory processing and increased phoneme processing, rhythm and phonological awareness skills (e.g.,22,23) on the one hand, and improved ability to visually decode symbols and quickly translate them into meanings and gestures on the other. Several studies have shown how musical perceptual skills correlate with phonological awareness24 and reading skills, and may also be predictive of children's reading developmental trajectories25,26,27,28. For example, an interesting meta-analysis29 showed that students who received music education scored significantly higher on reading tests than control students. Again, Standley and Hughes30 and Register31 gave music lessons to 4–5 year old children and compared them with a control group of children of the same age who did not receive music lessons, and showed that music lessons at an early age contributed to improvements in pre-reading and writing skills. Swaminathan et al.32 observed a large group of 166 native and non-native English-speaking adults who had an average of two years of private music lessons outside of school and found a positive correlation between music practice and reading skills, as also found by Schellenberg and Weiss21.
Despite this body of knowledge, not much is known about the plastic changes in the brain that underlie improved reading skills, particularly with regard to the visual encoding of orthographic and symbolic information such as words and musical notation. Here, we tested the hypothesis that musical literacy (the ability to read music fluently) acquired at the age of 10 or earlier, and continued practice of this ability, would have a direct impact on the ability to read letters and words. This would occur through the development of a right hemispheric reading area (used for pentagram reading). In addition, the intensive training of attention and eye shifting involving V5 and the oculomotor area33 would also act as an enhancing and protective factor for reading ability.
A previous ERP study by Proverbio et al.34 compared the ability to identify notes and letters (embedded in musical bars and words, respectively) in groups of musicians and matched controls. The results showed that the orthographic N170 was larger for words in musicians than in controls. In addition, the N170 response showed a bilateral distribution in musicians during the processing of both notes and words, whereas it was strongly left-sided in controls, as expected. Source reconstruction showed that the fusiform (BA37) and inferior occipital gyri (BA18) were activated in both hemispheres in musicians (for both word and music processing), whereas orthographic processing was restricted to the left hemisphere in controls. This neural pattern was associated with an enhanced ability to recognise both notes and letters in musicians compared to controls. We concluded that the neural mechanisms of word reading can be modified by musical training in childhood (from the age of ~ 8 years). The evidence of right hemisphere involvement for a function normally lateralised to the left seems to be a consequence of the neuroplastic effects of musical training on reading ability. This hypothesis is supported by the findings reported by Li and Hsiao35, who investigated the effects of musical reading on reading in an English word and Chinese character naming task presented in different visual fields. They found an effect of musical literacy on word reading lateralization: in fact, musicians performed significantly faster than controls for words presented in the left visual field (right hemisphere). The literature consistently demonstrates that bilateral neural mechanisms, primarily relying on the right OT cortex, underlie the ability to read musical notation. In different paradigms, the so-called Visual Note From Area (VNFA) has been localised in the right transverse occipital sulcus, right occipital gyrus, right inferior occipital gyrus, right occipitotemporal junction, right fusiform gyrus and right superior parietal cortex36,37 and supramarginal cortices34,38,39,40,41,42,43,44,45. If music education and word reading skills are linked, and if the former is able to influence the development and performance of reading skills, then it can be expected that treatment based on learning music skills might contribute to the improvement of language skills in individuals who are deficient in these skills. Indeed, Register et al.46 and Habib et al.47 investigated whether music could be a strategy to improve reading skills in students with specific reading difficulties and found significant improvements in reading tests administered before and after music training.
If it were true that the development of a right-sided orthographic area (necessary for encoding spatial relations in pentagrams, as originally reported by Proverbio et al.34 was the cause of the better performance of musicians in orthographic tasks and reading, then we would expect not only a bilateral activation of orthographic areas in musicians (during word reading), but also their better performance in independent reading tests. In addition, the Visual Word Form Area (VWFA) should show less activation in poor readers than in good readers. The VWFA, located in the medial part of the left fusiform gyrus, is known to underlie the ability to recognise letters and words, being more sensitive to letter strings than to other pictorial stimuli48,49,50,51,52,53, while also being sensitive to sub-lexical properties such as word familiarity or frequency of use54,55,56. The development and specialisation of this area for the recognition of written words enables rapid reading by increasing the perceptual capacity for words and making it sensitive to recurrent features of the writing system57. In this respect, the N170 component of ERP is deemed as the electromagnetic representation of VWFA activity. Research has proven that the amplitude of the N170 response is higher when presented with letters or words instead of other objects. Moreover, this response is typically focused over the left occipito/temporal area55,58.
The present ERP study aimed to investigate the pattern of brain electrical activity in a sample of professional musicians and controls in response to words and musical notation. Right hemispheric involvement in word reading (in addition to music reading) was expected in areas contralateral to the VWFA in musicians, as previously demonstrated by Proverbio et al.34. In the present study, a parallel comparison was made between good and poor readers within the large sample of participants. We hypothesised: (1) that professional musicians have developed a specialised visual area over the right OT for reading music notation, the activity of which is reflected in a bilateral N170 response to notes (obviously larger than in controls); (2) that musicians also automatically activate the bilateral orthographic areas for word processing (whereas the N170 is purely left-sided in non-musicians); (3) that musicians were better readers than controls and showed larger amplitudes of N170 and P300 components; that good readers, regardless of musical ability, showed bilateral involvement of orthographic areas; (4) that N170 amplitudes correlated with reading ability, thus proving to be a reliable marker of reading ability. It is important to note that in order to prevent transfer of perceptual strategies across tasks, the musical notation and word experimental sessions were independent and separated in time (in different experimental blocks).
Few studies have previously compared N170 and P300 potentials elicited by written words (reflecting orthographic and selective attention processes, respectively) in musicians vs. non-musicians. Proverbio and co-authors34 found larger N170 and N250 responses (selection negativity) to written words in musicians compared to control readers, as well as faster response times (RTs) and higher accuracy in letter identification tasks, but no larger P300 responses. Li and coauthors35,44 found earlier RTs, higher accuracy and larger N170 responses (also over the right hemisphere) in musicians, in word naming tasks, but did not quantify P300 responses. Overall, there is no consolidated evidence that the P300 is larger in musicians during visual reading tasks, whereas it is commonly reported that the P300 is larger in musicians compared to non-musicians in auditory or speech listening tasks (e.g.59) due to neuroplastic changes in the auditory cortex.
Material and methods
Participants
Sixty right-handed, age-matched, healthy students took part in this study. They were half male and half female, matched for age and socio-cultural status, and all native Italian speakers. Specifically, the musicians were 34 individuals (17 females) aged 18–28 years (22.8; SD = 2.68). Inclusion criteria were being a professional musician, having graduated (BA or Master) from a music conservatory in the Lombardy region. They specialised in a variety of instruments: piano, guitar, opera singing, saxophone, oboe, violin, electric bass, bassoon, double bass, euphonium, transverse flute, organ, cello, trumpet and composition. Their average number of years of music study was 12.5 years (SD = 4.26), with an average age of acquisition (AoA) of 10 years, confirming that the musicians were truly experienced and professional. Four musicians were excluded due to excessive EEG artifacts. The final sample for ERP analyses consisted of 30 musicians (15 females) aged between 18 and 28 years (22.7; SD = 2.64). Their lateral preference was assessed by administering the Edinburgh Inventory. The musicians' mean laterality score was 0.75 (mean = 0.7479, min = 0.4286, max = 1.0, SD = 1.18).
Participants were preliminarily interviewed about their reading habits and musicians self-reported an average number of printed pages read (per year) = 1250, equivalent to approximately 3.5 books (including college textbooks). Controls were 26 graduated (BA or master) University student of non-musical faculties in the Lombardy area (21 females). They aged 19–27 years (22.12; SD = 1.82); their average laterality score was 0.80 (mean = 0.7999, min = 0.4300, max = 1.0, SD = 1.145). No control participant was discarded because of EEG artifacts. Non-musicians reported an average number of printed pages read (in a year) = 2915, equivalent to about 8 books (including college textbooks). An ANOVA performed on the laterality scores showed no effect of group (p = 0.26). A further ANOVA on the number of pages read in 1 year showed a significant effect of group (p < 0.0002), as expected. Inclusion criteria for the control group were: never having studied music (except for a few years in junior high school), not currently able to read music, not playing or singing, and not studying a musical instrument as a hobby.
Inclusion criteria for all 60 participants were that they had never had a psychiatric or neurological disease or injury, were not currently taking drugs or narcotics, and were not predisposed to epilepsy. All participants had (self-reported) normal or corrected-to-normal vision and hearing. No participant suffered or had ever suffered from a learning or reading disorder (e.g. developmental dyslexia, alexia, autism, ADHD, etc.). All participants gave written informed consent before taking part in the study and were unaware of the specific purpose of the research. The experiment lasted approximately 3 h and participants volunteered or received academic credit. All methods were performed in accordance with the relevant guidelines and regulations (Declaration of Helsinki). The project was approved by the Ethics Committee of the University of Milan-Bicocca (protocol number RM-2021-370).
Reading tests and questionnaires
The study consisted of two experimental sessions, separated by a few days to reduce subject fatigue: the first session consisted of the assessment of inclusion criteria, the collection of demographic data and the administration of the reading and laterality tests; the second session consisted of the recording of EGG/ERP signals. Subjects were recruited through announcements on the university's social networks. Volunteers first provided some preliminary information, such as age, degree, eventual academic level in music attained, age of acquiring the ability to play an instrument, primary instrument, secondary instrument, and identification of presumed right-hand preference. After recruitment, both musicians and controls participated in an initial meeting via a digital platform, and the video was recorded for data analysis with the participant's consent.
In this session, a reading battery (in Italian) was administered, consisting of word and nonword reading lists from the Battery for the Assessment of Developmental Dyslexia and Dysorthographia60 and a text from the VALS-Assessment of Reading and Writing Difficulties in Adulthood61. The word reading test consisted of 4 runs of vertically ordered familiar words (28 words per set), administered individually. The nonword reading test consisted of 3 sets of 16 pronounceable nonwords. The text reading consisted of a one-page narrative (taken from Stefano Benni's novel "The Bar Under the Sea") that had to be read quickly but with appropriate prosody. The order in which the tests were administered was randomised across subjects. Reading accuracy and speed were calculated for each test (word, pseudoword and text reading). Participants were asked to read the text displayed on the screen as quickly as possible, but as accurately and clearly as possible. Five warning signals preceded the start of the test. "Attention", "Three, Two, One", "Go". Reading times were calculated from the start of the participant's reading to the end of vocal production. After each performance, all participants received the same general positive reinforcement and thank you. Following the administration of the reading tests, participants were administered the Edinburgh Inventory Questionnaire to assess their lateral preference.
Stimuli and procedure
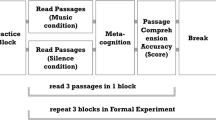
The stimuli consisted of 300 Italian words and 300 music bars of different length and complexity, presented in random order in the centre of a PC screen, approximately 114 cm from the subject's eyes (Fig. 1). The stimuli and procedure were the same as those used in the ERP study by Proverbio et al.34.

Time sketch of experimental procedure for the music and language conditions.
Words were typed in capitals in Arial Narrow and were 0° 30′ 11″ (1 cm) high and from 1° 15′ 27″ to 4° 31′ 37″ (2.5 to 9 cm) long. Music bars were 0° 45′ 16″ (1.5 cm) high and 4° 16′ 32″ (8.5 cm) long. Music bars varied in length from 4 to 8 notes, while words varied in length from 4 to 10 letters. Music fragments were selected from Mozart and Schumann real pieces for piano and violin. Two different experimental conditions were used: a note recognition task and a letter recognition task. Half of the participants completed the music task in the first half of the experiment and the orthographic task in the second half, while the order was reversed for the other half of the participants. In the orthographic task, 300 Italian words (half target, half non-target) were presented pseudorandomly in the centre of the screen. Stimuli lasted 1600 ms and the ISI ranged from 1000 to 1200 ms. All stimuli were matched for duration across trials and across target and non-target categories. Words were also matched for frequency of use (across target/non-target categories). Word frequencies were taken from a large online database of Italian words (ColFIS62).
The stimuli used as targets were 'mi', 'fa', 'sol', 'la' and 'si' of the middle piano octave (i.e. E4, F4, G4, A4 and B4) for the note recognition task and the letters B, G, L, M and S for the orthographic recognition task. At the beginning of each session, subjects were told what the targets were and which hand would be used to respond. Subjects were given both a verbal label and an enlarged visual representation of the isolated letter or note (within the pentagram), which remained in the subject's hand for free inspection until the experimental trial began. Participants sat comfortably in an acoustically and electrically shielded booth under scotopic luminance conditions in front of a high-resolution monitor placed outside the booth. They were asked to keep their gaze fixed on a 3 mm fixation point at the centre of the screen, minimising eye movements and avoiding any body movements. The task was to respond as quickly and accurately as possible by pressing a button on a joystick with their index finger each time they detected the target within the stimulus. The experimenter indicated the response hand, which alternated between runs, before the start of each sequence. Stimuli were presented in 12 runs of 50 stimuli each, half target and half non-target, randomly intermixed. The order of response hands was counterbalanced across participants, and the order of presentation of word and tone sequences was also randomised and counterbalanced. The two experimental sessions were preceded by two training sequences using exemplar stimuli that would not be repeated later in the experiment. Each sequence began with the appearance of the words 'Ready', 'Attention' and 'Go' written in block letters and ended with the words 'Thank you'.
EEG recordings and data analysis
EEG data were recorded using a standard 128-electrode EEG cap placed according to the 10–5 international system63 with EEProbe v2.2 software (ANT Neuro, Hengelo, The Netherlands) at a sampling rate of 512 Hz (bandpass 0.16–70 Hz). Horizontal (hEOG) and vertical (vEOG) eye movements were also recorded. Linked mastoids were used as reference lead. Electrode impedance was kept below 5 KOhm. Computerised artefact rejection and manual eye inspection were used to remove EEG segments contaminated by eye artefacts (saccades and blinks), muscle-related potentials or amplifier blockages. The computerised criterion for rejecting artefacts was a peak-to-peak amplitude exceeding 50 μV. EEG epochs were synchronised with stimulus onset. Evoked response potentials (ERPs) were averaged off-line from 100 ms before to 1500 ms after stimulus onset, and an off-line filter (band-pass 0.16–30 Hz) was applied to the ERPs. Data from reading tests, behavioural responses and EEG/ERP recordings were analysed.
The results of the reading tests were normed following the protocols of the battery for the assessment of dyslexics and dysgraphias60 and the VALS test61. The sample was divided into 'good readers' and 'poor readers' groups based on their average reading speed (number of syllables read per second), both within each group (musicians vs. controls) and considering the whole population. Correlation analyses were performed between the scores on the three reading tests, between word reading speed (syllables per second = reading ability) and TRs to target words, and between reading ability and the amplitude of N170 and P300 responses to target words.
ERP components were identified in the time window and scalp location where and when they reached maximum amplitude and according to previous literature. The mean area amplitude of the N170 was quantified at occipito-temporal sites (PPO9h-PPO10h). The N1 peak was defined as the most negative value between 170 and 210 ms. The mean area amplitude P300 was quantified in the 600–800 ms time window for notes and in the 450–650 ms time window for words at centro/parietal sites (CP1-CP2).
For each ERP component, repeated-measures ANOVAs were applied to individual ERP amplitudes recorded in musicians and controls, as a function of reading proficiency (poor vs. good readers), and stimulus type. In details, between-group factors were: Proficiency (poor vs. good readers) and Group (musicians vs. controls). Within-group factors were: stimulus type (notes vs. words), attention (non-targets, targets), hemisphere (left, right).
Further repeated-measures ANOVAs were performed on response times (RTs) and accuracy data. Between-group factors were: proficiency and Group (Musicians vs. Controls). Within-group factors were: stimulus type (note vs. word), targetness (non-target, target) and response hand (left, right).
Tukey and Fisher post-hoc comparisons were carried out to test differences among means. The effect size for the statistically significant factors was estimated using partial etasquared (ηp2) and the Greenhouse–Geisser correction was applied to account for non-sphericity of the data. All the ANOVAs were performed using Statistica software (version 10) by StatSoft.
Spearman Rho correlation analyses were performed between individual values of N170 recorded in response to words in the left and right hemispheres and proficiency as measured by syllable/sec reading speed. Furthermore, response times were correlated with the amplitude of the P300 response to target words as recorded at centro/parietal CP1-CP2 electrode pair. Correlation analyses also performed to assess the relationship between reading performances on the three reading scales.
Source reconstruction
Low-resolution electromagnetic tomography (LORETA) was performed on ERP waveforms at the N170 latency (170–210 ms) during selective attention to letters or notes in good and poor readers, musicians and controls. Further LORETAs were performed on ERP waveforms at the P300 latency (450–650 ms) during selective attention to letters, in musicians and controls. LORETA is an algorithm that provides discrete linear solutions to inverse EEG problems. The resulting solutions correspond to the 3D distribution of neuronal electrical activity that has the most similar orientation and strength between neighbouring neuronal populations (represented by adjacent voxels). This study used an improved version of this algorithm, the standardised weighted (sw)LORETA64. This version, referred to as swLORETA, incorporates a singular value decomposition based source field weighting method. The source space properties included a grid spacing (the distance between two computation points) of five points (mm) and an estimated signal-to-noise ratio, which defines the regularisation, with a higher value indicating less regularisation and therefore less blurring of the results, of three. Using a value of 3–4 for the calculation of SNR in Tikhonov's regularisation results in superior accuracy of solutions for each inverse problem evaluated. swLORETA was run on the grand-averaged group data to identify statistically significant electromagnetic dipoles (p < 0.05) where larger magnitudes correlated with more significant activation. As part of the LORETA analysis, the data were automatically referenced to the mean reference. A realistic boundary element model (BEM) was derived from a T1-weighted 3D MRI dataset. This was achieved by segmenting the brain tissue. This BEM model consisted of a homogeneous compartment with 3,446 vertices and 6,888 triangles. Advanced Source Analysis (ASA) uses a realistic head model consisting of three layers (scalp, skull and brain) and is generated using the BEM. This realistic head model consists of a set of irregularly shaped boundaries and the conductivity values for the compartments between them65. A number of points connected by planar triangles are used to approximate each boundary. The triangulation results in a more or less evenly distributed mesh of triangles, depending on the chosen grid spacing. A smaller value for the grid spacing results in a finer mesh and vice versa. For the three-layer realistic head model mentioned above, it is assumed that the segmentation includes current generators of brain volume, including both grey and white matter. The regional conductivities of the scalp, skull and brain were assumed to be 0.33, 0.0042 and 0.33 respectively. Source reconstruction solutions provided by the Montreal Neurological Institute were projected onto the 3D MRI of the Collins brain. SwLORETA was performed on the grand mean group data to identify statistically significant electromagnetic dipoles (p < 0.05), where larger magnitudes correlated with more significant activations. Probabilities of source activation based on Fisher's F-test were provided for each independent EEG source, with values reported on a "unit" scale in nA (the larger the value, the more significant). Different colours indicate different strengths of electromagnetic signals. Both segmentation and head model generation were performed using ASA software (ANT, Enschede, The Netherlands). Topographic distributions of surface voltage were generated by mapping isopotential lines, derived by interpolating voltage values between surface electrode sites at specific time latencies, onto a colourimetric scale.
Results
Reading tests
Table 1 shows the results of the three reading tests for musicians and controls. The participants were divided into two subgroups, 'poor readers' (1st to 50th percentile) and 'good readers' (51st to 100th percentile), on the basis of the performance obtained, and in particular the speed of reading syllables per second for words. It is important to note that the participants did not suffer from dyslexia or alexia, nor did they have any reading or learning disorder; they were normative readers, as evidenced by their performance (see Table 1).
The negligible number of errors made when reading words or non-words confirms that all participants were healthy, normo-typical readers. As slowing down, uncertainty or repetition were also considered errors, a few errors were observed when reading the whole page of an unfamiliar text with the constraint of being very fast. The slightly higher number of reading inaccuracies for text reading among musicians may be related to the significantly lower number of book pages per year that they self-reported. In addition, the slightly higher fluency of the controls compared to the normative data (especially in text reading) could be due to their cultural status (being university graduates rather than first year students).
Correlation analyses performed to assess the relationship between performance on the three scales showed a significant correlation between: word reading and non-word reading (r = 0.56, p < 0.05); between word reading and text reading (r = 0.73, p < 0.05); between non-word reading and text reading (r = 0.60, p < 0.05) (Fig. 2, left), indicating the reliability of the tests.

Correlation matrices displaying the relationship between the reading performances acquired in the three reading tests: word reading, nonword reading, and text reading in musicians (left column) and controls (right column).
The lower part of Table 1 shows the results of the three reading tests for the controls (non-musicians). On the basis of the performance obtained, and in particular the speed of reading syllables per second for words, the participants were divided into two subgroups of "poor readers" (1st to 50th percentile) and "good readers" (51st to 100th percentile). Correlation analyses performed to assess the relationship between performance on the three scales showed a significant correlation between: word and non-word reading (r = 0.58, p < 0.05); word and text reading (r = 0.58, p < 0.05); non-word and text reading (r = 0.66, p < 0.05) (Fig. 2, right).
An ANOVA performed on the whole sample to compare reading speed in the two subgroups revealed the strong effect of musicianship [F(1, 52) = 4.5236, p = 0.038], with musicians reading faster than controls regardless of proficiency (Fig. 3). The significance of the proficiency factor [F(1, 52) = 72.7, p < 0.00001] indicated faster reading times for good readers than for poor readers, regardless of stimulus type.

The data for reading tests are presented here, indicating reading speed for musicians and controls in comparison, for both good and poor readers. Additionally, reading speed (in syllables per second) is plotted as a function of stimulus type (word, non-word or text), and reading speed is compared between good and poor readers for each stimulus type.
The significance of stimulus type [F(2, 104) = 443, p < 0.00001, ε = 0.999] indicated faster reading times for words than for nonwords and for text than for words. In summary, musicians were more fluent readers than controls, regardless of ability level and stimulus type. This effect is rather surprising in view of the self-reported reading habits, which indicated that controls read twice as many books (in text) as musicians in one year. However, the number of pages read by the musicians did not take into account the number of pages of sheet music, and thus underestimated the massive attention-eye shifting exercise involved in reading sheet music, which is inherent in being a professional musician.
Behavioural data
Accuracy
The ANOVA performed on the percentages of correct responses during the EEG recording showed the significance of the group factor [F (1, 52) = 27.5, p < 0.000003; ηp2 = 0.35] with higher accuracy in musicians than in controls regardless of proficiency and target type (MUS = 98.5%, SD = 0.67; CON = 93.3%, SD = 0.7). The further significance of target type factor [F (1, 52) = 37.56, p < 0.000001, ε = 1; ηp2 = 0.42] indicated a higher accuracy for letters (98.78%, SD = 0.1) than notes (92.98%, SD = 0.96). The significant interaction of group x target type [F (1, 52) = 28.82, p < 0.000002, ε = 1; ηp2 = 0.36] showed a much lower accuracy for notes than for letters in controls (notes = 87.87%, SD = 1.41; letters = 98.82%, SD = 0.14; p < 0.0001), no difference in accuracy for the 2 target types in musicians (notes = 98.1%, SD = 1.29; letters = 98.82%, SD = 0.14), and no difference in accuracy with letters between groups (musicians = 98.82%, controls = 98.74%, SD = 0.16).
Reaction times (RTs)
The ANOVA performed on the mean response times during the EEG recording showed the significance of group factor [F (1, 52) = 19.83, p < 0.00005; ηp2 = 0.28] with faster RTs in musicians (665.5 ms, SD = 13.2) than in controls (753.8, SD = 14.4). Further significance of proficiency factor [F (1, 52) = 6.15, p < 0.016; ηp2 = 0.11] showed faster RTs in good readers (684.8 ms, SD = 14.4) than in poor readers (733.5 ms, SD = 13.27), regardless of target type. The factor target type also yielded significance [F (1, 52) = 430, p < 0.00001; ε = 1; ηp2 = 0.89] with faster RTs to letters (564 ms, SD = 8.79) than notes (854.4 ms, SD = 14.6). The ANOVA also showed the significant interaction of group x target type [F (1, 52) = 21.13, p < 0.00003; ε = 1; ηp2 = 0.29]. Post-hoc comparisons showed no difference in the groups’ mean RTs to letters, although musicians tended to be faster than controls in word reading, and musicians showed a strong advantage over controls in the note detection condition (see Fig. 4, left).

(Left) RTs analyses showed how good word readers were also good at detecting notes, regardless of musicianship. (Right) Scatterplot depicting the significantly negative correlation between the results of the three reading tests and word response times in all participants.
Correlation analyses were carried out between the reading test scores (word reading proficiency) and word response times in the experimental task. The two variables were found to be negatively significantly correlated, (Rho = − 0.4476, p < 0.05 in the whole sample), as can be seen in Fig. 4 (right). The correlation was more significant for musicians (Rho = − 0.56, p < 0.05) than for controls (Rho = − 0.4, p < 0.05).
Electrophysiological data
N170 component
The ANOVA performed on the amplitudes of the N170 component in the time window between 170 and 210 ms at PPO9h and PPO10h electrodes for good and poor readers (musicians vs. controls, respectively) showed the significance of the group factor with larger N170 responses recorded in musicians (− 4.11 µV, SE = 0.54) than in controls (− 0.50 µV, SE = 0.58), regardless of stimulus type or proficiency [F (1, 52) = 20.75, p < 0.00003; ηp2 = 0.28]. The ANOVA also revealed the stimulus type factor [F (1, 52) = 58.1, p < 0.00001; ε = 1; ηp2 = 0.53], with larger responses to words (− 3.83 µV, SE = 0.38 than to notes (− 0.77 µV, SE = 0.50), as shown in the ERP waveforms of Fig. 5. The interaction of group factor x stimulus type was also significant [F (1, 52) = 11.7, p < 0.001; ε = 1; ηp2 = 0.50]. Post-hoc comparisons showed significantly larger N170 responses to notes in musicians than controls (p < 0.0003), and significantly larger responses to words in musicians than controls (p < 0.02), regardless of proficiency (see graph in Fig. 6 for means and SE values). There was a significant stimulus x hemisphere interaction [F (1, 52) = 20.5, p < 0.00004; ε = 1; ηp2 = 0.28]. Post-hoc tests revealed a strong left-hemisphere asymmetry for the N170 for words (left = − 4.84 µV, SE = 0.46, right = − 2.83 µV, SE = 0.41) and a tendency for the N170 to be larger over right than left sites for notes (left = − 0.39 µV, SE = 0.57; right = − 1.16 µV, SE = 0.55).

Grand-average ERP waveforms recorded at left and right occipito/temporal sites in musicians vs. controls as a function of their reading proficiency. Poor readers showed a smaller right-sided hemispheric N70 response, especially if non-musicians.

(Top) Left: Interaction between musicianship and stimulus type. Mean amplitude values of N170 response recorded in the two groups of participants as a function of stimulus type (notes vs. words). Right: Interaction between targetness x hemisphere x proficiency. Mean amplitude values of N170 response recorded in the two groups of poor and good readers over the left and right occipito/temporal areas, in response to targets and non-targets. (Middle) Grand-average ERP waveforms recorded at left and right occipito/temporal sites in controls in response to target vs. non-target stimuli as a function of their reading proficiency. (Bottom) Correlational analyses between N170 amplitudes in response to words and word reading speed (syl/sec) in controls.
The further interaction of targetness x hemisphere x proficiency [F (1, 52) = 5.62, p < 0.001; ε = 1; ηp2 = 0.10]. Post-hoc comparisons showed an overall larger N170 in the left than in the right hemisphere, in good than in poor readers regardless of hemisphere, and a proficiency effect in the right hemisphere, with larger responses to targets over the right hemisphere in good than in poor readers, regardless of musicianship (see graph in Fig. 6 for means and SE values). However, the further interaction of musicianship x targetness x hemisphere x proficiency [F (1, 52) = 20.5, p < 0.00004; ε = 1; ηp2 = 0.12] showed that this effect was more true for controls (p < 0.001), as musicians showed a bilateral N170 response in all conditions, except for a tendency for poor readers to show a larger N170 to non-targets over the left than the right hemisphere (RH, p = 0.056). The topographic maps in Fig. 7a show the scalp distribution of the reading-specific N170 response, and clearly show the interaction between proficiency and hemispheric lateralization of the N170, with good readers also showing a N170 focus over the RH.

(Left) Isocolour topographical maps of N170 voltage distribution as recorded in good and poor readers (regardless of musicianship), in response to notes (upper row) and words (lower row). (Right) Same but recorded in the P300 range.
Correlation analyses were performed between individual values of N170 recorded in response to words in the left and right hemispheres and proficiency as measured by syllable/sec reading speed. In controls, there was no correlation (Rho = 0.07) between the two measures for the left hemisphere and an inverse correlation for the right hemisphere (the more negative the N170 response, the higher the proficiency; Rho = − 0.40, p < 0.05).
P300 component
The ANOVA performed on the amplitudes of the centro/parietal P300 component recorded in the 450–650 ms time window at CP1 and CP2 sites showed the significance of stimulus factor [F (1, 52) = 26.7, p < 0.00001; ε = 1; ηp2 = 0.34], with larger P300 amplitudes to words (4.90 µV, SE = 0.45) than notes (2.39 µV, SE = 0.38) in all participants, regardless of proficiency or musicianship (Fig. 7, right). Also significant was the group x proficiency interaction [F (1, 52) = 4.61, p < 0.0036; ε = 1; ηp2 = 0.08]. Post-hoc comparisons showed larger P300 recorded in good (5.0 µV, SE = 0.62) than poor musician readers (3.0 µV, SE = 0.66, p < 0.03), but no difference between the P300 recorded in good (2.84 µV, SE = 0.69) than in poor (3.74 µV, SE = 0.69) control readers. Post-hoc comparisons also showed larger P300 responses recorded in good musicians vs. good control readers (p < 0.02), but no difference between P300 responses recorded in poor musicians vs. poor controls. The ANOVA also revealed the significance of the stimulus x group interaction [F (1, 52) = 8.1, p < 0.006; ε = 1; ηp2 = 0.14]. Post-hoc comparisons showed larger P300 to words in musicians (5.95 µV, SE = 0.51) than in controls (2.72 µV, SE = 0.55, p < 0.01), as can be seen from the ERP waveforms in Fig. 8 (left). The stimulus targetness factor was also found to be significant [F (1, 52) = 36.5, p < 0.00001; ε = 1; ηp2 = 0.41], with larger P300s to target (4.46 µV, SE = 0.38) than non-target stimuli (2.84 µV, SE = 0.34). The significant interaction of targetness x group [F (1, 52) = 4.27, p < 0.04; ε = 1; ηp2 = 0.08] indicated the P300 was larger to targets than to non-targets in both groups, with a tendency for the P300 to be larger to targets in musicians than in controls (p = 0.06). Finally the ANOVA revealed the significance of stimulus type x targetness [F (1, 52) = 17.1, p < 0.0001; ε = 1; ηp2 = 0.25]. Post-hoc comparisons showed larger P300s to target than non-target stimuli, regardless of stimulus type, but with a larger effect for words (target = 6.21 µV, SE = 0.49; non-target = 3.61 µV, SE = 0.48; p < 0.0001) than for notes (target = 2.71 µV, SE = 0.43; non-target = 2.07 µV, SE = 0.39; p < 0.04). Correlation analyses between individual amplitude values of the P300 component in response to target words and word reading speed (syllables per second) showed the significance of the relationship in both musicians and controls (r = 0.412, p < 0.05), as can be seen in Fig. 8 (right).

(Left) Grand-average ERP waveforms recorded at midline central, parietal and left and right occipito/temporal sites, in musicians and controls in response to notes and words when target or non-target. (Right). Correlation between P300 amplitude values in response to target words and reading proficiency for the whole sample of participants.
Source reconstruction
N170 swLORETA
Eight swLORETA (standardised weighted low resolution electromagnetic tomography) source reconstructions were performed on the scalp-recorded voltage of N170 component (170–210 ms) as recorded in groups of poor or good readers, musicians or controls, in response to notes or words. The inverse solutions are shown in Figs. 9 and 10, while Table 2 lists the corresponding active electromagnetic dipoles according to the specific comparison.

Axial brain sections showing the location and strength of electromagnetic dipoles explaining the surface voltage of N170 response (170–210 ms), in musicians vs. controls, in good vs. poor readers, as a function of stimulus type (notes vs. words).

Coronal, axial and sagittal brain sections showing the location and strength of electromagnetic dipoles explaining the surface voltage of N170 response (170–210 ms) to words, in musicians as a function of their reading proficiency (good vs. poor readers). A higher reading proficiency was associated with a right-hemispheric involvement of OT cortex in reading.
Good vs. poor readers
Electromagnetic dipoles significantly active during word processing for good readers were the left fusiform gyrus (BA19) and the right middle (BA21) and inferior (BA20) temporal gyri (See Fig. 10). In contrast, the most active dipoles in poor readers were the left middle occipital gyrus (MOG, BA37), and no other right-sided visual regions.
Compared to good readers, poor readers had fewer active regions, not including the dorsolateral (BA9) and superior, left and right frontal (BA6) prefrontal cortex. Poor readers also showed significantly lower activations than good readers in areas involved in orthographic encoding (left BA37/19), reading (BA35), executive systems and attention (BA10 superior, BA10 middle), response selection (BA23), and eye movements (frontal eye fields FEF, BA8), as shown by the significance of non-parametric tests (Sign test: z = 2.04, p = 0.04, Wilcoxon test: z = 2.2, p = 0.028).
Electromagnetic dipoles that were significantly active during note processing for good readers were the right MOG (BA19) and for poor readers the right precuneus. Compared to good readers, poor readers had significantly fewer active regions, not including regions involved in shifting attention and eye movements (SFG, BA8), auditory processing in sensory (BA41) and associative (BA38) areas, executive systems, working memory and attention (bilateral BA 6, 9, 10, 11). Poor readers also showed significantly less activation than good readers in the cingulate gyrus (BA24) and SFG (BA6).
Musicians vs. controls
The electromagnetic dipoles that were significantly active during word processing for musicians were the left fusiform gyrus (FG BA19) and the right middle (BA21) and inferior (BA20) temporal gyrus (Fig. 10). In contrast, the most active visual dipoles in controls were the left middle occipital gyrus (MOG, BA37), and the right FG BA20. Compared to musicians, controls showed significantly weaker activations in areas involved in orthographic analysis (left FG BA19/37 and right FG BA20) reading (BA35 and BA38), response selection (BA23), executive systems, working memory and attention (i.e., left SFG BA10), right MFG (BA6), right dorsolateral prefrontal cortex (BA9, left superior and middle frontal gyri (BA10/11)), as demonstrated by the statistical significance of non-parametric tests applied to the observed magnitudes (Sign test: z = 2.0, p = 0.045, Wilcoxon test: z = 2.55, p = 0.01).
Electromagnetic dipoles that were significantly active during note processing in musicians were the right MOG BA37 and the left MOG BA19, while in controls it was the right precuneus BA7 (Fig. 9). Compared to musicians, controls had significantly fewer active regions, not including regions involved in auditory processing (Heschl gyri BA41), shifting of attention and eye movements (FEF BA8), executive systems, working memory and attention (right superior and middle frontal gyri, BA9 and BA11). Compared to musicians, controls showed significantly weaker activations in the superior temporal gyrus (BA22) and in the left and right superior frontal gyrus (BA6 and BA10), as shown by the significance of non-parametric tests (Sign test: z = 1.8, p = 0.07, Wilcoxon test: z = 2.02, p = 0.04).
P300 swLORETA
Two swLORETA source reconstructions were performed on the scalp-recorded voltage of P300 component (450–650 ms) as recorded in groups of musicians and non-musicians during word processing. Table 3 lists the corresponding active electromagnetic dipoles found in the two groups of readers.
Compared to musicians, controls had significantly fewer active regions, not including right hemispheric orthographic areas (BA18 and BA21) and regions involved in target selection and non-target suppression (the cingulate and posterior cingulate cortices BA 30 and BA23), as well as in shifting of attention and eye movements (FEF, BA8). Compared to controls, musicians showed significantly stronger cerebral activations in the left supramarginal gyrus (BA40) devoted to visual word recognition and in the left VWFA.
Discussion
The main aim of the present study was to investigate the neural correlates of note and word reading in people with different music proficiency to investigate how musical skills can influence and modify word reading ability and mechanisms. To this end, a sample of graduate non-musicians and musicians with a degree or diploma from a conservatoire was recruited. A word, non-word and text reading task was first administered to the participants. Event-related potentials were recorded during a letter/note detection task. Preliminary reading tests, showed how musicians were more proficient in language reading than controls. They were able to pronounce a higher number of syllable/sec than controls in all reading tests (word, non-word and text), regardless of the instrument played. Reading tests showed to be highly correlating among each other and with response times to target words recorded in the experimental session, which supports their reliability, and that of experimental paradigm in assessing reading proficiency. The higher reading proficiency of musicians, expected on the basis of previous literature (e.g.,24,25,26,27,28,29,30,31,34), was somewhat surprising given the self-reported reading habits, according to which musicians were used to read less than half as many books per year as non-music students. However, these estimates did not take into account the large number of musical scores that professional musicians read over the course of a year, which, if not full of words, are rich in symbols and equally stimulate reading, attention and ocular mechanisms44,66. Based on reading tests, the sample was further subdivided into poor and good readers, who coherently showed slower RTs to words and notes in the former and faster RTs to words and notes in the latter. The fact that word ability predicted the ability to detect notes embedded in musical bars suggests the existence of a common reading mechanism that benefits from musical literacy34.
ERP data showed larger orthographic N170 responses to words than to notes, larger N170 responses in good readers than in poor readers, and in musicians than in controls. The N170 component was found to be larger for word than for note recognition. These results are consistent with empirical evidence confirming the role of the occipitotemporal N170 in reflecting word recognition processes67,68. An interesting finding from the analysis of the N170 component is the significant interaction of the stimulus factor with the hemisphere factor. From the results, it would appear that there was not much difference in hemispheric activation between notes and words in the right hemisphere. On the contrary, the literature on the orthographic N170 provides evidence demonstrating that the processing of the orthographic properties of linguistic stimuli takes place in the VWFA, located in the left inferotemporal cortex69,70,71,72.
In this study, analyses showed a strong left-hemispheric asymmetry for N170 during word processing and a tendency for N170 to being larger over right than left sites to notation. The further interaction of targetness x hemisphere x proficiency showed overall larger N170s in the left than in the right hemisphere, in good than poor readers regardless of hemisphere, and a proficiency effect on the right hemisphere, with larger response to targets over the right hemisphere in good than poor readers, regardless of musicianship. This suggests the key role of right hemispheric specialization in enhanced reading skills. Again, the interaction of musicianship x targetness x hemisphere x proficiency showed that this effect was more pronounced for controls since musicians already showed a bilateral N170 response in all conditions, which correlated with their increased reading skills.
Correlation analyses performed between individual values of N170 to words and syllable/sec reading speed showed no correlation in controls over the left hemisphere, but a significant inverse correlation over the right hemisphere (the more negative the N170 response, the higher the reading proficiency). This finding further supports the causal hypothesis that an intensive training and specialization of the right orthographic area might lead to a superior performance (in good vs. poor readers, and in musicians vs. controls). This finding may suggest that early musical literacy in reading music does indeed lead to right hemisphere involvement in visual word processing. Right-hemisphere involvement in reading music notation is a robust notion widely supported by the previous literature34,38,39,40,41,42,43,44,45. The evidence for a difference in activation of the right fusiform gyrus in response to words between good and poor readers highlights the neuroplastic effects of music education on visual word recognition ability. The swLORETAs applied to N170 component, which were carried out on groups of musician and control readers, showed, among other things, an effect of musical literacy and proficiency on the lateralisation of the reading neural mechanism.
In addition, the results provide evidence for a reduced ability to discriminate stimuli in poor readers, as evidenced by a reduced amplitude of the P300 component, as well as slower RTs and reading times. This may be due to difficulties in sustaining attention, a poorer ability to shift attention and focus rapidly in search of the target, and a poorer ability to detect targets visually. A positive correlation was also found between P300 amplitude and reading speed (number of syllables per second). From these data, it can be concluded that shorter reading times are associated with a greater P300 amplitude, which reflects discrimination certainty73, due to a greater ability to discriminate and classify the stimulus, as well as attentiveness74,75,76 in musicians, as well as in good than poor readers. Indeed, overall, P300 response to words was found of greater amplitude in musician than control readers. Although the psychophysiological literature is lacking previous evidence of enhanced P300 to written words in musician as compared to non-musicians (see the previously mentioned ERP studies34,35,44) this effect might suggest enhanced perceptual and selective attention abilities, linked to the neuroplastic reading-related changes, such as the development of a right orthographic area (shown by N170 and behavioural data). Additionally, the constant eye movements involved in reading music might strengthen attentional shifting and reading skills. Both hypotheses were supported by the results of the source reconstruction applied to P300 potentials. The data showed stronger brain activity in musicians compared to control readers in regions involved in visual word recognition (i.e., the left supramarginal gyrus77), orthographic processing (the left B37 and bilateral BA18 and BA 21 areas), target selection (the posterior cingulate cortex78), and ocular/attentional shifting (FEF, BA879). This pattern of results further suggests that musical literacy improves visual word recognition by enhancing orthographic and letter recognition areas over posterior brain areas, and by providing more efficient attentional selection mechanisms. Specularly, the reduced amplitude of the P300 in poor readers could be due to a greater difficulty in detecting the targets, due to the inefficiency of the orthographic analysis mechanism (insufficient VWFA activity and lack of bilateral activation), or due to their difficulty in maintaining sustained attention. There is some empirical evidence in the literature for the presence of a reduced amplitude of the P300 component in individuals with attentional disorders80,81. For example, Papagiannopoulou and Lagopoulos82 found a reduced amplitude of the P300 component in children with dyslexia and impaired attentional resource allocation. In light of this evidence, the data from the present study regarding the lower P300 amplitude in subjects with lower reading ability appears to be consistent with the longer response times they showed in the note and word recognition tasks.
Word processing
In detail, during word processing, larger N170 magnitudes from VWFA sources (left fusiform/MOG BA19 and BA37) were found in good vs. poor readers (reading ability) and in musicians than in controls (musicianship and derived enhanced skills). On the other hand, a reduced activation of the VWFA was found in both poor readers and controls, suggesting less efficient orthographic processing. This is consistent with the poorest text reading performance of poor readers and their less efficient ability to detect target letters (as well as target notes). The reduced activation of the left FG in poor readers is strongly consistent with its role in reading performance.
There is literature to suggest that individuals with dyslexia have reduced VWFA activation83,84. At this regard, Maisog et al.85 conducted two meta-analyses to explore the neurological foundation of developmental dyslexia. The study found that dyslexic individuals have hypoactivity in the left extrastriate cortex, which is consistent with the results of the present investigation. The hypoactivation of the visual word form area (VWFA) and the homologous area in the right hemisphere among individuals with poor reading skills was concomitant with a diminished and less extensive neurometabolic activation in the prefrontal region. The reduced activation of the latter possibly arises from a resulting decrease in attentional capacity and working memory86, which may adversely affect the performance of weaker readers compared to stronger ones. There is much evidence to suggest that injury to the left occipitotemporal area can lead to impaired reading ability, including pure alexia87,88 or transient alexia89. Consistently, developmental dyslexia (in the absence of brain damage) is instead associated with insufficient or atypical activation of the left occipito-temporal ventral cortex activation for words88,90,91,92,93,94,95,96. This atypical or insufficient activity would result in smaller amplitudes of the left occipital/temporal N170 in response to words that symbols91,97, as compared to controls.
Most notably, the most powerful N170 source (after the VWFA) in good readers and in musicians was the right OT (middle and inferior temporal gyrus, BA20 and 21), that was not find active in poor readers and in controls. It is possible to relate the existence of the right orthographic area active during letter search to the development of a right-sided VNFA in musicians, and to the acquired enhanced reading skills.
Statistical analyses carried out on source reconstruction data showed that brain activations were more extended and more intense in good than poor readers, and in musicians than in controls. The areas of higher activity during word reading in good readers were: the parahippocampal gyrus (BA35), strongly connected with the VWFA98,99 and involved in reading100 and visuo-spatial processing101. Also more active were: the right middle temporal/angular gyrus (BA39) involved in grapheme-to-phoneme conversion and phonological processing102,103; the right cingulate cortex (BA23) engaged in target selection104; bilateral anterior brain areas, including the premotor cortex (BA6), dorsolateral prefrontal (BA9), middle and superior frontal cortices and frontal eye fields (BA8), know to control attentional selection, sustained attention, and attentional and ocular shifting105,106,107,108,109.
Note processing
During note processing, skilled readers and musicians showed the highest activation in the right middle occipital gyrus (BA19), implying this region as a putative VNFA. The literature has consolidated evidence that music reading particularly engages a specialized area of the right hemisphere of the occipito/temporal region. This area has been identified in different studies as the right transverse occipital sulcus, right occipital gyrus, right inferior occipital gyrus, right occipito/temporal junction, and right fusiform gyrus. The supporting studies notably include works by Sergent et al.38, Schön et al.39, Mongelli et al.40, Meinster et al.41, Proverbio et al.34,42, Wond and Gauthier43, Li et al.44, and Nakada et al.45. However, the same mechanism was not observed in controls, because of a lack of specialization and literacy. Musical notation reading in non-musicians (and in poor-readers) was subserved by an occipito/parietal area (the right cuneus, BA19), an area devoted to the processing of spatial properties of objects. It should be noted that in novice readers, note identification relies heavily on spatial analysis of note positions on the five pentagram lines.
A significant disparity between musicians and controls was observed in the neural network activated during the processing of musical notation. The network was substantially more extensive in musicians than in controls, encompassing the right premotor cortex, as well as the right and left dorsolateral prefrontal and superior frontal cortices, which may be associated with attentional maintenance, working memory, attentional focus, and attentional shifts.
It is worth noting that significant activation of frontal eye fields (BA8), which are involved in ocular movement and shifting of attention, was found only in musicians (and in good readers). Similarly, an activation of Heschl gyri (primary auditory area, BA41) was associated with note reading only in musicians. This area is thought to subserve the representation of sounds in music reading in musicians110.
Conclusions and future perspectives
Overall, the data demonstrated how musicians' improved reading proficiency discovered in independent reading tests was associated with better performance in orthographic and notation detection tasks, specifically with increased N170 and P300 amplitudes, and larger differences between targets and non-targets. From a neural standpoint, this was manifested in more extensive, bilateral activation111 of regions dedicated to orthographic processing, phonological processing, attentional shifting, target selection and eye movements. On the contrary, the diminished reading scores of both control and low-performing readers are linked to a unilateral and less prominent response in the N170 latency period. This is particularly associated with a decrease in activity in the VWFA, FEF, and frontal regions.
The evidence suggests that skilled readers, including musicians and good readers, show a unique connection between reading words and the right occipitotemporal cortex. It was discovered that reading proficiency was linked to the amplitude of N170 over the right hemisphere in control readers, indicating that greater responsiveness in the right OT cortex to words led to faster reading speeds (measured in syllables per second). This specific data directly associates the presence of an orthographic region in the right hemisphere with enhanced reading abilities.
Overall, it is suggested that the acquisition of music literacy could aid in improving reading skills in children who are at genetic risk for dyslexia or predisposed to it. This is due in part to the development of a bilateral orthographic area. Additionally, the intensive training in attentional and ocular displacement, which involves V5 and the oculomotor area, may serve as an enhancing and protective factor for reading ability33,112.
Given the insufficient activation of the left VWFA in surface dyslexic readers90, the functionality of a right homologous area for processing orthographic information would be a valuable neural aid. In addition, a possibly anomalous right-sided lateralization of phonological functions (described in children at genetic risk for dyslexia113,114) would benefit from the functionality on a right intra-hemispheric grapheme-to-phoneme conversion circuit. In both cases, music literacy might serve as a protective or rehabilitating factor for reading disorders.
It is worth noting that all study participants were fully right-handed. Therefore, functional hemispheric asymmetries cannot be attributed to individual differences but rather to their intensive music reading training (in comparison to a lack thereof). It was statistically shown that musicians' "word" training, resulting from book reading, was not as pronounced as that of control students.
The study clearly shows how (comparatively) poor readers did not show activation of the right OT cortex during word processing. A similar disadvantage was reported in face processing for unilateral versus bilateral cerebral activation, whereby males exhibited activation only in the right face fusiform area or FFA, leading to poorer performance than females in facial expression recognition tasks. This was reported by Proverbio115. Again, Koshik et al.116 reported that during tridimensional rotation of shapes, females exhibited activation in the right parietal cortex but not the left, which was found to correlate with poorer performance in mental rotation tasks.
Further investigations should be carried out on samples of adult dyslexic patients (musicians vs. non-musicians), although it would be complicated to disentangle the effects of compensating factors (neural plasticity) from the genetically inherited atipicality in neural wiring.
Data availability
The authors confirm that the data supporting the findings of this study are available within the article. Other information are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.
References
Schlaug, G., Jäncke, L., Huang, Y., Staiger, J. F. & Steinmetz, H. Increased corpus callosum size in musicians. Neuropsychologia 33(8), 1047–1055 (1995).
Schlaug, G. The brain of musicians: A model for functional and structural adaptation. Ann. N. Y. Acad. Sci. 930(1), 281–299 (2001).
Münte, T. F., Altenmüller, E. & Jäncke, L. The musician’s brain as a model of neuroplasticity. Nat. Rev. Neurosci. 3(6), 473–478 (2002).
Dalla Bella, S. Music and brain plasticity. In The Oxford handbook of music psychology (eds. Hallam, S. Cross, I. & Thaut, M.) 325–342 (2nd ed.). (Oxford University Press, 2016).
Peretz, I. & Zatorre, R. J. Brain organization for music processing. Annu. Rev. Psychol. 56, 89–114 (2005).
Shenker, J. J., Steele, C. J., Zatorre, R. J. & Penhune, V. B. Using cortico-cerebellar structural patterns to classify early- and late-trained musicians. Hum. Brain Mapp. 44(12), 4512–4522 (2023).
Steele, C. J., Bailey, J. A., Zatorre, R. J. & Penhune, V. B. Early musical training and white-matter plasticity in the corpus callosum: Evidence for a sensitive period. J. Neurosci. 33(3), 1282–1290 (2013).
Altenmüller, E. & Schlaug, G. Neurologic music therapy: The beneficial effects of music making on neurorehabilitation. Acoust. Sci. Technol. 34(1), 5–12 (2013).
Hutchinson, S., Lee, L. H. L., Gaab, N. & Schlaug, G. Cerebellar volume of musicians. Cerebral Cortex 13(9), 943–949 (2003).
Gaser, C. & Schlaug, G. Brain structures differ between musicians and non-musicians. J. Neurosci. 23(27), 9240–9245 (2003).
Schwenkreis, P. et al. Assessment of sensorimotor cortical representation asymmetries and motor skills in violin players. Eur. J. Neurosci. 26(11), 3291–3302 (2007).
Elbert, T., Pantev, C., Wienbruch, C., Rockstroh, B. & Taub, E. Increased cortical representation of the fingers of the left hand in string players. Science 270(5234), 305–307 (1995).
Hashimoto, I. et al. Is there training-dependent reorganization of digit representations in area 3b of string players?. Clin. Neurophysiol. 115(2), 435–447 (2004).
Bangert, M. & Schlaug, G. Specialization of the specialized in features of external human brain morphology. Eur. J. Neurosci. 24(6), 1832–1834 (2006).
Wong, P., Skoe, E., Russo, N. M., Dees, T. & Kraus, N. Musical experience shapes human brainstem encoding of linguistic pitch patterns. Nat. Neurosci. 10(4), 420–422 (2007).
Schneider, P., Sluming, V., Roberts, N., Bleeck, S. & Rupp, A. Structural, functional, and perceptual differences in Heschl’s gyrus and musical instrument preference. Ann. N. Y. Acad. Sci. 1060(1), 387–394 (2005).
Lappe, C., Herholz, S. C., Trainor, L. J. & Pantev, C. Cortical plasticity induced by short-term unimodal and multimodal musical training. J. Neurosci. 28(39), 9632–9639 (2008).
Habibi, A., Ilari, B., Heine, K. & Damasio, H. Changes in auditory cortical thickness following music training in children: Converging longitudinal and cross-sectional results. Brain Struct. Funct. 225(8), 2463–2474 (2020).
Schellenberg, G. E. Does exposure to music have beneficial side effects?’. In The Cognitive Neuroscience of Music (eds Peretz, I. & Zatorre, R. J.) (Oxford Academic Press, 2003).
Schellenberg, E. G. (2012). Cognitive performance after listening to music: A review of the Mozart effect. In Music, Health, and Wellbeing, 324–338.
Schellenberg, E. G. & Weiss, M. W. Music and cognitive abilities. In The Psychology of Music 3rd edn (ed. Deutsch, D.) 499–550 (Elsevier, Amsterdam, 2013). https://doi.org/10.1016/B978-0-12-381460-9.00012-2.
Gordon, R. L., Fehd, H. M. & McCandliss, B. D. Does music training enhance literacy skills? A meta-analysis. Front. Psychol. 6, 1777 (2015).
Forgeard, M. et al. The relation between music and phonological processing in normal-reading children and children with dyslexia. Music Percept. 25(4), 383–390 (2008).
Janurik, M., Surján, N. & Józsa, K. The relationship between early word reading, phonological awareness, early music reading and musical aptitude. J. Intell. 10(3), 50 (2022).
Anvari, S. H., Trainor, L. J., Woodside, J. & Levy, B. A. Relations among musical skills, phonological processing, and early reading ability in preschool children. J. Exp. Child Psychol. 83(2), 111–130 (2002).
Overy, K., Nicolson, R. I., Fawcett, A. J. & Clarke, E. F. Dyslexia and music: Measuring musical timing skills. Dyslexia 9(1), 18–36 (2003).
Goswami, U., Huss, M., Mead, N., Fosker, T. & Verney, J. P. Perception of patterns of musical beat distribution in phonological developmental dyslexia: Significant longitudinal relations with word reading and reading comprehension. Cortex 49(5), 1363–1376 (2013).
Flaugnacco, E. et al. Rhythm perception and production predict reading abilities in developmental dyslexia. Front. Human Neurosci. 8, 392 (2014).
Butzlaff, R. Can music be used to teach reading?. J. Aesthetic Educ. 34(3/4), 167–178 (2000).
Standley, J. M. & Hughes, J. E. Evaluation of an early intervention music curriculum for enhancing prereading/writing skills. Music Therapy Perspect. 15(2), 79–86 (1997).
Register, D. The effects of an early intervention music curriculum on prereading/writing. J. Music Ther. 38(3), 239–248 (2001).
Swaminathan, S., Schellenberg, E. G. & Venkatesan, K. Explaining the association between music training and reading in adults. J. Exp. Psychol. Learn. Memory Cognit. 44(6), 992 (2018).
Krafnick, A. J., Flowers, D. L., Luetje, M. M., Napoliello, E. M. & Eden, G. F. An investigation into the origin of anatomical differences in dyslexia. J Neurosci. 34(3), 901–908 (2014).
Proverbio, A. M., Manfredi, M., Zani, A. & Adorni, R. Musical expertise affects neural bases of letter recognition. Neuropsychologia 51(3), 538–549 (2013).
Li, S. T. K. & Hsiao, J. H. W. Music reading expertise modulates hemispheric lateralization in English word processing but not in Chinese character processing. Cognition 176, 159–173 (2018).
Stewart, L. A neurocognitive approach to music reading. Ann. N. Y. Acad. Sci. 1060, 377–386 (2005).
Stewart, L. et al. Becoming a pianist: An fMRI study of musical literacy acquisition. Ann. New York Acad. Sci. 999(1), 204–208 (2003).
Sergent, J., Zuck, E., Terriah, S. & MacDonald, B. Distributed neural network underlying musical sight-reading and keyboard performance. Science 257(5066), 106–109 (1992).
Schön, D., Anton, J. L., Roth, M. & Besson, M. An fMRI study of music sight-reading. Neuroreport 13(17), 2285–2289 (2002).
Mongelli, V. et al. Music and words in the visual cortex: The impact of musical expertise. Cortex 86, 260–274 (2017).
Meister, I. G. et al. Playing piano in the mind–an fMRI study on music imagery and performance in pianists. Brain Res. Cogn Brain Res. 19(3), 219–228 (2004).
Proverbio, A. M., Calbi, M., Manfredi, M. & Zani, A. Audio-visuomotor processing in the musician’s brain: An ERP study on professional violinists and clarinetists. Sci Rep. 29(4), 5866 (2014).
Wong, Y. K. & Gauthier, I. Holistic processing of musical notation: Dissociating failures of selective attention in experts and novices. Cogn. Affect Behav. Neurosci. 10(4), 541–551 (2010).
Li, T. K., Chan, H. Y. V., Li, L. & Hsiao, J. H. W. (2017). How does music reading expertise modulate visual processing of English words? An ERP study. In Annual Conference of the Cognitive Science Society, CogSci 2017. Cognitive Science Society.
Nakada, T., Fujii, Y., Suzuki, K. & Kwee, I. L. ‘Musical brain’revealed by high-field (3 Tesla) functional MRI. Neuroreport 9(17), 3853–3856 (1998).
Register, D., Darrow, A. A., Swedberg, O. & Standley, J. The use of music to enhance reading skills of second grade students and students with reading disabilities. J. Music Ther. 44(1), 23–37 (2007).
Habib, M. et al. Music and dyslexia: A new musical training method to improve reading and related disorders. Front. Psychol. 7, 26 (2016).
Cohen, L. et al. The visual word form area: Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain 123(2), 291–307 (2000).
Garrett, A. S. et al. Cortical activity related to accuracy of letter recognition. Neuroimage 11(2), 111–123 (2000).
Polk, T. A. et al. Neural specialization for letter recognition. J. Cognit. Neurosci. 14(2), 145–159 (2002).
Flowers, D. L. et al. Attention to single letters activates left extrastriate cortex. Neuroimage 21(3), 829–839 (2004).
Kuriki, S., Takeuchi, F. & Hirata, Y. Neural processing of words in the human extrastriate visual cortex. Cogn. Brain Res. 6(3), 193–203 (1998).
Pernet, C., Celsis, P. & Démonet, J. F. Selective response to letter categorization within the left fusiform gyrus. Neuroimage 28(3), 738–744 (2005).
Dehaene, S., Le Clec’H, G., Poline, J. B., Le Bihan, D. & Cohen, L. The visual word form area: A prelexical representation of visual words in the fusiform gyrus. Neuroreport 13(3), 321–325 (2002).
Proverbio, A. M., Zani, A. & Adorni, R. The left fusiform area is affected by written frequency of words. Neuropsychologia 46(9), 2292–2299 (2008).
Kronbichler, M. et al. The visual word form area and the frequency with which words are encountered: Evidence from a parametric fMRI study. Neuroimage 21(3), 946–953 (2004).
McCandliss, B. D., Cohen, L. & Dehaene, S. The visual word form area: Expertise for reading in the fusiform gyrus. Trends Cognit. Sci. 7(7), 293–299 (2003).
Bentin, S., Mouchetant-Rostaing, Y., Giard, M. H., Echallier, J. F. & Pernier, J. ERP manifestations of processing printed words at different psycholinguistic levels: Time course and scalp distribution. J. Cogn. Neurosci. 11(3), 235–260 (1999).
Benet, N., Krishna, R. & Kumar, V. Enhancement of processing capabilities of hippocampus lobe: A P300 based event related potential study. J. Audiol. Otol. 25(3), 119–123. https://doi.org/10.7874/jao.2021.00024 (2021).
Sartori, G., Job, R. & Tressoldi, P. E. Battery for the assessment of developmental dyslexia and dysorthographia. Batteria per la valutazione della dislessia e della disortografia evolutiva (Giunti OS, 1995).
George, F. & Pech-Georgel, C. (2017). VALS-Assessment of Reading and Writing Difficulties in Adulthood (Italian translation by A. G. De Cagno, K. Gaspary, S. Mantione). Trento, Erickson
Bertinetto, P. M., Burani, C., Laudanna, A., Marconi, L., Ratti, D., Rolando, C., et al. (2006). Corpus e Lessico di Frequenza dell’Italiano Scritto (CoLFIS). Scuola Normale Superiore di Pisa.
Oostenveld, R. & Praamstra, P. The five percent electrode system for high-resolution EEG and ERP measurements. Clin. Neurophysiol. 112(4), 713–719 (2001).
Palmero-Soler, E., Dolan, K., Hadamschek, V. & Tass, P. A. swLORETA: a novel approach to robust source localization and synchronization tomography. Phys. Med. Biol. 52(7), 1783–1800 (2007).
Zanow, F. & Knösche, T. R. ASA–Advanced Source Analysis of continuous and event-related EEG/MEG signals. Brain Topogr. 16(4), 287–290 (2004).
Puurtinen, M. Eye on music reading: A methodological review of studies from 1994 to 2017. J. Eye Mov. Res. https://doi.org/10.16910/jemr.11.2.2 (2018).
Simon, G., Petit, L., Bernard, C. & Rebaï, M. N170 ERPs could represent a logographic processing strategy in visual word recognition. Behav. Brain Funct. 3(1), 1–11 (2007).
Wang, F. et al. Distinct neural sources underlying visual word form processing as revealed by steady state visual evoked potentials (SSVEP). Sci. Rep. 11(1), 1–15 (2021).
Cohen, L. & Dehaene, S. Specialization within the ventral stream: The case for the visual word form area. Neuroimage 22(1), 466–476 (2004).
Devlin, J. T., Jamison, H. L., Gonnerman, L. M. & Matthews, P. M. The role of the posterior fusiform gyrus in reading. J. Cognit. Neurosci. 18(6), 911–922 (2006).
Price, C. J. & Devlin, J. T. The pro and cons of labelling a left occipitotemporal region: “The visual word form area”. Neuroimage 22(1), 477–479 (2004).
Proverbio, A. M., Zotto, M. D. & Zani, A. Greek language processing in naive and skilled readers: Functional properties of the VWFA investigated with ERPs. Cognit. Neuropsychol. 23(3), 355–375 (2006).
Picton, T. W. The P300 wave of the human event-related potential. J. Clin. Neurophysiol. 9(4), 456–479 (1992).
Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 118(10), 2128–2148 (2007).
Bledowski, C. et al. Localizing P300 generators in visual target and distractor processing: A combined event-related potential and functional magnetic resonance imaging study. J. Neurosci. 24(42), 9353–9360 (2004).
Orlandi, A. & Proverbio, A. M. ERP indices of an orientation-dependent recognition of the human body schema. Neuropsychologia 146, 107535 (2020).
Stoeckel, C., Gough, P. M., Watkins, K. E. & Devlin, J. T. Supramarginal gyrus involvement in visual word recognition. Cortex 45(9), 1091–1096 (2009).
Vossel, S., Weidner, R., Moos, K. & Fink, G. R. Individual attentional selection capacities are reflected in interhemispheric connectivity of the parietal cortex. Neuroimage 129, 148–158 (2016).
Buschman, T. J. & Miller, E. K. Serial, covert shifts of attention during visual search are reflected by the frontal eye fields and correlated with population oscillations. Neuron 63(3), 386–396 (2009).
Altınkaynak, M. et al. Diagnosis of Attention Deficit Hyperactivity Disorder with combined time and frequency features. Biocybernet. Biomed. Eng. 40(3), 927–937 (2020).
Kaiser, A. et al. Earlier versus later cognitive event-related potentials (ERPs) in attention-deficit/hyperactivity disorder (ADHD): A meta-analysis. Neurosci. Biobehav. Rev. 112, 117–134 (2020).
Papagiannopoulou, E. A. & Lagopoulos, J. P300 event-related potentials in children with dyslexia. Ann. Dyslexia 67(1), 99–108 (2017).
Koyama, M. S. et al. Cortical signatures of dyslexia and remediation: An intrinsic functional connectivity approach. PLoS One 8(2), e55454 (2013).
Centanni, T. M. et al. Disrupted left fusiform response to print in beginning kindergartners is associated with subsequent reading. NeuroImage Clin. 22, 101715 (2019).
Maisog, J. M., Einbinder, E. R., Flowers, D. L., Turkeltaub, P. E. & Eden, G. F. A meta-analysis of functional neuroimaging studies of dyslexia. Ann. N. Y. Acad. Sci. 1145(1), 237–259 (2008).
Bahmani, Z., Clark, K., Merrikhi, Y., Mueller, A., Pettine, W., Isabel Vanegas, M., et al. (2019). Prefrontal contributions to attention and working memory. In Processes of Visuospatial Attention and Working Memory, 129–153.
Turkeltaub, P. E. et al. Alexia due to ischemic stroke of the visual word form area. Neurocase 20(2), 230–235 (2014).
Chandregowda, A. et al. Neurodegeneration of the visual word form area in a patient with word form alexia. Neurol. Clin. Neurosci. 9(4), 359 (2021).
Arrington, C. N., Ossowski, A. E., Baig, H., Persichetti, E. & Morris, R. The impact of transcranial magnetic stimulation on reading processes: A systematic review. Neuropsychol. Rev. 33, 1–23 (2022).
Brem, S. et al. Visual word form processing deficits driven by severity of reading impairments in children with developmental dyslexia. Sci. Rep. 10(1), 1–14 (2020).
Salmelin, R. et al. Impaired visual word processing in dyslexia revealed with magnetoencephalograpy. Ann. Neurol. 40, 157–162 (1996).
Pugh, K. R. et al. Functional neuroimaging studies of reading and reading disability (developmental dyslexia). Ment. Retard. Dev. Disabil. Res. Rev. 6(3), 207–213 (2000).
Pugh, K. R. et al. The angular gyrus in developmental dyslexia: Task specific differences in functional connectivity in posterior cortex. Psychol Sci. 11, 51–56 (2000).
Girbau-Massana, D., Garcia-Marti, G., Marti-Bonmati, L. & Schwartz, R. G. Gray–white matter and cerebrospinal fluid volume differences in children with specific language impairment and/or reading disability. Neuropsychologia 56, 90–100 (2014).
Van der Mark, S. et al. Children with dyslexia lack multiple specializations along the visual word-form (VWF) system. Neuroimage 47(4), 1940–1949 (2009).
Rodrigues, A. P., Rebola, J., Pereira, M., van Asselen, M. & Castelo-Branco, M. Neural responses of the anterior ventral occipitotemporal cortex in developmental dyslexia: Beyond the visual word form area. Investig. Ophthalmol. Vis. Sci. 60(4), 1063–1068 (2019).
Fraga González, G. et al. Brain-potential analysis of visual word recognition in dyslexics and typically reading children. Front. Human Neurosci. 8, 474 (2014).
Van der Mark, S. et al. The left occipitotemporal system in reading: Disruption of focal fMRI connectivity to left inferior frontal and inferior parietal language areas in children with dyslexia. Neuroimage 54(3), 2426–2436 (2011).
Fan, Q., Anderson, A. W., Davis, N. & Cutting, L. E. Structural connectivity patterns associated with the putative visual word form area and children׳ s reading ability. Brain Res. 1586, 118–129 (2014).
Sefcikova, V., Sporrer, J. K., Juvekar, P., Golby, A. & Samandouras, G. Converting sounds to meaning with ventral semantic language networks: Integration of interdisciplinary data on brain connectivity, direct electrical stimulation and clinical disconnection syndromes. Brain Struct. Funct. 227(5), 1545–1564 (2022).
Baumann, O. & Mattingley, J. B. Functional organization of the parahippocampal cortex: Dissociable roles for context representations and the perception of visual scenes. J. Neurosci. 36(8), 2536–2542 (2016).
Booth, J. R. et al. Development of brain mechanisms for processing orthographic and phonologic representations. J. Cogn. Neurosci. 16(7), 1234–1249 (2004).
Proverbio, A. M. & Zani, A. Developmental changes in the linguistic brain after puberty. Trends Cogn. Sci. 9(4), 164–167 (2005).
Westendorff, S., Kaping, D., Everling, S. & Womelsdorf, T. Prefrontal and anterior cingulate cortex neurons encode attentional targets even when they do not apparently bias behavior. J. Neurophysiol. 116(2), 796–811 (2016).
Choi, W., Desai, R. H. & Henderson, J. M. The neural substrates of natural reading: A comparison of normal and nonword text using eyetracking and fMRI. Front. Human Neurosci. 8, 1024 (2014).
Corbetta, M., Shulman, G. L., Miezin, F. M. & Petersen, S. E. Superior parietal cortex activation during spatial attention shifts and visual feature conjunction. Science 270(5237), 802–805 (1995).
Bowling, J. T., Friston, K. J. & Hopfinger, J. B. Top-down versus bottom-up attention differentially modulate frontal–parietal connectivity. Human Brain Map. 41(4), 928–942 (2020).
Mecklinger, A., Bosch, V., Grünewald, C., Bentin, S. & von Cramon, D. Y. What have Klingon letters and faces in common? An fMRI study on content-specific working memory systems. Human Brain Map. 11(3), 146–161 (2000).
Japee, S., Holiday, K., Satyshur, M. D., Mukai, I. & Ungerleider, L. G. A role of right middle frontal gyrus in reorienting of attention: A case study. Front. Syst. Neurosci. 9, 23 (2015).
Bouhali, F., Mongelli, V., de Schotten, M. T. & Cohen, L. Reading music and words: The anatomical connectivity of musicians’ visual cortex. NeuroImage 212, 116666 (2020).
Patston, L. L., Hogg, S. L. & Tippett, L. J. Attention in musicians is more bilateral than in non-musicians. Laterality 12(3), 262–272 (2007).
Flaugnacco, E. et al. Music training increases phonological awareness and reading skills in developmental dyslexia: A randomized control trial. PLoS ONE 10(9), e0138715 (2015).
Leppänen, P. H., Pihko, E., Eklund, K. M. & Lyytinen, H. Cortical responses of infants with and without a genetic risk for dyslexia: II. Group effects. Neuroreport 10(5), 969–973 (1999).
Pihko, E. et al. Cortical responses of infants with and without a genetic risk for dyslexia: I. Age effects. Neuroreport 10(5), 901–905 (1999).
Proverbio, A. M. Sexual dimorphism in hemispheric processing of faces in humans: A meta-analysis of 817 cases. Soc. Cogn. Affect Neurosci. 16(10), 1023–1035 (2021).
Koscik, T., O’Leary, D., Moser, D. J., Andreasen, N. C. & Nopoulos, P. Sex differences in parietal lobe morphology: Relationship to mental rotation performance. Brain Cogn. 69(3), 451–459 (2009).
Acknowledgements
We express gratitude to all participating students and musicians, as well as their professors, for their kind support and cooperation throughout this study.
Author information
Authors and Affiliations
Contributions
A.M.P. conceived the idea and designed the study. M.M., G.A. and M.M.P. recruited and tested participants and analysed the data. All authors contributed to the statistical analyses and interpretation of the results. A.M.P. took the lead in writing the final manuscript. All authors provided critical feedback and helped shape the research, analysis and manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pantaleo, M.M., Arcuri, G., Manfredi, M. et al. Music literacy improves reading skills via bilateral orthographic development. Sci Rep 14, 3506 (2024). https://doi.org/10.1038/s41598-024-54204-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-54204-8
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
