
Abstract
Siberian apricot (Prunus sibirica L.) is a woody tree species of ecological, economic, and social importance. To evaluate the genetic diversity, differentiation, and structure of P. sibirica, we analyzed 176 individuals from 10 natural populations using 14 microsatellite markers. These markers generated 194 alleles in total. The mean number of alleles (13.8571) was higher than the mean number of effective alleles (6.4822). The average expected heterozygosity (0.8292) was higher than the average observed heterozygosity (0.3178). Shannon information index and polymorphism information content were separately 2.0610 and 0.8093, demonstrating the rich genetic diversity of P. sibirica. Analysis of molecular variance revealed that 85% of the genetic variation occurred within populations, with only 15% among them. The genetic differentiation coefficient and gene flow were separately 0.151 and 1.401, indicating a high degree of genetic differentiation. Clustering results showed that a genetic distance coefficient of 0.6 divided the 10 natural populations into two subgroups (subgroups A and B). STRUCTURE and principal coordinate analysis divided the 176 individuals into two subgroups (clusters 1 and 2). Mantel tests revealed that genetic distance was correlated with geographical distance and elevation differences. These findings can contribute to the effective conservation and management of P. sibirica resources.
Similar content being viewed by others


Introduction
Siberian apricot (Prunus sibirica L.) is a deciduous perennial woody tree species of the genus Prunus in the Rosaceae subfamily Prunoideae1. Given its white or pinkish flowers in spring, ripe yellow fruit in summer, and highly diverse leaf colors in autumn, the tree is considered to have particular ornamental value2,3. As a pioneer tree species in the restoration of vegetation, P. sibirica can tolerate cold, drought, and barren land and, thus, has high ecological value1,4. In addition, the seeds of this species contain amygdalin and high-quality oils, which have significant economic value5,6. Consequently, the government in north China promotes this tree as an economic forest species in semi-arid and arid areas, and breeding improved varieties with excellent characteristics is desirable.
As a source of genetic variation and potentially valuable agronomic traits, wild genetic resources can make a contribution to crop improvement7. P. sibirica is native to northern and northeastern China and has distribution in its neighboring countries of Russia and Mongolia8,9. Increased study and introduction of these wild germplasm resources will facilitate breed new varieties with improved characteristics. Such improvement is based on the germplasm collection, assessment, and conservation of genetic resources10. To date, reports on P. sibirica germplasm resources have been limited to parts of northern and northeastern China11,12,13,14,15,16. However, those populations located in less favorable and more peripheral habitats are also significant with unique adaptive genetic diversity17. Our incomplete understanding of these populations could limit the efficient utilization of P. sibirica genetic resources10,18. Furthermore, human interference is increasingly threatening the survival of these natural populations13,19, and the resulting habitat fragmentation will reduce the occurrence of genetic variation and the evolutionary potential of P. sibirica20. Given this scenario, it is urgent to devise and implement protection strategies designed for the sustainable protection of the wild resources of P. sibirica.
Assessing the genetic diversity of P. sibirica genetic resources is particularly important for breeding and conservation programs21. In the development of superior varieties, genetic diversity can reflect the improvement potential of species in the breeding process22. Moreover, genetic diversity is fundamental to sustaining the adaptive capacities of species, the loss of which can potentially compromise the survival potential of affected populations or species23. The distribution pattern of genetic diversity within a species is an essential reference for designing conservation measures24. In this respect, genetic markers can reflect differences in DNA sequences and are commonly applied to study the amount and distribution of genetic variation25. Compared with morphological and biochemical markers, molecular markers can identify variation at specific loci and assist in quantifying genetic diversity levels22. Among the various kinds of molecular markers, microsatellite markers have the favorable characteristics of co-dominant inheritance, high polymorphism, good repeatability, and regular distribution26,27. Consequently, microsatellite markers are widely applied to detect the genetic diversity of plants28,29,30,31, particularly in genus Prunus21,32,33,34.
In view of these considerations, we obtained genetic materials from 176 samples in 10 natural P. sibirica populations and applied 14 pairs of microsatellite markers to assess the level of genetic diversity. The nuclear of our research were to (1) assess the genetic diversity, differentiation, and structure of P. sibirica and (2) develop strategies to protect P. sibirica germplasm resources. The results of our research will contribute to the effective conservation and management of the wild P. sibirica resources.
Materials and methods
Plant materials
A total of 176 samples collected from 10 natural populations of P. sibirica were used in this study. Location information for 176 samples (Table S1) was obtained using GPS. With the exception of population R, distributed in the Zabaykalsky Krai region of Russia, all other populations are distributed in northeast China. The sampled populations cover a broad geographical span and ecological spectrum, with longitudes ranging from N 40°01′ to 53°05′, latitudes from 115°23′ to 123°01′, and elevations from 222 to 1292 m. We collected the fresh young leaves of trees in the field, froze them in liquid nitrogen, and stored them at − 80 °C until use.
DNA extraction and amplification
We extracted the total genomic DNA from 176 samples individually using the Plant Genome DNA Extraction Kit (TianGen, Beijing, China) and purified them by 1% agarose gel electrophoresis. NanoDrop2000 spectrophotometry (Thermo Fisher Scientific, Waltham, MA, USA) was used to test the concentration and purity of the extracted DNA, and the high quality DNA was stored at − 20 °C until use.
In our previous studies, we developed microsatellite markers for P. sibirica12,35. We used 14 pairs of microsatellite markers (Table 1) to amplify the extracted genomic DNA. PCR amplifications were performed in 20-µL mixtures comprising 20 ng template DNA, 0.25 mmol/L dNTPs, 2 mmol/L Mg2+, 1 U Taq polymerase, 10 × PCR buffer, and 0.15 µmol/L primers. Amplification was performed by the following parameters: (1) initiated with a 105 °C hot lid, (2) initially denatured at 94 °C for 5 min, (3) 35 cycles of 94 °C for 30 s, 55 °C for 30 s (annealing temperature depending on the primers used), and 72 °C for 30 s, (4) finally extended at 72 °C for 5 min, and (5) stored at 4 °C. The amplified products were visualized by non-denaturing polyacrylamide gel electrophoresis at 220 V for 90 min. After rinsing, silver staining, and development, the gel was imaged using a Bio-Rad gel detection system (Bio-Rad, Hercules, CA, USA).
Data analysis
We used Image Lab v 6.1 software to read the gel bands and obtained the raw genotype data after manual correction. The data were converted to different formats for further analysis using GenAlEx v 6.50236. POPGENE v 1.32 software37 was used to calculate genetic diversity parameters, including the number of alleles (Na), the number of effective alleles (Ne), observed heterozygosity (Ho), expected heterozygosity (He), and Shannon’s information index (I)38,39,40. Polymorphic information content (PIC) was determined using Cervus v 3.0 software41.
The results of analysis of molecular variance (AMOVA), pairwise FST analysis among populations, and principal coordinate analysis (PCoA) were computed by GenAlEx v 6.50236,42,43. In addition, we used GenAlEx v 6.502 to mantel test the correlations about genetic distance with geographical distance and genetic distance with elevation differences37.
Genetic structure was analyzed based on Bayesian clustering using STRUCTURE v 2.3.444,45. This software was performed by the following parameters: (1) used the admixture model to perform a Markov chain Monte Carlo (MCMC) simulation, and set at 200,000 and 100,000 corresponded to the number of repetitions and the length of the burn-in period, and (2) set the K value from 1 to 10 with 10 repetitions. The optimal value of K was estimated using Evanno’s ∆K method and Structure Harvester46,47. A geographical distribution diagram with the proportion of subgroups membership was plotted using ArcMap v 10.2.
We calculated genetic similarity coefficients between 176 individuals using the Simple Matching (SM) algorithm and constructed clustering relationship tree graphs using the unweighted pair group method with arithmetic means (UPGMA), both using Ntsys-pc v 2.10s43,48. We used the MEGA v 7 software to calculate the Nei’s genetic distances between the 10 natural populations and constructed the UPGMA clustering relationship tree49,50.
Ethical statement
The plant materials used in this article did not involve disputes. Plant materials were collected in compliance with institutional, national, and international guidelines and legislation. The plant material in our collection is preserved in the National Germplasm Repository of Prunus sibirica in Kazuo County, Liaoning Province, China.
Results
Genetic diversity
The extracted DNA of 176 P. sibirica samples were amplified by the 14 pairs of microsatellite primers, generating a total of 194 alleles (Table 1). The PIC of each locus was higher than 0.5, meaning that all 14 loci were highly polymorphic. The Na per locus ranged from 8 (L62) to 19 (Y65 and X47), with a mean of 13.8571 alleles. The Ne for each locus ranged from 3.2636 (L75) to 9.0865 (P40H), with an average of 6.4822. The Ho varied from 0.0114 (P57H) to 0.6989 (Y5), whereas the He ranged from 0.6936 (L75) to 0.8899 (P40H). And the mean value of Ho (0.3178) was lower than that of the He (0.8292), suggesting a lower level of heterozygosity in P. sibirica. The He value above 0.5 reflected that P. sibirica population was not affected by high-intensity selection and possessed high genetic polymorphism. The I for each locus ranged from 1.5161 (L75) to 2.4459 (P40H), with an average of 2.0610.
The genetic diversity parameters of the 10 P. sibirica populations are shown in Table 2. The Na per population ranged from 3.6429 (R population) to 8.2143 (NZD population), with a mean of 5.8500. The Ne per population ranged from 2.3912 (R population) to 5.3442 (NZW population), with a mean of 3.7703. The I value per population ranged from 0.8769 (R population) to 1.7759 (NZW population), with a mean value of 1.3677. The average of He (0.6590) was higher than that of Ho (0.3183). These comparisons indicated that the NZW population shows the highest genetic diversity within the assessed populations, whereas the R population shows the lowest.
Genetic differentiation
AMOVA showed that 15% of the genetic variation occurred among the 10 P. sibirica populations and 85% within them (Table 3). The genetic differentiation among populations was low, moderate, and high when the FST value was between 0.00 and 0.05, 0.05 and 0.15, and 0.15 and 0.25, respectively, greater than 0.25 indicating a large genetic differentiation. The genetic differentiation of P. sibirica detected by FST was high (FST = 0.151 > 0.15, p < 0.01), with the gene flow of 1.401 (Nm > 1), which indicated a moderate interchange of genes among these populations. Paired FST values (Table S2) indicate the degree of genetic differentiation between populations, with the highest differentiated being the R and HL populations (FST = 0.415 > 0.25) and the lowest differentiated being the LB and LC populations (FST = 0.002 < 0.05).
A UPGMA dendrogram generated based on Nei’s genetic distances of the 10 P. sibirica populations (Fig. S1) revealed that at a coefficient of 0.6, the P. sibirica populations could be classified into subgroups A and B, with subgroup B comprising populations HZ, HL, and HW, and the remaining populations grouped in subgroup A. The pairwise FST values (Table S2) within subgroup A ranged from 0.002 (LB and LC) to 0.287 (R and LC) and from 0.002 to 0.141 (NA and NZW) without the R population. The pairwise FST values within subgroup B ranged from 0.011 (HL and HW) to 0.065 (HW and HZ). With the exception of population R, which showed substantial genetic differentiation from the others, we found a moderate and low degree of genetic differentiation within subgroups (FST < 0.15).
Genetic structure
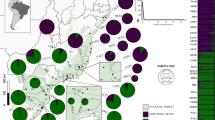
The STRUCTURE results revealed that ∆K reached a maximum value when K = 2 (Fig. 1), thereby indicating that the 176 P. sibirica samples can be organized into two subgroups (clusters 1 and 2). The subgroup membership probabilities of 176 individuals from the 10 populations are shown in Fig. 2. Moreover, the proportion of subgroups membership of 10 nature populations was shown in Fig. 3. We detected significant differences in the geographical distribution of two subgroup members, dividing the natural populations into eastern and western subgroups that separately correspond to subgroups A and B. Three populations NZD, NZW, and LB showed a relatively mixed, suggesting possible gene flow between subgroups.

Relationship between the rational cluster K and estimated value ΔK.

A genetic structure analysis for 176 Prunus sibirica individuals based on Bayesian simulation (K = 2). Red for cluster 1, and green for cluster 2. The ordinate is the Q value, the abscissa is the code of the individual and their natural population. (1), (2), (3), (4), (5), (6), (7), (8), (9), and (10) represent LK, LC, LB, NA, NZW, NZD, R, HL, HW, and HZ, respectively.

The proportions of subgroup memberships in each of the 10 Prunus sibirica populations.
With the increase of K value (K = 3, 4, and 5), clusters 1 was separated into smaller subgroups based on the geographical distribution (Figs. S2–S4). According to the genetic structure analysis, the R population showed a relatively pure genetic component, and the other populations in clusters 1 contain admixed genetic components (Figs. S6–S8). When K = 6, clusters 2 was separated into two subgroups, showing a certain mixed state (Figs. S5, S9).
Similarly, the findings of PCoA (Fig. S10) showed that the 176 P. sibirica samples could be divided into two subgroups in complete consistency with the structure result. Furthermore, the constructed UPGMA tree clearly shows that P. sibirica individuals can be divided into two subgroups (P1 and P2) under a coefficient of 0.83 (Fig. 4). At a genetic similarity coefficient of 0.84, the P2 subgroup can be subdivided into P21 and P22, the latter of which was completely consistent with cluster 2 of the structure result. Mantel test revealed the genetic distance of 176 P. sibirica individuals correlated with geographical distance (r = 0.077, p < 0.01) (Fig. S11) and elevational distribution (r = 0.112, p < 0.01) (Fig. S12), respectively.

Unweighted pair group method with arithmetic means (UPGMA) cluster analysis of 176 Prunus sibirica individuals based on Simple Match (SM) similarity coefficient.
Discussion
Genetic diversity
Genetic diversity reflects the evolutionary characteristics of species and their adaptation to the environment, which is typically determined based on the data of continuous traits or discrete allelic states51. In this study, we obtained discrete allelic data for 176 P. sibirica accessions from 10 natural populations based on analysis using 14 microsatellite markers. We evaluated the genetic diversity of P. sibirica in terms of allelic richness, heterozygosity, and allelic diversity. Compared with previous studies11,12,13,14,15,17, the present study significantly extends the sampling area of P. sibirica to the north (Zabaykalsky Krai), with a corresponding expansion of climate type from intermediary and warm to cold temperate zones. The mean genetic parameters of each locus detected in our research (Na = 13.857, He = 0.829, and I = 2.061) is comparable to that reported by Wang et al.15 (Na = 19.323, He = 0.774, and I = 2.062). These findings indicate that within the genus Prunus, P. sibirica has a slightly higher level of genetic diversity than apricot (Prunus armeniaca L.) (Na = 24.360, He = 0.732, and I = 1.837)21, and a significantly higher diversity compared with sweet cherry (Prunus avium L.) (Na = 9.800 and He = 0.700)52, peach (Prunus persica L.) (Na = 6.410 and He = 0.490)53 and alpine plum (Prunus brigantina Vill.) (Na = 5.040 and He = 0.430)54. Moreover, compared with xylophyta in the Rosaceae family, the diversity is lower than that of apple (Malus × domestica Borkh.) (Na = 23.060 and He = 0.830)55 and higher than that of pear (Pyrus pyrifolia) (Na = 7.120 and He = 0.780)56. The average genetic parameters of the populations detected in our research (Na = 5.850, He = 0.659, and I = 1.368) is higher than that of Prunus mira Koehne (Na = 3.800, He = 0.520, and I = 0.950)57. We speculate that this relatively high level of genetic diversity in P. sibirica is related to their area of distribution. Li et al.13 considered that P. sibirica might accumulate considerable intraspecific genetic variation to adapt to the diverse ecological conditions in its wide range of distribution. Climatic fluctuations over recent millions of years have tended to influence the genetic diversity of species. However, the center of P. sibirica distribution is East Asia, which is generally considered to have been a large-scale refuge during the Pleistocene, characterized by a relatively small range of environmental changes58. We thus speculate that this species may have retained a large proportion of its intraspecific genetic variation. Furthermore, Mehlenbacher et al.1 found that the hybridizations between any two true apricot species were successful, including those among P. sibirica, P. armeniaca, P. mandshurica, and P. mume. Thus, interspecific hybridization may also influence the accumulation of genetic variation in P. sibirica.
In the present study, we found that the natural populations of P. sibirica distributed in different geographical regions differ significantly in terms of genetic diversity, with the highest levels of genetic diversity being detected in the NZW and NZD populations distributed in the northeastern region of Inner Mongolia, followed by the LB, LK, and LC populations in western Liaoning, and then the HW, HZ, and HL populations in northern Hebei and the NA population in central Inner Mongolia, with the lowest diversity detected in R populations in southern Zabaykalsky Krai. The natural habitat of the NZW and NZD populations is located near the Changbai Mountains overlaps with the natural distribution range of the Manchurian apricot (Prunus mandshuria Skv.), and most germplasms are older and less disturbed by human activities, which may account for higher genetic diversity of these populations. Population R is the only population distributed within the cold temperate zone, and although has lower genetic diversity than other assessed populations, it has accumulated unique genotypes during the process of adapting to its marginal habitats. In addition, germplasm resources in western Liaoning have been widely reported12,13,14,15. In the present study, the ranking of the genetic diversity levels in the Liaoning populations (LC < LK < LB) is consistent with the previous findings12,15. Contrastingly, Li et al.13 were of opinion that the genetic diversity was higher in the LC population than in the LK population. However, we found the genetic diversity of the NA population with similar geographical distance was relatively low, which is also consistent with the results presented by Chen et al.12 and Wang et al.15.
Genetic differentiation and genetic structure
Woody plants tend to maintain a larger extent of variation within populations than between populations, particularly in the case of those species with a large geographical range, outcrossing, and seed dispersal mediated mainly by wind or animals59. This is consistent with our present findings for P. sibirica, the genetic variation of which mainly exists within species and populations. We detected a high degree of genetic differentiation at the species level (0.15 < FST < 0.25), and given because the existing gene flow (Nm = 1.401 > 1), there is a resistance to genetic drift, there was no further differentiation43,60. Consequently, the genetic structure was relatively stable. Given the extensive and discontinuous distribution of P. sibirica, we speculate that gene flow among populations is maintained by animal-mediated seed dispersal rather than a combination of pollen transmission and insect- and wind-mediated dispersal, which is also consistent with the views of Li et al.13. In the distribution area surveyed in the present study, rodents hoarded P. sibirica seeds at dispersed locations, and exploited these less often or consumed them only in the absence of alternatives, owing to the amygdalin content61. We speculate that the primary mode of gene flow is the moderate exchange of genes between the buried P. sibirica seeds and local populations through pollination after they have developed to reproductive maturity.
We established that the genetic structure of P. sibirica consists of two subgroups: the eastern and western subgroups, based on the STRUCTURE, clustering, and PCoA results. We found that whereas there was a moderate and low genetic differentiation among populations within the same subgroup (excepting the R population), there was a high differentiation between the populations belonging to different subgroups, among which the R population of the eastern subgroup and the HL population of the western subgroup were characterized by the highest genetic differentiation (FST = 0.415 > 0.25). Based on the common natural hybridization phenomena among plants of the Li subgenus apricot group1 and the distribution of plants of subgenus apricot in each region, we speculate that western subgroup may have undergone an introgression with P. armeniaca, which has thus influenced the genetic structure of the P. sibirica. Liu et al.14 divided P. sibirica germplasm into two subgroups and similarly believed that the northwestern subgroup was largely derived as a consequence of introgression between the northeastern subgroup and P. armeniaca. Mantel test results revealed the significant influence of geographical distance and elevation distribution in contributing to the genetic structure among P. sibirica in the study area (p < 0.001). Furthermore, the area surveyed in the present study falls within a semi-humid region, and given the high degree of genetic differentiation between populations in the western subgroups region lying in the warm temperate zone and those in the eastern subgroup region lying in the middle and cold temperate zone, we speculate that temperature also influences the genetic structure of P. sibirica.
Conservation remarks
As a consequence of frequent human activities, such as fruit and seed picking, grazing, and deforestation, large areas of P. sibirica habitat have been destroyed, and its natural distribution range continues to decline13. To maximize the conservation of P. sibirica genetic resources, we have developed conservation strategies for P. sibirica in the study area depending on our findings. The genetic variation of P. sibirica exists occurs mainly within populations, and thus conservation efforts should focus on maintaining and increasing the genetic diversity level within populations. In this regard, we established that the genetic diversity of the NZW and NZD populations distributed in northern Inner Mongolia was significantly higher than that of other assessed populations, and thus the in situ conservation of these two populations should be given priority22. To protect the existing populations, we recommend the establishment of protected areas or sites to enhance management, which would require cooperation with the local government in charge of forest management. Moreover, to protect the genetic resources of populations with relatively low levels of genetic diversity, we propose to improve the habitats of P. sibirica via complementary silvicultural actions using forest plants from the same or genetically related populations. Furthermore, we have collected genetic materials from the natural population and ex situ preserved in a germplasm conservation base. Lay a solid foundation basis for long-term breeding and genetic resource conservation of P. sibirica.
Conclusions
In this research, we used 14 microsatellite markers to analyze samples collected from 176 individuals in 10 natural populations of Prunus sibirica to assess their genetic diversity, differentiation, and structure. Our findings indicate that P. sibirica has a high level of genetic diversity, particularly within the NZW and NZD populations distributed in northeast Inner Mongolia. At the species level, there is a high degree of genetic differentiation and a relatively stable genetic structure. The genetic structure of P. sibirica can be affected by geographical distance and elevation distribution, with 10 natural populations dividing into two subgroups. This finding will contribute to the conservation research and the rational utilization of P. sibirica.
Data availability
The datasets generated during and analyzed during the current study are available from the corresponding author on reasonable request.
References
Mehlenbacher, S. A., Cociu, V. & Hough, F. L. Apricots (Prunus). Acta Hortic. 3, 65–110 (1991).
Hao, R. et al. A comparative analysis of characteristic floral scent compounds in Prunus mume and related species. Biosci. Biotechnol. Biochem. 78, 1640–1647 (2014).
Xing, X., Hao, P. & Dong, L. Color characteristics of Beijing’s regional woody vegetation based on Natural Color System. Color. Res. Appl. 44, 595–612 (2019).
Wang, Y. et al. The dynamic trend of soil water content in artificial forests on the Loess Plateau, China. Forests 7, 236 (2016).
Do, J., Hwang, J., Seo, H., Woo, W. & Nam, S. Antiasthmatic activity and selective inhibition of type 2 helper T cell response by aqueous extract of semen armeniacae amarum. Immunopharmacol. Immunotoxicol. 28, 213–225 (2006).
Wang, J. et al. Comprehensive evaluation of fuel properties and complex regulation of intracellular transporters for high oil production in developing seeds of Prunus sibirica for woody biodiesel. Biotechnol. Biofuels 12, 6 (2019).
Zamir, D. Improving plant breeding with exotic genetic libraries. Nat. Rev. Genet. 2, 983–989 (2001).
Liu, W. et al. Apricot germplasm resources and their utilization in China. Acta Hortic. 862, 45–49 (2010).
Wang, L. & Yu, H. Biodiesel from Siberian apricot (Prunus sibirica L.) seed kernel oil. Bioresour. Technol. 112, 355–358 (2012).
Liu, F. et al. Genetic diversity and population structure analysis of Dalbergia Odorifera germplasm and development of a core collection using microsatellite markers. Genes 10, 281 (2019).
Buer, H., Rula, S., Wang, Z., Fang, S. & Bai, Y. Analysis of genetic diversity in Prunus sibirica L. in Inner Mongolia using SCoT molecular markers. Genet. Resour. Crop Evol. 69, 1057–1068 (2022).
Chen, J. et al. Genetic diversity of Prunus sibirica L. superior accessions based on the SSR markers developed using restriction-site associated DNA sequencing. Genet. Resour. Crop Evol. 68, 615–628 (2021).
Li, M., Zhao, Z., Miao, X. & Zhou, J. Genetic diversity and population structure of Siberian apricot (Prunus sibirica L.) in China. Int. J. Mol. Sci. 15, 377–400 (2014).
Liu, S. et al. The complex evolutionary history of apricots: Species divergence, gene flow and multiple domestication events. Mol. Ecol. 28, 5299–5314 (2019).
Wang, Z. et al. High-level genetic diversity and complex population structure of Siberian apricot (Prunus sibirica L.) in China as revealed by nuclear SSR markers. PLoS ONE 9, e87381 (2014).
Wang, Z. et al. Phylogeography study of the Siberian apricot (Prunus sibirica L.) in northern china assessed by chloroplast microsatellite and DNA makers. Front. Plant Sci. 8, 1989 (2017).
Phillips, J., Asdal, Å., Brehm, J. M., Rasmussen, M. & Maxted, N. In situ and ex situ diversity analysis of priority crop wild relatives in Norway. Divers. Distrib. 22, 1112–1126 (2016).
Haliloglu, K., Turkoglu, A., Tan, M. & Poczai, P. SSR-based molecular identification and population structure analysis for forage pea (Pisum sativum var. arvense L.) landraces. Genes 13, 1086 (2022).
Li, W. et al. Genetic diversity, population structure, and relationships of apricot (Prunus) based on restriction site-associated DNA sequencing. Hortic. Res. 7, 69 (2020).
Hughes, A. R. & Stachowicz, J. J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proc. Natl. Acad. Sci. U. S. A. 101, 8998–9002 (2004).
Bourguiba, H. et al. Genetic structure of a worldwide germplasm collection of Prunus armeniaca L. reveals three major diffusion routes for varieties coming from the species’ center of origin. Front. Plant Sci. 11, 638 (2020).
White, T. L., Adams, W. T. & Neale, D. B. Forest Genetic (CAB International, 2007).
Pauls, S. U., Nowak, C., Bálint, M. & Pfenninger, M. The impact of global climate change on genetic diversity within populations and species. Mol. Ecol. 22, 925–946 (2013).
Francisco-Ortega, J., Santos-Guerra, A., Kim, S. C. & Crawford, D. J. Plant genetic diversity in the Canary Islands: A conservation perspective. Am. J. Bot. 87, 909–919 (2000).
Sunnucks, P. Efficient genetic markers for population biology. Trends Ecol. Evol. 15, 199–203 (2000).
Hartl, D. L. & Cochrane, B. J. Genetics: Analysis of Genes and Genomes with 8 edn (Jones & Bartlett Learning, 2019).
Sullivan, A. R., Lind, J. F., Mccleary, T. S., Romero-Severson, J. & Gailing, O. Development and characterization of genomic and gene-based microsatellite markers in North American red oak species. Plant Mol. Biol. Rep. 31, 231–239 (2013).
Ahn, J., Lee, J. & Hong, K. Genetic diversity and structure of Pinus densiflora Siebold & Zucc. populations in Republic of Korea based on microsatellite markers. Forests 12, 750 (2021).
Li, L. et al. Comparison of genetic diversity between ancient and common populations of Docynia delavayi (Franch.) Schneid. Gene 829, 146498 (2022).
Stoltz, S. S. & Husband, B. C. High genetic diversity in American chestnut (Castanea dentata) despite a century of decline. Conserv. Genet. 24, 25–39 (2023).
Waqar, Z. et al. Gene flow and genetic structure reveal reduced diversity between generations of a tropical tree, Manilkara multifida Penn., in Atlantic forest fragments. Genes 12, 2025 (2021).
Kwon, S. et al. Microsatellite variations and population structure in an on-farm collection of Japanese apricot (Prunus mume Sieb. et Zucc.). Biochem. Syst. Ecol. 42, 99–112 (2012).
Lacis, G., Rashal, I., Ruisa, S., Trajkovski, V. & Iezzoni, A. F. Assessment of genetic diversity of Latvian and Swedish sweet cherry (Prunus avium L.) genetic resources collections by using SSR (microsatellite) markers. Sci. Hortic. 12, 451–457 (2009).
Pérez, V., Larrañaga, N., Abdallah, D., Wünsch, A. & Hormaza, J. I. Genetic diversity of local peach (Prunus persica) accessions from La Palma Island (Canary Islands, Spain). Agronomy 10, 457 (2020).
Dong, S. et al. The complete chloroplast genome sequence of Prunus sibirica. Mitochondrial DNA B 5, 581–582 (2020).
Peakall, R. & Smouse, P. E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28, 2537–2539 (2012).
Yeh, F., Yang, R. & Boyle, T. Official Site of POPGENE, The User-Friendly Freeware for Population Genetic Analysis. https://sites.ualberta.ca/fyeh/popgene.html (1999).
Kimura, M. & Crow, J. F. The number of alleles that can be maintained in a finite population. Genetics 49, 725–738 (1964).
Lewontin, R. C. The apportionment of human diversity. Evol. Biol. 14, 381–398 (1972).
Nei, M. Analysis of gene diversity in subdivided populations. Proc. Nat. Acad. Sci. 70, 3321–3323 (1973).
Kalinowski, S. T., Taper, M. L. & Marshall, T. C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 16, 1099–1106 (2007).
Balloux, F. & Lugon-Moulin, N. The estimation of population differentiation with microsatellite markers. Mol. Ecol. 11, 155–165 (2002).
Sneath, P. & Sokal, R. Numerical Taxonomy, the Principle And Practice of Numerical Classification (W. H. Freeman and Company, 1973).
Falush, D., Stephens, M. & Pritchard, J. K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 7, 574–578 (2007).
Pritchard, J. K., Stephens, M. & Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 155, 945–959 (2000).
Earl, D. A. & VonHoldt, B. M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 4, 359–361 (2012).
Evanno, G., Regnaut, S. & Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 14, 2611–2620 (2005).
Rohlf, F. J. NTSYS-pc Numerical Taxonomy and Multivariate Analysis System (Applied Biostatistics Inc, 2009).
Kumar, S., Nei, M., Dudley, J. & Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 9, 299–306 (2008).
Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89, 583–590 (1978).
Hughes, A. R., Inouye, B. D., Johnson, M. T., Underwood, N. & Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 11, 609–623 (2008).
Barreneche, T. et al. SSR-based analysis of genetic diversity and structure of sweet cherry (Prunus avium L.) from 19 countries in Europe. Plants 10, 1983 (2021).
Chavez, D. J., Beckman, T. G., Werner, D. J. & Chaparro, J. X. Genetic diversity in peach [Prunus persica (L.) Batsch] at the University of Florida: Past, present and future. Tree Genet. Genomes 10, 1399–1417 (2014).
Liu, S. et al. Genetic diversity and population structure analyses in the Alpine plum (Prunus brigantina Vill.) confirm its affiliation to the Armeniaca section. Tree Genet. Genomes 17, 2 (2021).
Urrestarazu, J. et al. Analysis of the genetic diversity and structure across a wide range of germplasm reveals prominent gene flow in apple at the European level. BMC Plant Biol. 16, 130 (2016).
Xue, L. et al. The southwestern origin and eastward dispersal of pear (Pyrus pyrifolia) in East Asia revealed by comprehensive genetic structure analysis with SSR markers. Tree Genet. Genomes 14, 1–12 (2018).
Bao, W. et al. Genetic diversity and population structure of Prunus mira (Koehne) from the Tibet plateau in China and recommended conservation strategies. PLoS ONE 12, e0188685 (2017).
Liu, K. Quaternary history of the temperate forests of China. Quarter. Sci. Rev. 7, 1–20 (1988).
Hamrick, J. L., Godt, M. J. & Sherman-Broyles, S. L. Factors influencing levels of genetic diversity in woody plant species. New For. 6, 95–124 (1992).
Slatkin, M. Gene flow and the geographic structure of natural populations. Science 236, 787–792 (1987).
Li, D. et al. Effects of season and food on the scatter-hoarding behavior of rodents in temperate forests of Northeast China. Zookeys 1025, 73–89 (2021).
Funding
This study was supported by National Key Research and Development Program of China (SQ2019YFD100071), and Liaoning Province Wild apricot Germplasm Resource Preservation and Breeding National Permanent Scientific Research Base (2020132519). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception. L.W. and Y.S. surveyed and collected the samples. X.W. analyzed the data and wrote the original manuscript. S.D., Q.L., and J.C. wrote reviewed and edited. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, X., Wang, L., Sun, Y. et al. Genetic diversity and conservation of Siberian apricot (Prunus sibirica L.) based on microsatellite markers. Sci Rep 13, 11245 (2023). https://doi.org/10.1038/s41598-023-37993-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-37993-2
This article is cited by
-
Genetic diversity and population structure of wild and cultivated apricots collected from Kyrgyzstan
Genetic Resources and Crop Evolution (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
