
Abstract
Climate change is reshaping global ecosystems at an unprecedented rate, with major impacts on biodiversity. Therefore, understanding how organisms can withstand change is key to identify priority conservation objectives. Marine ectotherms are being extremely impacted because their biology and phenology are directly related to temperature. Among these species, sea turtles are particularly problematic because they roam over both marine and terrestrial habitats throughout their life cycles. Focusing on green turtles (Chelonia mydas) in the Mediterranean Sea, we investigated the future potential changes of nesting grounds through time, assuming that marine turtles would shift their nesting locations. We modeled the current distribution of nesting grounds including both terrestrial and marine variables, and we projected the potential nesting distribution across the Mediterranean basin under alternative future greenhouse gas emission scenario (2000–2100). Our models show an increase in nesting probability in the western Mediterranean Sea, irrespective of the climate scenario we consider. Contrary to what is found in most global change studies, the worse the climate change scenario, the more suitable areas for green turtles will potentially increase. The most important predictors were anthropogenic variables, which negatively affect nesting probability, and sea surface temperature, positively linked to nesting probability, up to a maximum of 24–25 °C. The importance of the western Mediterranean beaches as potential nesting areas for sea turtles in the near future clearly call for a proactive conservation and management effort, focusing on monitoring actions (to document the potential range expansion) and threat detection.
Similar content being viewed by others

Introduction
Anthropogenically induced climate change is quickly reshaping the world’s ecosystems with major impacts on biodiversity1. Marine ecosystems have been particularly hit, with roughly 66% of the global oceans being affected by multiple human pressures2. The consequences for marine biodiversity are severe. The Ocean Living Planet Index indicates that global ocean fish stocks have been overexploited by 29%, ocean species have decreased by 39%, and world coral reefs have decreased by 50%3. Among all threats affecting marine ecosystems, climate change is currently considered one of the most important4, with 14% of all marine species already heavily impacted by climatic factors5 and with increasingly severe impacts over time. The extreme heat waves in 2019 hit 57% of the ocean surface waters, with significant impacts on marine ecosystems and their ability to provide resources to coastal communities6.
Future projections return an overly grim outcast, even considering the near future, with the Mediterranean Sea representing one of the most vulnerable regions in the world when considering the impacts of global warming7. The Mediterranean has always been hugely exploited by humans, with important and potentially negative interactions between human activities and climate changes. The basin currently hosts more than 500 million inhabitants, a third of whom live along the coasts, and is the first tourist destination in the world with 360 million visitors per year8,9. The combination of climate change and human impacts clearly generate the starting baseline for a biodiversity collapse10, especially for species exploiting at the same time marine and terrestrial habitats as sea turtles.
Many species of vertebrates (birds, mammals, and reptiles) roam over both the marine and terrestrial realm and are potentially very sensitive to global changes. In fact, they exploit very different habitats during their life cycles, often migrating over long distances to reach nesting, feeding, and reproductive grounds11,12, being therefore exposed to multiple and often uncoupled threats. Many of these species (e.g., Mirounga leonina, Aptenodytes patagonicus) are responding to climate change by changing their phenology and/or distribution range13,14, while others (e.g., Fratercula arctica) seem unable to adapt fast enough11.
Single species responses are expected to lead to changes also in the pattern of species richness15, in community and ecosystem structure16, and in ecosystem services17. Although negative impacts are predominant, climate changes could also have positive effects on marine biodiversity. For instance, increased temperatures and CO2 concentrations seem to accelerate primary biomass production18, while milder winters might increase survival probabilities of many threatened species in temperate regions19,20.
Understanding the impact of global changes in marine systems is particularly challenging given their complexity and the huge number of factors that should be considered. Umbrella and/or keystone species can clearly represent a good proxy for global change effects, and marine turtles are particularly interesting in this context. In fact, during their life cycles, they exploit different habitats, nesting on the terrestrial realm and then using both the neritic and oceanic habitats, being potentially exposed to multiple, unrelated, and very different threats11. Being highly mobile and migratory species, they have the potential to adapt to changing conditions21 and can respond to changes by shifting their spatiotemporal distribution and/or changing their nesting and foraging behaviour22,23. However, sea turtles have a peculiar reproductive physiology, and therefore are particularly sensitive to climatic conditions during eggs incubation and development24, when climate change can directly affect sex ratio in hatchlings25. In fact, turtles have temperature-dependent sex determination, with females associated with high incubation temperatures (roughly above 29 °C)26 and males associated with lower temperatures, but extreme hot temperatures of the nests, ranged from 28.4 to 33.5 °C, can lead to decreased hatchling rates or even to complete nest failures27,28,29. Some authors have proposed a positive effect of increasing temperatures on sea turtle populations, suggesting that more female hatchlings would automatically translate into more breeding females, more nests and, consequently, population growth30,31. However, an excessive shortage of males can produce negative consequences (e.g., reductions in egg fertility, which may threaten population viability)32 and extreme temperatures may even lead to embryonic mortality33. Climate changes can also impact adult turtles. Being ectothermic, seawater temperature can radically impact physiology and behaviour of turtles34. In fact, below a thermal threshold of around 10 − 15 °C (depending on the species) their metabolic rates decrease, and turtles become less mobile35,36,37. On the other side, higher temperatures can induce indirect stress by promoting the growth of pathogens, with clear consequences on the biology of populations38. Overall, different analyses have predicted significant losses in nesting habitat for sea turtle populations in the Great Barrier Reef, Greece, and the Hawaiian Islands, even exceeding 65% of the areas suitable for nesting39,40,41,42. However, these projections, catastrophic in some cases, are often focused exclusively on climate and do not consider other important factors which can contribute to an even worse outcome. For example, although the relationship between climate change and sea level rise and coastal storms in the Mediterranean Sea is very weak43, the flooding of beaches, associated with an increasing prevalence and intensity of storms and sea level rise, will further reduce both hatching success and availability of nesting habitats27,44.
Currently, most research efforts in relation to climate change impacts in the Mediterranean have been related to loggerhead sea turtles (Caretta caretta), focusing on the current and potential distribution23, ecology45, and conservation of the species46. Much less is known about green turtles (Chelonia mydas), for which only a general overview of the ecology and conservation status exists47. A recent analysis48, considering only bioclimatic variables, found no appreciable change in the potential suitability for green turtle nesting grounds under different global change scenarios in the Mediterranean. As a species tightly linked to sea surface temperature, with an optimum temperature range for functioning > 25 °C49,50,51,52,53, these results are surprising. In fact, green turtles are strongly adapted to hot conditions with a distribution that is mostly tropical. In the Mediterranean, they are limited to the hottest areas, and particularly to the Levantine basin (Turkey, Syria, Cyprus, Lebanon, Israel, Egypt). Their nesting grounds are all in the easternmost part of the Mediterranean, with the vast majority of the nests located in Cyprus and Turkey54. Foraging areas extend to Greece and Libya47, while a few wandering green turtles can be occasionally found in the Adriatic Sea and very rarely in the western basin.
Using a species distribution modelling approach fully focused on the species ecology, and therefore including both marine and terrestrial variables, anthropogenic and natural factors, we provide here a thorough investigation of the impact of future climate change scenarios on the distribution of nesting grounds for green turtles in the Mediterranean basin. Our main hypothesis, contrary to what is available in the literature for the same species48, is that nesting ground availability for green turtles will potentially increase in the Mediterranean following hotter summers and winters, shifting their nesting locations over extended periods ranging from years to decades in response to climate change-induced warming and its associated impacts. If our hypothesis is confirmed, our results can be extremely important for a proactive management approach in which the conservation of the species and the management of potential conflicts with human coastal activities is planned and enforced well before the species’ arrival.
Methods
Nesting locations
We built a database on green turtles’ nesting locations searching Scopus, Web of Science, and Google Scholar with the following keywords: “sea turtle*” OR “marine turtle*” AND “nesting habitat” AND “Mediterranean”. We combined the list of references obtained from the 3 search engines and obtained a first set of 37,580 references. We also downloaded all documents referred in the first set of references and obtained a total of 38,590 peer reviewed papers and technical reports. We scanned all documents, and we retained only documents dealing with green turtles in the Mediterranean, looking for indications on nest location, excluding all dubious observations (nest attempted, nest possible but not proved, false crawls) and duplicate records (same date and same beach reported in different documents).
Environmental data and scenarios
To model the potential distribution for nesting sites we considered environmental, anthropogenic, and climatic variables spanning both the marine and the terrestrial realm. We considered a total of 14 variables directly linked to nesting ecology in green turtles and summarized each variable over May–August to specifically include the nesting season55 (Table 1).
We considered sea surface temperature and salinity because green turtles show a strong affinity to areas with warmer sea temperatures in the Mediterranean Sea, thriving at SST of roughly 27–29 °C and with highly variable salinity (range 1.2–35.5‰)49,51,52,53. Furthermore, the spatial distribution of adults is closely tied to sea surface temperature62,63, which can influence their selection of nesting sites64,65. We also considered several variables related to water chemistry that may positively and/or negatively influence nesting site selection66,67. Excessive concentrations of nitrogen, often used in agriculture and spilled in sea waters, cause water pollution, eutrophication, and acidification68, adversely affecting sea turtle livelihoods, foraging, and nesting habitats69. Dissolved oxygen concentration, on the contrary, was chosen as a parameter to determine the health of water bodies70. Poor water quality can negatively impact sea turtle health, altering their distribution71. On land, green turtle nests are clearly related to the presence of sandy beaches but are adversely affected by urbanization and other types of anthropogenic developments which usually besets sandy shores72. In fact, green turtles are often forced to use sub-optimal nesting habitats, competing for their nests with tourists and coastal residents73,74.
To limit the negative effects of multicollinearity on model calibration75, we performed a Variance Inflation Factor analysis (VIF) on the initial set of 14 variables and we excluded all variables with a VIF ≥ 3 obtaining a final database with 10 variables (Table 1).
We downloaded all available future layers at 2100 for the same variables from Bio-Oracle 76,77 (~ 9.2 km resolution) and from the SEDAC (Socioeconomic Data and Applications Center) 60 (1 km resolution). For future projections, we considered the following RCP (Representative Concentration Pathway) from CMIP-5 (Coupled Model Intercomparison Project): RCP2.6, a peak-and-decline scenario ending in very low greenhouse gas concentration levels by the end of the twenty-first century; RCP4.5 and RCP6.0, both with greenhouse gas concentrations stabilizing; and RCP8.5, a scenario of increasing emissions over time, leading to high greenhouse gas concentration levels77.
Green turtles are highly mobile animals, clearly influenced in their movements and in their ecology by environmental conditions occurring over large areas. Therefore, we applied to each variable considered in the model a moving window function in ArcGIS pro (ESRI ©) to summarize in each pixel the environmental characteristics of the neighbouring areas. For marine variables we considered a 20 km radius window to account for the mobility of the animals that typically swim within a mean distance of 20 km from beaches during nesting season78,79. For terrestrial variables we chose a 2 km radius to account for the limited movements when the animals are close to the shore (± 2 km)80. For all variables, the focal function assigned to the central pixel of the moving window the mean value calculated over all pixels inside the window.
Modelling approach
To model the current species distribution of green turtles’ nesting grounds over the Mediterranean shores we used an information theory approach81. We calibrated the model using a maximum entropy algorithm (Maxent v. 3.4.1)82 considering all available nesting locations and 10,000 random background points sampled along the coasts of the eastern Mediterranean Sea (Levantine Sea, Tunisian Plateau, and Aegean Sea)83. We tuned model parameters according to Muscarella et al.84, with regularization multiplier ranging from 0.5 to 5.0 (0.5 increments) and with five different combination of feature classes: linear only, linear and quadratic; linear, quadratic, and hinge; linear, quadratic, hinge, and product; linear, quadratic, hinge, product, and threshold. Based on Li et al.85, we estimated the corrected Akaike Information Criteria (AICc) for each combination of feature classes and regularization multipliers. Variable contribution for each explanatory variable was evaluated using a jackknife approach86.
We evaluated the predictive capacity of the final model using a split-plot approach: we randomly split the set of occurrences and background points and we used 80% of the occurrences/background to calibrate a model and the remnant 20% for evaluation. We repeated the same process for 10 replicates measuring the AUC value (Area Under the receiver operating characteristic Curve). We obtained the average and standard deviation AUC and tested the statistical significance87.
There are several ways to correct sampling bias. Spatial filtering may not be helpful when there are only a few presence points88. For this reason, we utilized clumping, which reduces the training sample size. Depending on the heterogeneity of the surrounding environment and the selected spatial resolution, it determines which areas in the climate landscape fall outside the range represented by the training data88.
We projected the final model over the entire coastline of the Mediterranean Sea both under current conditions and future scenarios. We converted the continuous models into binary predictions of presence and absence using a threshold corresponding to the 10th percentile of probability values measured over the occurrences, and we used the binary model to calculate the percentages of changes in species presences from current conditions to future scenarios for the entire Mediterranean Sea.
Ethical approval
All methods were carried out in accordance with relevant guidelines and regulations.
Results
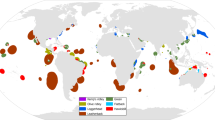
From the initial set of 38,590 references, only 23 reported information on green turtle nesting grounds, covering 5 countries, all in the eastern Mediterranean basin (Table 2). Overall, we obtained 178 nesting locations of green turtles going from 1982 to 2019, located from Turkey to the southern boundaries of Israel (Fig. 1). All nests were in the Levantine Sea ecoregion (Fig. 1), except a single nest in the Aegean Sea (but still in Turkey) collected in 2000.

Nesting locations of Chelonia mydas in the Mediterranean basin from 1982 to 2019.
Current and future nesting model
The final potential nesting probability model (AICc values in Appendix A, Supplementary material, Table A.2) showed a very good predictive power with a mean AUC = 0.943 (± 0.020; p value ≤ 0.001). The average sea surface temperature, salinity, and human population density explained 98.8% of the variance of the entire model. Sea surface temperature was by far the most important variable, with salinity and human population density having a marginal effect on the potential distribution of nests.
The probability of nesting increases with increasing sea surface temperatures and salinity (Fig. 2), being basically zero for temperatures below 21 °C and increasing up to a plateau corresponding to slightly more than 23 °C and 38‰ of salinity. On the contrary, the probability of nesting decreased with higher human population densities (Fig. 2).

Marginal response curves for three variables: (a) sea surface temperature (sst), (b) salinity (psu), (c) human population density (pop). The curve represents the mean response to green turtle nesting probability, and the shaded areas represent one standard deviation both calculated over 10 replicates.
Considering current climate, the highest probabilities of nesting are limited to the eastern Mediterranean basin, and particularly to the southern shores of Turkey, to Cyprus, the middle East, and to the easternmost coasts of Egypt. Overall, only 5% of the Mediterranean shores are currently suitable as nesting grounds for the green turtle (Fig. 3). Whatever future scenario we consider, suitability for nesting will increase responding positively to increasing temperatures. Considering RCP2.6, RCP4.5, and RCP6.0 at 2100 the increase in suitability will be limited to the coasts of Turkey, Middle East, and North Africa, going westward up to the southern Tunisia (Fig. 3). According to RCP8.5, suitability will increase to cover 67% of the Mediterranean basin, with the north African coasts being almost completely suitable (up to Algeria), and with the Italian coasts representing good habitat for the species (Fig. 3).

Distribution maps of environmental suitability for Chelonia mydas nesting habitat in the Mediterranean Sea from current, with only 5% of suitable areas, to future at 2100, with a 67% increase in the RCP scenario considering the highest levels of CO2 emissions. Percentages of change are calculated over the binary outputs (Appendix A in Supplementary material, Table A.3).
Discussion
Overall, in the vast majority of biological systems the impacts of ongoing and future climate changes are driving towards a huge loss of biodiversity89,90, with consequences also for human wellbeing91. Only a limited number of studies highlight a potentially positive effect, for example the increase in average carbon dioxide levels over the past century positively affected plant growth rates92,93 and primary biomass production18. In this study we have shown that future climate change can potentially lead to an increase in the nesting distribution of green turtles, which theoretically should expand their nesting ground towards the western Mediterranean Sea.
Green turtles have a global distribution that spans primarily the tropical, subtropical, and temperate regions of the Atlantic, Pacific, and Indian Oceans, with a small population also in the Mediterranean Sea66,94. Here, their nesting grounds have always been limited to the Levantine basin, with 99% of nesting records limited to Turkey and Cyprus, and with occasional nesting along the shores of Egypt, Israel, and Lebanon95. During the first half of the twentieth century, the Mediterranean population of green turtles have declined dramatically due to loss or deterioration of nesting beaches, adverse pressure from incidental catch in marine fisheries (particularly trawling), and marine pollution96. Most conservation efforts have been concentrated on creating protected areas to safeguard nesting beaches, controlling beach nourishment, and minimizing human influences. All these efforts have certainly contributed to a sharp reduction in these problems, and monitoring data now show an increase in nesting activities and a higher number of mature females in the Mediterranean97. These changes are still limited to the eastern Mediterranean basin, but vagrant individuals are constantly seen in the western Mediterranean in the last few years98,99.
The potential distribution we obtained with our model confirms that the current distribution of nesting grounds for the green turtle is limited to the easternmost part of the Mediterranean basin, the only part where SST is optimal100. The model closely follows existing nests, with only marginal suitability along the Egyptian shores where no nest has been recorded. We found a very narrow interval of sea surface temperatures that defines a beach as suitable for nesting, with optimal nesting grounds corresponding to a SST of 25 °C in front of the shore. This value corresponds strikingly well with the results obtained from empirical field studies, which suggests that female turtles during the nesting period actively select waters with temperatures > 25 °C49,50,51,52,53.
In our model's future projections suitable areas increase in the western basin, and the temperature increase reaches a plateau while still remaining below 26 °C. Since 1978 the trends in SST in the Mediterranean Sea increased by 1.4 °C101,102, and the basin is constantly warming by 0.35 °C per decade, especially in spring by 0.38 °C, followed by 0.32 °C in summer103, mostly in the Eastern Mediterranean104. All future scenarios project an increasingly hot Mediterranean Sea by the end of the twenty-first century, ranging from an average annual increase of 0.45 °C for the RCP2.6 scenario to 2.56 °C for the RCP8.5 scenario. In each scenario, the summer, a key season for sea turtles, displayed the maximum warning trend103. Our model will continue to predict high suitability even if temperatures are excessively high. Therefore, we recommend that in situations with a strong sampling bias towards some regions or environmental features, caution should be exercised to avoid over-interpreting the results. Additionally, it is advisable to reduce spatial clumping of records in datasets used for Maxent model calibration105.
Contrary to Arslan et al.48, who found no significant differences in the suitability of nesting areas for green turtles from the past to the future, we found that nesting habitat for the green turtle in the Mediterranean will increase in the future irrespective of the climate scenario we considered, and the increase in suitability is going to be stronger for more extreme scenarios. This discrepancy could possibly result from the different explanatory variables considered. However, it’s very important to consider an information theoretic approach or another type of variable reduction approach otherwise overfitting problems could easily drive the modelling results106, leading to a model that fit random noise in the data107 instead of ecological signals.
Clearly, the capacity of species to respond to environmental and climate changes depends on a combination of different elements, going from phenological plasticity to range shifts, and ultimately to rapid evolution of traits better suited to new conditions108. Moreover, marine turtles have persisted through dramatic climate change events during their past evolutionary history, clearly demonstrating the ability to adapt to changing conditions64,109. It is true, however, that current climate change is occurring at unprecedented speed110, questioning the evolutionary capacities of a k-selected species like the green turtle111. On the contrary, we do have empirical evidence that marine turtles, thanks to their high mobility, may respond to climatic changes by shifting their distribution range towards climatically suitable areas23,112,113. In fact, marine turtles may change the distribution of their nesting grounds, nest-site choice, and nest depth in the sand, adapting in situ by adjusting their pivotal temperature, and nesting in cooler months114,115,116,117,118. Moreover, sporadic nesting is thought to be a mechanism that enhances the ability of sea turtles to disperse and adapt to environmental changes, and new nesting populations can form in new regions where sporadic nesting is observed with increasing temperatures119. Indeed, to date more green turtle individuals have been sighted in the waters of the western Mediterranean, and the factor that has likely contributed to increased number of green turtles is rising sea temperatures98,99.
Range shifts may offer one of the most successful responses to climate change in marine organisms, and in marine turtles in particular120. However, especially considering human dominated landscapes like the Mediterranean basin, a high mobility may result in an increased exposure to threats, such as coastal development. In fact, around 40% of the total Mediterranean coastal zone are nowadays developed, with a projected increase with further coastal developments121. The human population in coastal areas grew from about 100 million in 1980 to 150 million in 2005. It could reach 200 million by 20308. Coastal development, human presence, and associated pressures can modify marine turtle nesting environment and affect females ‘reproductive output’122,123. The direct and indirect impacts of coastal development on marine turtles can potentially reduce their ability to use and colonize new areas124. Driving on the beach and the use of heavy machinery for beach cleaning purposes are common practices and are responsible for alterations in sand characteristics and the destruction of turtle clutches66. Beach furniture, sports courts, and artificial lighting on some nesting beaches reduce the habitat available for nesting, prevent females from accessing suitable nesting sites and cause disorientation of hatchlings through light pollution125. In addition, people on the beach at night may disrupt nesting activity, causing the abandonment of nesting attempts or the destruction through trampling of incubating nests125.
In conclusion, our results show a potential expansion of Chelonia mydas in the western Mediterranean in response to climate change. Therefore, as soon as green turtle arrives in the west, management efforts should focus on enhancing marine turtle’s resilience to changing environmental conditions by mitigating other non-climatic threats they currently experience112,126. The choice of a particular beach as nesting ground it is also negatively influenced by urbanization and other types of development, which should be included in future studies. To better inform future management of marine turtles, there is the need to understand future threats, as well as to couple predictions of marine turtle distribution with an assessment of exposure to these threats127.
Data availability
Data will be made available on request to corresponding author: chiara.mancino@uniroma1.it.
References
Tang, K. H. D. Implications of climate change on marine biodiversity. Glob. J. Agric. Soil Sci. 1(1), 1–6 (2020).
Díaz, S. M., Settele, J., Brondízio, E., Ngo, H., Guèze, M., Agard, J., & Zayas, C. (2019). The Global Assessment Report on Biodiversity and Ecosystem Services: Summary for Policy Makers.
Tanzer, J. et al. Living blue planet report Species (Habitats and Human Well-Being, 2015).
Hoegh-Guldberg, O. & Bruno, J. F. The impact of climate change on the world’s marine ecosystems. Science 328(5985), 1523–1528 (2010).
Luypaert, T., Hagan, J. G., McCarthy, M. L. & Poti, M. Status of marine biodiversity in the Anthropocene. In YOUMARES 9-The Oceans: Our research, our future (ed. Poti, M.) 57–82 (Springer, 2020).
Tanaka, K. R. & Van Houtan, K. S. The recent normalization of historical marine heat extremes. PLOS Clim. 1(2), e0000007 (2022).
Hilmi, N., Ali, E., Carnicer Cols, J., Cramer, W., Georgopoulou, E., Le Cozannet, G., & Tirado, C. IPCC AR6 WGII Cross-Chapter Paper 4: Mediterranean Region. In EGU General Assembly Conf. Abstracts (pp. EGU22–10590) (2022).
Bleu, P. State of Mediterranean Forests 2018 (Food & Agriculture Org, 2019).
Camiñas-Hernández, J.A., Kaska, Y., Hochscheid, S., Casale, P., Panagopoulou, A., Báez, J.C., Otero, M.D.M., Numa, C. and Alcázar, E., Conservation of marine turtles in the Mediterranean sea [brochure]. Centro Oceanogr. Málaga. (2020)
He, Q. & Silliman, B. R. Climate change, human impacts, and coastal ecosystems in the Anthropocene. Curr. Biol. 29(19), R1021–R1035 (2019).
Häkkinen, H., Petrovan, S. O., Sutherland, W. J. & Pettorelli, N. Terrestrial or marine species distribution model: Why not both? A case study with seabirds. Ecol. Evol. 11(23), 16634–16646 (2021).
Blondin, H. E. et al. Land-dependent marine species face climate-driven impacts on land and at sea. Mar. Ecol. Progr. Ser. 699, 181–198 (2022).
Narayanaswamy, B. E. & Foley, M. 3.9 Impacts and effects of ocean warming on deep sea communities. Explain Ocean Warm Causes Scale Effects Conseq. 199, 59895456 (2016).
Sanford, E., Sones, J. L., García-Reyes, M., Goddard, J. H. & Largier, J. L. Widespread shifts in the coastal biota of northern California during the 2014–2016 marine heatwaves. Sci. Rep. 9(1), 1–14 (2019).
Chust, G. et al. Are Calanus spp. shifting poleward in the North Atlantic? A habitat modelling approach. ICES J. Mar. Sci. 71(2), 241–253 (2014).
Bryndum-Buchholz, A. et al. Twenty-first-century climate change impacts on marine animal biomass and ecosystem structure across ocean basins. Glob. Change Biol. 25(2), 459–472 (2019).
Cheung, W. W. L. et al. Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Glob. Change Biol. 16, 24–3510 (2010).
Carozza, D. A., Bianchi, D. & Galbraith, E. D. Metabolic impacts of climate change on marine ecosystems: Implications for fish communities and fisheries. Glob. Ecol. Biogeogr. 28(2), 158–169 (2019).
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W. & Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 15(4), 365–377 (2012).
Al-Tawaha, A. R. M., Al-Tawaha, A., Sirajuddin, S. N., McNeil, D., Othman, Y. A., Al-Rawashdeh, I. M., & Al-Taey, D. K. A. Ecology and Adaptation of Legumes Crops: A Review. In IOP Conf. Series: Earth and Environmental Science (Vol. 492, No. 1, p. 012085). (IOP Publishing, 2020).
Patrício, A. R., Hawkes, L. A., Monsinjon, J. R., Godley, B. J. & Fuentes, M. M. Climate change and marine turtles: Recent advances and future directions. Endanger. Species Res. 44, 363–395 (2021).
Chatzimentor, A., Almpanidou, V., Doxa, A., Dimitriadis, C. & Mazaris, A. D. Projected redistribution of sea turtle foraging areas reveals important sites for conservation. Clim. Change Ecol. 2, 100038 (2021).
Mancino, C., Canestrelli, D. & Maiorano, L. Going west: Range expansion for loggerhead sea turtles in the Mediterranean Sea under climate change. Glob. Ecol. Conserv. 38, e02264 (2022).
Girondot, M. & Kaska, Y. A model to predict the thermal reaction norm for the embryo growth rate from field data. J. Therm. Biol. 45, 96–102 (2014).
Roberts, H. P. et al. Is the future female for turtles? Climate change and wetland configuration predict sex ratios of a freshwater species. Glob. Change Biol. 29(10), 2643–2654 (2023).
Ackerman, R. A. The nest environment and the embryonic development of sea turtles. In The Biology of Sea Turtles (eds Lutz, P. L. & Musick, J. A.) 83–106 (CRC Press, 1997).
Pike, D. A. Forecasting the viability of sea turtle eggs in a warming world. Glob. Change Biol. 20(1), 7–15 (2014).
Turkozan, O., Almpanidou, V., Yılmaz, C. & Mazaris, A. D. Extreme thermal conditions in sea turtle nests jeopardize reproductive output. Clim. Change 167(3–4), 30 (2021).
Türkozan, O., Yılmaz, C., Almpanidou, V., Godfrey, M. H. & Mazaris, A. D. Thermal conditions of green turtle (Chelonia mydas) nests in the largest rookery in the eastern Mediterranean. Endanger Species Res. 50, 63–73 (2023).
Boyle, M., Hone, J., Schwanz, L. E. & Georges, A. Under what conditions do climate-driven sex ratios enhance versus diminish population persistence?. Ecol. Evol. 4(23), 4522–4533 (2014).
Hays, G. C., Mazaris, A. D., Schofield, G. & Laloë, J. O. Population viability at extreme sex-ratio skews produced by temperature-dependent sex determination. Proc. R. Soc. B Biol. Sci. 284(1848), 20162576 (2017).
Hays, G. C., Laloë, J. O., Lee, P. L. & Schofield, G. Evidence of adult male scarcity associated with female-skewed offspring sex ratios in sea turtles. Curr. Biol. 33(1), R14–R15 (2023).
Türkozan, O., Almpanidou, V., Yılmaz, C. & Mazaris, A. D. Extreme thermal conditions in sea turtle nests jeopardize reproductive output. Clim. Change 167(3–4), 30 (2021).
Milton, S. L. & Lutz, P. L. Physiological and genetic responses to environmental stress. Biol. Sea Turt. 2, 163–197 (2003).
Hochscheid, S., Bentivegna, F. & Speakman, J. R. Long-term cold acclimation leads to high Q10 effects on oxygen consumption of loggerhead sea turtles Caretta caretta. Physiol. Biochem. Zool. 77(2), 209–222 (2004).
Southwood, A. L., Reina, R. D., Jones, V. S., Speakman, J. R. & Jones, D. R. Seasonal metabolism of juvenile green turtles (Chelonia mydas) at Heron Island, Australia. Can. J. Zool. 84(1), 125–135 (2006).
Hochscheid, S., Bentivegna, F., Bradai, M. N. & Hays, G. C. Overwintering behaviour in sea turtles: Dormancy is optional. Mar. Ecol. Progr. Ser. 340, 287–298 (2007).
Plotkin, P., Lutz, P. L., Musick, J. A. & Wyneken, J. Adult migrations and habitat use. Biol. Sea Turt. 2, 225–241 (2002).
Baker, J. D., Littnan, C. L. & Johnston, D. W. Potential effects of sea level rise on the terrestrial habitats of endangered and endemic megafauna in the Northwestern Hawaiian Islands. Endanger. Species Res. 2, 21–30 (2006).
Fish, M. R. et al. Construction setback regulations and sea-level rise: Mitigating sea turtle nesting beach loss. Ocean Coast. Manag. 51(4), 330–341 (2008).
Fuentes, M. M. P. B., Limpus, C. J., Hamann, M. & Dawson, J. Potential impacts of projected sea-level rise on sea turtle rookeries. Aquat. Conserv. Mar. Freshw. Ecosyst. 20(2), 132–139 (2010).
Katselidis, K. A., Schofield, G., Stamou, G., Dimopoulos, P. & Pantis, J. D. Employing sea-level rise scenarios to strategically select sea turtle nesting habitat important for long-term management at a temperate breeding area. J. Exp. Mar. Biol. Ecol. 450, 47–54 (2014).
Ibáñez, C. & Caiola, N. Sea-level Rise, Marine Storms and the Resilience of Mediterranean Coastal Wetlands: Lessons Learned from the Ebro Delta (Marine and Freshwater Research, 2021).
Van Houtan, K. S. & Bass, O. L. Stormy oceans are associated with declines in sea turtle hatching. Curr. Biol. 17(15), R590–R591 (2007).
Almpanidou, V. et al. Foraging grounds of adult loggerhead sea turtles across the Mediterranean Sea: Key sites and hotspots of risk. Biodiv. Conserv. 23, 1–18 (2022).
Abalo-Morla, S. et al. Assessing the use of marine protected areas by loggerhead sea turtles (Caretta caretta) tracked from the western Mediterranean. Glob. Ecol. Conserv. 38, e02196 (2022).
Casale, P. et al. Mediterranean sea turtles: Current knowledge and priorities for conservation and research. Endanger. Species Res. 36, 229–267 (2018).
Arslan, G., Ertürk, A. & Candan, O. Predicting the distribution of green turtle nesting sites over the Mediterranean with outcoming climate driven changes. J. Nat. Conserv. 71, 126320 (2023).
Hays, G. C. et al. Movements of migrating green turtles in relation to AVHRR derived sea surface temperature. Int. J. Rem. Sens. 22(8), 1403–1411 (2001).
Hays, G. C. et al. Water temperature and internesting intervals for loggerhead (Caretta caretta) and green (Chelonia mydas) sea turtles. J. Thermal Biol. 27(5), 429–432 (2002).
Hays, G. C., Broderick, A. C., Glen, F. & Godley, B. J. Climate change and sea turtles: A 150-year reconstruction of incubation temperatures at a major marine turtle rookery. Glob. Change Biol. 9(4), 642–646 (2003).
Esteban, N. et al. A global review of green turtle diet: sea surface temperature as a potential driver of omnivory levels. Mar. Biol. 167, 1–17 (2020).
Stubbs, J. L., Marn, N., Vanderklift, M. A., Fossette, S. & Mitchell, N. J. Simulated growth and reproduction of green turtles (Chelonia mydas) under climate change and marine heatwave scenarios. Ecol. Modell. 431, 109185 (2020).
Türkozan, O. & Kaska, Y. Turkey. In Sea Turtles in the Mediterranean: Distribution, Threats, and Conservation Priorities (eds Casale, P. & Margaritoulis, D.) 257–293 (IUCN/SSC Marine Turtle Specialist Group, Gland, 2010).
Broderick, A. C., Glen, F., Godley, B. J. & Hays, G. C. Estimating the number of green and loggerhead turtles nesting annually in the Mediterranean. Oryx 36(3), 227–235 (2002).
Fratianni, C., Simoncelli, S., Pinardi, N., Cherchi, A., Grandi, A., Dobricic, S., 2014. Mediterranean sea RR 1955–2015. Copernic. Monit. Environ. Mar. Serv.
Teruzzi, A., Bolzon, G., Cossarini, G., Lazzari, P., Salon, S., Crise, A., Solidoro, C. Mediterranean Sea biogeochemical reanalysis (CMEMS MED-Biogeochemistry). Copernic. Monit. Environ. Mar. Serv. (2019)
Sbrocco, E. J. & Barber, P. H. MARSPEC: Ocean climate layers for marine spatial ecology: ecological archives E094–086. Ecology 94(4), 979–979 (2013).
European Commission, Joint Research Centre (JRC). Global Shoreline Change Projections. European Commission, Joint Research Centre (JRC) (2019)
Gao, J. (2020). Global 1-km Downscaled Population Base Year and Projection Grids Based on the Shared Socioeconomic Pathways, Revision 01. NASA Socioeconomic Data and Applications Center (SEDAC).
Falchi, F. Light Pollution 147–159 (Urban Pollution Science and Management, 2018).
Spotila, J. R. & Standora, E. A. Environmental Constraints on the Thermal Energetics of Sea Turtles 694–702 (Copeia, 1985).
Seebacher, F. & Franklin, C. E. Physiological mechanisms of thermoregulation in reptiles: A review. J. Comp. Physiol. B 175, 533–541 (2005).
Hawkes, L. A., Broderick, A. C., Godfrey, M. H. & Godley, B. J. Investigating the potential impacts of climate change on a marine turtle population. Glob. Change Biol. 13(5), 923–932 (2007).
Mazaris, A. D., Kornaraki, E., Matsinos, Y. G. & Margaritoulis, D. Modeling the effect of sea surface temperature on sea turtle nesting activities by investigating seasonal trends. Nat. Resour. Model. 17(4), 445–465 (2004).
Lutz, P. L., Musick, J. A. & Wyneken, J. The biology of sea turtles Vol. II (CRC Press, 2002).
Bolten, A. B. & Witherington, B. E. Loggerhead sea turtles. Mar. Turt. Newsl. 104, 319 (2004).
Ngatia, L., Grace, J. M. III., Moriasi, D. & Taylor, R. Nitrogen and phosphorus eutrophication in marine ecosystems. Monit. Mar. Pollut. 1, 1–17 (2019).
Lutcavage, M. E. (2017). Human impacts on sea turtle survival. In The Biology of Sea Turtles. CRC press
Bozorg-Haddad, O. (ed.) Economical, Political, and Social Issues in Water Resources (Elsevier, 2021).
Brodie, J., Ariel, E., Thomas, C., O’Brien, D., & Berry, K. Links Between Water Quality and Marine Turtle Health. (TropWATER—Tropical Water & Aquatic Ecosystem Research, 2014).
Casale, P. Sea Turtles in the Mediterranean: Distribution, Threats and Conservation Priorities (IUCN, 2010).
Witherington, B. E. & Martin, R. E. Understanding, Assessing, and Resolving Light-Pollution Problems on Sea Turtle Nesting Beaches (Florida Marine Research Institute, 2000).
Vandersteen, J., Kark, S., Sorrell, K. & Levin, N. Quantifying the impact of light pollution on sea turtle nesting using ground-based imagery. Rem. Sens. 12(11), 1785 (2020).
Araújo, M. B. et al. Standards for distribution models in biodiversity assessments. Sci. Adv. 5(1), eaat4858 (2019).
Tyberghein, L. et al. Bio-ORACLE: A global environmental dataset for marine species distribution modelling. Glob. Ecol. Biogeogr. 21, 272–281 (2012).
Assis, J. et al. Bio-ORACLE v2.0: Extending marine data layers for bioclimatic modelling. Glob. Ecol. Biogeogr. 27(3), 277–284 (2017).
Chambault, P. et al. The influence of oceanographic features on the foraging behavior of the olive ridley sea turtle Lepidochelys olivacea along the Guiana coast. Progr. Oceanogr. 142, 58–71 (2016).
Sloan, K. A., Addison, D. S., Glinsky, A. T., Benscoter, A. M. & Hart, K. M. Inter-nesting movements, migratory pathways, and resident foraging areas of green sea turtles (Chelonia mydas) satellite-tagged in Southwest Florida. Front. Mar. Sci. 12, 1798 (2022).
Hart, K. M., Zawada, D. G., Fujisaki, I. & Lidz, B. H. Inter-nesting habitat-use patterns of loggerhead sea turtles: Enhancing satellite tracking with benthic mapping. Aquat. Biol. 11(1), 77–90 (2010).
Burnham, K. P. & Anderson, D. R. Practical use of the Information-Theoretic Approach 75–117 (Springer, 1998).
Phillips, S. J., Anderson, R. P., Dudík, M., Schapire, R. E. & Blair, M. E. Opening the black box: An open-source release of Maxent. Ecography 40(7), 887–893 (2017).
Spalding, M. D. et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 57(7), 573–583 (2007).
Muscarella, R. et al. ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 5(11), 1198–1205 (2014).
Li, Y., Li, M., Li, C. & Liu, Z. Optimized maxent model predictions of climate change impacts on the suitable distribution of cunninghamia lanceolata in China. Forests 11(3), 302 (2020).
Warren, D. L. & Seifert, S. N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 21(2), 335–342 (2011).
Raes, N. & ter Steege, H. A null-model for significance testing of presence-only species distribution models. Ecography 30(5), 727–736 (2007).
Phillips, S. J. et al. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 19(1), 181–197 (2009).
Antão, L. H. et al. Temperature-related biodiversity change across temperate marine and terrestrial systems. Nat. Ecol. Evol. 4(7), 927–933 (2020).
Pinsky, M. L., Comte, L., & Sax, D. F. (2022). Unifying Climate Change Biology Across Realms and Taxa. Trends in Ecology & Evolution.
Kosanic, A. & Petzold, J. A systematic review of cultural ecosystem services and human wellbeing. Ecosyst. Serv. 45, 101168 (2020).
Bent, E. Induced systemic resistance mediated by plant growth-promoting rhizobacteria (PGPR) and fungi (PGPF). Multigenic Induc. Syst. Resist. Plants 62, 225–258 (2006).
Lugtenberg, B. & Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63, 541–556 (2009).
Wallace, B. P. et al. Regional management units for marine turtles: A novel framework for prioritizing conservation and research across multiple scales. Plos one 5(12), e15465 (2010).
Camiñas, J. A. Sea Turtles of the Mediterranean Sea: Population Dynamics Sources of Mortality and Relative Importance of Fisheries Impacts (The Center for Oceanography of Malaga, 2004).
Kasparek, M., Godley, B. J. & Broderick, A. C. Nesting of the green turtle, Chelonia mydas, in the Mediterranean: A review of status and conservation needs. Zool. Middle East 24(1), 45–74 (2001).
Hochscheid, S., Kaska, Y. & Panagopoulou, A. Sea Turtles in the Mediterranean Region: MTSG Annual Regional Report 2018 (Draft Report of the IUCN-SSC Marine Turtle Specialist Group, 2018).
Casale, P., Hochscheid, S., Kaska, Y. & Panagopoulou, A. Sea Turtles in the Mediterranean Region (MTSG Annual Regional Report, 2020).
Jančič, M. et al. Apparent increasing importance of Adriatic Sea as a developmental habitat for Mediterranean green sea turtles (Chelonia mydas). Nat. Croat. Period. Musei Hist. Nat. Croat. 31(2), 225–240 (2022).
Mazaris, A. D., Kornaraki, E., Matsinos, Y. G. & Margaritoulis, D. Modeling the effect of sea surface temperature on sea turtle nesting activities by investigating seasonal trends. Natl. Resour. Model. 17(4), 445–465 (2004).
Rixen, M. et al. The western mediterranean deep water: A proxy for climate change. Geophys. Res. Lett. 32(12), 5985436 (2005).
Belkin, I. M. Rapid warming of large marine ecosystems. Progr. Oceanogr. 81(1–4), 207–213 (2009).
Shaltout, M. & Omstedt, A. Recent sea surface temperature trends and future scenarios for the Mediterranean Sea. Oceanologia 56(3), 411–443 (2014).
Skliris, N. et al. Decadal scale variability of sea surface temperature in the Mediterranean Sea in relation to atmospheric variability. Ocean Dyn. 62, 13–30 (2012).
Kramer-Schadt, S. et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 19(11), 1366–1379 (2013).
Heikkinen, R. K. et al. Methods and uncertainties in bioclimatic envelope modelling under climate change. Progr. Phys. Geogr. 30(6), 751–777 (2006).
Fielding, A. H. (2002). What are the appropriate characteristics of an accuracy measure? In ‘Predicting Species Occurrences: Issues of Accuracy and Scale’ (Eds JM Scott, PJ Heglund, ML Morrison, JB Haufler, MG Raphael, WA Wall and FB Samson.) pp. 271–281.
Aurelle, D. et al. Biodiversity, climate change, and adaptation in the Mediterranean. Ecosphere 13(4), e3915 (2022).
Hamann, M., Fuentes, M. M., Ban, N. C. & Mocellin, V. J. 13 climate change and marine turtles. Biol. Sea Turt. 3, 353 (2013).
Nasr, A. et al. A review of the potential impacts of climate change on the safety and performance of bridges. Sustain. Resilient Infrastruct. 6(3–4), 192–212 (2021).
Johnston, E. L., Clark, G. F. & Bruno, J. F. The speeding up of marine ecosystems. Clim. Change Ecol. 3, 100055 (2022).
Perez, E. A., Marco, A., Martins, S. & Hawkes, L. A. Is this what a climate change-resilient population of marine turtles looks like?. Biol. Conserv. 193, 124–132 (2016).
Mainwaring, M. C. et al. Climate change and nesting behaviour in vertebrates: A review of the ecological threats and potential for adaptive responses. Biol. Rev. 92(4), 1991–2002 (2017).
Davenport, J. Sustaining endothermy on a diet of cold jelly: energetics of the leatherback turtle Dermochelys coriacea. Bull. Brit. Herpetol. Soc. 62, 4–8 (1998).
Weishampel, J. F., Bagley, D. A. & Ehrhart, L. M. Earlier nesting by loggerhead sea turtles following sea surface warming. Glob. Change Biol. 10(8), 1424–1427 (2004).
Mitchell, N. J. & Janzen, F. J. Temperature-dependent sex determination and contemporary climate change. Sexual Dev. 4(1–2), 129–140 (2010).
Hawkes, L. A., Broderick, A. C., Godfrey, M. H., Godley, B. J. & Witt, M. J. The Impacts of Climate Change on Marine Turtle Reproductive Success 287–310 (Coastal conservation. Cambridge University Press, 2014).
Almpanidou, V., Markantonatou, V. & Mazaris, A. D. Thermal heterogeneity along the migration corridors of sea turtles: Implications for climate change ecology. J. Exp. Mar. Biol. Ecol. 520, 151223 (2019).
Carreras, C. et al. Sporadic nesting reveals long distance colonisation in the philopatric loggerhead sea turtle (Caretta caretta). Sci. Rep. 8(1), 1435 (2018).
Schofield, G. et al. Microhabitat selection by sea turtles in a dynamic thermal marine environment. J. Animal Ecol. 23, 14–21 (2009).
Lacroix, D. Adapting to global change in the Mediterranean Sea. Mediter. Region Clim. Change 647, 466995 (2016).
Brock, K. A., Reece, J. S. & Ehrhart, L. M. The effects of artificial beach nourishment on marine turtles: Differences between loggerhead and green turtles. Restor. Ecol. 17(2), 297–307 (2009).
Kamrowski, R. L., Limpus, C., Moloney, J. & Hamann, M. Coastal light pollution and marine turtles: assessing the magnitude of the problem. Endanger. Species Res. 19(1), 85–98 (2012).
Fuentes, M. M. et al. Conservation hotspots for marine turtle nesting in the United States based on coastal development. Ecol. Appl. 26(8), 2708–2719 (2016).
Dimitriadis, C., Fournari-Konstantinidou, I., Sourbès, L., Koutsoubas, D. & Mazaris, A. D. Reduction of sea turtle population recruitment caused by nightlight: Evidence from the Mediterranean region. Ocean Coast. Manag. 153, 108–115 (2018).
Fuentes, M. M., Pike, D. A., Dimatteo, A. & Wallace, B. P. Resilience of marine turtle regional management units to climate change. Glob. Change Biol. 19(5), 1399–1406 (2013).
Pikesley, S. K. et al. Modelling the niche for a marine vertebrate: A case study incorporating behavioural plasticity, proximate threats and climate change. Ecography 38(8), 803–812 (2015).
Acknowledgements
The authors acknowledge the work and effort of all studies used in this study. We would like to thank all researchers, organizations and associations that monitor beaches collecting and publishing nesting data along the coasts of the Mediterranean Sea.
Author information
Authors and Affiliations
Contributions
This work was carried out in collaboration among all authors. All Authors designed the study, C.M. performed the statistical analysis and wrote the original first draft. L.M. and S.H. supported and contributed to the analyses’ methodology. All authors contributed to the writing, read, and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mancino, C., Hochscheid, S. & Maiorano, L. Increase of nesting habitat suitability for green turtles in a warming Mediterranean Sea. Sci Rep 13, 19906 (2023). https://doi.org/10.1038/s41598-023-46958-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-46958-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
