
Abstract
Globally, 15–24% couples are unable to conceive naturally and 50% of cases of this problem are due to infertility in males. Of this, about 50% of male infertility problems are developed due to unknown reasons called as idiopathic infertility. It is well established that, reactive oxygen species (ROS) have negative impact on male fertility and are involved in 80% of total idiopathic male infertility cases. Medicinal plants are considered as an alternative approach for mitigating the health problems. The plants with good antioxidant capacity can improve the male infertility symptoms generated by ROS. Such medicinal plants can be used to alleviate the symptoms of male infertility with their diverse phytoconstituents. Mucuna pruriens is a well-accepted herb, with its seeds being used to improve the male fertility in various ways and one of the ways is by eliminating the ROS. In our field survey, another plant, Flemingia praecox, although less known, its roots are used in all problems related to the male fertility by tribal people of the Gadchiroli district of Maharashtra, India. The study was conducted to determine in vitro antioxidant potential of F. praecox and compared the results with the well-established male fertility improving plant M. pruriens with special emphasis on medicinally important roots of F. praecox and seeds of M. pruriens. The objective of the study was investigated by studying their total phenol (TPC) and flavonoid (TFC) content, antioxidant parameters (DPPH, FRAP, ABTS, DMPD, β-carotene bleaching and TAA) and finally DNA damage protection capacity of the plant extracts was studied. The plant parts used for the medicinal purposes have been investigated along with other major parts (leaves, stem and roots of both the plants) and compared with synthetic antioxidants, BHA, BHT and ascorbic acid. Moreover, the inhibition of two male infertility enzyme markers, PDE5 and arginase by F. praecox root and M. pruriens seed extract was also studied in vitro. The results showed that F. praecox possesses higher antioxidant activity than M. pruriens in the majority of studies as observed in TFC, DPPH, TAA, ABTS and DMPD assays. However, M. pruriens seeds showed best results in TPC, FRAP and DNA damage protection assay. F. praecox root extract also gave better PDE5 inhibition value than M. pruriens seeds. This study will help to establish the authenticity of F. praecox used by tribal people and will encourage its further use in managing the male infertility problems.
Similar content being viewed by others
Introduction
It is evident from previous large scale surveys that sperm count had declined by 50–60% globally during the last 60-year1,2,3. Male infertility is associated with greater incidence of cancer4, obesity, diabetes5, metabolic syndrome6 and also with mortality and can even cause problems in the health of future progeny5. Therefore, in a greater perspective, male infertility should not be seen only as a medical condition affecting fertility, but also general health and wellbeing7. The problem of male infertility is heterogeneous in origin which may be the consequence of genetic or environmental factors or both. The genetic factor includes, microdeletions in Y-chromosome, autosomal deletions, X-linked gene copy number variations, mutation in Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) gene, defects in DNA repair mechanisms, etc.8. Other factors contributing to this problem includes environmental or occupational exposure to toxicants, lifestyle like smoking, alcohol consumption, drugs, psychological stress9 and recreational drugs which acts at the level of hypothalamic–pituitary–gonadal axis or directly on spermatogenesis consequently causing infertility10. However, in most cases the causes of male subfertility are poorly understood or not known, called idiopathic male infertility11. Recently, it has been reported that the epigenetic modifications, like abnormal DNA methylation, small non-coding RNA, histone tail modification8, single nucleotide polymorphism12, etc. in reproduction-related genes are responsible factors for idiopathic male infertility. However, many of these genetic problems are linked with antioxidant or ROS genes12 which might influences the critical balance between antioxidant and ROS.
ROS are one of the most closely associated factors involved in deciding the male infertility. ROS or the free radicals (FR) are the molecules with at least one unpaired electron. It is generated as a result of oxygen metabolism. The unpaired electrons make ROS a highly reactive and damaging chemical13. Low levels of ROS are required in various events of fertilization13 however, its excessive production, because of any reason and if it is not counterbalanced by body’s own antioxidant defences like superoxide dismutase, catalase, glutathione peroxidase and glutathione, etc.14 then it will lead to oxidative stress (OS). Consequently, it will cause oxidative damage to spermatozoa by increasing lipid peroxidation in its plasma membrane and thus alter the sperm functioning15. About 80% of the idiopathic infertile male16,17 and 30–40% of males with known causes has been reported to have elevated levels of ROS called as male oxidative stress infertility (MOSI)18. This oxidative stress will results in the protein, lipid and DNA damage in and around sperm atmosphere resulting in decline of fertility. The good thing about OS is that it can be reversed by using oral antioxidants and thus provides a good opportunity for treatment9.
The plant based antioxidants can become a good alternative to mitigate the problem of MOSI. The major antioxidant compounds in the plants are phenolics and flavonoids which work by eliminating and preventing the production of ROS19. Phenolic compounds have potential to scavenge major ROS and FR by different ways (Fig. 1). Moreover, the chemical structure of phenolics is more crucial than their concentration as it determines the extent of their absorption in the plasma20. Flavonoids, in the same line, although having the better antioxidant potential than phenols21 but due to their low absorption through intestinal route it was thought to work in improving the male fertility in different ways. It is evident that flavonoids improve male fertility preferably by modulating the cell signalling pathways and improving22,23,24,25,26 (Fig. 2). Hence, the plants with high concentration of different phenolic compounds including flavonoids can be used as a good source of antioxidants to alleviate the problem of male infertility.

Phenol (blue) with its different mechanism of action against ROS (red).

Flavonoids contribute in improvement of male fertility by different mechanisms.
M. pruriens is a well-recognized plant and traditionally been used for improving the male fertility. It improves male fertility by reducing ROS level, restoring mitochondrial membrane potential, regulating apoptosis27, controlling unspecific generation of ROS28, reactivating the antioxidant defense system, managing stress29, reducing lipid peroxidation30, acting on hypothalamus–pituitary–gonadal (HPG) axis and increasing levels of hormones28. Moreover, it is also thought that the male specific hormone production is assisted by presence of an active component, levodopa in its seeds28. However, a study showed that, apart from levodopa, other more superior bioactive components must be present in its seeds31.
Genus Flemingia consists of 44 species and two varieties that are mostly distributed in old world tropics32. In India, it is represented by 27 species and one variety33. The genus Flemingia is not only known for its high concentration of flavonoids but also contains its good diversity including flavones, flavanones, isoflavones, isoflavanone, chalcones, dihydrochalcones, flavonols, santalin flavones, flavanol, chromone and diflavone34. Traditionally, genus Flemingia have been used in the treatment of diseases like epilepsy, insomnia, ulcer, pain, swelling and regardless of a long traditional use of some species, this genus has not been explored properly35. However, the selected taxon for the study, F. praecox var. robusta (F. praecox hereafter) is endemic to India and has been reported in various parts of India36,37 and its phytochemical study was not done before. Interestingly, the traditional medicinal practitioners in Gadchiroli district of Maharashtra, India use this plant against male infertility problems. Therefore, we hypothesizing that F. praecox must be having chemical properties specific for improving the male fertility.
To check this hypothesis, we have conducted in vitro antioxidant studies on both F. praecox (its leaves, stem and medicinally important roots) and the well-recognized and traditionally used plant M. pruriens (its roots, leaves, stem and medicinally important seeds) under the similar analytical conditions and compared the findings. The correlational studies were also performed to discuss probable action mechanisms of these plants on ROS. Finally, inhibition of two male infertility markers, Phosphodiesterase 5 (PDE5) and arginase by the plant extract were also studied in vitro.
Based on recent literature reviews, it was observed that among various Flemingia species recognized for their medicinal properties, the most important organ with medicinal use was its roots38,39,40,41,42,43,44,45,46,47 followed by leaves and stems35,48. A very few studies have demonstrated the use of its seeds for medicinal purposes35. Moreover, none of the work has shown its any organ with capacity to ameliorate the male reproductive health. Furthermore, the traditional medicinal practitioners of Gadchiroli district were also denied the use of its seeds in male infertility cases. Due to these reasons, and most importantly, the extremely limited availability, we have not included the seeds of F. praecox in our studies.
Results and discussion
Preliminary phytochemical availability test
We have tested the availability of phenols, flavonoids, glycosides, alkaloids, terpenoids, tannins, steroids and saponins (Table 1). These tests were performed because we did not find any previous studies in literature on phytochemistry of F. praecox. The results obtained were compared with the M. pruriens. Multiple tests were performed for various categories of secondary metabolites. All parts of F. praecox have shown positive results in all the tests performed for phenols, flavonoids and also to some extent tannins. However, terpenoids and glycosides are almost absent in the F. praecox extracts which is also the case with the M. pruriens extract. M. pruriens also showed presence of phenols and flavonoids except its seed and root which showed negative results in some of the tests. Alkaloids were present in the leaves of both the plants. The roots of F. praecox and leaves of M. pruriens also give positive results for the presence of steroids. M. pruriens seeds also possessed good foaming capacity indicating the presence of saponins.
Alkaloids are produced by the plants mainly for deterring herbivory by vertebrates which are also observed to have a negative role in pharmacological context49. Except phenolic and flavonoid glycosides all other glycosides are known to have adverse effects on50,51. Hence, the absence or low levels of glycosides and alkaloids in medicinally important plant parts i.e. seeds of M. pruriens and roots of F. praecox eliminates its possible side effects on the health. Previous data indicated the absence of alkaloids in aqueous and methanolic extract of M. pruriens leaf however, in our analysis all the tests performed showed its presence. Moreover, the terpenoids were absent in our analysis but they were found by other workers52. Previously alkaloids were isolated from M. pruriens leaves53 and seeds54 and its bioactivity was also studied which validates our positive results for alkaloids in M. pruriens55. Our results of saponins, tannins and flavonoids in M. pruriens were in accordance with results obtained by previous workers52. Steroids were reported in M. pruriens seeds56 however, in our analysis it was not detected in seeds but observed in the leaf.
Work has been done on various species of Flemingia like F.57,58,59, F. macrophyla60, F. chappar61, F. philippinensis62, F. faginea48, F. grahmiana63 F. stricta64, etc. but no phytochemical work has been found in the literature on the species F. praecox. However, these species have shown the presence of phenols, flavonoids, steroids, tannins, glycosides, alkaloids and saponins and in most of the species the terpenoids were not64,65 our preliminary analysis also, the terpenoids were not detected in F. praecox.
Quantification of phenolics
In the plant parts
Phenols and flavonoids are major groups of secondary metabolites that are known to have maximum shares in total antioxidant potential of any plant19. TPC and TFC were calculated first in all the plant parts (Fig. 3a and b) and then the plant parts that gave the highest values were fractionated by using various solvents with increasing polarity and again TPCs and TFCs of these fractions were estimated (Fig. 4a and b).

Total phenolics in methanolic extract of organs of M. pruriens (M.) and F. praecox (F.) (a) and total flavonoids in methanolic extract of parts of M. pruriens and F. praecox (b). Values are presented as means of three readings ± SD (standard deviation). Highest to lowest values are shown in alphabetical order. Means with the different letter are significantly different at 95% confidence interval (p < 0.05) according to Tukey’s multiple range test.

Total phenolics (a) and flavonoids (b) in different fractions made by sequential extraction of M. pruriens seeds (M.) and F. praecox roots (F.) in different solvents. Different letters represent significant differences at the p < 0.05 level.
In our analysis, the significantly highest phenolic content was observed in M. pruriens seeds which is 327.48 ± 3.81 mg of gallic acid equivalent per gram of plant extract (mg GAE/g) followed by F. praecox roots containing 199.00 ± 5.96 mg GAE/g. However, leaves of both the plants showed statistically similar concentrations of phenol that is 154.78 ± 1.98 and 160.23 ± 5.85 mg GAE/g in F. praecox and M. pruriens respectively. The roots of M. pruriens and stem of F. praecox contain its minimum concentration (91.38 ± 0.88 and 90.71 ± 0.77 mg GAE/g respectively).
Unlike phenols, the highest level of flavonoids was estimated in F. praecox root that is 360.93 ± 8.49 mg of quercetin equivalent per gram of extract (mg QE/g) followed by M. pruriens seeds containing 277.59 ± 16.14 mg QE/g. F. praecox leaves also have shown the better levels of flavonoid (216.48 ± 8.49 mg QE/g) than other remaining plant parts. F. praecox stems contain lowest flavonoid content among the rest of the plant parts of both the plants (36.85 ± 3.21 mg QE/g).
In the fractions
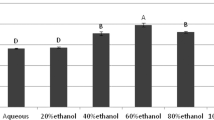
On the basis of results obtained, we have selected M. pruriens seeds and F. praecox roots which are also the parts that are being used for improving the male fertility and attempted to quantify the phenols and flavonoids from their serial fractions made in different solvents with increasing polarity (Fig. 4a and b). The serial fractionation was done in the following sequence, n-hexane → ethyl acetate → chloroform → acetone → methanol. In our study, acetone extracted the highest fraction of phenol from F. praecox roots (175.87 ± 9.536 mg GAE/g). Methanol also dissolved significantly higher phenols from F. praecox roots (131.65 ± 34.90 mg GAE/g) than the remaining solvents. Also ethyl acetate fraction of F. praecox root has shown good phenolic value (67.77 ± 2.830 mg GAE/g). Among the M. pruriens seed fractions, the highest phenolic value was obtained in its methanolic fraction (115.99 ± 0.270 mg GAE/g) which is also statistically similar to phenols obtained in the methanolic fraction of F. praecox root.
The TFC analysis of different solvent fraction showed that F. praecox root contain significantly higher flavonoids that are soluble in acetone (522.96 ± 6.55 mg QE/g) followed by methanolic fraction (281.914 ± 1.07 mg QE/g). The ethyl acetate also extracted considerable flavonoids from F. praecox roots (194.26 ± 6.68 mg QE/g). Among M. pruriens, its methanolic fraction contains a maximum flavonoid than other fractions (265.86 ± 12.05 mg QE/g) indicating methanol as the best solvent for flavonoid extraction from M. pruriens.
Previously, the phenol content of M. pruriens was quantified by using different extraction solvents and methods. Due to its medicinal property, phenols were quantified mostly from seed extracts made in water, ethanol and methanol and it was found in the range of 3.9 to 230 mg GAE/g66,67,68,69,70, clearly showing that solvent influences the phenolic extraction. These levels were much lower than our quantified results in crude methanolic seed extract. Some studies however showed a considerably high level of phenolics up to 3730 mg GAE/g of extract71.
As stated earlier, the species F. praecox was not studied in the context of its phytochemistry. This may be due to its very low population size or its rarity in nature. However, we studied its phytochemistry for the first time from its restored population in our experimental field. In literature, we found that most of the work was done on F. philippinensis72. In its leaves, the phenols were 40 mg GAE/g and the roots showed 49 mg GAE/g. Other species of Flemingia expectedly showed varied amounts of phenols which ranges from 12 mg GAE/g in F. strobilifera and F. vestita to 280 mg GAE/g in F. faginea47,48,57,73,74 In our studies on F. praecox the phenols were found in good concentration i.e. 199 mg GAE/g in roots and also its leaves contain considerably higher phenols than in the leaves of F. philippinensis72. Thus our observation indicates that F. praecox can be the better source of phenolic antioxidants among its other species.
The methanolic extract of M. pruriens seeds has significantly higher concentration of phenols than its other organs which may be the reason for using its seeds for improving the male fertility. On the other hand, methanolic extract of F. praecox roots also showed considerably higher levels of phenols than its other organs which here as well can be the reason for its use in alleviating the male infertility problems by conventional medicinal practitioners. Moreover, acetone and methanol fractions of F. praecox give significantly higher phenol estimates even better than the similar fractions of M. pruriens seeds. Thus it can be concluded that F. praecox and M. pruriens are both relatable in the context of its phenolic content and medicinal property.
Previous studies have reported the high levels of flavonoids and also new flavonoids have been discovered from time to time from different species of Flemingia. Moreover, in vitro and preclinical properties of these flavonoids have also been reported by various researchers42,75,76,77,78,79. Some researchers also worked on isolation and in vitro properties of new flavonoids from leaves of other F. praecox63,80,81.
In M. pruriens, previous studies indicated its flavonoids are in the range of 63 to 807 mg QE/g of aqueous extract70,71 and 423 mg QE/g of ethanolic extract82 again indicating the place of origin of the plant and extraction procedure affecting the quantified values. In our analysis we observed maximum flavonoids in the M. pruriens seeds than its other organs studied which again signifying the use of its seeds for the medicinal purpose.
Studies on the flavonoids in different species of Flemingia showed that its value was ranging from 0.75 to 52.76 mg rutin equivalent per g aqueous47,73 and from 7.69 to 30.58 mg QE/g of hydro-alcoholic74,83. In one more study on the F. faginea leafy shoot found to contain 33.31 mg QE/g flavonoids in its aqueous extract. However, our study on F. praecox showed significantly high concentration of flavonoids in both its roots and leaf. Moreover, the analysis of flavonoid content in different solvent fractions of its roots shown even higher flavonoids specially in acetone fraction followed by methanolic fraction, indicating the acetone as a better solvent for flavonoid isolation from F. praecox roots which is also specified earlier by Pisoschi et al.84 Comparison of flavonoids in M. pruriens and F. praecox indicates that F. praecox species under study is far better source of flavonoids than its counterpart M. pruriens. F. praecox contain at least about more than two folds of flavonoid concentration in its acetone fraction than any fractions of M. pruriens and about 30% more in its medicinally important organ root than M. pruriens seeds. Even the leaves of F. praecox contain considerably good concentration of flavonoids.
Flavonoids are important group of secondary metabolite consist of cyclized diphenylpropane structure19 and are secreted in plants in the form of pigments in flowers, fruits, seeds, and leaves for recruiting pollinators and seed dispersers, in defence as feeding deterrent and antimicrobial agents, and in UV protection85. However, due to its diverse structure it is also found to have important medicinal applications. It was observed that in contrast to simple phenols which have mainly antioxidant properties (Fig. 1)22,86. Flavonoids show antioxidant activity mostly by chelating free radical forming metal ions like Fe2+ by formation of coordinate bonds with them by its –C=O and –OH groups87. Along with its major property of modulation of cell signalling pathway it was reported that flavonoids have capacity to improve vascular endothelial function by increasing the production of nitric oxide (NO) through endothelial nitric oxide synthase (eNOS)88. Flavonoids also have a neuroprotective role as it stimulates neuronal nitric oxide synthase (nNOS)89,90 and also possesses antidiabetic properties due to its insulin production capacity therefore improving the diabetes mediated vascular dysfunction91. Moreover, another group of flavonoids, isoflavones, have prostate cancer inhibition capacity by hormone dependent signalling pathway92 along with various spermatogenesis promoting effects93 (Fig. 2). This male fertility improving role of flavonoids can be met by its high levels in the F. praecox.
Phenol and flavonoid detection in plant fractions by HPLC–MS/MS analysis
In F. praecox root, higher number of phenolic compounds were detected compared to the seeds of M. pruriens (Tables 2 and 3). Many of these compounds have been reported to have a positive impact on male fertility through various mechanisms. Some of these compounds possess antioxidant properties and may contribute to the reduction of oxidative stress21 or they are acting at different levels of male reproductive system.
For instance, the epigallocatechin and other catechins from F. praecox have been shown to reverse testicular damage94 possibly due to their active antioxidant95 or DNA damage protection properties96. Similar positive effects on male fertility have been reported for compounds such as Lespenefril24, Chrysin97 and rutin98, all found in F. praecox. Spermidine derivatives like N1, N5, N10-Tricoumaroyl spermidine have been associated with ameliorative effects on sperm disorders in diabetic mice99. Other compounds in F. praecox, including Ononin100, Procyanidin B7, Curcumin II101 and Mulberranol102 have spermatogenic effect by modulating testosterone and other sex hormone levels. Moreover, flavonoids like Isoliquiritigenin have been reported to ameliorate sexual dysfunction103, while Licocoumarin A has been identified as an estrogen modulator104. Lastly, Xanthohumol has demonstrated the capacity to inhibit the growth and invasion of prostate cancer cells105.
On the other hand, in M. pruriens seed extract, the reported phenolic compound 5-(3′,4′,5′-Trihydroxyphenyl)-gamma-valerolactone which has been reported to possess neuroprotective properties106 and thus might have implication in the psychogenic male infertility. Another compound, the isoflavonoid, Luteolin have a well-known positive role in the process related to steroidogenesis, apoptosis and in stress response107. However, Beclomethasone dipropionate another compound detected in the M. pruriens seeds has been associated with negative effects on the reproductive function of male rats108. These findings collectively suggest that the presence of these phenolic and flavonoid compounds in both F. praecox roots and M. pruriens seeds may contribute to the improvement of male fertility, although through different mechanisms and at various levels within the male reproductive system.
In vitro antioxidant capacity
The high concentrations of phenols and flavonoids in the medicinally used M. pruriens seeds and F. praecox roots also indicated the possibility of having high antioxidant values. Antioxidants are a major primary defence system against ROS and FR. It is well established from previous research that ROS and FR are the important contributory factors in various diseases including male infertility17,109,110,111. To check this hypothesis we studied the antioxidant properties of the plant parts of M. pruriens and F. praecox by DPPH (2,2-diphenyl-1-picrylhydrazyl), ABTS (2,2′-azino-bis-(3-ethylbenzothiazoline-6-sulfonic) acid) and DMPD (N, N-dimethyl-p-phenylenediamine) free radical scavenging assay, β-carotene bleaching, FRAP (Ferric ion reducing antioxidant power) and phosphomolybdenum antioxidant assay. The results obtained were compared with artificial antioxidants, butylated hydroxyanisole (BHA), butylated hydroxytoluene (BHT) and ascorbic acid.
DPPH⋅ scavenging activity
The DPPH radical scavenging activity is based on the reduction of purple coloured DPPH⋅ to its yellow hydrazine product (DPPH-H) by hydrogen or electron donating capacity of the plant compounds19,112. We have studied DPPH⋅ scavenging activity of plant parts (Fig. 5a) and the different fractions of M. pruriens seeds and F. praecox roots (Fig. 5b). The study revealed that F. praecox root has best scavenging activity among all the plant parts with IC50 (half-maximal inhibitory concentration) value 7.34 ± 0.315 µg and is also statistically similar in its scavenging activity to artificial antioxidants, ascorbic acid (5.24 ± 0.29 µg) and BHA (4.3 ± 0.12 µg) and even better than BHT (13.7 ± 0.31 µg). Other workers studied DPPH⋅ scavenging activity in different species of Flemingia, mostly in its roots47,48,57,74,83 and leaves63. In leaves of F. grahmiana, Gumula et al. showed IC50 value 5.9 µg whereas others studies on roots of various species of Flemingia, the IC50 for DPPH⋅ scavenging was ranged from best in F. faginea (15.04 µg)48 to the least in F. vestita (287 µg)74. However, among M. pruriens, its seed possesses the highest scavenging activity with IC50 value 18.34 ± 0.182 µg which is statistically similar to BHT. In previous studies on alcoholic and hydro-alcoholic extract of M. pruriens seeds, the DPPH⋅ scavenging activity in terms of IC50, was in range of 5.1 to 61.02µg67,68,69,70. Among the fractions of M. pruriens seed and F. praecox root, best DPPH⋅ scavenging activity was observed in methanol and acetone fractions of F. praecox root with IC50 values 7.21 ± 0.26 µg and 8.15 ± 0.83 µg respectively followed by methanol fraction of M. pruriens seeds having IC50 value 11.79 ± 0.51 µg. These all values are statistically similar to standards used at p < 0.05. Hexane and chloroform fraction of M. pruriens seeds did not show any scavenging activity at the used concentration.

DPPH radical scavenging capacity of plant parts of M. pruriens and F. praecox (a) and the sequentially extracted fractions (b) of M. pruriens seeds and F. praecox roots. Different letters represent significant differences at the p < 0.05 level. ND not detected.
ABTS⋅+ scavenging activity
ABTS⋅+ scavenging activity demonstrates the capacity of the phytochemicals to neutralize ROS by hydrogen atom transfer (HAT) or single electron transfer (SET) mechanism19. The best HAT or SET capacity again shown by F. praecox roots having ABTS•+ scavenging IC50 value 3.63 ± 0.112 µg (Fig. 6a). Statistical tests shows that ABTS•+ scavenging potential of F. praecox root is statistically similar (p < 0.05) to ascorbic acid (2.50 ± 0.125 µg) and BHT (3.10 ± 0.832 µg). The artificial antioxidant, BHA have shown the best ABTS•+ scavenging activity with IC50 value 2.14 ± 0.066 µg. Previous work on other species of Flemingia like F. faginia48 and F. vestita74 found the ABTS⋅+ scavenging IC50 value 67.33 µg and 11.49 µg respectively. These activities shown by other species are much lower than our studied species F. praecox. In the case of M. pruriens, seeds (8.64 ± 0.149 µg) and stem (9.43 ± 0.196 µg) have shown better scavenging activity than its other organs but significantly lower than its counterpart F. praecox root. At the end, M. pruriens root (14.60 ± 0.273 µg) and F. praecox stem (14.00 ± 0.482 µg) have shown lowest capacity to scavenge ABTS radicals. However, a large range of ABTS⋅+ scavenging values were observed by other workers in M. pruriens which is 6.009 µg as found by Njemuwa et al.69 to 137 µg which was observed by Chittasupho et al.70.

ABTS (a), DMPD (b) radical scavenging and β-carotene bleaching inhibition activity (c) of plant parts of M. pruriens and F. praecox. Different letters represent significant differences at the p < 0.05 level. ND not detected.
DMPD⋅+ scavenging
Another radical scavenging activity investigated with DMPD radical cation (DMPD•+) which is based on the HAT and SET mechanism of radical scavenging. This assay is less sensitive to hydrophobic and more specific for the hydrophilic antioxidants. This is opposite to DPPH⋅ and ABTS⋅+ scavenging assay19. The best scavenging results were again observed in F. praecox roots (Fig. 6b) with IC50 value 62.86 ± 0.64 µg which is significantly better than BHA (114.58 ± 4.93 µg). However, we found poor scavenging activity in all other plant organs of both F. praecox and M. pruriens. Among M. pruriens, the better scavenging capacity was shown by its leaf (228.79 ± 35.25 µg). Previous studies on other M. pruriens species have reported 41% DMPD⋅+ scavenging at 40µg113 and about 85% DMPD⋅+scavenging at 100 µg aqueous extract of its raw seeds114. We did not find any previous study on other organs of M. pruriens and on any species of Flemingia with respect to DMPD⋅+ scavenging. In our study, the obtained highest DMPD⋅+ scavenging activity of F. praecox root indicates that it contain abundant hydrophilic antioxidants as compared to M. pruriens which showed significantly lower DMPD⋅+ scavenging activity. This indicates that the antioxidant capacity of M. pruriens is mostly governed by hydrophobic antioxidants and less by hydrophilic antioxidants.
β-carotene bleaching protection assay
Lipid peroxidation protection or peroxyl radical (ROO⋅) scavenging property of the plant extracts was assessed by β-carotene bleaching assay. In this assay, the ROO⋅ generated by thermal autoxidation of linoleic acid reacts with β-carotene and causes its discolouration. This discolouration is prevented when antioxidants in plants neutralises the ROO⋅ to ROOH. This activity is an important indicator of capacity of plant extract to protect fragile sperm membrane susceptible to lipid peroxidation by ROO⋅115. In our study we observed that at a concentration of 30 µg, F. praecox roots have maximum capacity to protect β-carotene from bleaching (29.46 ± 1.40%) followed by F. praecox leaf (19.77 ± 2.71%) (Fig. 6c). M. pruriens seed protected 14.85 ± 0.30% β-carotene from bleaching. M. pruriens stem and root as well as ascorbic acid did not show any protection against the bleaching at this concentration. β-carotene bleaching protection activity of M. pruriens was also reported previously where the workers observed 5.4% bleaching protection activity by 100 µg methanolic extract of M. pruriens116 and 59.35% protection by 200 µg processed extract of another species, M. gigantia117. No study was found on any F. praecox species with respect to their in vitro β-carotene bleaching protection activity. Among the standards, BHA showed the maximum level of β-carotene protection from bleaching (73.01 ± 0.60%) followed by BHT (64.85 ± 0.38%).
Ferric ion reducing (Fe3+ → Fe2+) antioxidant power assay (FRAP)
The reducing capacity of the plant extracts was determined by FRAP assay which is based on the SET mechanism. High reducing capacity of plant extract is an indicator of its potential antioxidant capacity19. The results of TAA are presented in Table 4 (SF1b and c). The highest FRAP value was observed in M. pruriens seed (A700 = 0.194 ± 0.006 absorption units (AU)) followed by F. praecox root (A700 = 0.143 ± 0.003AU). Among standards, ascorbic acid showed the best reducing capacity (A700 = 1.148 ± 0.025AU). In previous studies on M. pruriens, FRAP activity was found to be 561 mg ascorbic equivalent/g68. However, in our analysis it was observed to have 155.90 ± 5.42 mg ascorbic equivalent/g (SF1a). The FRAP studies on two species of Flemingia, F. vestita and F. macrophylla showed FRAP values, although calculated differently, 9.28 mg GAE/g of hydroalcoholic extract74 and an IC50 of 23.05 µg/mL of aqueous extract47 respectively. Stem and leaf of both M. pruriens and F. praecox and roots of M. pruriens have shown minimum and statistically similar activity indicating their low antioxidant capacity.
Total antioxidant activity (TAA)
To determine total antioxidant activity (TAA) of the plant extract phosphomolybdenum method was used. This method evaluates both water-soluble and fat-soluble antioxidants from the plant extract118. In our study we get the highest TAA value in F. praecox root (A695 = 0.251 ± 0.002AU) followed by M. pruriens seed (A695 = 0.137 ± 0.006AU) as can be seen in Table 5 (SF2a and b). However, among the standards used, the highest TAA value was found in ascorbic acid (A695 = 0.641 ± 0.005AU) followed by BHA (A695 = 0.568 ± 0.03AU) and BHT (A695 = 0.317 ± 0.006AU). Previous studies were not found on M. pruriens and F. praecox in context of the performed assay. This result shows that F. praecox root might contain both water-soluble and fat-soluble antioxidants in abundance than M. pruriens seeds.
Any plant sample contains hundreds of compounds and its antioxidant property depends upon their physicochemical properties. Therefore, the antioxidant capacity of the plant extract or any sample should not be concluded on the basis of any single antioxidant test model. To evaluate the overall antioxidant potential of the plant extract thus required multiple antioxidant tests to be performed112. Our attempt of conducting multiple tests revealed that F. praecox root and M. pruriens seed have very high antioxidant capacity as revealed by DPPH⋅, ABTS⋅+, DMPD⋅+, FRAP, β-carotene bleaching and TAA assay. This property of F. praecox roots and M. pruriens seeds might be the major contributing factor in improving male fertility. Moreover, the phytochemically unexplored plant, F. praecox which is used by the tribal people have shown exceptional antioxidant properties even better than the conventionally used, M. pruriens. In some antioxidant aspects like DPPH, DMPD and β-carotene bleaching assay, it is showing even higher activity than the well-known synthetic antioxidants such as BHA, BHT and ascorbic acid (Table 6) thus giving the promising alternative as a rich source of natural antioxidants to prevent damage caused by oxidative stress to sperms and other important male reproductive physiological parameters.
DNA damage protection activity
DNA protection capacity of all the plant extracts against damaging agent, Fenton’s reagent was assessed by agarose gel electrophoresis. Fenton’s reagent generates highly reactive, DNA damaging hydroxyl radical (HO⋅). This radical is known to damage the DNA by oxidizing 2-deoxyribose to malonaldehyde116. Therefore, the plant antioxidants are used to assess their HO⋅ radical scavenging capacity and to protect DNA against the damage caused by the radical (Fig. 7a and b). The highest DNA protection was governed by M. pruriens seed extract which protected 98.88% DNA at 50 µg concentration followed by its stem and leaf which protected 90.92% and 87.73% respectively. In F. praecox, its roots protected the maximum 65.63% DNA followed by its leaf which showed 18.34% protection (Fig. 8 and SF3). The F. praecox stem did not show any protection against DNA damage. Previous studies shows that M. pruriens have capacity to scavenge HO• radical and protect DNA in dose dependent manner116. One study shows the methanolic extract of the M. pruriens has DNA protection capacity at IC50 value of 38 µg66. Not much work has been done before on DNA protection activity of F. praecox. However, in one study where Kim et al. isolated bioactive compound, auriculasin form F. philippinensis which showed 90.9% DNA protection at 60 µM concentration83.

DNA damage protection activity of methanolic extracts of M. pruriens seeds (a) and F. praecox roots (b). L1 = Plasmid DNA (pDNA), control; L2 = pDNA + Fenton’s Reagent. In (a) L3 = pDNA + FR + M. pruriens Seed Extract; L4 = pDNA + FR + M. pruriens Leaf Extract; L5 = pDNA + FR + M. pruriens Stem Extract; L6 = pDNA + FR + M. pruriens Root Extract. In (b) L3 = pDNA + FR + F. praecox Root Extract; L4 = pDNA + FR + F. praecox Leaf Extract; L5 = pDNA + FR + F. praecox Stem Extract. Arrows indicate distinct forms of plasmid DNA: OC (open circular); SC (supercoiled).

DNA damage protection activity of methanolic extracts of M. pruriens and F. praecox plant organs (ND not detected).
Correlation study
The different antioxidant assays studied represent their different modes of action towards different ROS and FR84. To know whether their modes of actions are correlated to their properties of eliminating ROS and FR, we studied the correlation among their activity by using Pearson’s correlation (Fig. 9 and SF4). The TPC was found positively correlated to only TFC (r = 0.76) at p < 0.05 significance level possibly because phenolics and flavonoids are structurally related. On the other hand, flavonoids have shown strong positive correlation (p < 0.01) with DPPH (r = 0.92) and ABTS (r = 0.90) showing their ability of HAT and SET to eliminate ROS and FR. Previously, researchers have also found good correlation between total polyphenols (including flavonoids) and DPPH, ABTS activities but have noted comparatively lower correlation with DMPD radical scavenging activity122. Moreover, flavonoids also displayed good correlation with β-carotene bleaching, TAA and FRAP at p < 0.05 significance level representing that they are involved in lipid peroxidation protection. These results indicate that flavonoids are better antioxidants with a wider spectrum of scavenging mechanisms than phenols which is also evident from the result of previous work21. TAA is also strongly correlated with DPPH (r = 0.89) and DMPD (r = 0.88) at p < 0.01 significance level. This might be attributed to capacity of TAA assay to measure both hydrophobic and hydrophilic antioxidants118 and thus showing cumulative activity based on the principle of both DMPD and DPPH assay which are known to be more specific to hydrophilic and hydrophobic antioxidants respectively19. Similarly, β-carotene bleaching and ABTS activity are strongly correlated (r = 0.90). This indicating the similar mechanism of HAT might be required for scavenging ROO⋅ in β-carotene bleaching assay and ABTS assay19. Finally, DNA damage protection although is positively correlated with all the assays except β-carotene bleaching and ABTS, its correlation was found to be non-significant. This suggesting that, DNA damage protection assay which is mainly based on HO• scavenging property of the compound123 may be having quite different mechanism of action towards scavenging HO⋅ than other assays tested. FRAP assay is based on ferric ion reducing capacity of antioxidants19 and under OS ferric ions can generate DNA damaging HO⋅ by Fenton’s reaction116. Our results of FRAP assay showed best activity in M. pruriens seeds and similar results were also observed in DNA damage protection assay indicating both the activities are correlated and M. pruriens seed’s metabolites may have active involvement in improving male fertility by protecting sperm DNA from HO⋅.

Similarity matrices (correlation study) between the antioxidant assays that were represented as heatmap and hierarchical clustering tree.
Collectively, our results indicate that flavonoids are the major contributing factor for the antioxidant capacity of the plant extract. Therefore, F. praecox roots being the most flavonoid rich part of the plant can serve as a better source of antioxidants than conventionally used M. pruriens seeds to protect from ROS and FR and to repair and improve the male fertility.
PDE5 and arginase inhibition activity
Both PDE5 and arginase enzymes are considered as negative regulators of erection and their over activity or expression can cause erectile dysfunction by independent mechanisms. PDE5 is known to terminate cyclic nucleotide signalling required to mediate relaxation of smooth muscle necessary for the penile erection124. Medicines like sildenafil, vardenafil and tadalafil are effective inhibitors of PDE5 thus helping in the management of erectile dysfunction125. Here we have attempted to study whether our plant extracts have any capacity to inhibit the PDE5 activity (Fig. 10). Our study revealed that F. praecox root extract at 100 µg concentration inhibited the PDE5 activity by 13.12% whereas at the same concentration M. pruriens showed inhibition activity of 4.85%. This result suggests that F. praecox might contain more effective PDE5 inhibitors than M. pruriens. However, sildenafil citrate has shown nearly similar inhibition percent to M. pruriens at 100 nM concentration (4.48%).

PDE5 inhibition activity of methanolic extract of F. praecox and M. pruriens. Different letters represent significant differences at the p < 0.05 level.
Another biomarker for erectile dysfunction studied is arginase which works by competing for the l-arginine, the substrate for the nitric oxide synthase (NOS) needed for the synthesis of nitric oxide (NO). NO is an important molecule for penile cavernosal tissue relaxation and erection126. Therefore, arginase inhibitors can enhance L-arginine bioavailability to NOS. Our study showed that both M. pruriens seed and F. praecox root extract have nearly similar arginase inhibition capacity with their IC50 value calculated to be 144.41 ± 46 µg and 146.20 ± 29.68 µg respectively (Table 7).
Conclusion
As it is evident from the previous work that ROS has a huge impact on the male fertility and its effect can be reversed with the help of antioxidants from natural sources like M. pruriens. Therefore, the aim of the present investigation was set to examine a similar role of a less explored but traditionally effective and endemic plant, F. praecox and its activity was compared with the activity of M. pruriens. This aim was investigated with the help of examining their antioxidant parameters like phenolic, flavonoid content, DPPH, ABTS, DMPD radical scavenging capacity, β-carotene bleaching protection, FRAP and TAA activity along with DNA damage protection capacity. The second aim was to directly examine comparative inhibition potential of both the plants against infertility markers PDE5 and arginase enzymes.
The study has identified F. praecox is having better antioxidant activity than M. pruriens in majority of the antioxidant assays suggesting that antioxidant potential of F. praecox may be the contributing factor for its fertility improving activity. Another significant observation about F. praecox is that its radical scavenging capacity is better than artificial antioxidants thus implicating its further use as a source of dietary antioxidants or can be used in combination with available antioxidants for better synergistic effects for improving the male fertility. The presence of a higher number of phenolic compounds in F. praecox roots compared to M. pruriens seeds, along with the diverse mechanisms by which these compounds positively influence male fertility, emphasises their potential role in enhancing male reproductive health. Finally. the finding of better PDE5 inhibition activity and similar arginase inhibition values of F. praecox in relation to its counterpart, M. pruriens is again encouraging for its further preclinical and clinical trials to study its actual potential.
Materials and methods
Plant material collection and processing
For collection of plants, all relevant permits or permissions have been obtained. The study also complies with local and national regulations. F. praecox C.B. Clarke Ex Prain was collected from Gadchiroli district of Maharashtra, India and identified by D. L. Shirodkar, botanist from Botanical Survey of India (BSI), Pune and deposited in the herbarium of BSI, Pune with identification No. BSI/WRC/Iden. Cer./2021/0911210004872. F. praecox is extremely rare in the natural habitat therefore, very few seeds of it were collected from Gadchiroli district of Maharashtra, India and then it was planted and grown for two years till its further successful seed setting has occurred. Later, its leaf, stem and roots were harvested, cleaned, washed, chopped, dried in a hot air oven at 45 °C and powdered in a mechanical grinder. M. pruriens L. was collected from RTM Nagpur University Educational Campus, Nagpur, India and identified by Prof. N. M. Dongarwar, taxonomist in Department of Botany, RTM Nagpur University, Nagpur (identification No. 187). Its seeds, leaves, stem and roots were collected and processed in a similar way like that of F. praecox. All samples were extracted in methanol by soxhlet. Also the seed of M. pruriens and roots of F. praecox were extracted sequentially in different solvents like n-hexane, ethyl acetate, chloroform, acetone and methanol with their increasing polarity then filtered and used for further analysis.
Preliminary phytochemical analysis
Preliminary phytochemical tests were done as per the standardized protocols127,128,129.
Test for phenols
Ferric chloride test
Three to four drops of 5% FeCl3 solution was added in 2 mL of crude extract of plants. Appearance of bluish black colour confirms the presence of phenols.
Test for flavonoids
Lead acetate test
1 mL of 10% lead acetate solution was added to 1–2 mL of plant extract. The appearance of blue colour confirms the presence of flavonoids.
Shinoda test
For this test, in the aqueous extract of plants some pieces of magnesium metal ribbons were added followed by addition few drops of concentrated HCl which within a minute or two gives pink, crimson or magenta colour that shows presence of flavonoids.
Alkaline reagent test
For this test 2 mL of 2% NaOH was added to 1–2 mL of aqueous extract of plants that give intense yellow colour. Addition of 3 mL of 5% HCl to it turns reaction mixture colourless indicates presence of flavonoids.
Test for alkaloids
Hager’s test
Freshly prepared Hager’s reagent (1 g picric acid in 100 mL hot water) when added to plant extract gives yellow precipitate indicating presence of alkaloids.
Dragendorff’s test
Few drops of Dragendorff’s reagent were added to the plant extract which gives orange, red or creamy precipitate confirms presence of alkaloids.
Mayer’s test
Mayer’s reagent (potassium mercuric iodide) when reacted with alkaloids in plant extract (2 mL) gives yellow, whitish or creamy precipitate.
Wagner’s test
1 mL of Wagner’s reagent added to 2 mL of plant extract, reddish brown precipitate confirms the presence of alkaloids.
Test for steroids
Salkowski test
In this test, 2 mL of extract is used and 2 mL chloroform and 1–2 mL concentrated sulphuric acid were added to it, the reddish brown colour at the junction of aqueous and chloroform layer indicates presence of steroids.
Test for tannins
Bramer’s test
2-3drops of 5% FeCl3 solution was added to diluted plant extract. Appearance of green or bluish black precipitate indicates presence of tannins.
Lead acetate test
In this test, to the 2 mL of extract 10% lead acetate solution was added. Appearance of white precipitation confirms the presence of tannins.
Potassium dichromate test
In 2 mL of plant extract, formation of red or dark coloured precipitate after addition of 10% potassium dichromate confirms the presence of tannins.
Gelatin test
1 mL of 1% gelatin solution in 10% NaCl was prepared and added to 2 mL of extract. Formation of white precipitate indicates presence of tannins.
Test for saponins
Foam test
5 mL of aqueous extract or 500 mg of dry extract was heated and shaken with 5 mL distilled water. Foam produced persisted for 10 min indicates presence of saponins.
Olive oil test
In 5 mL of extract a few drops of olive oil was added and the solution was shaken vigorously. Formation of emulsion confirms presence of saponins.
Test for glycosides
Keller-kiliani test
To the 2 mL of plant extract 1 mL of glacial acetic acid was added followed by addition of a few drops of FeCl3 and at the end 1 mL of H2SO4 added slowly and the solution allowed to settle. A reddish brown colour ring appears at the junction of two layers and the upper layer turns bluish green. These results suggest the presence of cardiac steroidal glycosides (aglycon).
Legal’s test
2 mL of concentrated extract mixed with 2 mL of pyridine, few drops of 2% freshly prepared sodium nitroprusside solution and few drops of 20% NaOH. Blue or pink coloration indicates presence of aglycon moiety.
Liebermann’s test
2 mL of extract was heated with 2 mL of acetic anhydride. After its cooling a few drops of concentrated H2SO4 was added from the sides of the test tube. Appearance of the blue or green colour precipitate indicates presence of glycosides.
Test for terpenoids
Acetic anhydride test
2 mL of acetic anhydride was added to 2 mL of extract followed by addition of 2–3 drops of concentrated H2SO4. The deep red coloration indicated the presence of terpenoids.
Chloroform test
In this test, to the 2 mL of plant extract, 2 mL chloroform was added and the solution was evaporated in a water bath to make its concentrate. Later 3 mL H2SO4 was added and the solution was boiled. The grey colour will appear when the terpenoids are present.
Total phenol content (TPC)
TPC was estimated by Folin-ciocalteu method130. In brief, 2.5 mL of 10% Folin-ciocalteu reagent and 2 mL of 7.5% sodium carbonate were added to 500 µg of extract. The reaction mixture was incubated at 45 °C for 45 min and the blue coloured phosphomolybdic/phosphotungstic acid complex was measured at 760 nm. The TPC value was calculated using gallic acid standard and presented as mg GAE/g of extract.
Total flavonoid content (TFC)
TFC was determined by aluminium chloride method131 with slight modification. 200µL of 5% sodium nitrite was added to 200 µg of extract and allowed to react for 5 min. 300µL of 10% aluminium chloride was added to the mixture and after 5 min, 2 mL of 1 M NaOH was added and the absorbance of the orange-red aluminium complex was taken at 510 nm. The TFC value was calculated using the quercetin standard and presented as mg QE/g of extract.
Phenol and flavonoid detection in plant fractions by HPLC–MS/MS analysis
One gram of dried Flemingia root powder and Mucuna seed powder were macerated in HPLC grade Methanol for 48 h. The extract was filtered by Whatman filter paper no. 1 and clear filtrate was used for the metabolome analysis by HRLC-MS/MS. The metabolomics data generated was then searched for the phenol and flavonoid compounds. Detailed set up procedure for HPLC–MS/MS instrument for the analysis is given in supplementary data file.
2, 2-Diphenyl-1-picrylhydrazyl radical (DPPH⋅) scavenging assay
DPPH⋅ scavenging assay was done as per the procedure explained by Tuba and Gulcin132 with some modification as per Kedare and Singh133. Purple coloured DPPH⋅ solution was prepared in methanol till the absorbance was achieved to 0.950 ± 0.025 at 517 nm. 3 mL methanol was added to 4, 8, 12, 16 and 20 µg of plant extract followed by addition of 1 mL DPPH⋅ solution. The reaction mixture vortexed and incubated at RT for 30 min in the dark. Absorbance of the pale yellow hydrazine product measured at 517 nm with blank containing only methanol. IC50 values of samples were calculated along with the ascorbic acid, BHA and BHT standards.
2, 2-azinobis (3-ethylbenzothiazoline-6-sulfonic acid radical (ABTS⋅+) scavenging assay
ABTS⋅+ scavenging activity of the plant extracts were determined by first generating ABTS radical cation (ABTS⋅+) by mixing 7 mM ABTS and 2.45 mM potassium persulfate in deionized water and kept at room temperature for overnight (12–16 h) and finally the absorbance of ABTS⋅+ was adjusted to 0.750 ± 0.025 at 734 nm. Later 3 mL methanol and 1 mL ABTS⋅+ solution was added to 2, 4, 6, 8 and 10 µg of plant extract. After 10 min of incubation at RT, the absorbance of decolorized/scavenged ABTS⋅+ was measured at 734 nm with blank containing only methanol134. IC50 values of samples were calculated along with the ascorbic acid, BHA and BHT standards.
N, N-dimethyl-p-phenylenediamine dihydrochloride radicle (DMPD⋅+) scavenging assay
DMPD cation radical (DMPD⋅+) generated by reacting DMPD with ferric chloride in acetate buffer. For this 500µL of 100 mM DMPD was added to 50 mL of 0.1 M acetate buffer (pH 5.3) and then 100µL of ferric chloride added to generate DMPD⋅+. Finally the absorbance of this solution was adjusted by using acetate buffer or ferric chloride to 0.900 ± 0.100 at 505 nm. Now, 2 mL of the DMPD⋅+ solution was added to 10, 20, 30, 40 and 50µL of extract and incubated at RT for 10 min and discoloration is noted at 505 nm by using acetate buffer as blank135. IC50 values of samples were calculated along with the ascorbic acid, BHA and BHT standards.
Ferric ion reducing (Fe3+ → Fe2+) antioxidant power assay (FRAP)
The FRAP assay for formation of intense perl’s prussian blue complex of the Fe2+–ferricyanide complexes from yellow coloured Fe3+–ferricyanide complexes by the reducing power of plant extract was also performed132. Briefly, different concentrations of plant extracts (5, 10, 20, 30, 40 and 60 µg) was taken and reacted with 2.5 mL of 1% potassium ferricyanide in 2.5 mL sodium phosphate buffer (0.2 M; pH 6.6) and incubated at 50 °C for 20 min. Then 2.5 mL of 10% trichloroacetic acid was added. 2.5 mL of this reaction mixture was taken then diluted with 2.5 mL distilled water and 0.5 mL of 0.1% ferric chloride was added. The absorption of the complex was measured at 700 nm.
β-carotene bleaching protection assay
A β-carotene bleaching assay was done by using protocol of Duan et al.136. Shortly, 1 mg/mL β-carotene solution was prepared in chloroform and 4 mL of it was added to 45µL of linoleic acid and 365 µL of tween-20. Chloroform was evaporated and slowly 100 mL oxygenated distilled water was added and vortexed to form emulsion and to initiate the β-carotene bleaching. 4 mL of it was added to 30 µg of the plant extract and delay in discolouration by plant extract was noted after 60 min of incubation for 45–50 °C at 470 nm.
Phosphomolybdenum method for total antioxidant activity (TAA)
In this method different concentration of plant extract (20, 40, 60, 80 and 100 µg) was reacted with 5.4 mL phosphomolybdenum reagent made up of 28 mM sodium phosphate, 4 mM ammonium molybdate and 0.6 M sulfuric acid. The reaction mixture then incubated at high temperature of 95 °C for 90 min, cooled at room temperature and subsequently the absorbance of green phosphate/Mo(V) complex formed noted at 695nm118.
DNA damage protection activity
DNA damage protection capacity of the plant extract from Fenton’s reagent was determined by using plasmid DNA as explained by Kim83 with some modifications. Briefly, in the sequence, reaction mixture of 3 µl of Plasmid DNA (0.35 µg/mL), 9 µl of 50 mM sodium phosphate buffer (pH 7.4), 2 µl of 1 mM FeSO4, 50 µg of sample and 3 µl of 30% H2O2 was prepared. Then the reaction mixture incubated at 37 °C for 30 min in the dark. 5 µl of it was loaded in 0.8% agarose gel with 1 µl of 6 × DNA loading buffer for electrophoresis for 60 min at 85 V and 90 mA. The bands generated were analyzed by using Image Lab software and percent DNA protection was calculated by comparing with control containing only plasmid DNA.
In vitro PDE5 inhibition activity
Rat lung homogenate was used as a source of PDE5 enzyme137. The homogenate (10% w/v) was centrifuged at 13,000 rpm for 20 min and supernatant was used as a source of enzyme for inhibition assay. The reaction mixture was prepared in the following sequence. 100 µg of plant extract in 5% DMSO was added to 2 mL of 20 mM Tris–HCl (pH 8.0) containing 5 mM MgCl2 followed by the addition of 100µL of enzyme extract. Finally, 100µL of 5 mM 4-nitrophenyl phenylphosphonate substrate was added to initiate the reaction. After incubation of 60 min at 37 °C, the absorbance of the hydrolysed product from substrate was noted at 400nm138 using 5% DMSO as blank and compared with the sildenafil as a positive control.
In vitro arginase inhibition activity
Arginase inhibition capacity of the plant extract was determined by using lung tissue homogenate as a source of arginase by method developed by Iyamu et al.139. The reaction mixture including 100µL enzyme extract, 100µL of 100 mM MnCl2, 1 mL of 50 mM Tris–HCl (pH 7.5) and 50µL of 0.5 M arginine substrate/ substrate with 50µL plant extract (1 mg/mL)/ substrate with 50µL DMSO (5%) incubated at 37ºC for 60 min. Then the reaction was stopped by adding 1 mL of 0.72 M HCl, the solution was centrifuged for 5 min at 5000 rpm. 1 mL of supernatant was mixed with 2 mL of 6% ninhydrin in ethanol. Finally, the solution was incubated at 60 °C for 30 min, cooled at RT and the formation of a reddish complex was noted at 505 nm. The inhibition percent was calculated by comparing the result of sample with control.
All experimental protocols were approved by the Institutional Animal Ethical Committee of Smt. Kishoritai Bhoyar College of Pharmacy, Kamptee, Nagpur, Maharashtra (IAEC approval No. 853/IAEC/22-23/23). Quarantine procedures and animal maintenance followed the recommendations of CPCSEA (Committee for the Purpose of Control and Supervision of Experiments on Animals) guidelines for laboratory animal facilities, and the methods are reported in accordance with ARRIVE guidelines.
Statistical analysis
All the analyses were performed in triplicate experiments (n = 3). The results of TPC, TFC, TAA and FRAP were calculated as mean of observations ± SD. Whereas for DPPH, ABTS and DMPD radical scavenging activities, the means of IC50 ± SD was calculated. β-carotene bleaching assay, DNA damage protection assay and in-vitro PDE5 and arginase inhibition capacity were calculated as mean of percent protection/inhibition ± SD. For defining the statistical significance between the observations, analysis of variance (ANOVA) and Tukey’s post-hoc test was applied (p < 0.05) and for Pearson’s correlational studies between antioxidant tests, multcompview and metan package in R were used.
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information file.
References
Carlsen, E., Giwercman, A., Keiding, N. & Skakkebaek, N. E. Evidence for decreasing quality of semen during past 50 years. BMJ 305, 609–613 (1992).
Swan, S. H., Elkin, E. P. & Fenster, L. The question of declining sperm density revisited: An analysis of 101 studies published 1934–1996. Environ. Health Perspect. 108, 961 (2000).
Mishra, P., Negi, M. P. S., Srivastava, M., Singh, K. & Rajender, S. Decline in seminal quality in Indian men over the last 37 years. Reprod. Biol. Endocrinol. 16, 1–9 (2018).
Hanson, B. M., Eisenberg, M. L. & Hotaling, J. M. Male infertility: A biomarker of individual and familial cancer risk. Fertil. Steril. 109, 6–19 (2018).
Ding, G. L. et al. The effects of diabetes on male fertility and epigenetic regulation during spermatogenesis. Asian J. Androl. 17, 948–953 (2015).
Ferlin, A. et al. Sperm count and hypogonadism as markers of general male health. Eur. Urol. Focus 7, 205–213 (2021).
Tvrda, E., Agarwal, A. & Alkuhaimi, N. Male reproductive cancers and infertility: A mutual relationship. Int. J. Mol. Sci. 16, 7230 (2015).
Gunes, S., Arslan, M. A., Hekim, G. N. T. & Asci, R. The role of epigenetics in idiopathic male infertility. J. Assist. Reprod. Genet. 33, 553–569 (2016).
Agarwal, A. et al. Male infertility. Lancet 397, 319–333. https://doi.org/10.1016/S0140-6736(20)32667-2 (2021).
Sansone, A. et al. Smoke, alcohol and drug addiction and male fertility. Reprod. Biol. Endocrinol. 16, 3 (2018).
Punab, M. et al. Causes of male infertility: A 9-year prospective monocentre study on 1737 patients with reduced total sperm counts. Hum. Reprod. 32, 18–31 (2017).
Kothandaraman, N., Agarwal, A., Abu-Elmagd, M. & Al-Qahtani, M. H. Pathogenic landscape of idiopathic male infertility: New insight towards its regulatory networks. NPJ Genom. Med. 1, 16023 (2016).
Agarwal, A., Virk, G., Ong, C. & du Plessis, S. S. Effect of oxidative stress on male reproduction. World J. Mens. Health 32, 1 (2014).
Adewoyin, M. et al. Male infertility: The effect of natural antioxidants and phytocompounds on seminal oxidative stress. Diseases 5, 9 (2017).
Sikka, S. Relative impact of oxidative stress on male reproductive function. Curr. Med. Chem. 8, 851–862 (2001).
Ko, E. Y., Sabanegh, E. S. & Agarwal, A. Male infertility testing: Reactive oxygen species and antioxidant capacity. Fertil. Steril. 102, 1518–1527 (2014).
Wagner, H., Cheng, J. W. & Ko, E. Y. Role of reactive oxygen species in male infertility: An updated review of literature. Arab J. Urol. 16, 35 (2018).
Agarwal, A. et al. Male oxidative stress infertility (MOSI): Proposed terminology and clinical practice guidelines for management of idiopathic male infertility. World J. Mens. Health 37, 296–312 (2019).
Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 94, 651–715 (2020).
Cipolletti, M., Fernandez, V. S., Montalesi, E., Marino, M. & Fiocchetti, M. Beyond the antioxidant activity of dietary polyphenols in cancer: The modulation of estrogen receptors (ERs) signaling. Int. J. Mol. Sci. 19, 2624 (2018).
Chun, O. K., Kim, D. O. & Lee, C. Y. Superoxide radical scavenging activity of the major polyphenols in fresh plums. J. Agric. Food Chem. 51, 8067–8072 (2003).
Spencer, J. P. E. et al. Contrasting influences of glucuronidation and O-methylation of epicatechin on hydrogen peroxide-induced cell death in neurons and fibroblasts. Free Radic. Biol. Med. 31, 1139–1146 (2001).
Dumoulin, M. et al. A camelid antibody fragment inhibits the formation of amyloid fibrils by human lysozyme. Nature 424, 783–788 (2003).
Jamalan, M., Ghaffari, M. A., Hoseinzadeh, P., Hashemitabar, M. & Zeinali, M. Human sperm quality and metal toxicants: protective effects of some flavonoids on male reproductive function. Int. J. Fertil. Steril. 10, 215–222 (2016).
Adana, M. Y. et al. Naringenin attenuates highly active antiretroviral therapy-induced sperm DNA fragmentations and testicular toxicity in Sprague-Dawley rats. Andrology 6, 166–175 (2018).
Hussein, M. M. A. et al. Amelioration of titanium dioxide nanoparticle reprotoxicity by the antioxidants morin and rutin. Environ. Sci. Pollut. Res. 26, 29074–29084 (2019).
Singh, A. P., Sarkar, S., Tripathi, M. & Rajender, S. Mucuna pruriens and its major constituent L-DOPA recover spermatogenic loss by combating ROS, loss of mitochondrial membrane potential and apoptosis. PLoS One 8, e54655 (2013).
Shukla, K. K. et al. Mucuna pruriens improves male fertility by its action on the hypothalamus-pituitary-gonadal axis. Fertil. Steril. 92, 1934–1940 (2009).
Shukla, K. K. et al. Mucuna pruriens reduces stress and improves the quality of semen in infertile men. Evid. Based Complement. Altern. Med. 7, 137–144 (2010).
Ahmad, M. K. et al. Effect of Mucuna pruriens on semen profile and biochemical parameters in seminal plasma of infertile men. Fertil. Steril. 90, 627–635 (2008).
Misra, L. & Wagner, H. Extraction of bioactive principles from Mucuna pruriens seeds. Indian J. Biochem. Biophys. 44, 56–60 (2007).
Gavade, S. K., Surveswaran, S., van der Maesen, L. J. G. & Lekhak, M. M. Taxonomic revision and molecular phylogeny of Flemingia subgenus Rhynchosioides (Leguminosae). Blumea J. Plant Taxon. Plant Geogr. 64, 253–271 (2019).
Thacker, K. D., Gavade, S. K., Lekhak, M. M., Gondaliya, A. D. & Rajput, K. S. Comparison of petiole anatomy in Flemingia and its potential for delimitation of species. Flora 278, 151790 (2021).
Li, H., Zhai, F. & Liu, Z. Chemical constituents and bioactivities of the plants of genus Flemingia Roxb. et Ait. (Leguminosae). Comb. Chem. High Throughput Screen 15, 611–622 (2012).
Gahlot, K., Lal, V. K. & Jha, S. Phytochemical and pharmacological potential of Flemingia Roxb. ex W.T.Aiton (Fabaceae). Int. J. Phytomed. 3, 294–307 (2011).
Ravi, G., Rehman, A., Koppula, S. & Veeranjaneyulu, D. Flemingia praecox var. robusta (Mukerjee) An. Kumar (Fabaceae)-An addition to the flora of Telangana 1 2 3 4. J. Indian Bot. Soc. 103, 62–66 (2023).
Gavade, S. K., van der Maesen, L. J. G. & Lekhak, M. M. Taxonomic revision of the genus Flemingia (Leguminosae: Papilionoideae) in India. Webbia 75, 141–218 (2020).
Muliar, G., Paul, A. & Kamaruz Zaman, M. Flemingia vestita benth-a highly valued medicinal and edible tuber of Meghalaya. Curr. Trends Pharm. Res. 9, 35–46 (2022).
Madan, S., Gullaiya, S., Nath Singh, G. & Kumar, Y. Flemingia strobilifera: Review on phytochemistry and pharmacological aspects. Int. J. Phytopharm. 4, 255 (2013).
Gahlot, K., Lal, V. & Jha, S. Anticonvulsant potential of ethanol extracts and their solvent partitioned fractions from Flemingia strobilifera root. Pharmacogn. Res. 5, 265 (2013).
Wang, Y. et al. Inhibition of tyrosinase activity by polyphenol compounds from Flemingia philippinensis roots. Bioorg. Med. Chem. 22, 1115–1120 (2014).
Xie, G. et al. New flavonoids with cytotoxicity from the roots of Flemingia latifolia. Fitoterapia 104, 97–101 (2015).
Roy, B. & Tandon, V. Effect of root-tuber extract of Flemingia vestita, a leguminous plant, on Artyfechinostomum sufrartyfex and Fasciolopsis buski: A scanning electron microscopy study. Parasitol. Res. 82, 248–252 (1996).
Anil Kumar, K., Dewan, B. & Rama, T. Evaluation of anti-ulcerogenic properties from the root of Flemingia strobilifera. J. Basic Clin. Pharm. 2, 33–9 (2010).
Tandon, V. & Das, B. In vitro testing of anthelmintic efficacy of Flemingia vestita (Fabaceae) on carbohydrate metabolism in Rallietina echinobothrida. Methods 42, 330–338 (2007).
Fu, M. Q. et al. Chemical constituents from roots of Flemingia philippinensis. Chin. Herb. Med. 4, 8–11. https://doi.org/10.3969/j.issn.1674-6384.2012.01.003 (2012).
Wang, B. S. et al. Antioxidant and antityrosinase activity of Flemingia macrophylla and Glycine tomentella roots. Evid. Based Complement. Alternat. Med. 2012, 1–7 (2012).
Ouedraogo, W. R. C. et al. Phytochemical study, antioxidant and vasodilatation activities of leafy stem extracts of Flemingia faginea Guill. & Perr. (Barker), a medicinal plant used for the traditional treatment of arterial hypertension. Pharmacol. Res. Mod. Chin. Med. 7, 100231 (2023).
Matsuura, H. N. & Fett-Neto, A. G. Plant alkaloids: Main features, toxicity, and mechanisms of action. Plant Toxins https://doi.org/10.1007/978-94-007-6728-7_2-1 (2015).
Bartnik, M. & Facey, P. C. Chapter 8 glycosides. Pharmacogn. Fundam. Appl. Strateg. https://doi.org/10.1016/B978-0-12-802104-0.00008-1 (2017).
Williams, D. J., Pun, S., Chaliha, M., Scheelings, P. & O’Hare, T. An unusual combination in papaya (Carica papaya): The good (glucosinolates) and the bad (cyanogenic glycosides). J. Food Compos. Anal. 29, 82–86 (2013).
Agbafor, K. N. & Nwachukwu, N. Phytochemical analysis and antioxidant property of leaf extracts of Vitex doniana and Mucuna pruriens. Biochem. Res. Int. 2011, 1–4 (2011).
Ghosal, S., Singh, S. & Bhattacharya, S. K. Alkaloids of Mucuna pruries chemistry and pharmacology. Planta Med. 19, 280–284 (1971).
Kumar, P. et al. Antiproliferative effect of isolated isoquinoline alkaloid from Mucuna pruriens seeds in hepatic carcinoma cells. Nat. Prod. Res. 30, 460–463 (2016).
Anosike, C. A., Igboegwu, O. N. & Nwodo, O. F. C. Antioxidant properties and membrane stabilization effects of methanol extract of Mucuna pruriens leaves on normal and sickle erythrocytes. J. Tradit. Complement. Med. 9, 278 (2019).
Shanmugavel, G. & Krishnamoorthy, G. Nutraceutical and phytochemical investigation of Mucuna pruriens seed. Pharma Innov. J. 7, 273–278 (2018).
Pizon, J. R. L., Nuñeza, O. M., Uy, M. M. & Senarath, W. T. P. S. K. GC-MS analysis and evaluation of in-vitro antioxidant potential and total phenolics content of wild hops (Flemingia strobilifera (L.) W. T. Aiton). Int. J. Biosci. 8, 25–32 (2016).
Mahajon, B., Remadevi, R., Sunil Kumar, K. N. & Ravishankar, B. Preliminary analysis of botanical and phytochemical features of lamalu—Root of Flemingia strobilifera (L.) W.T. Aiton. J. Tradit. Med. Clin. Naturop. 3, 1–6 (2014).
Mohini Nemkul, C., Bajracharya, G. B. & Shrestha, I. Phytochemical evaluation and in vitro antimicrobial activity of the roots of Flemingia strobilifera (l.) R. Br. J. Plant Resour. 17, 98–103 (2019).
Saio, V. & Syiem, D. Phytochemical analysis of some traditionally used medicinal plants of north-east India. J. Sci. Environ. Today 1, 6–13 (2015).
Sudhakar, Y. & Padmaja, Y. Investigation of analgesic, anti-inflammatory and antipyretic potential of ethanolic extract of arial parts of Flemingia chappar Graham. Int. J. Adv. Pharm. Biol. Chem. 3, 42–53 (2014).
Sun, F., Li, Q. & Xu, J. Chemical composition of roots Flemingia philippinensis and their inhibitory kinetics on aromatase. Chem. Biodivers. 14, e1600193 (2017).
Gumula, I. et al. Flemingins G-O, cytotoxic and antioxidant constituents of the leaves of Flemingia grahamiana. J. Nat. Prod. 77, 2060–2067 (2014).
Biozid, M. et al. Anti-oxidant effect of Flemingia stricta Roxb. leaves methanolic extract. Eur. J. Biol. Res. 8, 224–231 (2018).
Yang, R. Y. et al. Chemical constituents of the stems of Flemingia strobilifera. Chem. Nat. Compd. 52, 139–141 (2016).
Rajeshwar, Y., Senthil Kumar, G. P., Gupta, M. & Mazumder, K. Studies on in vitro antioxidant activities of methanol extract of Mucuna pruriens (Fabaceae) seeds. Eur. Bull. Drug Res. 13, 31–39 (2005).
Dhanani, T., Singh, R., Shah, S., Kumari, P. & Kumar, S. Comparison of green extraction methods with conventional extraction method for extract yield, L-DOPA concentration and antioxidant activity of Mucuna pruriens seed. Green Chem. Lett. Rev. 8, 43–48 (2015).
Iamsaard, S. et al. Evaluation of antioxidant capacity and reproductive toxicity of aqueous extract of Thai Mucuna pruriens seeds. J. Integr. Med. 18, 265–273 (2020).
Njemuwa, N. N., Dickson, N. U., Elizabeth, A. E., Uchenna, R. M. & Ogbonnaya, C. N. Evaluation of the antioxidant and anti-diabetic effect of Mucuna puriens extract. Eur. J. Med. Plants https://doi.org/10.9734/EJMP/2019/V27I230110 (2019).
Chittasupho, C. et al. Development of jelly loaded with nanogel containing natural l-dopa from Mucuna pruriens seed extract for neuroprotection in Parkinson’s disease. Pharmaceutics 14, 1079 (2022).
Jimoh, M. A., Idris, O. A. & Jimoh, M. O. Cytotoxicity, phytochemical, antiparasitic screening, and antioxidant activities of Mucuna pruriens (fabaceae). Plants 9, 1–13 (2020).
Li, L., Deng, X., Zhang, L., Shu, P. & Qin, M. A new coumestan with immunosuppressive activities from Flemingia philippinensis. Fitoterapia 82, 615–619 (2011).
Hsieh, P.-C. et al. Activities of antioxidants, α-glucosidase inhibitors and aldose reductase inhibitors of the aqueous extracts of four Flemingia species in Taiwan. Bot. Stud. 51, 293–302 (2010).
Marboh, V. & Mahanta, C. L. Characterisation and antioxidant activity of sohphlang (Flemingia vestita), a tuberous crop. J. Food Sci. Technol. 57, 3533 (2020).
Cardillo, B., Gennaro, A., Merlini, L., Nasini, G. & Servi, S. New chromenochalcones from Flemingia. Phytochemistry 12, 2027–2031 (1973).
Li, H. et al. A new benzofuran derivative from Flemingia philippinensis Merr. et Rolfe. Molecules 17, 7637–7644 (2012).
Madan, S. et al. Isoflavonoids from Flemingia strobilifera (L) R. Br. roots. Acta Pol. Pharm. 66, 297–303 (2009).
Rao, K. N. & Srimannarayana, G. Fleminone, a flavanone from the stems of Flemingia macrophylla. Phytochemistry 22, 2287–2290 (1983).
Kang, W. J. et al. New chalcone and pterocarpoid derivatives from the roots of Flemingia philippinensis with antiproliferative activity and apoptosis-inducing property. Fitoterapia 112, 222–228 (2016).
Tjahjandarie, T. S. et al. Cytotoxicity evaluation of two new chalcones from the leaves of Flemingia macrophylla (Willd.) Merr. Phytochem. Lett. 44, 78–81 (2021).
Tanjung, M. et al. Two new flavanones from the leaves of Flemingia lineata (L.) Aiton. Nat. Prod. Sci. 28, 40–43 (2022).
Theansungnoen, T. et al. Phytochemical analysis and antioxidant, antimicrobial, and antiaging activities of ethanolic seed extracts of four Mucuna species. Cosmetics 9, 14 (2022).
Kim, J. Y. et al. Antioxidant activities of phenolic metabolites from Flemingia philippinensis merr et. rolfe and their application to DNA damage protection. Molecules 23, 816 (2018).
Pisoschi, A. M., Pop, A., Cimpeanu, C. & Predoi, G. Antioxidant capacity determination in plants and plant-derived products: A review. Oxid. Med. Cell. Longev. 2016, 1–36 (2016).
Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 126, 485–493 (2001).
Spencer, J. P. E., Rice-Evans, C. & Williams, R. J. Modulation of pro-survival Akt/protein kinase B and ERK1/2 signaling cascades by quercetin and its in vivo metabolites underlie their action on neuronal viability. J. Biol. Chem. 278, 34783–34793 (2003).
Samsonowicz, M. & Regulska, E. Spectroscopic study of molecular structure, antioxidant activity and biological effects of metal hydroxyflavonol complexes. Spectrochim. Acta A Mol. Biomol. Spectrosc. 173, 757–771 (2017).
Grassi, D. et al. Tea, flavonoids, and cardiovascular health: Endothelial protection. Am. J. Clin. Nutr. 98, 1660S-1666S (2013).
Sokolov, A. N., Pavlova, M. A., Klosterhalfen, S. & Enck, P. Chocolate and the brain: Neurobiological impact of cocoa flavanols on cognition and behavior. Neurosci. Biobehav. Rev. 37, 2445–2453 (2013).
Vauzour, D., Vafeiadou, K., Rodriguez-Mateos, A., Rendeiro, C. & Spencer, J. P. E. The neuroprotective potential of flavonoids: A multiplicity of effects. Genes Nutr. 3, 115–126 (2008).
Babu, P. V. A., Liu, D. & Gilbert, E. R. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem. 24, 1777–1789 (2013).
Ko, K. P. Isoflavones: Chemistry, analysis, functions and effects on health and cancer. Asian Pac. J. Cancer Prev. 15, 7001–7010 (2014).
Ye, R. J. et al. Interplay between male reproductive system dysfunction and the therapeutic effect of flavonoids. Fitoterapia 147, 104756 (2020).
Guvvala, P. R., Ravindra, J. P., Rajani, C. V., Sivaram, M. & Selvaraju, S. Protective role of epigallocatechin-3-gallate on arsenic induced testicular toxicity in Swiss albino mice. Biomed. Pharmacother. 96, 685–694 (2017).
Hassan, E., Kahilo, K., Kamal, T., Hassan, M. & Saleh Elgawish, M. The protective effect of epigallocatechin-3-gallate on testicular oxidative stress in lead-induced toxicity mediated by Cyp19 gene/estradiol level. Toxicology 422, 76–83 (2019).
Chen, M., Liu, W., Li, Z. & Xiao, W. Effect of epigallocatechin-3-gallate (EGCG) on embryos inseminated with oxidative stress-induced DNA damage sperm. Syst. Biol. Reprod. Med. 66, 244–254 (2020).
El-Sisi, A. E., El-Sayad, M. E. & Abdelsalam, N. M. Protective effects of mirtazapine and chrysin on experimentally induced testicular damage in rats. Biomed. Pharmacother. 95, 1059–1066 (2017).
Jahan, S. et al. Ameliorative effects of rutin against cisplatin-induced reproductive toxicity in male rats. BMC Urol. 18, 107 (2018).
Wang, J.-Y. et al. Effect of spermidine on ameliorating spermatogenic disorders in diabetic mice via regulating glycolysis pathway. Reprod. Biol. Endocrinol. 20, 45 (2022).
Wang, T.-Q., Zhang, X. & Yang, J. Dynamic protective effect of Chinese herbal prescription, Yiqi Jiedu decoction, on testis in mice with acute radiation injury. Evid. Based Complement. Altern. Med. 2021, 1–16 (2021).
Tsao, C.-W., Ke, P.-S., Yang, H.-Y., Chang, T.-C. & Liu, C.-Y. Curcumin remedies testicular function and spermatogenesis in male mice with low-carbohydrate-diet-induced metabolic dysfunction. Int. J. Mol. Sci. 23, 10009 (2022).
Inanc, M. E. et al. Protective role of the dried white mulberry extract on the reproductive damage and fertility in rats treated with carmustine. Food Chem. Toxicol. 163, 112979 (2022).
Saikia, Q., Hazarika, A. & Kalita, J. C. Isoliquiritigenin ameliorates paroxetine-induced sexual dysfunction in male albino mice. Reprod. Toxicol. 117, 108341 (2023).
Powers, C. N. & Setzer, W. N. A molecular docking study of phytochemical estrogen mimics from dietary herbal supplements. Silico Pharmacol. 3, 4 (2015).
Venè, R. et al. Xanthohumol impairs human prostate cancer cell growth and invasion and diminishes the incidence and progression of advanced tumors in TRAMP mice. Mol. Med. 18, 1292–1302 (2012).
Angelino, D. et al. 5-(Hydroxyphenyl)-γ-valerolactone-sulfate, a key microbial metabolite of flavan-3-ols, is able to reach the brain: Evidence from different in silico, in vitro and in vivo experimental models. Nutrients 11, 2678 (2019).
Couture, R. et al. Luteolin modulates gene expression related to steroidogenesis, apoptosis, and stress response in rat LC540 tumor Leydig cells. Cell Biol. Toxicol. 36, 31–49 (2020).
dos Borges, C. S. et al. Long-term adverse effects on reproductive function in male rats exposed prenatally to the glucocorticoid betamethasone. Toxicology 376, 15–22 (2017).
Halliwell, B. Reactive oxygen species in living systems: Source, biochemistry, and role in human disease. Am. J. Med. 91, S14–S22 (1991).
Brieger, K., Schiavone, S., Miller, F. J. & Krause, K. H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 142, w13659–w13659 (2012).
Sharma, R. K. & Agarwal, A. Role of reactive oxygen species in male infertility. Urology 48, 835–850 (1996).
Alam, M. N., Bristi, N. J. & Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 21, 143–152 (2013).
Aware, C. et al. Processing effect on l-dopa, in vitro protein and starch digestibility, proximate composition, and biological activities of promising legume: Mucuna macrocarpa. J. Am. Coll. Nutr. 38, 447–456 (2019).
Patil, R. R., Rane, M. R., Bapat, V. A. & Jadhav, J. P. Phytochemical analysis and antioxidant activity of Mucuna sanjappae: A possible implementation in the Parkinson’s disease treatment. J. Pharm. Med. Res. 2, 48–51 (2016).
Aitken, R. J. A free radical theory of male infertility. Reprod. Fertil. Dev. 6, 19–24 (1994).
Siddhuraju, P. & Becker, K. Studies on antioxidant activities of mucuna seed (Mucuna pruriens var utilis) extract and various non-protein amino/imino acids through in vitro models. J. Sci. Food Agric. 83, 1517–1524 (2003).
Vadivel, V. & Biesalski, H. K. Total phenolic content, antioxidant activity, and type II diabetes related functionality of traditionally processed ox-eye bean [Mucuna gigantea (Willd) DC.] seeds: An indian underutilized food legume. Food Sci. Biotechnol. 20, 783–791 (2011).
Prieto, P., Pineda, M. & Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 269, 337–341 (1999).
Agarwal, A., Virk, G., Ong, C. & Plessis, S. S. D. Effect of oxidative stress on male reproduction. World J. Mens. Health 32, 1–17 (2014).
Aitken, R. Free radicals, lipid peroxidation and sperm function. Reprod. Fertil. Dev. 7, 659 (1995).
Maneesh, M. & Jayalekshmi, H. Role of reactive oxygen species and antioxidants on pathophysiology of male reproduction. Indian J. Clin. Biochem. 21, 80–89 (2006).
Fernández-Pachón, M. S., Villaño, D., García-Parrilla, M. C. & Troncoso, A. M. Antioxidant activity of wines and relation with their polyphenolic composition. Anal. Chim. Acta 513, 113–118 (2004).
Imlay, J. A., Chin, S. M. & Linn, S. Toxic DNA damage by hydrogen peroxide through the Fenton reaction in vivo and in vitro. Science 240, 640–642 (1988).
Gratzke, C. et al. Anatomy, physiology, and pathophysiology of erectile dysfunction. J. Sex. Med. 7, 445–475 (2010).
Samplaski, M. K. & Nangia, A. K. Adverse effects of common medications on male fertility. Nat. Rev. Urol. 12, 401–413 (2015).
Masuda, H. Significance of nitric oxide and its modulation mechanisms by endogenous nitric oxide synthase inhibitors and arginase in the micturition disorders and erectile dysfunction. Int. J. Urol. 15, 128–134 (2008).
Shaikh, J. R. & Patil, M. Qualitative tests for preliminary phytochemical screening: An overview. Int. J. Chem. Stud. 8, 603–608 (2020).
Evans, W. C. Trease and Evans’ Pharmacognosy 16th edn, 1–603 (Elsevier Health Sciences, 2009).
Harborne, J. B. Phytochemical Methods: a Guide to Modern Techniques of Plant Analysis (Chapman and Hall, 1998).
Ordoñez, A. A. L., Gomez, J. D., Vattuone, M. A. & Isla, M. I. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 97, 452–458 (2006).
Wolfe, K., Wu, X. & Liu, R. H. Antioxidant activity of apple peels. J. Agric. Food Chem. 51, 609–614 (2003).
Ak, T. & Gülçin, I. Antioxidant and radical scavenging properties of curcumin. Chem. Biol. Interact. 174, 27–37 (2008).
Kedare, S. B. & Singh, R. P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 48, 412 (2011).
Mandade, R., Sreenivas, S. A. & Choudhury, A. Radical scavenging and antioxidant activity of Carthamus tinctorius extracts. Free Radic. Antioxid. 1, 87–93 (2011).
Fogliano, V., Verde, V., Randazzo, G. & Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 47, 1035–1040 (1999).
Duan, X. J., Zhang, W. W., Li, X. M. & Wang, B. G. Evaluation of antioxidant property of extract and fractions obtained from a red alga, Polysiphonia urceolata. Food Chem. 95, 37–43 (2006).
Temkitthawon, P. et al. Kaempferia parviflora, a plant used in traditional medicine to enhance sexual performance contains large amounts of low affinity PDE5 inhibitors. J. Ethnopharmacol. 137, 1437–1441 (2011).
Ademiluyi, A. O., Oyeleye, S. I., Ogunsuyi, O. B. & Oboh, G. Phenolic analysis and erectogenic function of African Walnut (Tetracarpidium conophorum) seeds: The impact of the seed shell on biological activity. J. Food Biochem. 43, e12815 (2019).
Iyamu, E. W., Asakura, T. & Woods, G. M. A colorimetric microplate assay method for high-throughput analysis of arginase activity in vitro. Anal. Biochem. 383, 332–334 (2008).
Acknowledgements
We acknowledge the support from University Grant Commission, India for the fellowship received to SDK (UGC-Ref. No.: 853/(OBC) (CSIR-UGC NET DEC. 2016)).
Author information
Authors and Affiliations
Contributions
Conceptualization by D.P.G. and S.D.K., analysis by S.D.K. and S.S.U., interpretation of data and writing original draft by S.D.K., reviewing and editing by D.P.G. and S.D.K. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kumbhare, S.D., Ukey, S.S. & Gogle, D.P. Antioxidant activity of Flemingia praecox and Mucuna pruriens and their implications for male fertility improvement. Sci Rep 13, 19360 (2023). https://doi.org/10.1038/s41598-023-46705-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-46705-9
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
