
Abstract
The fundamental relationship between the mesoscopic structure of neuronal circuits and organismic functions they subserve is one of the major challenges in contemporary neuroscience. Formation of structurally connected modules of neurons enacts the conversion from single-cell firing to large-scale behaviour of an organism, highlighting the importance of their accurate profiling in the data. While connectomes are typically characterized by significant sparsity of neuronal connections, recent advances in network theory and machine learning have revealed fundamental limitations of traditionally used community detection approaches in cases where the network is sparse. Here we studied the optimal community structure in the structural connectome of Caenorhabditis elegans, for which we exploited a non-conventional approach that is based on non-backtracking random walks, virtually eliminating the sparsity issue. In full agreement with the previous asymptotic results, we demonstrated that non-backtracking walks resolve the ground truth annotation into clusters on stochastic block models (SBM) with the size and density of the connectome better than the spectral methods related to simple random walks. Based on the cluster detectability threshold, we determined that the optimal number of modules in a recently mapped connectome of C. elegans is 10, which precisely corresponds to the number of isolated eigenvalues in the spectrum of the non-backtracking flow matrix. The discovered communities have a clear interpretation in terms of their functional role, which allows one to discern three structural compartments in the worm: the Worm Brain (WB), the Worm Movement Controller (WMC), and the Worm Information Flow Connector (WIFC). Broadly, our work provides a robust network-based framework to reveal mesoscopic structures in sparse connectomic datasets, paving way to further investigation of connectome mechanisms for different functions.
Similar content being viewed by others


Introduction
Complexity of biological and social systems driven by collective behaviour of their agents is commonly studied using network (or graph) representation, where nodes represent agents and edges correspond to pairwise coupling between them. The resulting dimensionality reduction frequently allows to extract the most valuable information about hidden relationships governing static and dynamic properties of a system. One of the most striking and practically important examples of such information is the mesoscopic organization of the network in modules or communities.
The nervous system is no exception in this regard as it can be represented as a structural connectome, that is, a graph, where vertices are nerve cells and edges reflect direct structural connections (wiring) between them. Similarly to most of real-world networks, the connectome is extremely sparse, that is, its number of theoretically possible connections between neurons greatly exceeds the factual amount of connections1.
Such a reduction of excessive edges is a consequence of network modularity, a tendency to form assortative communities (modules) with relatively loose inter-connections. Like an effective team work of people where complex problem requires distribution of tasks among specialized groups of participants, mesoscopic organization of neurons in a connectome serves to facilitate certain functions of the nervous system, such as “fire together wire together” principle,2. Thus, accurate detecting of modules (communities) in the connectome data can help to establish a conversion between micro-level single neuron interactions and macro-level organism behaviour.
Community detection is an extremely hot topic in various fields such as technological3,4, biological5,6, social7,8 and economical9,10,11 fields. A widely used approach in community detection is a spectral decomposition of a linear operator defined on a network: information about communities is then encoded in several leading eigenvectors12,13. It was shown that all commonly used matrices (adjacency, Laplacian, modularity, non-backtracking, see “Methods”) readily classify nodes as long as the network density is sufficient14,15. In particular, the modularity operator is one of the most efficient instruments that successfully detects communities in stochastic networks of various nature8,16,17,18. The modularity operator can be used to extract mesoscopic organization in C. elegans19.
Sparse graphs are a special case where most of the traditional community detection methods suffer from fundamental limitations. Namely, at a given cluster strength there is a critical network density below which community detection becomes a very difficult problem20. Furthermore, traditionally used operators (adjacency, Laplacian, modularity) turn out to fail above this threshold, since their leading eigenvectors rapidly become uncorrelated with the intrinsic community structure upon decrease of network density. This behaviour is explained as the emergence of vertices with anomalously high degree (hubs), which eventually perturbs the spectral edge of these operators because of the “Lifshitz tails” in the spectral density of sparse graphs21,22,23. Localization on hubs, and not on the communities, is thus a major drawback for all conventional spectral methods in the sparse regime.
To address this issue, Krzakala et al. proposed20 to make use of the non-backtracking (Hashimoto) random walks on the associated directed graph. By construction, such walks cannot revisit the same node immediately at the following step and, as a result, they do not localize on hubs. The leading eigenvectors of the non-backtracking operator (the transfer matrix of non-backtracking walks) encode for the community structure of the graph up to the theoretical resolution limit20. Due to their intrinsic ability to deal with sparse graphs, the non-backtracking walks have received increasing attention in the analysis of biological datasets. Recently we have shown that this approach allows to annotate compartments in the three-dimensional chromatin organization at the single-cell level24.

Clusterization of C. elegans connectome by means of the spectrum of the non-backtracking flow matrix: 0. A simplified scheme of the C. elegans nervous system; 1. Representation of the connectome as the adjacency matrix; 2. Construction of non-backtracking walks on the connectome network; 3. Normalization by the out-degree towards the non-backtracking flow operator; 4. The spectrum of the flow matrix (new data25) with the orange dots representing the eigenvalues outside of the spectral radius (pale red), which are used for identification of the communities.
In this work, we examine neuronal connectivity in Caenorhabditis elegans—one of the simplest organisms with a structural connectome first mapped by White et al. in 198626, which has been completely described by now25. The nervous system is a prominent part of C. elegans with almost one third of all cells in its body being neurons. Importantly, the morphology, location and connectivity of each neuron are remarkably invariant between individuals27 (it is worth noting that there is now a growing concern on to what extent this is actually true28,29,30,31), which arguably makes this organism a convenient model for studying neuronal connectivity and related functions. Curiously, due to the elongated shape of the nervous system of the worm, the adjacency matrix of the C. elegans connectome is similar to a single-cell chromosome contact map, which describes the spatial proximity of loci in individual conformations of chromosomes24,32. The so-called scaling in both types of matrices, being a generic polymer (or worm-like chain) feature, is a major source of sparsity in the data. This prompts one to apply to the C. elegans connectome the clustering procedures that has been specifically designed for reliable detection of communities in sparse networks.
Here we study the mesoscopic organization of C. elegans connectome by means of the non-backtracking walks. Namely, we construct the Newman’s non-backtracking flow operator, which describes the transfer probability of a random walk on the associated directed connectome with prohibited immediate revisiting. The isolated part of the flow matrix spectrum is known to encode for communities and can be used by the clustering algorithm. We ran simulations of community detection on stochastic block models (SBMs) of the corresponding size and density as the connectome and demonstrated that the non-backtracking flow matrix outperformed all traditional operators, in the full agreement with asymptotic results of20. In particular, we show better performance of non-backtracking walks over the modularity operator and other approaches, which were previously used for spectral clustering of the C. elegance connectome19,33.
We consider two C. elegans connectome datasets: “old”34,35 and “new”25. The difference in data completeness between these datasets is quite significant: the number of the edges has approximately doubled in the new connectome (for more details see “Methods”). We reveal that despite the evident difference in the network density, the modular organization of the two connectomes is rather similar, as reflected by the spectrum of the non-backtracking flow matrix. To establish the detectable amount of communities for each connectome data we propose an algorithm based on the theoretical detectability threshold for SBM-like graphs. In the new data25 our approach reveals 10 detectable communities in the C. elegans connectome, matching the number of isolated eigenvalues in the flow matrix spectrum. The biological interpretation of the communities in the complete connectome suggests that the found clusters highly correlate with the co-localization of neurites (so-called, contactome) in the nervous system of the worm, can be further associated with specific neuronal functions and also overlap with the anatomically defined ganglia. Importantly, we demonstrate that the non-backtracking clusters are much better interpretable than partitions by other spectral algorithms, as well as the modules reported previously19,33. Ultimately our integrative analysis of the mesoscopic structure of the structural connectome and related functions has revealed three neuronal compartments in the C. elegans: A. Worm Brain, B. Worm Movements Controller, C. Worm Information Flow Connector.
Stochastic block model and non-backtracking random walks
Here we provide a network theory background underlying clustering methods, which is instructive for the definition of the model. The particular connection with several widely used spectral clustering approaches is described in the “Methods” section. Stochastic block model (SBM) is a commonly used benchmark for community detection in real-world networks, with several important results obtained for asymptotically large networks15,16,20,36,37,38. By definition, a SBM is a generalization of an Erdös–Renyi random graph on N nodes, where all edges are generated independently with the probability p that depends on the type of the nodes that it connects. The nodes in a graph belong to k different types (clusters), \(G_i, i = 1, 2, \ldots , k\). Thus, each pair of nodes \((i, j): i \in G_r, j \in G_t\) gets independently connected by an edge with some probability \(w_{rt}\), which can be written as a matrix of pairwise cluster probabilities \(W = \{w_{rt}\}\) with \((r, t) = 1, 2, \ldots , k\). The corresponding entry in the adjacency matrix \(A_{ij}\) is 1 with probability \(w_{rt}\) and 0 otherwise. In the simplest version of the model (the planted SBM), all off-diagonal elements of the matrix W are the same and equal to \(w_{out}\), while all diagonal elements of W are equal to \(w_{in}\):
The assortative community structure corresponds to \(w_{in}>w_{out}\). In the connectome context, the neurons belonging to the same cluster have a preferentially higher probability to be connected with a link than the neurons from different clusters. Still, some of the neurons within the same cluster in the structural connectome are not connected (clusters are not always cliques), allowing one to make use of stochastic models.
Importantly, for SBMs there is a certain threshold on the minimally allowed difference \(\Delta w = w_{in} - w_{out}\) between the probabilities in order for the cluster structure to be resolved15,36. Following conventional notation, let us introduce the rescaled cluster affinities, \(c_{in}=Nw_{in}\) and \(c_{out}=Nw_{out}\), which scale linearly with the number of the inner and outer edges of a typical community. The detectability rule suggests that the SBM clusters are asymptotically resolved (\(N \gg 1\)) as long as
where \(c=(c_{in}+c_{out})/2\) is the average of \(c_{in}, c_{out}\). For dense networks \(c_{in}, c_{out}, c \sim O(N)\) and, thus, condition (2) is satisfied at any small \(\Delta w>0\). In the sparse case, \(c \sim O(1)\), the threshold (2) provides a practically important condition on the parameters \(\Delta w\) and k for the cluster structure to be resolved.
Spectral methods, such as Laplacian, adjacency or modularity, have been widely used to uncover the community structure in relatively dense stochastic block model networks12,14,16,37,38,39. The leading non-trivial eigenvectors of the corresponding operators provide dimensionality reduction of the system and these latent coordinates are then used by some conventional clustering algorithm (such as k-means) to perform partitioning into specified number of clusters12. However, as it was noted in20, for sparse networks the leading eigenvectors become uncorrelated with true community structure above the theoretical threshold (2). This is because of the abundance of star-like sub-graphs (hubs) in a sparse network, which are identified by these operators instead of cyclic subgraphs associated with the internal structure of communities. Indeed, as these operators are related to random walks on a graph, true clusters interfere with hubs in their spectrum. As a result, it turns out that the spectral methods that exploit random-walk-related operators (such as modularity, adjacency or Laplacian) fail to find communities in rather sparse networks, despite of the network parameters satisfying the detectability condition (2).
To overcome this difficulty, the spectrum of the Hashimoto matrix B can be utilized, which is a transfer matrix of non-backtracking walks on a graph. It is defined on the edges of the directed graph, \(i \rightarrow j, k \rightarrow l\), as follows
It is seen from (3) that the non-backtracking operator prohibits returns to the point which a walker visited at the previous step, thus effectively circumventing localization on the hubs. Notably, matrix B is non-symmetric and has a complex spectrum. For Poissonian graphs, the spectrum of B is constrained within a circle in the complex plane, whereas real eigenvalues of B lying out of the circle are relevant to the community structure even in sparse networks. Associating the corresponding eigenvectors with the network partitioning allows detecting communities all the way down to the theoretical limit (2). Interestingly, a “reluctant” version of the non-backtracking operator allows exploring the hanging trees of the network39, which the original operator B ignores by construction.
In37 the corresponding flow operator was proposed, which conserves the probability flow at each step of the non-backtracking walker (see Fig. 1):
where \(d_j\) is the degree of the vertex j. While the powers of non-backtracking matrix B count the non-backtracking walks of particular length on a graph, the flow matrix F is the transfer matrix of the non-backtracking probability. Similarly to the non-backtracking matrix, the bulk of the spectrum of F lies in the complex plane within a circle of the radius
but, as shown in37, has a more clear edge of the spectral band. Importantly, the amount of isolated eigenvalues in the spectrum of the flow matrix corresponds to the number of clusters in SBM network37. In what follows, we will use the flow matrix (4) for the purpose of the connectome clustering.
The flow matrix F defines the non-backtracking probability flow along the edges. While one is interested in the classification of the nodes, the eigenvectors of F have to be carefully translated from the space of edges to the space of nodes. This is conventionally performed using the relation between the quadratic forms of modularity and flow operators24,37. From this correspondence one can see that contribution \(u_i\) to the i-th node of the graph comes from the in-flow along all the directed edges adjacent to i. This procedure can be formally written as follows
where \(v_{j \rightarrow i}^{F}\) is the component of the eigenvector of the flow matrix, corresponding to the directed edge \(j\rightarrow i\). The element of the adjacency matrix \(A_{ij}\) is non-zero as long as there is an edge between i to j. Using (6) one can switch from edges to nodes representation of the non-backtracking flow and perform clustering of the nodes, e.g. using k-means on leading vectors \(u_i\). Trivially, isolated vertices in a graph have undefined values of the flow, and they are not involved in graph clustering.
Clustering the connectome of a worm: how many clusters are detectable?
The nervous system is one of the most complex parts of the nematode C. elegans as the neurons constitute one third of all cells in this organism. The graph of the hermaphrodite connectome consists of \(N=302\) vertices representing neurons and \(C=4887\) edges (chemical synapses, see “Methods”) between them representing structural connections, as recorded in the new dataset25. Since only \(11\%\) of the theoretically possible number of edges are present in the network, one may conclude that we deal with a rather sparse network. In the old dataset, in contrast, the network density is less than \(5\%\), thus increasing the role of sparsity, as we will see below.
In order to obtain communities in C. elegans connectome we implemented the spectral clustering approach, based on the leading non-trivial eigenvectors of the non-backtracking flow matrix (4). The spectrum of the actual network corresponding to the new data is shown in the Fig. 1. Its isolated part, which is essential for the spectral clustering, consists of the maximal (trivial) eigenvalue and 9 smaller eigenvalues that lie on the real line outside the bulk, constrained by a circle of the radius (5). This picture suggests that there are 10 communities in the network, encoded in the corresponding eigenvectors37. As we further found, the amount of isolated eigenvalues is invariant in both datasets analysed (see Fig. S2), despite the two-fold difference in the network density. This implies the existence of a stable mesoscopic structure, as revealed by the non-backtracking spectrum.
Then we asked—and this is not trivial—how many clusters out of 10 can be reliably resolved in the given data. To this end, we suggest an approach based on the detectability threshold (2). Namely, we note that for a given mean cluster strength \(\Delta w\) the condition (2) establishes the maximum number of clusters that can be resolved in the sparse network of a given size N and the average link probability \(w=c/N\). Therefore, the critical number of clusters is related to the network parameters as follows
To find \(k_{max}\) in the C. elegans connectome we cluster the network into consecutive number of clusters \(k = 2,3,4, \ldots , 10\) using the eigenvectors of the non-backtracking flow matrix and determine \(c_{in}\) and \(c_{out}\) as the average amount of intra- and inter-links in the detected clusters. For each dataset we can determine the maximal amount of clusters \(k_{max}\), such that \(c_{in}-c_{out}\) is still greater than \(k\sqrt{c}\) (see the sketch in Fig. 2, explaining the procedure). Thus, the resulting value of \(k_{max}\) provides the number of detectable communities, according to the detectability condition (2). An hierarchy of resulting community structures for different k is shown in Fig. 1.
While the total number of isolated eigenvalues is invariant in both datasets, the detectability condition clearly suggests a strong sensitivity of the amount of resolvable clusters \(k_{max}\) to the network density of the sparse experimental data (see Fig. 2). In the old and incomplete connectome data34,35 only \(k_{max}=7\) clusters can be resolved, thus, the remaining 3 modules cannot be established due to strong sparsity of the data. At the same time, based on the same detectability condition applied to the new connectome data25 with doubled density of synaptic connections, we conclude that all \(k_{max}=10\) communities can be reliably recovered using the information from the flow matrix eigenvectors.

Graphical representation of the condition (2) as a criterion for the optimal number of clusters that can be detected (a) in the old34,35 and (b) new25 connectome data. The intersection point of blue and orange curves provides the maximal amount of clusters \(k_{max}\) at which the detectability condition (2) is satisfied.
Non-backtracking flow outperforms other spectral methods in clustering of connectomes
Having found the detectable number of modules, we next compared the performance of the non-backtracking flow matrix with traditional clustering operators, such as the normalized Laplacian and modularity matrix, on artificial networks with statistical properties similar to the experimental dataset25. To this end, we generated a family of stochastic block models with blocks similar to the ones we have obtained in the C. elegans connectome. Namely, we fixed the network size, \(N=279\), the outer-cluster probability, \(w_{out}= 0.05\), and the total number of clusters, \(k=7\). Furthermore, the sizes of the simulated blocks were chosen to match the sizes of the clusters in the original data. The only parameter subject to variation was the inner-cluster probability, \(w_{in} = \{0.05, \ldots , 0.22, \ldots , 0.6\}\). For each value of \(w_{in}\) we generated 200 random SBMs. We ran the spectral clustering on \(k=7\) leading eigenvectors of four operators: non-backtracking flow, normalized Laplacian, Laplacian and modularity.
The partitions predicted by the four operators were then assessed using the AMI scores, see Fig. S1. The results demonstrate distinctively better performance of the non-backtracking flow and normalized Laplacian over the modularity and Laplacian in prediction of the ground truth cluster structure of the simulated SBMs. The flow operator slightly outperformed the normalized Laplacian, especially in the region of intermediate relative cluster strengths, \(w_{in}/w_{out} \approx 4-5\), which correspond to the empirical value. Such a moderate difference in prediction scores between the two operators is the result of a small network size N. As it was previously shown, in the limit of large (asymptotic) networks the non-backtracking operator outperforms the normalized Laplacian as well20. Notably, the empirical value \(w_{in}/w_{out} \approx 4.4\), labeled by red dots on the AMI curves, is close to the detectability limit, as in this critical region AMI abruptly decays to zero (see Fig. S1). For large \(N \gg 1\) there is an associated phase transition15,36, thus highlighting criticality of the worm connectome.
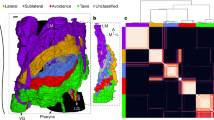
Next we turned to performance analysis of different clustering algorithms on the experimental network. We compute the Q-scores for each of the annotations into clusters, which is equal to the modularity score of the partition and is defined on the basis of the Newman-Girvan modularity operator (see “Methods”). Using this metric we reveal a better quality of non-backtracking flow clusters for various values of the total number of clusters k (see Fig. 3a). As one can infer from Fig. 3a and b, for all k, except for \(k=8\), the estimated quality of clustering by the flow operator outperforms other spectral approaches. In particular, the leading eigenvectors of the modularity matrix provide much poorer annotation into clusters, despite the quality metric being based on the modularity. This result is due to the sparsity issue discussed above; the leading spectrum of the non-backtracking flow matrix much better approximates the optimum of modularity function of a sparse graph than the leading spectrum of the modularity matrix itself. At the same time, we see that the normalized Laplacian produces annotations with similar, but steadily lower quality compared to the non-backtracking flow operator, in accord with the analysis of the SBM networks above (Fig. S1). Additionally, we have clustered the connectome using Infomap algorithm40. Interestingly, the Infomap suggests that the optimal number of clusters equals to \(k=3\) and provides a similar value of the modularity as obtained by the leading eigenvectors of the flow matrix (\(Q \approx 0.38\)), see Fig. 3a. However, it fails to find all the clusters that evidently exist in the network (Fig. 2).
To complement our analysis of Q-scores and better understand the (dis-)similarity of predictions by the flow matrix and other operators we further compute the pairwise relative overlaps between the clusters for the particular value of \(k=10\) (see Fig. S3). With the overlap threshold in \(80\%\) we find that 5 out of 10 modularity clusters poorly correspond to the flow matrix clusters, which translates into approximately \(10\%\) difference in the Q-scores (Fig. 3a). In the full agreement with the analysis above, only 3 clusters of normalized Laplacian overlap with the flow matrix clusters by less than \(80\%\). Importantly, the smallest 10th cluster of the flow operator (consisting of as few as 7 neurons) is not resolved by normalized Laplacian. It is not fully resolved by modularity operator neither: despite an apparently high overlap with a modularity cluster (consisting of 4 neurons), the modularity fails to capture the other 3 neurons of the 10th flow matrix cluster. Indeed, tiny clusters in sparse networks present a particular challenge for traditional algorithms.
Additionally, we compare the structural clusters with ganglia. While ganglia represent the groups defined purely by anatomy and cannot be used as the ground truth for the structural partitions, they provide an important biological benchmark to estimate the technical noise produced by different clustering algorithms. We find that for sufficiently large number of clusters \(k>4\), the AMI score for the non-backtracking flow takes the highest value compared to all other algorithms, Fig. 3c. It is also worth noting that enrichment of the edges in the new dataset25 has significantly increased the mutual information between the structural modules and the ganglia (Table S1). Still, similarity between the cluster groups obtained from old and new data is rather strong, see Fig. 3a. Furthermore, as Fig. 3b suggests, all flow matrix clusters in the new data have a statistically significant overlap with at least one ganglia (\(p \le 10^{-3}\)).
As another biological benchmark, we consider partitioning of the C. elegans nervous system into six groups corresponding to different neuronal functions: motor neurons (head, body, sublateral, sex specific), sensory neurons, interneurons (see Fig. 3c). Noticeably, three functional types (BM, HM and SM) are located within a particular group of clusters (\(p \le 10^{-5}\)), as derived by the flow matrix; body motor neurons belong to 5th–8th clusters, head motor neurons mostly locate in the 4th cluster, sublateral motor neurons locate in the 2nd cluster. The group of interneurons belongs to the 1st and the 10th cluster (at \(p \approx 10^{-3}\)) and sensory neurons are spread between the 3rd, 4th and 9th clusters.
Put together, the statistical analysis of clusters obtained on real and simulated connectomes suggests that the non-backtracking flow operator outperforms conventional clustering approaches on networks of size and composition similar to the C. elegans connectome.
Comparison with previously reported connectome modules
It is instructive to compare the results of the our algorithm (flow matrix) with other partitions reported in the literature. Here we analyse the results obtained by different approaches on the old dataset34, since to the best of our knowledge there have been no attempts to cluster the new connectome data in the literature.
Two open-source alternative annotations are considered, which were obtained by two different algorithms: iterative modularity maximization (IMMA)19 and Erdos-Renyi mixture model (ERMM)33. The IMMA approach is based on maximization of the modularity score (\(k=6\) modules were found for the weighted connectome); the ERMM is a non-deterministic algorithm that fits an arbitrary, not planted, SBM network to the real connectome data (\(k=9\) modules were found). First we compute the Q-scores of the clusters as obtained by the algorithms and compare to the quality of clusters produced by the flow matrix. We find that the flow matrix clusters produce a significantly higher quality of partition with the modularity value of 0.32, which is to be compared with 0.27 and 0.19 for ERMM and IMMA clusters, respectively.
Next, we compute the mutual information between partitions predicted by various methods and biological benchmarks. As Table S1 demonstrates, the IMMA algorithm yields worse agreement with the ganglia (AMI \(=0.31\)) as compared to the flow matrix clusters (AMI \(=0.34\)). At the same time, the ERMM approach has the same AMI score with ganglia (AMI \(=0.34\)) and the mutual information between the ERMM and flow matrix clusters is sufficiently high (AMI\(=0.45\)). Also both the flow matrix and ERMM algorithm grouped command interneurons from the lateral ganglion together \((p \le 10^{-5})\). Besides that, motor neurons were united into structural clusters by all three methods, see Fig. S5. All three methods successfully split parts of sensory neurons and interneurons, but only flow matrix was able to isolate the polymodal neurons \((p \le 10^{-4})\).
Overall, we conclude that the previous partitions of C. elegans connectome, based on the old dataset34, display lower quality of clustering (Q-values) and worse biological interpretability than can be achieved using the leading part of the spectrum of the non-backtracking flow matrix.

Quality comparison of different spectral approaches on the new connectome data25. (a) Q-values (15) of partitions into k clusters by various operators. (b) Partitions of the C. elegans connectome into \(k=10\) clusters obtained by different spectral methods. Each of the ten clusters inferred by the flow operator is annotated with a unique color according to its compartmental affiliation. (c) AMI score between ganglia and structural partitions for different number of clusters k.
Cross-talk between the clusters is determined by neuronal programs
On the basis of the complementary nature of detected communities (functional roles and anatomical locations), we reveal the following neuronal compartments. This classification is supported by similar neuronal functions and/or 3D coordinates in the worm body: the Worm Brain (1st–4th); the Worm Movement Controller (5th–8th); the Worm Information Flow Connector (9th and 10th). It should be noted that the neurons listed below are the names of the neuron sets, for example, VA contains twelve individual neurons VA1–VA12 or ADA contains ADAL and ADAR. For detailed description of cluster elements see Fig. 4.

(a) Cosine similarity measure between WB, WMC and WIFC compartments for the old34,35 and new connectome data25. (b) P-value of overlaps between the FM clusters and ganglia. (c) P-value of overlaps between the FM clusters and functional groups (BM body motor neurons, HM head motor neurons, I interneurons, S sensory neurons, SSM sex specific motor neurons, SM sublateral motor neurons).
-
Worm Brain (1st, 2nd, 3rd and 4th clusters) Similarly to the multifunctional organization of the brain of more complex organisms, we found that neurons in these four clusters had a common anatomical position and were involved in complex multimodal processes26. Based on the corresponding 3D model (see Fig. 4), one can note that the 3rd and 4th clusters are closer to the nose of the worm and the 1st and 2nd clusters are located behind them. This anatomical similarity between the clusters is consistent with their close functions. Accordingly, all polymodal neurons of the head part of the worm (IL1, IL1D, IL1V, OLQD, OLQV, RIM, ASH) and sensory neurons of the anterior ganglion (IL2D, IL2, IL2V, OLL) are located in the 3rd cluster. Wherein bilaterally symmetric neurons split between these two clusters, thus 27 out of 28 neurons of the 3rd cluster are right neurons (R), while 21 out of 29 neurons of the 4th cluster are left ones (L). The majority of interneurons (ADA, RIA, RIB, RIF, RIG, RIH, RIR, RIS, RIV, URB, URX, SAAD, SAAV, AVK, SDQ, SIAD, SIAV, SIBD, SIBV) belong to the 1st and 2nd clusters. Such a “layered” organization is consistent with intuition about how the signals received by the worm’s nervous system should be processed.
-
Worm Movements Controller (5th, 6th, 7th and 8th clusters) These four clusters contain the ventral cord neural group, which is split between them according to the anatomical positions of the neurons. Namely, neurons in the head half fall into the 5th and 6th clusters, while neurons in the tail body half fall into the 7th and 8th clusters (Fig. 4). Together, almost \(83\%\) of neurons in these four clusters belong to the ventral cord (AS, DA, DB, DD, VA, VB, VC, VD, AVE), which motoneurons are located along the entire body of the worm and split exactly into two groups in accordance with which half of the body of the worm these neurons belong to: 5th and 6th clusters correspond to the head part and 7th and 8th clusters are responsible for the tail movements (bright and dark green colors Fig. 4). The remaining \(17\%\) are represented by neurons of the tail ganglia: pre-anal ganglion, and dorsorectal ganglion (PV, LUA, PWV, PDA, PDB, DVA, DVB, DBC) and belong to the 8th cluster, which is responsible for the tail part of the ventral cord (Fig. 3b). Excitatory motor neurons in the ventral cord function as motor rhythm generators and underlie body undulation during reversal and forward movements42. That is why we refer this pair as a worm movements controller. The connection probabilities between these four clusters are reasonably low (\(\approx 5\%\) in average), which is tenable if we interpret them as disjoint parts of the movement control system, and the dense connections between motor neurons from opposite parts of the worm body are not functionally significant.
-
Worm Information Flow Connector (9th and 10th clusters) Almost \(53\%\) of the neurons in these two clusters belong to the lateral ganglion: sensory neurons (ADF, ADL, ASE, ASG, ASH, ASI, AFD, AWA, AWB, AWC, ASJ, ASK), interneurons (AIA, AIB, AIN, AIY, AIZ, AUA, AVJ) and command interneurons (AVA, AVD, PVC). Command interneurons, which are located in the 10th cluster, by definition receive a convergence of integrative sensory inputs and output to a multifarious group of pattern-generating efferent neurons43. This is consistent with the contact probabilities between the 10th cluster and the Worm Movements Controller. For example, there is an evidence that ablation of AVB or AVA command neurons leads to impairment of spontaneous forward or backward movements44, suggesting they are one of the most critical regulators for the directional motion. The distribution of contacts between the 10th command cluster and other clusters clearly shows its significant role in the information flow integration processes: it receives information about the outer environment from the interneurons located in the 1st, 2nd and 9th clusters (Fig. 3c) and coordinates the behavior of the worm through dense contacts with the worm movements controller. Therefore, one can propose that the 10th cluster plays the role of a “command post” coordinating movement of a worm (clusters 5th–8th), and responsible for the implementation of the worm’s motor programs.
The non-backtracking connectome clusters largely correspond to the contactome modules
In two recent studies29,30 a network of \(10^5\) membrane contacts (the contactome) from the C. elegans nerve ring was generated and analysed. The contactome data contains information about spatial neurite contacts, thus providing an important biological benchmark to compare the modules of the structural connectome.
The authors in29,30 have clustered the contactome using classical methods into several neuronal groups or strata that we statistically compare with the flow matrix clusters (see Fig. 5a,b). We find that all clusters of the Worm Brain compartment (1–4) have a significant intersection with four contactome stratas (Anterior, Lateral, Sublateral, Avoidance)29; the p-value of the intersection is \((p \le 10^{-5})\). The same analysis for the second contactome dataset from30 shows a similar result: the Worm Brain clusters intersect significantly with the first three strata.
Interestingly, the Taxis strata29 and Stratum 430 have a statistically significant overlap with our 9th cluster \((p \le 10^{-5})\); our 10th cluster has a statistically significant overlap with Avoidance strata29 and Strata 430 \((p \le 10^{-4})\). These are the clusters comprising the Worm Information Flow Connector with vast majority of the neurons belonging to the head of the worm. Since only head neurons comprised the contactome dataset (nerve ring), the other neurons of the worm were left unassigned there. Accordingly, we see that the clusters 5–8 from the Worm Movements Controller (WMC) have significant intersections with Unclassified and Unassigned strata from the contactome datasets. All together, we conclude that the contactome modules significantly overlap with the connectome clusters of brain neurons that fall into the Worm Brain and Worm Information Flow Connector.

Three-dimensional visualization of the detected clusters in the connectome. On the basis of the complementary analysis of functional roles and anatomical locations, we identify three compartments: WB, WMC, WIFC. Original 3D coordinates were taken from the Caltech Wormbase project41.
Next, we asked whether the same quantitative correspondence between contactome and connectome modules can be established using other clustering algorithms. For that we calculate the AMI score between the corresponding partitions, see Fig. S4 and Table S1. The maximal value of AMI score is achieved for 6 clusters, since the neurons from the other 4 clusters are missing in the contactome dataset. Indeed, as Fig. 6 shows, the clusters from 5 to 8 contain neurons that are left unassigned in both datasets of the contactome. Accordingly, at \(k=6\) the flow matrix yields connectome clusters that demonstrate the best quantitative agreement with the contactome modules among all the clustering methods (flow matrix: AMI = 0.51 for the Brittin et al. and AMI = 0.47 for the Moyle et al.; IMMA algorithm: AMI = 0.4 for the Brittin et al. and AMI = 0.43 for the Moyle et al.; ERMM algorithm: AMI = 0.41 for the Brittin et al. and AMI = 0.36 for the Moyle et al.). Furthermore, as Fig. S4 suggests, the flow matrix clusters appear to correspond better to the contactome modules than the normalized Laplacian and modularity clusters (e.g., the FM clusters overlap by more than 20% stronger with the contactome modules than the clusters produced by the other two spectral methods

This analysis highlights that the non-backtracking algorithm yields interpretable connectome communities that strongly overlap with the contactome modules derived from the neurite spatial contacts. The obtained non-backtracking communities correspond to the neurite contacts much better than the previously reported clusters of C. elegans connectome.
Conclusion
In this paper we performed a detailed analysis and demonstrated applicability of the spectrum of non-backtracking random walks to the problem of the structural connectome clustering on the model example of C. elegans. This clusterization method outperforms traditional clustering methods on simulated models in the regime of low density of connections, virtually circumventing the sparsity issue and related emergence of hubs. Our analysis of the C. elegans connectome has shown that already on a relatively small network size (\(N=279\)) the communities produced by the non-backtracking method—as compared to previously reported modules in the connectome—result in a better clustering quality and biological interpretability in the terms of functions and neurite spatial contacts.
We applied the non-backtracking clustering scheme for two versions of the C. elegans connectome (old34 and new25). While in both datasets \(k=10\) isolated eigenvalues can be seen in the spectrum of the non-backtracking flow matrix, the detectability condition suggests that only in the new connectome all \(k=10\) clusters can be reliably detected (see Fig. 2). Based on the complete C. elegans connectome25, we reveal ten interpretable communities that we further classify into three distinct compartments: (i) four multifunctional head clusters full of ring neurons (’Worm Brain’); (ii) four clusters responsible for movements control in the head and tail halves of the worm (’Worm Movements Controller’); (iii) one cluster made up from the command interneurons and one cluster from the lateral ganglion consisting of sensory neurons and interneurons (’Worm Information Flow Connector’).
Comparison with the recently mapped contactome modules reveals strong interpretability of the non-backtracking communities in terms of the neurite spatial contacts. We find that the ’Worm Brain’ compartment (clusters 1–4) consists of the neurons with axons that project into the anterior part of the nerve ring, while the ’Worm Information Flow Connector’ (clusters 9–10) compartment stores the amphid neurons that project axons into the posterior part of the worm head. Since the other ’Worm Movements Controller’ compartment (clusters 5–8) contains the neurons that are unclassified in the contactome, we conclude that contactome modules strongly reproduce the connectome clusters. Such a result supports Peter’s rule as a principle of synaptic specificity in the C. elegans brain.
Broadly, our study highlights deep interconnections between anatomical locations (metric space embedding), mesoscopic structure (topological embedding) and biological functions of the neurons that determine behaviour of an organism. In order to derive these relationships accurately it is important to precisely resolve the topological modules in the connectome network. In the framework of the one of the simplest model organisms we demonstrate that the cluster analysis of the connectome should be performed by taking into account the intrinsic sparsity of the network, which is addressed by the non-backtracking operator informed by the corresponding detectability thresholds. Given universality of our approach, we believe it can be further extended to connectomes of more complex organisms with larger networks, where the effect of sparsity would be even more dramatic.
Methods
Data
We have worked with old and new open-access data from the C. elegans connectome analysis project34,35 and Cook et al. paper25. Both datasets are updated and revised versions of the wiring data originally published in26. Neuron interactions, locations, sensory endings, and neuromuscular junctions, as well as the structure of the connectome, have been well studied and have been found to be invariant with respect to the type of animal26,27, however, there is now growing concern that the C. elegans connectome is not invariant28,29,31. Connectome 3D model used for the reconstruction of cluster elements anatomical positions was taken from the Caltech Wormbase project41.
The two versions of the connectome (34,35 and25) have significantly different number of edges: in the refined data of Cook et al.25 there is almost twice more synaptic contacts as compared to the Chen et al.34 previous work (6334 vs 2990). The new hermaphrodite connectome from25 is a network with 302 vertices and 6334 edges: 1447 edges are formed by gap junctions only; 4887 contain only chemical synapses. The old hermaphrodite connectome34,35 is a network with 302 vertices and 2990 edges: 796 edges are formed by gap junctions and 1962 contain only chemical synapses.
The entire nervous system is broken down into two large disconnected components and two isolated neurons (CANL, CANR) and additionally the VC06 neuron is isolated in the old connectome data34,35. Twenty of the neurons in one of the components are located within the worm pharynx, which has its own separate nervous system, and the remaining 280 (or 279 for34,35) neurons (excluding two isolated neurons) are located in various ganglia along the worm body. During the preprocessing stage, all connections in the connectome are made undirected and unweighted. Furthermore, we have divided the graph into two subgraphs according to contact types: chemical synapses or gap junctions and analyzed the connectome formed only by the synaptic contacts, because these two types of connections are fundamentally different in nature and their functions are also distinct.
There is a large body of knowledge on individual neurons that produce node-wise features. In this work, we have used the classification of neurons into ten anatomically defined ganglia (posterolateral, ventral, pre-anal, lateral, dorsorectal, dorsal, retrovesicular, ventral cord, anterior and lumbar ganglia) and six functional groups (body motor neurons, head motor neurons, interneurons, sensory neurons, sex specific motor neurons, sublateral motor neurons) from25,45.
As another benchmark for testing biological validity of our clusters, we have used the contactome adjacency matrices29,30, because contactome itself contains information about axon position and metric structure of the C. elegans nervous system. Contactomes have fewer number of described neurons (170 neurons in the29 stratas and 181 in the30), therefore we added all missing neurons to the stratas Unassigned and Unclassified correspondingly. Full contactomes descriptions could be found in the Github folder.
From stochastic block model to non-backtracking random walks
One of the most popular methods for community detection (in particular, of the connectome19) is optimization of modularity. In fact, it can be shown that the generalized modularity functional provides the entropy of a Poisson weighted stochastic block model with quenched degrees (configuration model). Such models, for example, describe the results of single-cell contact counting experiments in chromatin networks, as was shown by us recently24. If the degrees of all vertices \(d_i=\sum _j A_{ij}\) are kept fixed, without additional imposed cluster structure, the expected weight of the edge under random degree-preserving randomization is simply \(P_{ij}=\frac{d_i d_j}{\sum _{i} d_i}\) for \(i \ne j\). Assuming that the stochastic blocks are superimposed over the configuration model, each entry \(A_{ij}\) of the adjacency matrix of the observed network becomes a Poisson random variable with the mean \(P_{ij} w_{rt}\), such that the nodes i and j are assigned to the groups \(G_r\) and \(G_t\), respectively. Thus, the total statistical weight of A conditioned on the cluster probability matrix W, quenched degrees \(d_i\) and group labels \(g_i\) can be factorized into the product of the Poisson probabilities and written down as follows
which produces the following entropy
where \(\gamma\) is some parameter that depends on \(w_{in}\) and \(w_{out}\) of the planted SBM (1) as follows
Clearly, the entropic functional (9) up to the parameter \(\gamma\) is nothing but the modularity functional, which is widely used in clustering tasks, for connectome clustering as well19. It is important to note that generally the parameter \(\gamma\) have to be chosen self-consistently with the cluster parameters of the partition (10), for which the iterative procedure has been recently proposed24.
Modularity optimization has been originally proposed and proved to be useful for clusterization of scale-free networks, since, as we have shown above, it explicitly conserves the scale-free property of the degree distribution under stochastic randomization. Although most of the real-world networks are scale-free, modularity is one of the most popular approaches in spectral clustering. However, if one relaxes the degrees preservation assumption, the background probability becomes uniform \(P_{ij}=p\) and the underlying graph is assumed to be simply a G(N, p) Erdos–Renyi graph. Then the second term in (9) does not depend on cluster labels of the nodes, and maximization of the entropy for a given amount of clusters corresponds to maximization of the adjacency functional
which is trying to maximize the internal weight of the clusters. In a more general problem setting of a manifold learning, one is looking for the optimal representation (embedding) of N vertices in a low-dimensional space described by a set of coordinates \(g_i, i=1,2, \ldots , N\) (suppose, the latent space is one-dimensional for simplicity). As long as close points in the original high-dimensional space should be eventually put close in the latent space, the natural functional to be minimized is
which can be written as a quadratic form of the graph Laplacian, \(L=D-A\)
Of course, a similar functional over latent coordinates can be written for the modularity functional (9) as well.
Thus, we see that statistical inference of the optimal cluster structure is associated with optimization of a certain functional over partition of graph nodes. However, finding the global maximum of (9), (11), (13) is a very difficult computational task. To overcome this difficulty, spectral methods are used, which rely on the fact that the most essential information about the optimal partition is encoded in the first non-trivial eigenvectors of the corresponding operator. Indeed, the quadratic form associated with the manifold learning problem can be approximated by projecting the coordinates to the leading eigenvectors of the operator.
Modularity matrix
In46 the modularity matrix of a graph was defined as
where A is the adjacency matrix, \(d = (d_1, \dots , d_n)^T\) is the degree-vector comprised of the vertices degrees and \(C = \frac{1}{2}\sum _{i=1}^n d_i\) is the total number of edges in the network.
We computed a quantitative measure of modularity for each partition of graphs into several communities, using the standard Newman’s modularity (Q value):
By notation, A is the adjacency matrix of connectome (\(A_{ij} = 1\), if neurons i, j are connected, and 0, otherwise). The degree of each vertex i is given by \(d_i= \sum _{j} A_{ij}\). C is the total number of edges on the connectome graph, equal to \(C=\frac{1}{2}\sum _i d_i\) and \(\delta\) is the Kronecker delta and \(g_i\) is the label of the community to which vertex i is assigned. As we see, (15) is different from the entropic functional (9) by a particular normalization coefficient used.
Laplacian and normalized Laplacian
Laplacian is widely used in spectral manifold learning methods, a framework known as Laplacian Eigenmaps. The graph Laplacian matrix is defined as
where A is the adjacency and D is the diagonal matrix of degrees. Though Laplacian is related to many physical phenomena, such as heat propagation, a more direct connection with random walks is provided by the Normalized Laplacian (or Random Walks Laplacian), \(L_{RW}=D^{-1}L\), which is also frequently used for clustering. Note that \(L_{RW}\) is non-symmetric, however, its spectrum is real. Obviously, \(L_{RW}\) has the same set of eigenvalues as the symmetric normalized Laplacian
Similarity measures
In order to assess the similarity between different partitions and biological benchmarks we use the adjusted mutual information score (AMI), defined as follows. Suppose that we have a set S and two partitions of S: U and V, the elements of the partitions are called clusters. Let us denote the probability that some random object falls into a cluster \(U_{i}\) of U as \(P_{U(i)}\) which is equal to \(\frac{|U_i|}{|S|}\). The entropy calculated for the partition U is equal to \(H(U)=-\sum _{i=1}^R P_U(i)\log P_U(i)\). Using the introduced notation, we can express the mutual information for U and V as
Importantly, this measure of similarity tends to be larger when the two partitions have a larger number of clusters even when we use the same number of elements for clustering. To avoid such biases one can use the adjusted mutual information which is defined as
where \(E\{MI(U,V)\}\) is the expected value of the mutual information of V and U.
Therefore, AMI is 0 when the similarity is equal to its expected value under random permutation of the vertices between the groups and 1 for identical partitions.
Data availability
Additional table with 280 C. elegans neurons and their functions distributed among flow matrix clusters (obtained from old34,35 and new25 connectome data) is available in the Data folder on the Github Project Page (link: https://github.com/kipolovnikov/celegans_nonbacktracking).
Code availability
Source code and data available on GitHub project page.
References
Karrer, B., Newman, M. E. J. & Zdeborová, L. Percolation on sparse networks. Phys. Rev. Lett. 113(20), 208702 (2014).
Hebb, D. O. The first stage of perception: Growth of the assembly. Org. Behav. 4, 60–78 (1949).
Albert, R., Jeong, H. & Barabási, A. L. Internet: Diameter of the world-wide web. Nature 401(6749), 130 (1999).
Broder, A. et al. Graph structure in the web. Comput. Netw. 33(1–6), 309–320 (2000).
Dekker, J., Marti-Renom, M. A. & Mirny, L. A. Exploring the three-dimensional organization of genomes: Interpreting chromatin interaction data. Nat. Rev. Genet. 14(6), 390 (2013).
Ravasz, E., Somera, A. L., Mongru, D. A., Oltvai, Z. N. & Barabási, A. L. Hierarchical organization of modularity in metabolic networks. Science 297(5586), 1551–1555 (2002).
Redner, S. How popular is your paper? An empirical study of the citation distribution. Eur. Phys. J. B 4(2), 131–134 (1998).
Chen, J., Zaïane, O. R., & Goebel, R. Detecting communities in social networks using max-min modularity. in Proceedings of the 2009 SIAM International Conference on Data Mining, 978–989 (2009).
Piccardi, C., Calatroni, L. & Bertoni, F. Communities in Italian corporate networks. Physica A 389(22), 5247–5258 (2010).
Polovnikov, K., Kazakov, V. & Syntulsky, S. Core-periphery organization of the cryptocurrency market inferred by the modularity operator. Physica A 540, 123075 (2020).
Polovnikov, K., Pospelov, N., & Skougarevskiy, D. Ownership concentration and wealth inequality in Russia. in Proceedings of the 31st International Joint Conference on Artificial Intelligence (2022).
Von Luxburg, U. A tutorial on spectral clustering. Stat. Comput. 17(4), 395–416 (2007).
Krivelevich, M. & Sudakov, B. The largest eigenvalue of sparse random graphs. Combin. Probab. Comput. 12(1), 61–72 (2003).
Nadakuditi, R. R. & Newman, M. E. The largest eigenvalue of sparse random graphs. Phys. Rev. Lett. 108(18), 188701 (2012).
Decelle, A., Krzakala, F., Moore, C. & Zdeborová, L. Inference and phase transitions in the detection of modules in sparse networks. Phys. Rev. Lett. 107(6), 065701 (2011).
Newman, M. Finding community structure in networks using the eigenvectors of matrices. Phys. Rev. E 74(3), 036104 (2006).
Norton, H. K. et al. Detecting hierarchical genome folding with network modularity. Nat. Methods 15(2), 119 (2018).
Sales-Pardo, M., Guimera, R., Moreira, A. A. & Amaral, L. A. N. Extracting the hierarchical organization of complex systems. Proc. Natl. Acad. Sci. 104(39), 15224–15229 (2007).
Pan, R. K., Chatterjee, N. & Sinha, S. Mesoscopic organization reveals the constraints governing Caenorhabditis elegans nervous system. PLoS ONE 5(2), e9240 (2010).
Krzakala, F. et al. Spectral redemption in clustering sparse networks. Proc. Natl. Acad. Sci. 110(52), 20935–20940 (2013).
Reichardt, J. & Bornholdt, S. Statistical mechanics of community detection. Phys. Rev. E 74(1), 016110 (2006).
Arenas, A., Fernandez, A. & Gomez, S. Analysis of the structure of complex networks at different resolution levels. N. J. Phys. 10(5), 053039 (2008).
Nechaev, S. K. & Polovnikov, K. Rare-event statistics and modular invariance. Phys. Uspekhi 61(1), 99 (2018).
Polovnikov, K., Gorsky, A., Nechaev, S., Razin, V. & Ulianov, S. V. Non-backtracking walks reveal compartments in sparse chromatin interaction networks. Sci. Rep. 10(1), 1–11 (2020).
Cook, S. J. et al. Whole-animal connectomes of both Caenorhabditis elegans sexes. Nature 571(7763), 63–71 (2019).
White, J. G. et al. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. B 314(1165), 1–340 (1986).
Hall, D. H. & Russell, R. L. The posterior nervous system of the nematode Caenorhabditis elegans: Serial reconstruction of identified neurons and complete pattern of synaptic interactions. J. Neurosci. 11(1), 1–22 (1991).
Yemini, E. et al. Neuropal: A multicolor atlas for whole-brain neuronal identification in C. elegans. Cell 184(1), 272–288 (2021).
Brittin, C. A., Cook, S. J., Hall, D. H., Emmons, S. W. & Cohen, N. A multi-scale brain map derived from whole-brain volumetric reconstructions. Nature 591(7848), 105–110 (2021).
Moyle, M. W. et al. Structural and developmental principles of neuropil assembly in C. elegans. Nature 591(7848), 99–104 (2021).
Witvliet, D. et al. Connectomes across development reveal principles of brain maturation. Nature 596(7871), 257–261 (2021).
Nagano, T. et al. Single-cell hi-c reveals cell-to-cell variability in chromosome structure. Nature 502(7469), 59–64 (2013).
Pavlovic, D. M., Vértes, P. E., Bullmore, E. T., Schafer, W. R. & Nichols, T. E. Stochastic blockmodeling of the modules and core of the Caenorhabditis elegans connectome. PLoS ONE 9(7), e97584 (2014).
Chen, B. L., Hall, D. H. & Chklovskii, D. B. Wiring optimization can relate neuronal structure and function. Proc. Natl. Acad. Sci. 103(12), 4723–4728 (2006).
Varshney, L. R., Chen, B. L., Paniagua, E., Hall, D. H. & Chklovskii, D. B. Structural properties of the Caenorhabditis elegans neuronal network. PLoS Comput. Biol. 7(2), e1001066 (2011).
Decelle, A., Krzakala, F., Moore, C. & Zdeborová, L. Asymptotic analysis of the stochastic block model for modular networks and its algorithmic applications. Phys. Rev. E 84(6), 066106 (2011).
Newman, M. Spectral community detection in sparse networks. arXiv:1308.6494 (2013).
Newman, M. E. J. Modularity and community structure in networks. Proc. Natl. Acad. Sci. 103(23), 8577–8582 (2006).
Singh, A. & Humphries, M. D. Finding communities in sparse networks. Sci. Rep. 5(1), 1–7 (2015).
Rosvall, M., Axelsson, D. & Bergstrom, C. T. The map equation. Eur. Phys. J. Spec. Top. 178(1), 13–23 (2009).
Harris, T. W. et al. Wormbase: A comprehensive resource for nematode research. Nucleic Acids Res. 38(suppl1), D463–D467 (2010).
Wen, Q., Gao, S. & Zhen, M. Caenorhabditis elegans excitatory ventral cord motor neurons derive rhythm for body undulation. Philos. Trans. R. Soc. B 373(1758), 20170370 (2018).
Kawano, T. et al. An imbalancing act: Gap junctions reduce the backward motor circuit activity to bias C. elegans for forward locomotion. Neuron 72(4), 572–586 (2011).
Wicks, S. R., Roehrig, C. J. & Rankin, C. H. A dynamic network simulation of the nematode tap withdrawal circuit: Predictions concerning synaptic function using behavioral criteria. J. Neurosci. 16(12), 4017–4031 (1996).
Altun, Z. F. & Hall, D. H. Handbook of C. elegans Anatomy. WormAtlas.http://www.wormatlas.org/handbook/contents.htm (2005).
Newman, M. E. J. & Girvan, M. Finding and evaluating community structure in networks. Phys. Rev. E 69(2), 026113 (2004).
Acknowledgements
All authors thank A. Gorsky and E. Burnaev for stimulating discussions on the project and A. Moiseeva for help with illustrations. This work was supported by the Russian Science Foundation, Grant №21-75-30024. AO, AC and KP acknowledge the support of Idea Foundation grant in collection and analysis of the connectome data.
Author information
Authors and Affiliations
Contributions
K.P. conceptualized the study and design. Data collection and analysis was performed by A.O. and A.C. All authors participated in writing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Onuchin, A.A., Chernizova, A.V., Lebedev, M.A. et al. Communities in C. elegans connectome through the prism of non-backtracking walks. Sci Rep 13, 22923 (2023). https://doi.org/10.1038/s41598-023-49503-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-49503-5
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
