
Abstract
Extraintestinal pathogenic Escherichia coli (ExPEC) producing extended-spectrum β-lactamases (ESBL) cause serious human infections due to their virulence and multidrug resistance (MDR) profiles. We characterized 144 ExPEC strains (collected from a tertiary cancer institute) in terms of antimicrobial susceptibility spectrum, ESBL variants, virulence factors (VF) patterns, and Clermont’s phylogroup classification. The developed multiplex recombinase polymerase amplification and thermophilic helicase-dependent amplification (tHDA) assays for blaCTX-M, blaOXA, blaSHV, and blaTEM detection, respectively, were validated using PCR-sequencing results. All ESBL-ExPEC isolates carried blaCTX-M genes with following prevalence frequency of variants: blaCTX-M-15 (50.5%) > blaCTX-M-55 (17.9%) > blaCTX-M-27 (16.8%) > blaCTX-M-14 (14.7%). The multiplex recombinase polymerase amplification assay had 100% sensitivity, and specificity for blaCTX-M, blaOXA, blaSHV, while tHDA had 86.89% sensitivity, and 100% specificity for blaTEM. The VF genes showed the following prevalence frequency: traT (67.4%) > ompT (52.6%) > iutA (50.5%) > fimH (47.4%) > iha (33.7%) > hlyA (26.3%) > papC (12.6%) > cvaC (3.2%), in ESBL-ExPEC isolates which belonged to phylogroups A (28.4%), B2 (28.4%), and F (22.1%). The distribution of traT, ompT, and hlyA and phylogroup B2 were significantly different (P < 0.05) between ESBL-ExPEC and non-ESBL-ExPEC isolates. Thus, these equipment-free isothermal resistance gene amplification assays contribute to effective treatment and control of virulent ExPEC, especially antimicrobial resistance strains.
Similar content being viewed by others

Introduction
Extended-spectrum β-lactamases (ESBLs) in Enterobacteriaceae are categorized by World Health Organization as the most critical cause of antimicrobial resistance (AMR) requiring discovery of new antibiotics1. In addition, most ESBL-producing Enterobacteriaceae are also multidrug-resistant (MDR), burdening the treatment2. Extraintestinal pathogenic Escherichia coli (ExPEC) is a major ESBL-producing organism which in addition to intestine, infects, urinary tract, bloodstream, meningitis, and wounds and causes sepsis. ESBL associated AMR in ExPEC is not only disseminated in health care settings but also in community-acquired infections3. The global increase of ESBL-ExPEC strains is causing clinical and economic losses similar in magnitude to that caused by pathogenic E. coli. Unlike for, intestinal pathogenic E. coli or commensal E. coli, defining the origin or primary reservoir of ExPEC is the major challenge in its treatment4. Moreover, the influence of AMR and virulence factors (VF) genes on ExPEC pathogenicity has become a serious global concern. Thus, researchers majorly rely on ExPEC genotyping to explore the association between AMR genes, VF, and their phylogenetic distribution.
The distribution of ESBL-producing E. coli (ESBL-E coli) in extraintestinal infections is diverse and varies among different geographical regions. The clonal spread of E. coli ST131 (associated with ExPEC infections, particularly urinary tract and blood stream infections) contributed to globally disseminated MDR clone5. Among the ESBL genes, blaCTX-M-15 is highly prevalent, followed by CTX-M, TEM, SHV, PER, VEB, GES, BES, TLA, and OXA genes6. The commensal E. coli in healthy cattle, pigs, and chickens serve as a reservoir of AMR genes7. The prevalence of CTX-M genes is higher in uropathogenic E. coli (UPEC) than in the commensal isolates from healthy volunteers8. Moreover, ESBLs producing Enterobacterales (ESBL-Enterobacterales) can colonize long-term (> 12 months) as intestinal microbiota9 enhancing the spread of ESBL AMR in a given health system, including humans, animals, and environments10. The widespread presence of ESBL genes in systemic infections also significantly impacts therapeutic and mortality outcomes. Clinical and Laboratory Standard Institute (CLSI) recommends phenotypic screening and confirmatory testing for ESBLs production as a part of regular clinical treatment of microbial infections11. However, the genotypic methods of ESBL screening are more advantageous for epidemiological management and help in overcoming challenges associated with phenotypic expression variance12.
According to Ambler’s molecular classification, ESBLs are serine β-lactamases comprising three significant class A genes (blaCTX-M, blaTEM, and blaSHV), and a class D blaOXA gene. Recently, various newly discovered isothermal nucleic acid amplification techniques, which do not require expensive thermal cycling machines, have been extensively applied for rapid and simple detection of AMR genes13. In 2021, a 10 min naked-eye rapid multiplex recombinase polymerase amplification (RPA) lateral flow assay was developed to detect the three common ESBLs genes (blaCTX-M, blaOXA, and blaSHV)14. However, the use of engineered recombinase enzyme from E. coli K12 or BL21 strains (carrying their own blaTEM gene) proved to be a major setback; the use of these RPA kits was plagued with cross reactivity and false positive results15. Helicase-dependent amplification technique, like RPA, utilizes a primer pair to amplify a specific DNA/RNA target at a set temperature. To unwind double-stranded DNA and generate a new amplicon, the RPA kit (TwistDx) uses recombinase and DNA polymerase with strand displacement activity (Sau polymerase), whereas the IsoAmp® II Universal tHDA kit (New England Biolabs) employs a thermostable helicase Tte-UvrD from Thermoanaerobacter tengcongenesis and Bacillus stearothermophilus DNA polymerase (Bst-DNA polymerase)16. The optimum amplification conditions for RPA and tHDA are 37–42 °C for 20–40 min and 60–65 °C for 90–120 min, respectively17. Both, RPA and tHDA, are portable nucleic acid detection assays ideally suited for use in point-of-care or resource-limited field settings13.
The ExPEC pathogenicity is influenced by several VF genes, with wide functions, ranging from bacterial colonization to virulence18. Moreover, most VF genes are encoded by plasmids, pathogenicity islands (PAI)18,19 or other mobile genetic elements18 which horizontally transfer to other pathogenic and non-pathogenic bacteria. E. coli is classified into eight phylogenetic groups (A, B1, B2, C, D, E, F, and Clade I) using the easier quadruplex PCR (which is less complex and more rapid than multilocus sequence typing or ribotyping methods)20. The association of ESBLs, VF genes, and phylogroups has been investigated in both ExPEC and commensal E. coli. Milenkov et al., found that blaCTX-M-15 was the most prevalent ESBL gene in E. coli specimens isolated from healthy pregnant Madagascar woman. In addition, 90% of these isolates belonged to phylogenetic groups A, B1, and C (which are associated with commensalism and carry a few VF genes involved in adhesion and iron acquisition). While, 10% of these isolates belonged to the extraintestinal virulent phylogenetic groups B2, D, and F21. Persistent long-term carriage (average 3.5 months) of ESBL-Enterobacterales (which belonged to extraintestinal virulence associated phylogroup B2/D/F) was observed in travelers visiting tropical areas, 3 or more months after their return. Whereas, commensal associated phylogroups A/B1/E persisted for shorter carriage durations (approx. 0.5 months)21. Phylogroup B2 of uropathogenic E. coli (UPEC) exhibited maximum AMR and carried six VFs22. ESBL-ExPECs carrying VF genes, isolated from bloodstream infections, showed the following order of phylogroup predominance: B2 (45.8%) > B1 (18.8%) > E (14.6%). The ESBL blaCTX-M-15 and VF traT genes were the most predominant23. E. coli, the most frequent pathogen, second only to group B streptococci, causing neonatal meningitis in early-onset infections, belonged to extraintestinal phylogroup B2; > 70% of this pathogenic E. coli strain carry kpsII, K1, neuC, iucC, sitA, and vat genes. In contrast, E. coli obtained from healthy individuals belonged to groups A and D; they carry < 27% of VF genes24,25.
We aimed to develop isothermal amplification assays to characterize ESBL genes (blaCTX-M, blaOXA, blaSHV, and blaTEM) in ESBL-ExPEC strains. The sensitivity and specificity of these assays were validated using nucleotide sequencing. These simple isothermal platforms can be established in low resources settings for monitoring ESBL in all clinical samples. Moreover, we characterized and phylogenetically analyzed VF genes (traT, ompT, iutA, fimH, iha, hlyA, papC, and cvaC) found in the ESBL-ExPEC clinical isolates. Identifying associations between ESBL genes, VF genes, and ESBL-ExPEC phylogroups is important for improved global AMR surveillance, targeted antibiotic treatment, and infection control.
Results
Identification of ESBLs, their variants, and antimicrobial susceptibility pattern
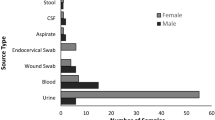
Of the 144 ExPEC isolated from various extraintestinal specimens including blood, urine, pus, body fluid, and sputum, 95 ESBL- and 49 non-ESBL-ExPEC strains were identified by combination disk method given in CLSI guideline11. All 95 ESBL-producing E. coli were subjected to genotyping. We found blaCTX-M genes (n = 95) in single or in combination with blaTEM (n = 40), blaOXA (n = 29), and blaSHV (n = 2) genes (Supplementary Table S1). In addition, 21 out of 49 phenotypically screened non-ESBL-producing E. coli harbored blaTEM-1 gene, while none of blaTEM, blaOXA, blaSHV, and blaTEM genes were observed in the rest. The PCR-sequencing results of CTX-M variants revealed the following order of prevalence: blaCTX-M-15 (50.5%) > blaCTX-M-55 (17.9%) > blaCTX-M-27 (16.8%) > blaCTX-M-14 (14.7%). The blaSHV was found only in 2 isolates. The antibiotic susceptibility pattern of most common ESBL–ExPEC variants (CTX-M-15, CTX-M-27, CTX-M-14, and CTX-M-55 types) showed 100% resistance to cefotaxime and cefdinir as described in Table 1. The following order (high–low) was observed in MDR toward ceftriaxone, cefepime, ciprofloxacin, ceftazidime, levofloxacin, and gentamycin: blaCTX-M-15 isolates > blaCTX-M-55 > blaCTX-M-27 isolates.
Development of multiplex RPA and tHDA assays
The optimum concentration of blaCTX-M, blaOXA, and blaSHV primers required in multiplex RPA reaction were 0.2 µM CTX-M, 0.1 µM OXA and 0.1 µM SHV (Fig. 1A), respectively. All products were amplified at 37–39 °C (Fig. 1B) at 25–30 min (Fig. 1C). The DNA template concentration ranging at 12.5–25 ng showed the apparent bands of all 3 amplicons (Fig. 1D). The blaSHV amplicon was absent at 41℃ and incubation duration < 25 min. Finally, the following multiplex RPA reaction condition was chosen as optimum: 37 °C for 25 min using 25 ng template DNA.

Optimization of the RPA assay. Agarose gel electrophoresis shows the blaCTX-M, blaOXA, and blaSHV amplicons (A) when 0.2 µM CTX-M, 0.1 µM OXA, 0.1 µM SHV and 0.2 µM CTX-M, 0.05 µM OXA and 0.05 µM SHV were used; (B) at 37–41 °C; (C) at 10–30 min incubation; and (D) at different DNA template concentrations (1.5–25 ng) [M, 100 bp Marker; C+, positive DNA control; C−, no template control].
The tHDA was employed instead of RPA to amplify 111 bp of blaTEM. The blaTEM amplicon was observed at 65–67 °C (Fig. 2A), with 30–90 min incubation (Fig. 2B). The optimum DNA template and primers concentrations were 50–100 ng (Fig. 2C) and 0.025–0.05 µM, respectively (Fig. 2D). The following tHDA condition was selected as optimum: 65 °C for 30 min using 50 ng template DNA and 0.025 µM primers.

Optimization of the tHDA assay. Agarose gel electrophoresis showed the blaTEM amplicons (A) at 59–67 °C; (B) at 15–90 min; (C) at different DNA template concentrations (0.05–100 ng); and (D) at different primers concentrations (0.025–0.075 µM) [M, 100 bp Marker; C+, positive DNA control; C−, no template control].
Limit of detection (LOD) and specificity of multiplex RPA and tHDA assays
The DNA templates were isolated from E. coli clinical strain ESBL120 carrying blaCTX-M, strain KP125 carrying blaOXA, strain EC137 carrying blaTEM, and K. pneumoniae ATCC 700,603 carrying blaSHV. The LOD of multiplex RPA assays for blaCTX-M, blaOXA, blaSHV genes (Fig. 3A–C) was 5 ng, 0.5 ng, and 0.5 ng, respectively. The sensitivity was 10–100 times higher than that of tHDA assay for blaTEM (LOD of 50 ng) (Fig. 3D).

(A–C) LODs for blaCTX-M, blaOXA, and blaSHV detection by RPA assay; (D) LODs for blaTEM detection by tHDA; (E) Specificity of the RPA assay for blaCTX-M, blaOXA, and blaSHV genes; and (F) Specificity of the tHDA assay for blaTEM gene [M, 100 bp Marker; C+, positive DNA control; C−, no template control].
Amplicons were absent (Fig. 3E and F) in RPA and tHDA assays of the DNA of the following organisms: E. coli ATCC 25,922, Proteus mirabilis ATCC 25,933, Pseudomonas aeruginosa ATCC 27,853, Acinetobacter baumannii ATCC 19,606, Staphylococcus aureus ATCC 25,923 and Enterococcus faecalis ATCC 29,212. A 214 bp amplicon was produced from DNA of K. pneumoniae ATCC 700,603, which naturally contains blaSHV.
Validation of multiplex RPA and tHDA assays
A total of 95 ESBL-ExPEC and 49 non-ESBL-ExPEC were subjected to multiplex RPA and tHDA assays. The results of blaCTX-M, blaOXA, and blaSHV genes from multiplex RPA assays were in concordance with PCR-sequencing results exhibiting 100% sensitivity (95% CI = 96.19–100%, 88.06–100%, 15.81–100%, respectively), 100% specificity, positive predictive value (PPV), and negative predictive value (NPV) (95% CI: 92.75–100%, 96.84–100%, and 97.44–100%, respectively) (Table 2). While tHDA assay showed eight false-negative samples of blaTEM gene yielding 86.89% (95% CI = 93.28–100), 100% (95% CI = 83.41–96.13%), 100% (95% CI = 28.93–45.42%), and 91.21% (95% CI = 77.37–92.78%) sensitivity, specificity, PPV, and NPV, respectively. The identification accuracy for blaCTX-M, blaOXA, and blaSHV genes were 100%, and blaTEM gene was 94.44%.
Distribution of VF genes, phylogroups among ESBL variants
In the 95 ESBL-ExPEC clinical isolates, the VF genes exhibited the following order in frequency: traT (67.4%) > ompT (52.6%) > iutA (50.5%) > fimH (47.4%) > iha (33.7%) > hlyA (26.3%) > papC (12.6%) > cvaC (3.2%), respectively (Table 3). All VF genes were distributed among CTX-M variants, excepting papC and cvaC that were not found in the CTX-M-27 variant. The traT gene was found frequently in CTX-M-15 (75%), CTX-M-14 (64.3%), and CTX-M-55 (76.5%). While iha gene was predominate in CTX-M-27 (70.6%). Most common phylogroups among ESBL-ExPEC strains included: A (28.4%), B2 (28.4%), F (22.1%). The CTX-M-14, 15, and 55 (35.7%, 29.2%, and 47.1%) were predominant in phylogroup A, while most of CTX-M-27 belonged to phylogroup B2 (70.6%). Only CTX-M-15, and CTX-M-55 variants were found in the rare phylogroups B1, and E, respectively.
Relationships of VF genes, phylogenetic groups, and antibiotic resistance
ESBL-ExPEC and non-ESBL-ExPEC isolates predominantly carried traT and ompT, respectively (Table 4). Surprisingly, cvcC was absent in our non-ESBL isolates. The association of traT, ompT, and hlyA were significantly different (P < 0.05) between the ESBL-ExPEC and non-ESBL-ExPEC isolates. Phylogroup predominance was as follows: phylogroup B2 (35.4%) > A (23.6%) > F (19.4%). ESBL-ExPEC were predominated by phylogroups A and B2, while non-ESBL-ExPEC were predominated by phylogroup B2. Only phylogroup B2 was significantly different between ESBL and non-ESBL groups (P < 0.05). Phylogroup Clades I was absent in all clinical isolates. Analysis of variance by Friedman’s test revealed a significant difference in VF gene distribution (P = 0.000). Three VF genes (hlyA, iha, and ompT) were distributed differently across phylogroups (Table 5). Pairwise analysis of phylogroup showed that hlyA was associated with phylogroup A and iha was associated with phylogroups F, A, and B2 (P < 0.05). Whereas, ompT was associated with phylogroups B2 and F (P < 0.05).
Discussion
ExPEC is a major cause of morbidity and mortality in both hospital and community-acquired infections. Apart from epidemiological factors, acquisition of VFs and AMR, is likely to contribute to the global pandemic of ExPEC lineages4,26. Moreover, plasmid-mediated horizontal transfer of ESBL genes occur easily among species, causing widespread infection6. Accumulating evidence suggested that VFs helps gastrointestinal pathogens outcompete the commensal microbiota and impair host immunity via inducing colonization, resistance, and invasion27.
ESBL-Enterobacteriaceae, especially E. coli was one of the most frequent isolates in blood stream infection samples collected from a pediatric oncology center28. The MDR E. coli from febrile neutropenic cancer patients showed high resistance to ampicillin, cefepime, ceftriaxone, and cephradine29. Here, all ESBL-ExPEC isolated from cancer patients carried blaCTX-M, showing the following predominance: blaCTX-M1 group (68.4%; 50.5% blaCTX-M-15 and 17.9% blaCTX-M-55) > blaCTX-M9 group (31.5%; 16.8% blaCTX-M-27 and 14.7% blaCTX-M-14). MDR was observed in all CTX-M variants. They were resistant to ceftriaxone, cefepime, ciprofloxacin, ceftazidime, levofloxacin, and gentamycin. Similarly, ESBL-ExPECs (from tertiary hospitals in Thailand) predominantly carried blaCTX-M1 (71.23%) and blaCTX-M9 (38.95%)30. The global pathogenic E. coli ST 131 strain harbors blaCTX-M-15 (67.6%), blaCTX-M-27 (20.6%), and blaCTX-M-14 (11.8%)31. Globally, the blaCTX-M-15 is frequently reported ESBL gene, especially in the bloodstream and urinary tract infections23,32,33,34. The blaCTX-M-55 is present in most E. coli isolated from pork and fecal samples14,35.
Several assays for genotyping ESBL genes, exist: (1) Kanokudom et al.,14 developed a naked-eye rapid multiplex RPA assay for detecting blaCTX-M, blaOXA, and blaSHV in pork E. coli isolates. Amplicons were visualized on a single-stranded tag hybridization chromatographic printed-array strip (STH-PAS, a commercial lateral flow assay strip); (2) Higgins et al.,36 set up a portable loop-primer endonuclease cleavage-loop-mediated isothermal amplification (loop-primer endonuclease cleavage-LAMP) assay for detecting blaCTX-M-1 and blaCTX-M-15 in porcine fecal E. coli isolates; and (3) Wang et al. developed a probe-based real-time PCR assay for detecting blaCTX-M, blaTEM, and blaSHV in broiler chicken E. coli isolates37.
Here, we developed the following isothermal assays to detect common ESBL genes: (1) a multiplex RPA assay to detect blaCTX-M, blaOXA, and blaSHV genes; and (2) a tHDA assay to detect blaTEM gene. The amplicons were visualized by agarose gel electrophoresis method, which is a cheap and widely used technique in general molecular laboratory. These simple isothermal platforms for nucleic acid amplification utilize only one pair of primers (like PCR) and the commonly available heating instruments. Moreover, compared to other isothermal platforms, the RPA kit (TwistAmp Basic kit) is a highly stable lyophilized reagent with long shelf life38. The lyophilized pellets in RPA kit are stable for at least one year when stored at temperatures below − 15 °C or at 2–8 °C, and up to 6 months at room temperature (22–28 °C)39. However, the RPA kit is slightly more expensive (~ $4.3 USD) than other amplification assays, such as PCR and LAMP (expired patents)38. Further, RPA patent is set to expire in 202340, making way for the development of a new cost-effective in-house RPA formula which will allow for its large scale applications.
The nucleotide compositions and length of primers affect the optimum annealing temperature and incubation time in multiplex RPA reactions. Here high temperatures (leading to poor primers binding) or short incubation times did not yield blaSHV amplicons, probably because the blaSHV primers have lower GC contents and shorter lengths than blaCTX-M and blaOXA primers. High primer concentrations increases the chance of primer-dimer and non-specific amplicon formation in multiplex RPA reactions, while low concentrations may lead to low yields. Therefore, good primer designing and extensive optimization are critical for effective multiplexed isothermal amplification41. The LOD of our multiplex RPA-gel electrophoresis assay in detecting blaCTX-M, blaOXA, and blaSHV genes was slightly lower than that of the previous multiplex RPA-Lateral Flow Assay (LFA)14. RPA amplicons must be purified before loading onto the agarose gel to remove protein contaminants42. This RPA post-amplification purification step causes amplicon loss. Moreover, the detection sensitivity using agarose gel electrophoresis depends on efficacy of various steps (pre-loading, pre-casting, and post-staining)43. Thus, the RPA post-amplification purification is a major drawback for gel electrophoresis detection44. Several methods for RPA products purification exist, including heat denaturation (65 °C or 95 °C for 10 min), sodium dodecyl sulfate treatment, proteinase K digestion, protein sedimentation via high-speed centrifugation and purification using commercial DNA purification kits44. Although agarose gel electrophoresis is a common method for visualization of amplification products, it is time consuming due to gel preparation, electrophoresis, gel staining, and imaging steps. These limitations can be circumvented in the future—by using various in-house detection methods (which employ bridge flocculation, SYBR green I or lateral flow assays)—to develop a cost-effective, simple, equipment-free, rapid, and naked-eye assay for detecting the desired genes39,44.
Assessing LOD of tHDA assay for blaTEM required 10x–100 × the template concentration used in RPA assays for blaCTX-M, blaOXA, blaSHV genes. The high-temperature tHDA operating condition (60–65 °C) may affect the primer binding efficiency; however specificity could be higher than mesophilic isothermal amplification38. Moreover, compared to RPA and PCR, we used lower primer concentrations (0.1–1 μM vs. 75–100 nM) for tHDA assay. The tHDA amplicon is generally < 150 nucleotides long, due to helicase processivity which limits the multiplexing capacity41,45. Our multiplex RPA assay showed 100% sensitivity and specificity for blaCTX-M, blaOXA, and blaSHV genes. However, tHDA had 86.89% sensitivity and 100% specificity for blaTEM. The higher LOD and lower primer concentrations used may have caused tHDA assay to have lower sensitivity, as compared to the RPA assay. The false-negative results may occur for samples with low quantity and quality of DNA (old/degraded DNA).
ExPEC strains, compared to commensal E. coli strains, have complex phylogenetic structure and diverse VFs4. ExPEC strains, such as uropathogenic E. coli (UPEC), neonatal meningitis E. coli (NMEC), sepsis-associated E. coli (SEPEC), and avian pathogenic E. coli (APEC), have several VFs. ExPEC VFs—such as adhesins, toxins and iron acquisition, lipopolysaccharides, capsules, and invasins—facilitate their colonization and systemic infection. UPEC prevalently causes urinary tract infection and secondary bacteremia18. Here, three VF genes (traT, ompT, and hlyA) were significantly associated with ESBL-ExPEC strains. traT was the most prevalent one, present in CTX-M-14, 15, and 27 variants. Other prevalent VF genes were aerobactin acquisition (iutA) and adhesins (fimH and iha). While a few cvaC existed only in our ESBL-ExPEC clinical isolates. The serum resistance gene traT, which inhibits the classical pathway of complement activity, is present in all pathotypes with more prevalence in UPEC46,47,48. The outer membrane protein (ompT) associated with UPEC enables intracellular survival and evasion from the host defense. Hemolysin A (hlyA) is a membrane lysis toxin present in UPEC strains. The afimbrial adhesin (afa) was found associated with bacteremia mortality23.
Only phylogroup B2 was significantly different between ESBL-ExPEC and non-ESBL-ExPEC. Similarly, the extended-spectrum cephalosporin resistant E. coli isolates belonging to phylogroup B2 carried ESBL and/or plasmid-mediated AmpC genes; blaCTX-M-15, blaCTX-M-14, blaCTX-M-55 were the most prevalent49. The phylogroup A and F are commensal and avian associated strains50, respectively. They are the second and third abundant phylogroups in our E. coli isolates. In addition, only three VF genes (ompT, iha, and hlyA) were distributed differently across phylogroups, especially in B2, F, and A. The phylogroup B2 was the most prevalent in clinical isolates (from many systemic infections) and carried a greater number of VFs than other phylogroups4. Although phylogroup F was less virulent than B2, its clinical significance in mediating AMR in ExPEC is established49,51,52.
In conclusion, most ESBL-ExPECs possessed a MDR pattern belonging to phylogroup B2 and carried more VF genes. The traT, ompT, and hlyA were found associated with ESBL-ExPEC strains. The relationship of VF genes was first demonstrated across phylogroups with diverse ompT, iha, and hlyA. The E. coli pathotypes should be characterized in the future. The identification and characterization of AMR, VF genes, and their pathotypes and phylogenetic analysis may contribute to developing novel strategies for treating E. coli-mediated systemic infection. Moreover, our multiplex RPA and tHDA assays, presented here, are simple, rapid, and reliable in detecting the most common ESBLs genes. These assays are beneficial for strategizing targeted therapy in low resource settings (using common heating equipments) and epidemiological control.
Materials and methods
Bacterial isolates
A total of 144 E. coli clinical isolates from various specimens including blood, urine, pus, body fluid, and sputum were collected from a tertiary cancer institute in Bangkok (Thailand) between February 2017–September 2018. All isolates were preserved in skimmed milk at − 80 °C until use.
The sample size used here was calculated using the Buderer method,53 which helps evaluate sensitivity and specificity of diagnostic tests at a 95% confidence interval. The study was approved by the Ethical Committee of the National Cancer Institute, Thailand (certificate number 020/2562).
Antimicrobial susceptibility testing and ESBLs detection
Ninety-five ESBL-ExPEC and forty-nine non-ESBL-ExPEC isolates were selected consecutively and non-duplicate from clinical samples by phenotypic ESBLs screening. Their antimicrobial susceptibility to various antibiotics—including, amikacin (30 µg), gentamicin (10 µg), ampicillin (10 µg), amoxicillin/clavulanic acid (20/10 µg), ceftazidime (30 µg), cefotaxime (30 µg), cefdinir (5 µg), cefepime (30 µg), ciprofloxacin (5 µg), ceftriaxone (30 µg), doripenem (10 µg), ertapenem (10 µg), imipenem (10 µg), meropenem (10 µg), levofloxacin (5 µg), and piperacillin/tazobactam (1.25/23.75 µg)—was determined using disk diffusion method. The ESBLs confirmatory testing was carried out using the combination disk method according to the CLSI guidelines11. ESBLs was considered to be produced when size of either inhibition zones of cefotaxime/clavulanic acid (30/10 µg) or ceftazidime/clavulanic acid (30/10 µg) were ≥ 5 mm compared to that of cefotaxime (30 µg), or ceftazidime (30 µg).
DNA extraction
DNA was isolated from E. coli pure colonies cultured on MacConkey agar by boiling method. Briefly, bacterial isolates were suspended in 400 µL of Tris–EDTA buffer (TE buffer), vortex mixed, and boiled at 95°C for 10 min. The supernatant was separated by centrifugation at 12,000 rpm for 5 min. DNA was precipitated by adding 1/10 volume of 3 M NaOAc, pH 5.2, and 2–3 volumes of chilled absolute ethanol and centrifuged at 12,000 rpm for 15 min. The bacterial DNA was washed with 70% ethanol, dried, and dissolved in 50 µL of TE buffer. DNA was stored at − 20 °C for subsequent amplification.
PCR amplification and DNA Sequencing
All E. coli clinical isolates were investigated for the presence of ESBL genes (blaCTX-M, blaOXA, blaSHV, and blaTEM). Further, the genes were sequenced to identify the variants. The primer sequences and amplicon sizes of each ESBL gene are described in Supplementary Table S2. The PCR reaction was carried out in a total volume of 25 µL comprising 50 ng of DNA, 1.5 mM MgCl2, 0.2 mM dNTPs, 0.4 µM of each primer, 1 × Standard Taq reaction buffer, and 1.25 U Taq polymerase (New England Biolabs, UK). The amplification of each ESBL gene were performed as previously described54,55,56,57. Positive control of blaTEM, blaCTX-M, blaOXA, and blaSHV were derived from E. coli EC137 and K.pneumoniae KP125, respectively (kindly provided by Prof. Visanu Thamlikitkul, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, Thailand). All PCR products were examined using 2% agarose gel electrophoresis. The amplified fragments were sequenced (Bioneer Corporation, South Korea) and aligned with the GenBank database using the BLASTn program (https://blast.ncbi.nlm.nih.gov/) and Clustal Omega (https://www.ebi.ac.uk/Tools/msa/ ustalo/), respectively.
Multiplex RPA reaction for bla CTX-M, bla OXA, and bla SHV genes
The RPA primers for blaCTX-M, blaOXA, and blaSHV, genes described previously were used (Supplementary Table S3). The multiplex RPA reaction was performed using the TwistAmp Basic reaction kit (TwistDx, UK). The RPA master mix contained 0.2 µM of blaCTX-M primers, 0.1 µM each of blaOXA and blaSHV primers, 29.5 µL rehydration buffer, 50 ng of DNA template, and sterile distilled water added to obtain a final volume of 47.5 µL. The reaction was vortex mixed, and then transferred to a freeze-dried tube, and finally mixed with 2.5 µL of 280 mM MgOAc. The reaction was incubated at 37 °C for 25 min. To explore the optimum conditions of the multiplex RPA reactions for blaCTX-M, blaOXA, and blaSHV, the following conditions were tried: (a) two sets of CTX-M, OXA, and SHV primer concentrations—0.2, 0.1, 0.1 and 0.2, 0.05, 0.05 µM; (b) three reaction temperatures—37 °C, 39 °C, and 41 °C; and (c) five incubation durations—10, 15, 20, 25, and 30 min. The RPA amplicons were purified using a GeneJet PCR purification kit (Thermo Fisher Scientific Inc., USA) and detected using 2% agarose gel electrophoresis.
tHDA reaction for bla TEM gene
The blaTEM gene of E. coli (accession no. MG515250.1) was used for primer design using Primer3plus program (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). The sequences were as follows: forward primer, 5′-TGAGTGATAACACTGCGGCCAACTTAC-3′; reverse primer, 5′-CCCTACGATCAAGGCGAGTTACATGAT-3′. The tHDA reaction was performed in a total volume of 50 µL using IsoAmp® II Universal tHDA Kit (New England Biolabs, Inc., USA). The reaction mixer contained 1X Annealing buffer II, 4 mM MgSO4, 40 mM NaCl, 3.5 µL IsoAmp® dNTP Solution, 0.075 µM of each primer, and 3.5 µL IsoAmp® Enzyme Mix. The reaction was overlaid with mineral oil and incubated at 67 °C for 75 min. The tHDA condition was optimized by varying primer concentration (0.025, 0.050, and 0.075 µM), temperature (63 °C, 65 °C, 67 °C, 69 °C, 71 °C) and incubation time (30, 45, 60, 75, and 90 min). The tHDA amplicon of 111 bp was detected using 2% agarose gel electrophoresis.
Analytical LOD and specificity of multiplex RPA and tHDA assays
The template DNA from E. coli EC120, K. pneumoniae KP125, K. pneumoniae ATCC 700,603, E. coli EC137 harboring blaCTX-M, blaOXA, blaSHV, and blaTEM genes, respectively, were diluted to 100, 50, 5, 0.5, 0.05 ng/µL concentrations. The LOD was assessed as the lowest DNA concentration required for amplicon production using the optimized (in this study) multiplex RPA and tHDA assays. The specificity was also evaluated using DNA isolated from E. coli ATCC 25,922, K. pneumoniae ATCC 700,603, Proteus mirabilis ATCC 25,933, Pseudomonas aeruginosa ATCC 27,853, Acinetobacter baumannii ATCC 19,606, Staphylococcus aureus ATCC 25,923, and Enterococcus faecalis ATCC 29,212.
Validation of multiplex RPA and tHDA assays
All 144 clinical isolates were examined for the presence of blaCTX-M, blaOXA, and blaSHV genes (multiplex RPA assay) and blaTEM gene (tHDA assay). The sensitivity, specificity, positive and negative predictive values were calculated by comparing with nucleotide sequencing results obtained using MEDCALC® easy-to-use statistical software (https://www.medcalc.org/calc/diagnostic_test.php).
Detection of virulence genes
Eight virulence genes, viz. traT, ompT, iutA, fimH, hlyA, iha, papC, and cvaC, which are commonly associated with ExPEC were characterized by multiplex PCR methods. Three positive control genes (fimH, hlyA, and iutA) were synthesized and commercially cloned in GeneArt vectors (Invitrogen, USA). The recombinant plasmids were then transformed into a DH5α competent cell using Subcloning Efficiency™ DH5α Competent Cell (Invitrogen, USA). The primers for these three virulence genes (designed here) and five primers sets for traT, ompT, cvaC, iha, and papC genes (described previously) are tabulated in Supplementary Table S4. The first pool consists of fimH, hlyA, and iutA, while the second pool comprises cvaC, iha, traT, ompT, and papC. A total 25 µL reaction comprising 1 × Standard Taq reaction buffer and 1.5 U Taq polymerase (New England Biolabs, UK), 0.2 µM of each set of primers (forward and reverse), 0.2 mM dNTPs, ~ 100 ng DNA template and ultra-pure water were used. The optimized PCR condition for virulence genes was: initial denaturation at 94 ℃ for 15 min followed by 29 cycles at 94 ℃ for 1 min, annealing at 56 ℃ for 1 min, extension at 72 ℃ for 1 min, and post extension at 72 ℃ for 10 min. All amplicons were analyzed using 2% agarose gel electrophoresis.
Phylogroup analysis
Quadruplex PCR was performed by targeting arpA, chuA, yjaA, and TspE4.C2 (Clermont et al.). An additional PCR was carried out as described previously20 using specific primers for groups C, E, and cryptic Clades. Previously published primers and the amplicon size are illustrated (Supplementary Table S5). The phylogroups B1, B2, and F were assigned based on quadruplex results, while specific primers were used to differentiate phylogroups C, E, and other cryptic Clades from A, D, and Clade I20. A total reaction of 25 µL consisting of 2.5 µL 10 × buffer, 0.2 mM of each dNTPs, 1 µL DNA template (100 ng), 1.5 U Taq polymerase (New England Biolabs, UK), 0.2 µM primers except for internal control (trpBA = 0.12 µM) and Milli Q water was used. We followed the same PCR conditions as described by Clermont et al. 201320. PCR products were analyzed using 2% agarose gel electrophoresis.
Statistical analysis
The association of VF genes and phylogroups was compared between ESBL and non-ESBL-ExPEC isolates using Mann–Whitney U test. The distribution of VF genes across phylogroups was tested using Friedman’s and Kruskal–Wallis tests. P value < 0.05 was considered statistically significant.
Ethical approval
The study protocol was approved by Research Committee of National Cancer Institute (certificate number 020/2562).
Data availability
The datasets generated during the current study are available in the genbank NCBI repository, with the accession number including OP999005-OP999011. These datasets were derived from the following public domain resources: https://www.ncbi.nlm.nih.gov/nuccore/OP999005, https://www.ncbi.nlm.nih.gov/nuccore/OP999006, https://www.ncbi.nlm.nih.gov/nuccore/OP999007, https://www.ncbi.nlm.nih.gov/nuccore/OP999008, https://www.ncbi.nlm.nih.gov/nuccore/OP999009, https://www.ncbi.nlm.nih.gov/nuccore/OP999010, https://www.ncbi.nlm.nih.gov/nuccore/OP999011.
References
World Health Organization. WHO publishes list of bacteria for which new antibiotics are urgently needed. https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (2017).
Umadevi, S. et al. Prevalence and antimicrobial susceptibility pattern of ESBL producing gram negative bacilli. J. Clin. Diagn. Res. 5, 236–239 (2011).
Khanfar, H. S., Bindayna, K. M., Senok, A. C. & Botta, G. A. Extended spectrum beta-lactamases (ESBL) in Escherichia coli and Klebsiella pneumoniae: Trends in the hospital and community settings. J. Infect. Dev. Ctries. 3, 295–299. https://doi.org/10.3855/jidc.127 (2009).
Riley, L. W. Distinguishing pathovars from nonpathovars: Escherichia coli. Microbiol. Spectr. 8, 4–8. https://doi.org/10.1128/microbiolspec.AME-0014-2020 (2020).
Shaik, S. et al. Comparative genomic analysis of globally dominant ST131 clone with other epidemiologically successful extraintestinal pathogenic Escherichia coli (ExPEC) lineages. MBio 8, e01596-e1617. https://doi.org/10.1128/mBio.01596-17 (2017).
Castanheira, M., Simner, P. J. & Bradford, P. A. Extended-spectrum β-lactamases: An update on their characteristics, epidemiology and detection. JAC-antimicrob. Resist. 3, dlab092. https://doi.org/10.1093/jacamr/dlab092 (2021).
Ewers, C. et al. Genomic diversity and virulence potential of ESBL-and AmpC-β-lactamase-producing Escherichia coli strains from healthy food animals across Europe. Front. Microbiol. 12, 626774. https://doi.org/10.3389/fmicb.2021.626774 (2021).
Qin, X. et al. Comparison of adhesin genes and antimicrobial susceptibilities between uropathogenic and intestinal commensal Escherichia coli strains. PLoS One 8, e61169. https://doi.org/10.1371/journal.pone.0061169 (2013).
Armand-Lefèvre, L. et al. Dynamics of extended-spectrum beta-lactamase-producing Enterobacterales colonization in long-term carriers following travel abroad. Microb. Genomics 7, 000576. https://doi.org/10.1099/mgen.0.000576 (2021).
Diallo, O. O. et al. Antibiotic resistance surveillance systems: A review. J. Glob. Antimicrob. Resist. 23, 430–438. https://doi.org/10.1016/j.jgar.2020.10.009 (2020).
CLSI. Performance Standards for Antimicrobial Susceptibility testings; Clinical and Laboratory Standards Institute: Twenty-eight informational supplement, M100-S28. (2018).
Su, M., Satola, S. W. & Read, T. D. Genome-based prediction of bacterial antibiotic resistance. J. Clin. Microbiol. 57, e01405-e1418. https://doi.org/10.1128/JCM.01405-18 (2019).
Vincent, M., Xu, Y. & Kong, H. Helicase-dependent isothermal DNA amplification. EMBO Rep. 5, 795–800. https://doi.org/10.1038/sj.embor.7400200 (2004).
Kanokudom, S. et al. Rapid detection of extended spectrum β-lactamase producing Escherichia coli isolated from fresh pork meat and pig cecum samples using multiplex recombinase polymerase amplification and lateral flow strip analysis. PLoS One 16, e0248536. https://doi.org/10.1371/journal.pone.0248536 (2021).
Piepenburg, O. TwistAmp DNA Amplification Kits-Combined Instruction Manual. Twist. Ltd, Cambridge, UK (2013).
Piepenburg, O., Williams, C. H., Stemple, D. L. & Armes, N. A. DNA detection using recombination proteins. PLoS Biol. 4, e204. https://doi.org/10.1371/journal.pbio.0040204 (2006).
An, L. et al. Characterization of a thermostable UvrD helicase and its participation in helicase-dependent amplification. J. Biol. Chem. 280, 28952–28958. https://doi.org/10.1074/jbc.M503096200 (2005).
Sarowska, J. et al. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 11, 1–16. https://doi.org/10.1186/s13099-019-0290-0 (2019).
Cyoia, P. S. et al. Presence of virulence genes and pathogenicity islands in extraintestinal pathogenic Escherichia coli isolates from Brazil. J. Infect. Dev. Ctries. 9, 1068–1075. https://doi.org/10.3855/jidc.6683 (2015).
Clermont, O., Christenson, J. K., Denamur, E. & Gordon, D. M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 5, 58–65. https://doi.org/10.1111/1758-2229.12019 (2013).
Milenkov, M. et al. Prevalence, risk factors, and genetic characterization of extended-spectrum beta-lactamase Escherichia coli isolated from healthy pregnant women in Madagascar. Front. Microbiol. https://doi.org/10.3389/fmicb.2021.786146 (2021).
Allami, M., Bahreini, M. & Sharifmoghadam, M. R. Antibiotic resistance, phylogenetic typing, and virulence genes profile analysis of uropathogenic Escherichia coli isolated from patients in southern Iraq. J. Appl. Genet. 63, 401–412. https://doi.org/10.1007/s13353-022-00683-2 (2022).
Daga, A. P. et al. Escherichia coli bloodstream infections in patients at a university hospital: Virulence factors and clinical characteristics. Front. Cell. Infect. Microbiol. 9, 191. https://doi.org/10.3389/fcimb.2019.00191 (2019).
Bonacorsi, S. & Bingen, E. Molecular epidemiology of Escherichia coli causing neonatal meningitis. Int. J. Med. Microbiol. 295, 373–381. https://doi.org/10.1016/j.ijmm.2005.07.011 (2005).
Wijetunge, D. S. S. et al. Characterizing the pathotype of neonatal meningitis causing Escherichia coli (NMEC). BMC Microbiol. 15, 1–15. https://doi.org/10.1186/s12866-015-0547-9 (2015).
Riley, L. W. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 20, 380–390. https://doi.org/10.1111/1469-0691.12646 (2014).
Kitamoto, S., Nagao-Kitamoto, H., Kuffa, P. & Kamada, N. Regulation of virulence: The rise and fall of gastrointestinal pathogens. J. Gastroenterol. 51, 195–205. https://doi.org/10.1007/s00535-015-1141-5 (2016).
Thacker, N. et al. Epidemiology of blood stream infections in pediatric patients at a tertiary care cancer centre. Indian J. Cancer 51, 438. https://doi.org/10.4103/0019-509X.175311 (2014).
Tohamy, S. T., Aboshanab, K. M., El-Mahallawy, H. A., El-Ansary, M. R. & Afifi, S. S. Prevalence of multidrug-resistant Gram-negative pathogens isolated from febrile neutropenic cancer patients with bloodstream infections in Egypt and new synergistic antibiotic combinations. Infect. Drug Resist. 11, 791. https://doi.org/10.2147/IDR.SI63293 (2018).
Bubpamala, J. et al. Phenotypic and genotypic characterizations of extended-spectrum beta-lactamase-producing Escherichia coli in Thailand. Infect. Drug Resist. 11, 2151. https://doi.org/10.2147/IDR.S174506 (2018).
Muller, A. et al. Hospital-diagnosed infections with Escherichia coli clonal group ST131 are mostly acquired in the community. Sci. Rep. 11, 1–6. https://doi.org/10.1038/s41598-021-85116-6 (2021).
Choi, H. J. et al. Characteristics of Escherichia coli urine isolates and risk factors for secondary bloodstream infections in patients with urinary tract infections. Microbiol. Spectr. 10, e01660-e1722. https://doi.org/10.1128/spectrum.01660-22 (2022).
Hossain, M. et al. Genotype–phenotype correlation of β-lactamase-producing uropathogenic Escherichia coli (UPEC) strains from Bangladesh. Sci. Rep. 10, 1–13. https://doi.org/10.1038/s41598-020-71213-5 (2020).
Hassuna, N. A., Khairalla, A. S., Farahat, E. M., Hammad, A. M. & Abdel-Fattah, M. Molecular characterization of extended-spectrum β lactamase-producing E. coli recovered from community-acquired urinary tract infections in Upper Egypt. Sci. Rep. 10, 1–8. https://doi.org/10.1038/s41598-020-59772-z (2020).
Seenama, C., Thamlikitkul, V. & Ratthawongjirakul, P. Multilocus sequence typing and blaESBL characterization of extended-spectrum beta-lactamase-producing Escherichia coli isolated from healthy humans and swine in Northern Thailand. Infect. Drug Resist. 12, 2201. https://doi.org/10.2147/IDR.S209545 (2019).
Higgins, O. et al. Portable differential detection of CTX-M ESBL gene variants, bla CTX-M-1 and bla CTX-M-15, from Escherichia coli isolates and animal fecal samples using loop-primer endonuclease cleavage loop-mediated isothermal amplification. Microbiol. Spectr. 11, e03316-e3322. https://doi.org/10.1128/spectrum.03316-22 (2023).
Wang, Z. et al. Prevalence of extended-spectrum β-lactamase-resistant genes in Escherichia coli isolates from central china during 2016–2019. Animals 12, 3191. https://doi.org/10.3390/ani12223191 (2022).
Pumford, E. A. et al. Developments in integrating nucleic acid isothermal amplification and detection systems for point-of-care diagnostics. Biosens. Bioelectron. 170, 112674. https://doi.org/10.1016/j.bios.2020.112674 (2020).
Lobato, I. M. & O’Sullivan, C. K. Recombinase polymerase amplification: Basics, applications and recent advances. Trac Trends Anal. Chem. 98, 19–35. https://doi.org/10.1016/j.trac.2017.10.015 (2018).
Piepenburg, O., Williams, C. H., Armes, N. A. & Stemple, D. L. Recombinase polymerase amplification. TwistDix Inc. U.S. Patent 7,666,598. https://patents.google.com/patent/US7666598B2/en (2010).
Mayboroda, O., Katakis, I. & O’Sullivan, C. K. Multiplexed isothermal nucleic acid amplification. Anal. Biochem. 545, 20–30. https://doi.org/10.1016/j.ab.2018.01.005 (2018).
Babu, B. et al. A rapid assay for detection of Rose rosette virus using reverse transcription-recombinase polymerase amplification using multiple gene targets. J. Virol. Methods 240, 78–84. https://doi.org/10.1016/j.jviromet.2016.11.014 (2017).
Motohashi, K. Development of highly sensitive and low-cost DNA agarose gel electrophoresis detection systems, and evaluation of non-mutagenic and loading dye-type DNA-staining reagents. PLoS One 14, e0222209. https://doi.org/10.1371/journal.pone.0222209 (2019).
Li, J., Macdonald, J. & von Stetten, F. Correction: Review: A comprehensive summary of a decade development of the recombinase polymerase amplification. Analyst 145, 1950–1960. https://doi.org/10.1039/C9AN90127B (2020).
Barreda-García, S., Miranda-Castro, R., de-Los-Santos-Álvarez, N., Miranda-Ordieres, A. J. & Lobo-Castañón, M. J. Helicase-dependent isothermal amplification: A novel tool in the development of molecular-based analytical systems for rapid pathogen detection. Anal. Bioanal. Chem. 410, 679–693. https://doi.org/10.1007/s00216-017-0620-3 (2018).
Neamati, F., Firoozeh, F., Saffari, M. & Zibaei, M. Virulence genes and antimicrobial resistance pattern in uropathogenic Escherichia coli isolated from hospitalized patients in Kashan, Iran. Jundishapur J. Microbiol. https://doi.org/10.5812/jjm.17514 (2015).
Lee, M. Y. et al. Dissemination of ST131 and ST393 community-onset, ciprofloxacin-resistant Escherichia coli clones causing urinary tract infections in Korea. J. Infect. 60, 146–153. https://doi.org/10.1016/j.jinf.2009.11.004 (2010).
Zhao, R. et al. Phylogenetic distribution of virulence genes among ESBL-producing uropathogenic Escherichia coli isolated from long-term hospitalized patients. J. Clin. Diagn. Res. JCDR 9, DC01. https://doi.org/10.7860/JCDR/2015/13234.6157 (2015).
Bortolami, A. et al. Diversity, virulence, and clinical significance of extended-spectrum β-lactamase-and pAmpC-producing Escherichia coli from companion animals. Front. Microbiol. 10, 1260. https://doi.org/10.3389/fmicb.2019.01260 (2019).
Murase, T. & Ozaki, H. Relationship between phylogenetic groups of Escherichia coli and Pathogenicity among Isolates from chickens with Colibacillosis and healthy chickens. Poult. Sci. 101, 102007. https://doi.org/10.1016/j.psj.2022.102007 (2022).
Vangchhia, B. et al. Phylogenetic diversity, antimicrobial susceptibility and virulence characteristics of phylogroup F Escherichia coli in Australia. Microbiology 162, 1904–1912. https://doi.org/10.1099/mic.0.000367 (2016).
Zhuge, X. et al. Chicken-source Escherichia coli within phylogroup F shares virulence genotypes and is closely related to extraintestinal pathogenic E. coli causing human infections. Transbound. Emerg. Dis. 68, 880–895. https://doi.org/10.1111/tbed.13755 (2021).
Buderer, N. M. F. Statistical methodology: I. Incorporating the prevalence of disease into the sample size calculation for sensitivity and specificity. Acad. Emerg. Med. 3, 895–900. https://doi.org/10.1111/j.1553-2712.1996.tb03538.x.
Stürenburg, E., Lang, M., Horstkotte, M. A., Laufs, R. & Mack, D. Evaluation of the MicroScan ESBL plus confirmation panel for detection of extended-spectrum β-lactamases in clinical isolates of oxyimino-cephalosporin-resistant Gram-negative bacteria. J. Antimicrob. Chemother. 54, 870–875. https://doi.org/10.1093/jac/dkh449 (2004).
Hasman, H., Mevius, D., Veldman, K., Olesen, I. & Aarestrup, F. M. β-Lactamases among extended-spectrum β-lactamase (ESBL)-resistant Salmonella from poultry, poultry products and human patients in The Netherlands. J. Antimicrob. Chemother. 56, 115–121. https://doi.org/10.1093/jac/dki190 (2005).
Costa, D. et al. Detection of Escherichia coli harbouring extended-spectrum β-lactamases of the CTX-M, TEM and SHV classes in faecal samples of wild animals in Portugal. J. Antimicrob. Chemother. 58, 1311–1312. https://doi.org/10.1093/jac/dkl415 (2006).
Kim, J., Kwon, Y., Pai, H., Kim, J.-W. & Cho, D.-T. Survey of Klebsiella pneumoniae strains producing extended-spectrum β-lactamases: Prevalence of SHV-12 and SHV-2a in Korea. J. Clin. Microbiol. 36, 1446–1449. https://doi.org/10.1128/JCM.36.5.1446-1449.1998 (1998).
Acknowledgements
We thank to Second Century Fund (C2F), Postdoctoral Fellowship, Chulalongkorn University, Bangkok, 10330, Thailand.
Funding
This research project was partially supported by Research Unit of Innovative Diagnosis of Antimicrobial Resistance, Ratchadapisek Sompoch Endowment fund, and the Second Century Fund (C2F), Chulalongkorn University. The funders had no role in study design, data collection, and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
This study was conceived and designed by N.C. Experimental work and data analysis were done by N.U. and T.A., and the drafted manuscript was done by N. Some reagents were supported by Y.A. Funding acquisition, statistical analysis, editing, and finalizing the manuscript were done by N.C. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ullah, N., Assawakongkarat, T., Akeda, Y. et al. Detection of Extended-spectrum β-lactamase-producing Escherichia coli isolates by isothermal amplification and association of their virulence genes and phylogroups with extraintestinal infection. Sci Rep 13, 12022 (2023). https://doi.org/10.1038/s41598-023-39228-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-39228-w
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
