
Abstract
Facial mimicry as well as the accurate assessment of one's performance when judging others’ emotional expressions have been suggested to inform successful emotion recognition. Differences in the integration of these two information sources might explain alterations in the perception of others’ emotions in individuals with Social Anxiety Disorder and individuals on the autism spectrum. Using a non-clinical sample (N = 57), we examined the role of social anxiety and autistic traits in the link between facial mimicry, or confidence in one’s performance, and emotion recognition. While participants were presented with videos of spontaneous emotional facial expressions, we measured their facial muscle activity, asked them to label the expressions and indicate their confidence in accurately labelling the expressions. Our results showed that confidence in emotion recognition was lower with higher social anxiety traits even though actual recognition was not related to social anxiety traits. Higher autistic traits, in contrast, were associated with worse recognition, and a weakened link between facial mimicry and performance. Consequently, high social anxiety traits might not affect emotion recognition itself, but the top-down evaluation of own abilities in emotion recognition contexts. High autistic traits, in contrast, may be related to lower integration of sensorimotor simulations, which promote emotion recognition.
Similar content being viewed by others


Introduction
The expression “her face says it all” exemplifies the fundamental contribution of nonverbal signals and cues in the communication of inner states1, 2. According to social-functional approaches, emotional expressions are crucial in guiding social interactions by informing about others’ states, evoking coordinated emotional responses, and incentivizing social behavior3, 4. The accurate identification of an observed emotional expression is a key component in the interpretation of an expresser’s emotional state, yet links between emotion recognition and other processes underlying emotion perception are still not well described. Facial mimicry, that is the mirroring of an observed expression, is one process that has been suggested to promote the recognition of others’ emotions5, 6 (see Facial mimicry paragraph for further details). In contrast to this bottom-up information channel, the top-down assessment of one’s recognition performance, a metacognitive process, might also provide relevant feedback about emotion processing7 (see Metacognition paragraph for further details). Importantly, alterations in the processing of others’ emotions, as well as in mimicry and metacognition8, 9 have been reported for various mental health and neurodevelopmental conditions, such as Social Anxiety Disorder (SAD) and Autism Spectrum Disorder (ASD). Research in both clinical populations has further revealed a link between alterations in processing other's emotional expressions and social interaction difficulties10, 11. The current study examines the putative associations between facial mimicry and confidence in emotion recognition abilities (i.e., a metacognitive judgment) with actual emotion recognition performance, as well as their potential alterations associated with social anxiety and autistic traits.
Mimicry and emotion recognition
When observing an individual expressing an affective state via the face, people tend to automatically mirror the observed facial expression—a phenomenon called facial mimicry12. Distinct changes in activity over two muscle regions, the Zygomaticus Major and the Corrugator Supercilii (for simplicity referred to as “zygomaticus” and “corrugator” hereinafter) have been consistently reported in response to videos of emotional displays: Strongest evidence has been found for an increase in zygomaticus activity when happy facial expressions were viewed, together with a decrease in corrugator activity5, 13,14,15,16,17,18. Enhanced corrugator activity, in contrast, has been linked to the perception of anger displays5, 13, 16,17,18,19 and, less pronounced, for sadness displays5, 17, 18.
Importantly, instead of being only an epiphenomenon, facial mimicry has been proposed to aid emotion recognition5, 20, 21. In line with seminal theories on emotion22, 23, peripheral signals, such as facial expressions, can not only inform the producer about the physiological effects of emotions via interoceptive pathways24. “Facial feedback”6 might also act as an information source when another individual’s expression is automatically mirrored, serving as a sensorimotor simulation of another person’s emotional state25. This view was supported by studies that showed a decline in emotion recognition performance if facial mimicry was voluntarily26 or artificially27 blocked. Yet, recent meta-analyses suggest that the effects of facial feedback on affective judgments, including emotion recognition, are not consistent28, 29. Moreover, facial mimicry does not seem to be a requirement for successful emotion recognition: A study in patients with Möbius syndrome has shown that, despite facial paralysis, these patients could still accurately recognize emotional expressions30.
Metacognition and emotion recognition
Metacognition describes the monitoring of one’s own cognitive processes and has been claimed to be an immanent feature of human social interactions31. Nevertheless, it is scarcely researched in the domain of emotion recognition. According to the few available studies on emotion recognition in healthy adults, a reliable metacognitive resolution (i.e., a clear subjective discrimination between correct and incorrect recognition), together with a general overconfidence has been found32, 33. Furthermore, only direct trial-by-trial ratings, which can be used to estimate ‘relative meta-accuracy’, and not global beliefs about one’s abilities, were found to be predictive of performance in emotion recognition tasks7.Thus, while global metacognitive beliefs seem to be biased, confidence in one’s emotion recognition skills (i.e., a metacognitive judgment) can act as a reliable feedback mechanism in an emotion recognition context.
Emotion recognition alterations in SAD and ASD
While emotion recognition difficulties have sporadically been reported in SAD34, most research did not find deficits35 or even found a higher sensitivity, reflected by an emotion detection at lower expression intensities, to emotional expressions36, 37. Heightened attention to social cues also stands at the basis of established theoretical models of SAD38, 39 and has predominantly received support in form of a “negativity bias”40,41,42. In other words, negative expressions automatically attract more attention and are avoided at the same time, they are integrated more strongly in judging the self in social interactions, they are remembered better, and even ambiguous expressions are more likely to be judged negatively. Correspondingly, not clinically diagnosed individuals with high social anxiety trait levels have shown an emotion recognition advantage43, and specifically better recognition of negative expressions44, 45. For individuals on the autism spectrum, in contrast, difficulties in visual emotion recognition paradigms in which emotional facial or bodily expressions had to be matched to samples or labelled have mainly been described for all basic emotions, and, particularly, for fear46,47,48 (however, see49). Thus, quite specific particularities in facial emotion recognition have been associated with SAD and ASD. Factors that could be linked to, and potentially even contribute to, the occurrence of those particularities are, however, not well described yet.
In past research, individual differences in autistic traits and social anxiety traits have also been related to the usage of different strategies to recognize emotional expressions. When labelling full-body emotional expressions, high compared to low socially anxious individuals have been shown to attend to faces less, and more to expressive hands, thus using different visual cues50. In a study comparing recognition of sadness in static facial expressions versus point light displays, only individuals with low autistic traits, compared to individuals with high autistic traits, showed a recognition advantage for sad faces. Fear, in contrast, could be better recognized in point-light-displays by individuals with low autistic traits, and in faces by individuals with high autistic traits51. These findings suggest that, depending on clinical trait levels, different features might be used to identify others’ emotional states. Recently, it has even been suggested more broadly that emotions reach awareness via different pathways in individuals on the autism spectrum compared to neurotypical individuals52. Differences in processing facial emotional expressions, despite unimpaired emotion matching performance, have already been reported in autistic children on a neural level53. In our study with healthy participants, we aimed to explore whether the link between emotion recognition and two processes that have been suggested to promote emotion recognition, namely facial mimicry and metacognitive judgments, differs depending on social anxiety and autistic traits.
Altered mimicry in emotion recognition in SAD and ASD
Studies investigating the effects of social anxiety (disorder) on facial mimicry have reported inconsistent results: while some studies found intact mimicry in non-clinical but high socially anxious individuals19, 54, others demonstrated enhanced mimicry of negative expressions and diminished mimicry of positive ones55, 56 or stronger differential muscle activity between happy and angry expressions, for both the zygomaticus and the corrugator57. The literature on ASD gives a clearer picture: reduced automatic mimicry in individuals on the autism spectrum has been reported in many studies8, 58. Importantly, this reduction could not be explained by a generally lower facial expressiveness or an inability to mimic expressions, but by a mismatch between observed and produced facial muscle activity patterns59,60,61. Only few studies have described differences in facial mimicry alterations between different emotion categories, and findings are inconsistent. Namely, mimicry of angry, but not happy, facial expressions was reduced with higher autistic trait levels in females in one study62, while reduced mimicry of happy, but not sad, expressions has been related to higher autistic traits in another study63. Whether observed reductions in facial mimicry in high autistic trait levels are also linked to differences in emotion recognition performance has, however, not directly been investigated yet.
Altered metacognition in emotion recognition in SAD and ASD
Reduced metacognitive abilities have been proposed as a shared characteristic in different psychiatric disorders9, 64. Theoretical accounts on the development and maintenance of SAD have highlighted the importance of a negatively biased view on one’s own performance in a social context38, 39, together with an excessive monitoring of the self65. This global negative judgment of one’s own abilities might have evolved via repeated underestimation of (social) abilities66. However, metacognitive abilities in emotion recognition have yet not been directly tested in individuals with SAD. In contrast, the few studies on metacognitive judgments of social cognition in individuals on the autism spectrum have suggested a complex pattern of alteration. Some studies reported no differences between neurotypical individuals and individuals on the autism spectrum in calibrating confidence judgments to emotion recognition performance, that is, higher confidence rating for more accurate or faster recognition67, 68. A more recent study, however, found evidence for both an over- and underconfidence in contrast to actual performance in social cognitive tasks, including emotion recognition, in individuals on the autism spectrum compared to neurotypical individuals69. Both expressing low confidence in accurate trials as well as high confidence in incorrect trials should be reflected in a reduced metacognitive sensitivity, which Fleming and Lau70 defined as “the extent to which confidence discriminates between correct and incorrect trials” (p. 2). Given the limited knowledge about metacognition in the domain of emotion recognition and its relation to SAD and ASD, the current study aimed to explore two assumptions: (1) whether the negatively biased assessment of one’s performance in social situations in people with high social anxiety trait levels also translates to emotion recognition, and (2) whether the decreased metacognitive sensitivity related to higher autistic traits in the social-cognitive domain also specifically holds for emotion recognition performance.
Objectives of the current study
In the current study, we examined whether social anxiety and autistic traits modulate the links between facial mimicry and emotion recognition as well as between confidence judgments in own emotion recognition skills and emotion recognition. To investigate this, our participants first passively viewed naturalistic video clips of different facial expressions of emotion while we measured their facial muscle activity. In a subsequent task, participants indicated how strongly they associated the expressions with distinct emotion categories and were asked how confident they were in their judgments. Despite sharing social interaction difficulties in the global disorder definitions, previous research on emotion recognition, facial mimicry, and metacognitive judgments showed specific alterations associated with SAD and ASD. Therefore, we also expected to find distinct modulations for the two trait dimensions.
More specifically, confirming the suggested heightened sensitivity to social cues, higher levels of social anxiety were expected to be related to a recognition advantage of facial expressions, resulting in higher accuracy rates. According to the negativity bias findings, this advantage should be specifically pronounced for negative expressions (i.e., anger, fear, and sadness). As individuals with SAD have been found to report a generalized underconfidence in their social skills, we expected that, despite an improved emotion recognition performance, elevated social anxiety traits would be related to reduced overall confidence in the performance. Moreover, based on observations that confidence judgments were predictive of emotion recognition accuracy in healthy subjects, we would like to explore whether the scaling of confidence judgments to actual emotion recognition performance is altered depending on social anxiety traits. In line with a proposed facilitating role of facial mimicry in emotion recognition, stronger facial mimicry might be assumed with higher social anxiety traits. Empirical evidence for both relationships, social anxiety (disorder) and facial mimicry as well as facial mimicry and emotion recognition, is, however, inconclusive. Therefore, we aimed to directly test whether the role of facial mimicry in emotion recognition is altered depending on an individual’s social anxiety traits.
Regarding autistic traits, we expected to observe an overall worse recognition of naturalistic dynamic facial expressions in association with higher levels of autistic traits, which should be most pronounced for fearful faces. Higher autistic trait levels were further expected to be associated with less facial mimicry, and this reduction was expected to be strongest for negative expressions. Furthermore, as automatic facial mimicry has been suggested to be impaired in ASD, the information about facial muscle activity might also be less well integrated in emotion recognition. Accordingly, we explored whether a positive relationship between facial mimicry and emotion recognition would be less pronounced in individuals with higher autistic trait levels. Lastly, extending on the few findings in clinical samples, we expected lower metacognitive sensitivity in relation to higher autistic traits. Hence, confidence judgments should be less predictive of actual emotion recognition accuracy in individuals with higher autistic trait levels. Given the little and inconclusive evidence on metacognition in emotion recognition in healthy and clinical populations, our analyses regarding this research question were explorative.
Methods
Participants
Fifty-seven healthy participants were recruited from the Leiden University student population (50 female and seven male). Their ages ranged from 18 to 30 years old (M = 22.75, SD = 3.27) and they all reported normal or corrected-to-normal vision. None of the participants reported current or past psychological or neurological disorders. Participation in the study was voluntary and written consent was obtained prior to the experiment. Participants received either two university credits or a monetary reward of six euros as reimbursement. The study has been executed in accordance with the Declaration of Helsinki and approved by the local ethics committee of the Faculty of Social and Behavioral Sciences at Leiden University (# 2020-02-10-M.E. Kret-V1-2117).
In the scope of a Master thesis project, an a priori power analysis was run for this study, treating clinical traits as a categorical variable (low vs. high trait levels). Based on a similar previous study that found significant group effects with medium effect sizes71, we estimated our ideal sample size with the Power Analysis for General ANOVA application (PANGEA)72. With 30 participants per group, hence 60 participants in total, a group effect of d = 0.50 should be found with a power of 0.901. Because of the COVID-19 regulations in the Netherlands, we had to stop data collection prematurely and ended up with 57 participants in total (56 participants: power of 0.879). For the analyses in this manuscript, we treated the clinical trait dimensions as continuous variables, thereby increasing the validity of the approach as well as statistical power73.
Stimuli
Following the call for more naturalistic stimuli in research on emotion perception, we chose the FEEDTUM database74 as a source for our stimuli. This database encompasses videotaped spontaneous (i.e., non-instructed) reactions to video clips inducing the six different basic emotions and neutral control expressions. All depicted individuals provided informed consent for the usage of the videos for research purposes, including distribution and publication of the material, in the original study. Permission to use the material under CC-by and to publish example images in scientific journals, such as in Fig. 1, was granted to the first author of this study by the creators of the database. Based on the choice of stimuli in a previous study investigating facial mimicry and emotion recognition in depression71, we included facial expressions of anger, fear, happiness, sadness, surprise, and neutral. Disgust was not included, which is a basic emotion that (next to surprise) is typically less investigated in studies on emotion recognition alterations in SAD and ASD35, 46. For each facial expression, video clips of ten individuals (five females and five males) were selected based on the following decision pipeline: first, videos were judged on their quality, and blurry or shaky videos were excluded. Second, individuals wearing glasses or individuals with hair in front of their eyes were excluded as these features made their facial expressions more difficult to recognize. Lastly, all remaining video clips were evaluated by the automated facial expression recognition software FaceReader 7.175 to ensure that the emotion label of the stimulus provided by the database could also be detected in the video. After this selection procedure, the video clips were cut to a uniform length of 2 s (500 ms neutral expression followed by 1500 ms of each category’s expression). Lastly, the video clips were standardized by removing the original backgrounds using Adobe After Effects76, and by replacing them with a uniform gray colored background (RGB color code: 145, 145, 145). This led to a total of 60 two-second videos with a grey background showing ten individuals (five males and five females) per facial expression.

Illustration of a Facial Emotion Recognition task trial. Each trial started with a fixation screen and ended with an intertrial interval (ITI) screen. The dotted line indicates that this sequence was repeated until all 60 videos were presented once.
Procedure
Participants were brought to a quiet room, in which they were given written and verbal instructions about the experimental procedure. After filling in the informed consent form, electrodes were attached to the participants’ faces for the facial electromyography recordings (see Measurements section). During the tasks, participants were seated in 50 cm distance of a Philips screen with a resolution of 1920 × 1080 pixels (23.6"), on which the stimuli (720 × 480 pixels, average visual angle: 22.12° horizontal and 14.85° vertical) were presented using E-Prime 3.0 software77. The grey background colour of all screens was set to the background colour of the stimuli (RGB color code: 145, 145, 145). The same 60 emotional facial expression videos were presented in a random order in two consecutive tasks, a passive viewing task during which the participants’ facial muscle activity was recorded, and a facial emotion recognition task. The rationale behind the two separate tasks was to avoid that participants would be biased in their perception and their facial mimicry responses in the passive viewing task by being aware of the possible emotion category labels (i.e., top-down modulation). In the passive viewing task, participants were instructed to only look at the stimuli without performing any action. Each trial started with the presentation of a black fixation cross against a grey background for one second, and was followed by one of the 60 video stimuli for two seconds. The end of a trial was marked by a grey inter-trial interval (ITI) screen, which appeared with a jittered duration of either 5750, 6000, or 6250 ms. To account for the possibility of missing observations due to noisy data, participants viewed each of the 60 videos twice, in two separate blocks, resulting in 120 trials in total. Between the blocks, participant could take a self-paced break. The passive viewing task lasted around 20 min in total. After the passive viewing task, the electrodes were detached from the participants’ faces and the experiment continued with the facial emotion recognition task. In this second task, the participants viewed all 60 video stimuli once again (thus three times in total) but were now instructed to answer questions about them. Similar to the passive viewing task, each trial started with the fixation cross screen for one second and one of the 60 video stimuli (2 s) was presented afterwards. Once the video disappeared, participants were asked to judge the displayed expression. More specifically, on the first question screen, they were asked to rate the expression according to its representativeness of the six expression categories that could be displayed: anger, fear, happiness, neutral, sadness, and surprise ('To what degree does the expression relate to the emotions below?'). Each expression category was accompanied by horizontal sliders ranging from 'not at all' to 'very much' and participants had to move the slider to indicate their judgment. The values of the sliders ranged from 0 to 100 in steps of 10 (0, 10, 20, etc.), which were not visible to the participants. The next screen contained three questions: (1) 'How intense was the expression displayed in the video?' to measure perceived emotional intensity; (2) 'Are you confident about your decision?' to measure confidence in own performance; and (3) 'Did you find the trial simple?' to measure simplicity71. Again, all questions were accompanied by a slider ranging from 'not at all' to 'very much' with underlying values ranging from 0 to 100 in steps of 10. Thus, higher scores indicated higher ratings on perceived intensity, confidence, and simplicity, respectively. After the second question screen, a grey inter-trial interval (ITI) screen appeared for three seconds. In total, the participants completed 60 trials, rating all stimuli from the first task, plus three additional practice trials showing a different individual to familiarize them with the task. After 30 trials, participants could take a self-paced break and the entire facial emotion recognition task lasted approximately 25–30 min. A visualization of one facial emotion recognition trial can be found in Fig. 1.
Importantly, only the ratings on the association of the displayed expression with the expression categories (first question screen) and the confidence ratings on the second screen were relevant to answer our hypotheses. As we did not formulate specific hypotheses about alterations in perceived emotional intensity ratings related to social anxiety or autistic traits, explorative analyses on this rating variable can be found in the Supplemental Materials S1. Furthermore, the simplicity ratings provided insights in how difficult emotion recognition with this novel stimulus set was perceived. Overall, happy expressions received the highest simplicity ratings (M = 69.39, SD = 27.15), followed by surprised expressions, (M = 62.51, SD = 25.35), fearful expressions (M = 50.93, SD = 26.19), angry expressions (M = 45.05, SD = 25.66), neutral expressions (M = 36.42, SD = 33.74), and sad expressions (M = 36.19, SD = 25.56). Some might consider differences in simplicity between emotion categories as potential confound in predicting emotion recognition accuracy (i.e., higher simplicity ratings might systematically be linked to more accurate choices). Yet, these differences have been proposed to arise due to factors that are inherently linked to the specific emotional expression, such as a higher familiarity with increased exposure in daily life78, 79. We aimed for higher ecological validity in examining facial emotion recognition by using spontaneous and non-acted expressions in the current study. Controlling for simplicity, in contrast, would detach our results from emotion recognition in everyday life, which is why we decided against it. Lastly, participants filled in the Liebowitz Social Anxiety Scale (LSAS)80 and the Autism Spectrum Quotient (AQ; see Measurements section)81. Upon completion of the experiment, participants were given a written and verbal debriefing about the goal of the study and were reimbursed. In total, the experiment lasted around 55 min, including instructions and the attachment of the facial electromyography electrodes.
Measurements
Facial electromyography (fEMG)
Facial electromyography (fEMG) was measured on the left side of the face of all our participants, following the guidelines of Fridlund and Cacioppo82. To specify, two reusable 4 mm Ag/AgCl surface electrodes were placed over the Corrugator Supercilii region, which allowed us to measure mimicry responses to sad, fearful, and angry expressions (according to the EMFACS definition) as well as to happiness expressions (as shown in previous research). Other two electrodes were placed over the Zygomaticus Major region, which allowed us to measure mimicry responses to happy expressions. Additionally, a ground electrode was added on the top of the forehead. The signal was transmitted and amplified using a Biopac MP150 system combined with a BioNomadix 2 channel wireless EMG amplifier. Data recordings were made in AcqKnowledge 4.383 using a sampling rate of 1000 Hz. Event markers as defined in the E-Prime77 tasks were sent via a parallel port and saved within an event marker channel. For data preprocessing, the EMG recordings were loaded into the PhysioData Toolbox84 in which they were rectified and filtered with a 28 Hz high cut-off filter, a 500 Hz low cut-off filter, and a 50 Hz notch filter. For each trial, separate epochs were defined for the fixation period (1 s), the first 500 ms of stimulus presentation in which a neutral expression was shown (later defined as baseline), the subsequent 1500 ms in which the emotional expression was shown (later defined as response), and the first 1500 ms of the blank screen after stimulus presentation. Within these epochs, the EMG signal was downsampled by calculating the average signal within consecutive 100 ms time bins. The combined data from all subjects was then exported into MATLAB for further preprocessing. First, an automated artifact detection, which was inspired by Dignath et al.85, was conducted. More specifically, for each subject and each muscle region, we checked the distribution of the EMG signal for extreme values (± 3.5 SDs) in the time bins regarding the absolute value of each time bin and the relative differences between subsequent bins. This was performed in relation to (1) the entire time interval of interest per trial (5 baseline time bins and 15 response time bins) and (2) the distribution of baseline time bins in the same position across trials. If more than 50% of all time bins (20) or more than 50% of the baseline time bins (5) belonging to one trial had extreme values, this trial was entirely excluded from the analysis. Otherwise, the respective time bins were replaced with missing values. Across all subjects, 17 trials (0.002%) were excluded from the corrugator analysis and 150 trials (0.02%) from the zygomaticus analysis. After excluding the marked time bins, a baseline correction was performed by subtracting the mean EMG activity of all baseline time bins belonging to one trial from the respective response time bins. The baseline-corrected EMG data was then z-scored for each participant and each muscle region. Furthermore, the data was summarized on a trial level by averaging the last second of each trial’s response window (last 10 time bins) for each participant and each muscle region as well as by averaging across the two presentations of each of the 60 stimuli to end up with the same amount of observations as for the rating data (trial-averaged data; used as predictor in generalized linear mixed models). Lastly, the data was also summarized on a participant level by creating the average of the same time window across trials for each participant, each muscle region and each emotion category (category-averaged, used as outcome variable in linear models).
Questionnaires
Social anxiety traits measure
We used the Liebowitz Social Anxiety Scale (LSAS)80 to measure self-reported social anxiety traits in our non-clinical sample. The LSAS is designed to assess fear and avoidance levels of individuals with social phobia in a range of social interaction and performance scenarios. The questionnaire contains 24 items in total. Respondents score the items for fear and avoidance separately on a 4-point Likert scale, fear: 0 (= None), 1 (= Mild), 2 (= Moderate), 3 (= Severe); avoidance: 0 (= Never), 1 (= Occasionally), 2 (= Often), 3 (= Usually). The scores are all added up to a total sum of all subscales, with higher scores indicating a higher severity of social anxiety symptoms. One participant had missing data for one questionnaire item and another participant had missing data for two questionnaire items, which were estimated using the mice-package86 for multiple imputation. The LSAS showed an excellent internal consistency in our sample (α = 0.91, 95% CI [0.88, 0.95]). LSAS scores ranged from 7 to 73 (M = 38.53, SD = 17.53), with 30 participants (52.63%) exceeding a score of 30. This score has been described as the best cut-off to discriminate between non-anxious individuals and individuals with SAD87, 88. Thus, a broad spectrum of social anxiety trait levels was covered in our sample, with half of the participants showing an indication of clinically relevant social anxiety. The average LSAS scores were considerably higher compared to the healthy validation sample of the LSAS self-report version (M = 13.49, SD = 23.70)89. With a skewness of 0.21 and a kurtosis of 2.08, the distribution of the LSAS scores showed to be slightly platykurtic, yet close to normal (see Fig. S1A in the Supplemental Material).
Autistic traits measure
The Autism-Spectrum Quotient (AQ)81 is a self-report questionnaire, which was created to measure traits associated with the autism spectrum. The AQ consists of 50 items in total and can be divided into five subscales (10 items each) assessing different domains: social skill, attention switching, attention to detail, communication, and imagination. Respondents indicate how strongly each item applies to them based on a 4-point Likert scale ranging from 1 (= definitely agree), 2 (= slightly agree), 3 (= slightly disagree), and 4 (definitely disagree), and some items are reversely scored to prevent response biases. All item scores are added up to a total sum score, with higher scores reflecting higher autistic trait levels. One participant did not complete the AQ and was therefore excluded from all analyses investigating effects of autistic traits. Furthermore, we had to estimate three single item scores using the mice-package86 for multiple imputation as one participant did not respond to one item and another participant did not respond to two items. Internal consistency of the AQ in our sample was good, α = 0.83, 95% CI [0.76, 0.89]. The range of AQ scores was between 2 – 39 (M = 16.38, SD = 7.34), which is highly similar to meta-analytic results on AQ scores in general population samples (M = 16.94, 95% CI [11.6, 20.0])90. Only 3 participants (5.26%) had a higher AQ score than 32, which indicates autistic trait levels of clinical significance. The skewness and kurtosis of the AQ score distribution were 1.05 and 4.17 respectively, thus showing a positive skew (see Fig. S1B in the Supplemental Material).
Data analysis
Spearman's rank correlation revealed that autistic traits and social anxiety traits, reflected by the scores on the two questionnaires, were not significantly associated with each other, rs = 0.04, p = 0.784. Our sample showed both variability within each trait dimension that was similar to studies with larger samples (see Questionnaire section) and independence between the trait dimensions, allowing for separate analyses for the two trait dimensions. Emotion recognition accuracy was calculated by determining the expression category with the highest slider score and comparing it to the predefined category of the stimulus for each trial71. If there was a match between the presented and the perceived expression category, a trial was scored as correct (1) whereas it was scored as incorrect (0) in case of a mismatch. Trials in which two expression categories received the same slider scores were discounted from the analysis. To check the robustness of this approach, we re-ran all analyses on accuracy with a relative accuracy score, which was calculated by subtracting the mean score of all other expression categories from score of the correct expression category91. The results were overall highly similar and are reported in the Supplemental Materials S1. All analyses were performed in R 4.0.192, using the lmerTest package93 for fitting the (generalized) linear mixed models ([G]LMMs), the multcomp package94 for general hypotheses testing, the sjPlot package95 for creating the tables and both the sjPlot package and ggplot296 for creating the plots.
Behavioural analysis
Accuracy and confidence
In order to test whether social anxiety traits were associated with better emotion recognition performance for negative expressions and whether emotion recognition accuracy was generally reduced with higher autistic traits, we calculated two binomial GLMMs on accuracy with emotion category (anger, fear, happiness, sadness, surprise, and neutral), the respective trait dimension (autistic traits or social anxiety traits), and their interaction as fixed effects. Both participant ID and the identity of the stimulus were added as random effects (random intercept). Furthermore, we fitted two LMMs on emotion recognition confidence to test the association between confidence level and (i) social anxiety traits, (ii) autistic traits. The models had the same fixed and random effects structure as the accuracy models. Coefficients for the emotion categories (main effects and interactions) were calculated by contrasting each single category against the mean of all categories (sum coding) to determine significant deviations from mean accuracy (main effect) or the mean effect of a trait dimension (interaction). For the neutral category, coefficients were calculated and tested (z-tests) using general hypotheses testing. Lastly, we explored whether the relation between confidence and emotion recognition was altered depending on an individual's clinical trait levels as well as the presented expression. To do so, we added emotion recognition confidence and all 2-way interactions as well as the 3-way interactions with emotion category and the respective clinical trait dimension to the accuracy models. The resulting model fits were the following:
-
1.
LSAS * EMOTION CATEGORY—> EMOTION RECOGNITION ACCURACY
-
2.
AQ * EMOTION CATEGORY—> EMOTION RECOGNITION ACCURACY
-
3.
LSAS * EMOTION CATEGORY—> EMOTION RECOGNTION CONFIDENCE
-
4.
AQ * EMOTION CATEGORY—> EMOTION RECOGNTION CONFIDENCE
-
5.
LSAS * EMOTION CATEGORY * CONFIDENCE—> EMOTION RECOGNITION ACCURACY (exploratory)
-
6.
AQ * EMOTION CATEGORY * CONFIDENCE—> EMOTION RECOGNITION ACCURACY (exploratory)
Metacognitive sensitivity
To examine how well an individual's confidence ratings could distinguish between accurate and inaccurate trials in the emotion recognition task, we calculated the hit and false alarm rate pairs with increasing confidence levels (11, according to points on the Likert scale) for each subject and employed the area under the type 2 ROC curve (AUROC2) approach according to Fleming and Lau70. More specifically, each confidence level was taken as a criterion to distinguish between low and high confidence trials; starting with a criterion in which only zeroes were regarded as low confidence ratings and all higher values were regarded as high confidence ratings, up until a criterion in which all trials below the highest confidence rating (100) were regarded as low confidence trials and only the highest rating was regarded as high confidence. The resulting probabilities for hits, p(high confidence|correct), and false alarms, p(high confidence|incorrect), were plotted against each other for each confidence level. The resulting area under this ROC2 curve was taken as an index for the subject's metacognitive sensitivity, describing how well an individual's confidence ratings were scaled to actual emotion recognition accuracy. The link to each clinical trait was then tested with a correlational analysis.
Facial EMG analysis
Facial muscle activity (mimicry)
By measuring facial muscle activity over the Corrugator Supercilii and Zygomaticus major regions, we could assess mimicry responses to angry, happy, sad and fearful expressions, with neutral expressions acting as a reference category. In order to examine whether social anxiety traits are associated to an enhanced mimicry of specifically angry (negative) expressions, we fitted a linear model on the category-averaged corrugator activity (i.e., taking the mean corrugator activity of all trials belonging to the same emotion category) with emotion category, social anxiety traits and their interaction as fixed effects. We also aimed to explore zygomaticus activity for mimicry of happy expressions and, therefore, used the same independent variables to predict category-averaged zygomaticus activity (i.e., taking the mean zygomaticus activity of all trials belonging to the same emotion category). By replacing social anxiety traits with autistic traits in the other two linear models on category-averaged corrugator and zygomaticus activity, we then tested whether typical mimicry patterns are indeed reduced with higher autistic traits (i.e., less corrugator activity for negative expressions (specifically anger), less zygomaticus activity for happy expressions and less decrease in corrugator activity for happy expressions). Coefficients for the emotion categories (main effects and interactions) were calculated by contrasting the respective category against the neutral reference category. Since the residual plots of the two model fits on zygomaticus activity showed violation of the normality assumption as well as heterogeneous error estimates, we calculated non-parametric estimates of the predictor effects and confidence intervals, using 1000 bootstrap iterations, for these two models. As neither main effects of clinical traits nor interaction effects with emotion category were found, the results, including significant effects of emotion category on EMG activity, are reported in the Supplemental Materials (Tables S7–S10).
Link between facial muscle activity (mimicry) and emotion recognition accuracy
As outlined in the introduction (see Mimicry and Emotion Recognition section), the association between facial muscle activity and emotions, including mimicry of expressions, is specific for each emotion category. Emotion recognition should only be informed by facial muscle activity that is in line with the assumed mimicry responses for the specific emotion (e.g., increase in zygomaticus activity and decrease in corrugator activity for happy expressions, and increase in corrugator for sad expressions). Therefore, we ran separate analyses for the emotion categories anger, happiness, sadness, and fear to investigate the relationship between intraindividual differences in facial muscle activity and emotion recognition accuracy in the context of varying clinical trait levels. Binomial GLMMs were fitted on emotion recognition accuracy with the trial-averaged EMG activity over the two muscle regions (corrugator and zygomaticus) as distinct predictors, as well as one of the clinical trait scores and both two-way interactions (8 models in total). Similar to the behavioural accuracy models, random intercepts were added for the subject as well as the stimulus identity. This resulted in the following models:
-
1.
LSAS * CORRUGATOR—> ANGER RECOGNITION ACCURACY
-
2.
LSAS * CORRUGATOR + LSAS * ZYGOMATICUS—> HAPPINESS RECOGNITION ACCURACY
-
3.
LSAS * CORRUGATOR—> SADNESS RECOGNITION ACCURACY
-
4.
LSAS * CORRUGATOR—> FEAR RECOGNITION ACCURACY
-
5.
AQ * CORRUGATOR—> ANGER RECOGNITION ACCURACY
-
6.
AQ * CORRUGATOR + AQ * ZYGOMATICUS—> HAPPINESS RECOGNITION ACCURACY
-
7.
AQ * CORRUGATOR—> SADNESS RECOGNITION ACCURACY
-
8.
AQ * CORRUGATOR—> FEAR RECOGNITION ACCURACY
Ethical Approval
The study was reviewed and approved by the Psychology Ethics Committee of Leiden University (2020-02-10-M.E. Kret-V1-2117).
Informed consent
Written consent was obtained from all participants.
Results
Behavioural results
Accuracy in emotion recognition
Social anxiety traits
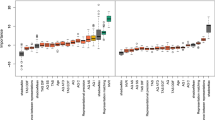
The first binomial GLMM on emotion recognition accuracy included emotion category, social anxiety traits, and their interaction as predictors. Results showed a significant main effect of emotion category, χ2(5) = 702.880, p < 0.001. Emotion recognition performance for happy, surprised, and neutral expressions was significantly better than average recognition performance, happy: OR = 10.834, z = 10.797, p < 0.001, surprise: OR = 3.027, z = 7.654, p < 0.001, neutral: OR = 2.337, z = 6.420, p < 0.001. In contrast, sad and fearful expressions were significantly worse recognized than average, OR = 0.232, z = − 14.336, p < 0.001, and OR = 0.066, z = − 23.445, p < 0.001 respectively. All the other effects or interactions were not significant. This suggests that recognition accuracy was predicted by the emotional content displayed in the video, independent of the level of social anxiety traits (see Fig. 2A). An overview of the model fit can be found in the Supplemental Material (see Table S1).

Predicted emotion recognition accuracies depending on (A) social anxiety trait levels and (B) autistic trait levels by emotion category (anger, fear, sadness, surprise happiness, neutral). In the model fits, accuracy was coded binomial (0–1 values). For illustrative purposes, predicted accuracies for mean values as well as mean values ± 1 SD of the continuous variables social anxiety traits and autistic traits are depicted in percentages. Whiskers represent confidence intervals and significant effects are marked with an asterisk. The dashed horizontal line indicates mean predicted accuracy (across all categories and trait levels).
Autistic traits
The second binomial GLMM on emotion recognition accuracy included emotion category, autistic traits, and their interaction as predictors. Results showed a significant effect of emotion category, χ2(5) = 666.374, p < 0.001, a significant effect of autistic traits, χ2(1) = 8.985, p = 0.003, and a significant interaction between autistic traits and emotion category, χ2 (5) = 21.606, p = 0.001. The overall negative association between autistic traits and emotion recognition accuracy, OR = 0.763, z = − 2.998, p = 0.003, was most pronounced for fearful expressions, OR = 0.637, z = − 3.292, p = 0.001. Recognition of sad expressions was less negatively affected by autistic traits compared to the overall performance, OR = 1.322, z = 2.821, p = 0.005, and similarly recognition of surprised expressions, OR = 1.314, z = 2.067, p = 0.039 (for all other effects and more detailed information, see Fig. 2B and Table S2 in the Supplemental Material). Thus, the expected overall negative association between autistic traits and emotion recognition performance seems specifically pronounced for fearful facial expressions.
Confidence in emotion recognition
Social anxiety traits
In the first LMM on confidence in emotion recognition, with emotion category, social anxiety traits, and their interaction as predictors, significant effects of both emotion category, F(5, 3344) = 118.666, p < 0.001, and social anxiety, F(1,3344) = 5.362, p = 0.024, could be observed. Compared to the average confidence judgments across emotion categories, participants were significantly more confident in judging happy expressions, β = 0.614, t(3344) = 19.695, p < 0.001, neutral expressions, β = 0.193, z = 6.186, p < 0.001, and surprised expressions, β = 0.084, t(3344) = 2.699, p = 0.007. For the other emotional expressions, confidence was significantly reduced compared to the average, angry: β = − 0.247, t(3344) = − 7.932, p < 0.001, fearful: β = − 0.287, t(6761) = − 9.209, < 0.001, sad: β = − 0.356, t(3344) = − 11.438, p < 0.001. The association between social anxiety traits and confidence was negative, β = − 0.132, t(3344) = − 2.316, p = 0.024, and did not vary by emotion category (i.e., no interaction). Thus, independent of the displayed expression, confidence judgments were significantly lower with higher social anxiety trait levels (for a detailed description of the model fit, see Fig. 3A and Table S3 in the Supplemental Material).

Predicted confidence in emotion recognition depending on (A) social anxiety trait levels and (B) autistic trait levels by emotion category (anger, fear, sadness, surprise happiness, neutral). For illustrative purposes, predicted accuracies for mean values as well as mean values ± 1 SD of the continuous variables social anxiety traits and autistic traits are depicted. Whiskers represent confidence intervals and significant effects are marked with an asterisk. The dashed horizontal line indicates mean predicted confidence (across all categories and trait levels).
Autistic traits
In the LMM that included autistic traits instead of social anxiety traits to predict confidence in emotion recognition, the effect of emotion category was significant, F(5, 3285) = 118.164, p < 0.001, as was the interaction between emotion category and autistic traits, F(5, 3285) = 9.531, p < 0.001. While for neutral and happy facial expressions confidence was significantly reduced with higher autistic traits compared to the average effect of autistic traits on confidence ratings, β = − 0.148, z = − 4.663, p < 0.001, and β = − 0.104, t(3285) = − 3.271, p = 0.001 respectively, this effect was reversed for displays of fear and sadness. For those two categories, autistic traits were associated with higher confidence ratings compared to the average effect of autistic traits on confidence, fear: β = 0.102, t(3285) = 3.229, p = 0.001; sadness: β = 0.118, t(3285) = 3.715, p < 0.001 (see Fig. 3B and Table S4 in the Supplemental Material).
Link between confidence and emotion recognition
Social anxiety and autistic traits
When exploring the link between confidence and accuracy in emotion recognition in relation to the clinical trait dimensions on a trial level and by emotion category, neither of the trait dimension had a substantial impact on this link (see Tables S5 and S6 in the Supplemental Material for the entire model fits).
Metacognitive sensitivity
According to Mahalanobis distance measures, two participants had to be excluded from the correlation analysis between social anxiety traits and AUROC2, and three participants had to be excluded from the correlation analysis between autistic traits and AUROC2. After excluding these bivariate outliers, all distributions did not majorly deviate from normality. The two correlational analyses between the clinical trait dimension and metacognitive sensitivity (AUROC2) revealed that autistic traits, but not social anxiety traits, were significantly related to metacognitive sensitivity (see Fig. 4). As expected, metacognitive sensitivity was reduced with higher autistic traits, r = − 0.489, t(51) = − 4.008, p < 0.001 and rs = − 0.476, p < 0.001 and no significant relation was found for social anxiety, r = − 0.222, t(53) = − 1.66, p = 0.103 and rs = − 0.251, p = 0.064.

Relationship between (A) social anxiety traits and (B) autistic traits and metacognitive sensitivity, which was indexed by the area under the type 2 ROC curve (AUROC2). The blue line indicates the estimated linear relationship, with significant relationships being marked by an asterisk.
Facial electromyography (fEMG) results
Facial mimicry in emotion recognition
Social anxiety traits
There was no significant interaction between corrugator activity and social anxiety traits in predicting emotion recognition accuracy of negative facial expressions, (i.e., anger, fear, and sadness, see Tables S11, S13 and S14 for the three model fits). The model on sad expressions did, however, reveal that accuracy was higher when the corrugator muscle was more strongly activated, χ2 (1) = 4.631, p = 0.031, OR = 1.585. Furthermore, both zygomaticus activity as well as its interaction with social anxiety traits were significant predictors in the model on happy expressions, χ2(1) = 4.331, p = 0.037, OR = 6.240, and χ2(1) = − 2.017, p = 0.044, OR = 0.213 respectively (Table S12 in the Supplemental Material). Hence, while the significant effect of zygomaticus activity hints towards a facilitating role of mimicry of smiles in emotion recognition, this link seems to be weakened with higher social anxiety traits. When examining the predicted value plot (see Fig. 5A), this effect, however, seems to be mainly driven by stronger variation in accuracies (i.e., also inaccurate responses) in individuals with lower social anxiety traits when the zygomaticus was not strongly activated. Otherwise, recognition of happy expressions was at ceiling and not much variation in relation to social anxiety trait levels could be observed.

(A) The relationship between predicted accuracy in recognizing happy expressions and the corresponding filtered, baseline-corrected and z-scored zygomaticus activity depending on social anxiety trait levels. (B) The relationship between predicted accuracy in recognizing sad expressions and the corresponding filtered, baseline-corrected and z-scored Corrugator activity depending on autistic trait levels. (C) The relationship between predicted accuracy in recognizing happy expressions and the corresponding filtered, baseline-corrected and z-scored zygomaticus activity depending on autistic trait levels. In the model fits, accuracy was coded binomial (0–1 values). For illustrative purposes, predicted accuracies for mean values as well as mean values ± 1 SD of the continuous variables social anxiety traits and autistic traits are depicted. Shaded areas represent confidence intervals.
Autistic traits
Similar to the models including social anxiety traits, there was no significant interaction between autistic traits and corrugator activity in accurately recognizing fearful or angry facial expressions (see Tables S15 and S17 in the Supplemental Material for the model fits). Contrasting the other negative expressions, the model on the recognition of sad expressions showed a significant interaction between corrugator activity and autistic traits as well as a significant main effect of corrugator activity. More specifically, in line with the social anxiety model, sad facial expressions were better recognized with higher corrugator activity, χ2(1) = 4.556, p = 0.033, OR = 1.597. This relationship was, however, weaker for higher autistic traits, interaction: χ2(1) = 4.142, p = 0.042, OR = 0.668 (see Fig. 5B and Table S18). Furthermore, significant effects of zygomaticus activity and of corrugator activity as well as a significant interaction between zygomaticus activity and autistic traits were found in the model on happy expressions, zygomaticus: χ2(1) = 5.300, p = 0.021, OR = 14.184, corrugator: χ2(1) = 4.679, p = 0.031, OR = 0.069, autistic traits*zygomaticus: χ2(1) = 5.503, p = 0.019, OR = 0.137. Thus, in addition to the activation of the zygomaticus, which was already described previously, a relaxation of the corrugator might facilitate the recognition of happy expressions. Similar to the model including social anxiety traits, autistic traits had a negative effect on the positive association between zygomaticus activity and accuracy in the recognition of happy expressions (see Fig. 5C and Table S16 in the Supplemental Material). This interaction again seemed to be driven by cases of low zygomaticus activity associated with inaccurate responses, but this time linked to low autistic trait levels.
Discussion
In our study, we provided evidence that autistic traits and social anxiety traits are distinctly related to bottom-up (i.e., mimicry) and top-down (i.e., metacognition) components in emotion perception. Specifically, while individuals with higher social anxiety traits had significantly less confidence in their performance regarding all emotion categories, despite an unaltered actual emotion recognition performance, individuals with higher autistic traits were less accurate in the recognition of emotions, and in particular of fearful expressions. Furthermore, individuals with higher autistic traits seemed to be less able to calibrate their confidence judgments to their actual emotion recognition performance, as they displayed a poorer metacognitive sensitivity. Unexpectedly, we did not observe alterations in emotion-specific facial muscle mimicry with regard to either social anxiety or autistic traits. Yet, we found indications that mimicry of frowning, indexed by corrugator activation, might facilitate the recognition of sad expressions, whereas mimicry of smiling, indexed by zygomaticus activation (and potentially relaxation of the corrugator), might support the recognition of happy expressions. Crucially, both links were less pronounced with higher autistic traits, while there was only weak evidence for a negative effect of social anxiety traits on the link between zygomaticus activity and accuracy in recognizing happy expressions.
Contradicting our expectations, we did not find a negativity bias (i.e., an improved recognition of negative expressions) with higher social anxiety traits reflected in our main analysis on the recognition accuracy of the displayed facial expressions. When using relative accuracy scores as an outcome (see Table S20 in the Supplemental Material), however, a better recognition of angry facial expressions with higher social anxiety trait levels could be observed. Given that an improved recognition of negative expressions in SAD was also not consistently found in the literature35, the effect seems unstable and additional factors might play a role. For example, a heightened sensitivity to negative expressions in social anxiety (disorder) might only occur under brief exposure times or when actual interactions with the expresser could be expected97. In this study, the presentation time was 2 s and the participants were not engaged in any interaction. Effects related to biases in early visual attention (< 500 ms) or to the fear of being negatively judged by an interaction partner were, therefore, highly unlikely. Importantly, social anxiety traits had the expected impact on the confidence judgment with regard to emotion recognition in our study. For all expression categories, confidence was reduced with higher social anxiety traits. The underconfidence in performance did, however, not affect the general positive link between confidence in emotion recognition and actual performance. Thus, while participants seemed to be able to calibrate their confidence ratings according to their recognition performance, a relative reduction in the confidence scores might have occurred with higher social anxiety traits. This observation might be a reflection of self-related negative beliefs about one’s own social skills in high socially anxious people, which were likely formed in a public setting66 and translated to a more global negative social skill assessment.
Theoretical models on SAD highlight low confidence in own social performance as a relevant cognitive bias in the development and maintenance of the disorder38, 39, 98. Evidence for this bias has been found in various studies contrasting social performance and subjective evaluations in real-life scenarios99,100,101. The retrospective evaluation of one’s performance in a social situation, so-called post-event processing, has been especially suggested to contribute to negative beliefs about one’s social skills102. In both highly socially anxious individuals103 and individuals with a SAD diagnosis104, negatively-biased post-event processing has been shown to be more frequent, and positively related to social anxiety (symptoms). The lower confidence in emotion recognition associated with higher social anxiety traits in our study might also arise from doubts in one’s own ability to recognize another person’s emotional state correctly.
Facial muscle responses to emotional expressions were not found to be altered depending on social anxiety traits in our sample. This suggests that not only explicit emotion labelling but also implicit, automatic processes, namely facial mimicry, seem to be comparable across varying levels of social anxiety traits. In our study, there was also little evidence to assume that the link between facial muscle activity and emotion recognition accuracy would be modulated by social anxiety traits. The weakened positive association between zygomaticus responses and the accurate labelling of happy expressions with higher social anxiety trait levels in our study was most likely due to stronger variability in accuracies (i.e., also inaccurate responses despite close to ceiling performance overall) when zygomaticus activity was low. Additionally, the effect could not be reproduced in the analyses with relative accuracy as an outcome (see Supplemental Material S1). Taken our findings related to social anxiety traits together, heightened social anxiety trait levels were not associated with poorer emotion recognition performance or alterations in the link between facial mimicry and emotion recognition. Yet, confidence in emotion recognition was lower with higher social anxiety trait levels, which indicates that negative beliefs about one’s skills might also exist in the domain of emotion recognition. In order to overcome this and other cognitive biases, Metacognitive Training can be a useful tool in the treatment of SAD105.
In line with previous studies describing worse performance in emotion recognition tasks associated with ASD, we observed overall reduced accuracies with higher autistic traits, which became most apparent for fearful expressions46,47,48. The recognition of sad expressions, on the other hand, was not as strongly affected by autistic traits in the main analysis, and even improved with higher autistic trait levels in the relative accuracy analysis (see Supplemental Material S1). This observation is compatible with a previous study, which reported a better recognition of sad facial expressions with higher autistic trait levels106. Confidence in recognizing displays of sadness as well as fear was rated higher with higher autistic traits. Neutral and happy facial expressions, in contrast, received lower confidence ratings with higher autistic trait levels. Given the negative impact of autistic trait levels on the recognition of fear displays, higher confidence ratings seem particularly surprising. The examination of the relationship between confidence and accuracy in emotion recognition (i.e., metacognitive sensitivity) revealed, however, that participants with higher autistic trait levels in our study were less able to scale their confidence according to their actual performance. Previous research on alterations in metacognitive judgments in ASD has already described a complex pattern of both over- and underconfidence in the social-cognitive domain69.
Our hypothesis that higher autistic trait level would result in reduced facial mimicry responses was also not confirmed. Even though we did not explicitly instruct participants to mimic, individuals with higher autistic trait levels seemed to automatically show unaltered facial muscle activation patterns, contradicting findings in clinical populations59 as well as in healthy individuals with high autistic traits62. Importantly, it has been suggested that mimicry in ASD might especially be reduced for shorter presentation durations107 and occur with a delay rather than not at all108, which we did not examine in our study. We did, however, observe a modulation in the link between facial mimicry and emotion recognition by autistic traits. In the recognition of sad expressions, increased activity of the corrugator, indicating mimicry of sadness, was less predictive of accurate recognition whereas the same applied to stronger zygomaticus activity in the recognition of happiness. While a sole evaluation of the latter effect would be difficult due to the ceiling performance in happiness recognition (see previous paragraph) as well as to a lack of reproducibility of a result when using a relative accuracy score (see Supplemental Material S1), the robust results concerning sadness recognition support the presence of a meaningful modulation.
It seems, thus, that facial mimicry plays a less informative role in emotion recognition, at least of sad expressions, in association with higher autistic trait levels. This observation is in line with past research that did not find an effect of automatic, intentional or externally induced mimicry on reports of the participant’s own emotional experience in individuals on the autism spectrum, while neurotypical participants were considerably influenced by mimicry109. According to the idea of the existence of two routes in emotion recognition, a fast one involving proprioceptive (bottom-up) information and a long one involving knowledge-based (top-down) information26, the fast route might have been less employed in the recognition of sadness in participants with higher autistic traits. Since recognition performance of particularly sad expression was less negatively affected by higher autistic traits, judgments via the alternative, long route could have resulted in similarly successful judgments.
Previous studies have already reported qualitative differences in the recognition of sadness compared to other emotion expressions related to ASD. For example, recognition of sadness in static faces compared to point-light-displays has been found to be only improved in individuals with low but not with high autistic traits51. Moreover, while dynamic information (i.e., videos) generally improved emotion recognition for both autistic and neurotypical individuals, individuals on the autism spectrum recognized dynamic sad expressions worse compared to static ones110. Information that facilitates the recognition of sadness in neurotypical individuals might not serve individuals on the autism spectrum in the same way. Why this is specifically the case for sadness should be investigated in future studies.
Taken our findings related to autistic traits together, feedback from multiple sources might not be integrated beneficially in emotion recognition. On the one hand, confidence in emotion recognition does not seem to be scaled to actual performance. Internal feedback, in other words, the “feeling” how well one performed, might not be informative of actual performance in autism and, thus, cannot assist successful learning. Our findings suggest that, on the other hand, a simulation of observed expression might not be as informative for emotion processing in ASD compared to a neurotypical population. This claim is supported by research showing a reduced access to bodily signals (i.e., interoceptive accuracy) next to a heightened sensitivity to those signals in autism111, which seems to be driven by comorbid alexithymia112, 113. Consequently, while interventions targeting metacognitive abilities could help overcome the gap between actual performance and subjective judgments in individuals on the autism spectrum, a training focusing on the integration of information from the bodily component of an emotional experience could indirectly benefit emotion recognition and other social skills.
In addition to the results specific to the trait dimensions, our findings also add to the current discussion on the general role of facial mimicry in emotion recognition. Recent meta-analyses have described no robust relationship between facial mimicry and emotion recognition28, as well as broader affective judgments29. Our study, in contrast, revealed a link between facial mimicry responses to happy and sad expressions and associated recognition accuracy. More specifically, stronger activation of the zygomaticus and relaxation of the corrugator predicted better recognition of happiness, and stronger activation of the corrugator predicted better recognition of sadness. In some instances, sensorimotor simulation (i.e., activating facial muscle patterns that are congruent to observed emotional facial expressions) might indeed become a relevant mechanism in understanding others’ emotions. Similar to previous literature, the effects were not robust in our study (see relative accuracy analysis in Supplemental Material S1), varied depending on clinical trait levels (see paragraphs above), and we did not observe significant relationships for all expression categories (e.g., not for anger). Our study therefore corroborates evidence that an embodiment of observed emotional expressions does not seem necessary for a successful recognition114. Yet, facial feedback can become informative under certain conditions29, and our results highlight that individual differences should additionally be considered.
Despite our efforts to create a more naturalistic emotion recognition scenario by displaying spontaneous, dynamic facial expressions of emotion, participants still observed standardized stimuli in a controlled lab setting in our study. This limits the generalizability of our results as the interpretation of emotional expressions has been shown to be highly context-dependent115, 116. In contrast to natural scenarios, the same stimuli were also presented repeatedly (three times) in different blocks. While the repeated presentation allowed us to obtain EMG responses without priming participants with emotion category words, learning effects might have occurred. For example, emotion recognition could have been facilitated or expressions could have been perceived as less intense. More importantly, our study did not involve a real social context. Without an interaction partner who receives and responds to expressions from the participant, the social communicative function of emotional expressions, including a bidirectional coordination of affective states3, may get lost117. This limitation might also affect trait dimension-specific modulations in emotion perception. For example, in a real social situation, higher social anxiety levels have been associated with an enhanced mimicry of polite, but not enjoyment smiles16. Furthermore, ASD was argued to specifically become apparent in alterations in interpersonal dynamics (i.e., during bidirectional information exchange)118, 119. Consequently, future studies on emotion perception should be conducted in real social situations that allow for reciprocity and affect coordination.
In addition to that, even though our observations on the impact of trait levels can give us hints with regard to alterations in clinical populations, we still collected data from a non-clinical sample. Once clinical symptoms that have a severe impact on an individual’s life come into play, emotion processing might be altered differently, both qualitatively and quantitatively. Half of the participants in our sample had social anxiety trait levels that are considered clinically relevant (i.e., above 30; see Questionnaires section). While these high social anxiety trait levels for non-diagnosed individuals might result in findings that are comparable to clinical populations, this might be less applicable for our results regarding autistic traits. For example, while sadness recognition was observed to be least impacted by autistic trait levels in our study, a reduced perceptual sensitivity has been specifically described for sad facial expressions in individuals on the autism spectrum120. This emotion-specific recognition impairment has been shown to extend to difficulties in interpreting sadness from animations, which, in turn, has been related to worse daily social functioning in individuals on the autism spectrum121. Thus, in order to provide meaningful insights, results from studies including healthy participants with variations on clinical trait dimensions should always be confirmed in clinical populations as well as related to actual day-to-day experiences.
Moreover, while our sample was not gender-balanced, gender differences in mimicry and its integration in emotion recognition have been reported in past research26, 122, as well as in autistic traits and social anxiety traits90, 123. Given the predominance of female participants in our sample, our findings cannot be easily generalized to the male population. Future studies should therefore examine whether similar effects to the ones described in the current study can be observed in a more balanced or even exclusively male sample. Lastly, as we did neither manipulate facial mimicry nor metacognition, our study does not allow for causality assumptions in their role in emotion recognition. Within an emotion processing context, information is likely to flow bidirectionally and recent findings support a context-dependent influence of emotion recognition on facial mimicry124. Furthermore, a more fine-grained investigation of potential mediatory processes in the course of emotion perception and interpretation, such as the integration of interoceptive information24, 125, might benefit the understanding of variability in emotion processing and enable the formalization of testable theoretical models126.
In conclusion, our study provides evidence for distinct modulations of facial mimicry and metacognitive judgments in emotion recognition by autistic traits and social anxiety traits in a majorly female sample. Higher social anxiety traits were predominantly related to an underconfidence in emotion recognition, despite an unaltered performance, whereas higher autistic traits were associated with an overall worse recognition performance as well as a poorer calibration of performance judgments, and a less pronounced link between facial mimicry and emotion recognition. These trait dimension-specific patterns might also translate to the linked clinical disorders, which, however, still has to be confirmed in future studies. Importantly, particularities in processing others’ emotions have been shown to contribute to social interactions difficulties experienced by individuals on the autism spectrum and by individuals with SAD. Hence, evidence-based interventions targeting condition-specific alterations in distinct components (i.e., metacognitive beliefs and bodily feedback) hold the promise to facilitate daily social encounters and improve the quality of life in the two clinical populations.
Data availability
Following the university policy, all data and code from this project will be made available on the DataverseNL repository upon paper publication (https://doi.org/10.34894/8UBNPL).
References
Frith, C. Role of facial expressions in social interactions. Philos. Trans. R. Soc. B Biol. Sci. 364, 3453–3458 (2009).
Tracy, J. L., Randles, D. & Steckler, C. M. The nonverbal communication of emotions. Curr. Opin. Behav. Sci. 3, 25–30 (2015).
Keltner, D. & Kring, A. M. Emotion, social function, and psychopathology. Rev. Gen. Psychol. 2, 320–342 (1998).
van Kleef, G. A. & Côté, S. The social effects of emotions. Annu. Rev. Psychol. 73, 629–658 (2021).
Künecke, J., Hildebrandt, A., Recio, G., Sommer, W. & Wilhelm, O. Facial EMG responses to emotional expressions are related to emotion perception ability. PLoS One 9, (2014).
Buck, R. Nonverbal behavior and the theory of emotion: The facial feedback hypothesis. J. Pers. Soc. Psychol. 38, 811–824 (1980).
Kelly, K. J. & Metcalfe, J. Metacognition of emotional face recognition. Emotion 11, 896–906 (2011).
Davies, H., Wolz, I., Leppanen, J., Fernandez-aranda, F. & Schmidt, U. Facial expression to emotional stimuli in non-psychotic disorders: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 64, 252–271 (2016).
Rouault, M., Seow, T., Gillan, C. M. & Fleming, S. M. Psychiatric symptom dimensions are associated with dissociable shifts in metacognition but not task performance. Biol. Psychiatry 84, 443–451 (2018).
Trevisan, D. A. & Birmingham, E. Are emotion recognition abilities related to everyday social functioning in ASD? A meta-analysis. Res. Autism. Spectr. Disord. 32, 24–42 (2016).
Gilboa-Schechtman, E. & Shachar-Lavie, I. More than a face: A unified theoretical perspective on nonverbal social cue processing in social anxiety. Front. Hum. Neurosci. 7, 1–14 (2013).
Dimberg, U. Facial reactions to facial expressions. Psychophysiology 19, 643–647 (1982).
Sato, W., Fujimura, T. & Suzuki, N. Enhanced facial EMG activity in response to dynamic facial expressions. Int. J. Psychophysiol. 70, 70–74 (2008).
Rymarczyk, K., Biele, C., Grabowska, A. & Majczynski, H. EMG activity in response to static and dynamic facial expressions. Int. J. Psychophysiol. 79, 330–333 (2011).
Rymarczyk, K., Zurawski, Ł, Jankowiak-Siuda, K. & Szatkowska, I. Do dynamic compared to static facial expressions of happiness and anger reveal enhanced facial mimicry?. PLoS ONE 11, 1–15 (2016).
Dijk, C., Fischer, A. H., Morina, N., Eeuwijk, C. Van & Kleef, G. A. Van. Effects of social anxiety on emotional mimicry and contagion: Feeling negative, but smiling politely. J. Nonverbal. Behav. 42, 81–99 (2018).
Hess, U. & Blairy, S. Facial mimicry and emotional contagion to dynamic emotional facial expressions and their influence on decoding accuracy. Int. J. Psychophysiol. 40, 129–141 (2001).
Olszanowski, M., Wróbel, M. & Hess, U. Mimicking and sharing emotions: A re-examination of the link between facial mimicry and emotional contagion. Cogn. Emot. 34, 367–376 (2020).
Peter-Ruf, C. et al. Emotion regulation in high and low socially anxious individuals: An experimental study investigating emotional mimicry, emotion recognition, and self-reported emotion regulation. J. Depress. Anxiety Disord. 1, 17–26 (2017).
Drimalla, H., Landwehr, N., Hess, U. & Dziobek, I. From face to face: The contribution of facial mimicry to cognitive and emotional empathy. Cogn. Emot. 33, 1672–1686 (2019).
Sato, W., Fujimura, T., Kochiyama, T. & Suzuki, N. Relationships among facial mimicry, emotional experience, and emotion recognition. PLoS ONE 8, 1–8 (2013).
Damasio, A. The somatic marker hypothesis and the possible functions of the prefrontal cortex. Philos. Trans. R. Soc. Lond. Ser. B 351, 1413–1420 (1996).
James, W. What is an emotion?. Mind 9, 188–205 (1884).
Critchley, H. D. & Garfinkel, S. N. Interoception and emotion. Curr. Opin. Psychol. 17, 7–14 (2017).
Wood, A., Rychlowska, M., Korb, S. & Niedenthal, P. Fashioning the face: Sensorimotor simulation contributes to facial expression recognition. Trends Cogn. Sci. 20, 227–240 (2016).
Stel, M. & van Knippenberg, A. The role of facial mimicry in the recognition of affect. Psychol. Sci. 19, 984–986 (2008).
Neal, D. T. & Chartrand, T. L. Embodied emotion perception: Amplifying and dampening facial feedback modulates emotion perception accuracy. Soc. Psychol. Pers. Sci. 2, 673–678 (2011).
Holland, A. C., O’Connell, G. & Dziobek, I. Facial mimicry, empathy, and emotion recognition: A meta-analysis of correlations. Cogn. Emot. 1–19. https://doi.org/10.1080/02699931.2020.1815655 (2020).
Coles, N. A., Larsen, J. T. & Lench, H. C. A meta-analysis of the facial feedback literature: Effects of facial feedback on emotional experience are small and variable. Psychol. Bull. 145, 610–651 (2019).
Bogart, K. R. & Matsumoto, D. Facial mimicry is not necessary to recognize emotion: Facial expression recognition by people with Moebius syndrome. Soc. Neurosci. 5, 241–251 (2010).
Frith, C. The role of metacognition in human social interactions. Philos. Trans. R. Soc. B Biol. Sci. 367, 2213–2223 (2012).
Dentakos, S., Saoud, W., Ackerman, R. & Toplak, M. E. Does domain matter? Monitoring accuracy across domains. Metacogn. Learn. 14, 413–436 (2019).
Bègue, I. et al. Confidence of emotion expression recognition recruits brain regions outside the face perception network. Soc. Cogn. Affect Neurosci. 14, 81–95 (2019).
Montagne, B. et al. Reduced sensitivity in the recognition of anger and disgust in social anxiety disorder. Cogn. Neuropsychiatry 11, 389–401 (2006).
Bui, E. et al. Heightened sensitivity to emotional expressions in generalised anxiety disorder, compared to social anxiety disorder, and controls. Cogn. Emot. 31, 119–126 (2017).
Arrais, K. C. et al. Social anxiety disorder women easily recognize fearful, sad and happy faces: The influence of gender. J. Psychiatr. Res. 44, 535–540 (2010).
Joormann, J. & Gotlib, I. H. Is this happiness i see? Biases in the identification of emotional facial expressions in depression and social phobia. J. Abnorm. Psychol. 115, 705–714 (2006).
Rapee, R. M. & Heimberg, R. G. A cognitive-behavioral model of anxiety in social phobia. Behav. Res. Ther. 35, 741–756 (1997).
Clark, D. M. & Wells, A. A cognitive model of social phobia. in Social phobia: Diagnosis, assessment, and treatment. 69–93 (The Guilford Press, 1995).
Amin, N., Foa, E. B. & Coles, M. E. Negative interpretation bias in social phobia. Behav. Res. Ther. 36, 945–957 (1998).
Hirsch, C. R. & Clark, D. M. Information-processing bias in social phobia. Clin. Psychol. Rev. 24, 799–825 (2004).
Machado-de-Sousa, J. P. et al. Facial affect processing in social anxiety: Tasks and stimuli. J. Neurosci. Methods 193, 1–6 (2010).
Hunter, L. R., Buckner, J. D. & Schimdt, N. B. Interpreting facial expressions: The influence of social anxiety, emotional valence, and race. J. Anxiety Disord. 23, 1–7 (2009).
Richards, A. et al. Anxiety-related bias in the classification of emotionally ambiguous facial expressions. Emotion 2, 273–287 (2002).
Gutiérrez-García, A. & Calvo, M. G. Social anxiety and threat-related interpretation of dynamic facial expressions: Sensitivity and response bias. Pers. Individ. Dif. 107, 10–16 (2017).
Uljarevic, M. & Hamilton, A. Recognition of emotions in autism: A formal meta-analysis. J. Autism Dev. Disord. 43, 1517–1526 (2013).
Frank, R., Schulze, L., Hellweg, R., Koehne, S. & Roepke, S. Impaired detection and differentiation of briefly presented facial emotions in adults with high-functioning autism and asperger syndrome. Behav. Res. Ther. 104, 7–13 (2018).
Sucksmith, E., Allison, C., Baron-Cohen, S., Chakrabarti, B. & Hoekstra, R. A. Empathy and emotion recognition in people with autism, first-degree relatives, and controls. Neuropsychologia 51, 98–105 (2013).
Mazzoni, N., Ricciardelli, P., Actis-Grosso, R. & Venuti, P. Difficulties in recognising dynamic but not static emotional body movements in autism spectrum disorder. J. Autism Dev. Disord. 52, 1092–1105 (2022).
Kret, M. E., Stekelenburg, J. J., de Gelder, B. & Roelofs, K. From face to hand: Attentional bias towards expressive hands in social anxiety. Biol. Psychol. 122, 42–50 (2017).
Actis-Grosso, R., Bossi, F. & Ricciardelli, P. Emotion recognition through static faces and moving bodies: A comparison between typically developed adults and individuals with high level of autistic traits. Front. Psychol. 6, (2015).
Arnaud, S. Emotional consciousness in autism. J. Conscious. Stud. 27, 34–59 (2020).
Corbett, B. A. et al. A functional and structural study of emotion and face processing in children with autism. Psychiatry Res. Neuroimaging 173, 196–205 (2009).
Dijk, C., van Emmerik, A. A. P. & Grasman, R. P. P. P. Social anxiety is related to dominance but not to affiliation as perceived by self and others: A real-life investigation into the psychobiological perspective on social anxiety. Pers. Individ. Dif. 124, 66–70 (2018).
Dimberg, U. Social fear and expressive reactions to social stimuli. Scand. J. Psychol. 38, 171–174 (1997).
Vrana, S. R. & Gross, D. Reactions to facial expressions: Effects of social context and speech anxiety on responses to neutral, anger, and joy expressions. Biol. Psychol. 66, 63–78 (2004).
Dimberg, U. & Thunberg, M. Speech anxiety and rapid emotional reactions to angry and happy facial expressions: Personality and Social Sciences. Scand. J. Psychol. 48, 321–328 (2007).
Trevisan, D. A., Hoskyn, M. & Birmingham, E. Facial expression production in autism: A meta-analysis. Autism Res. 11, 1586–1601 (2018).
McIntosh, D. N., Reichmann-Decker, A., Winkielman, P. & Wilbarger, J. L. When the social mirror breaks: Deficits in automatic, but not voluntary, mimicry of emotional facial expressions in autism. Dev. Sci. 9, 295–302 (2006).
Rozga, A., King, T. Z., Vuduc, R. W. & Robins, D. L. Undifferentiated facial electromyography responses to dynamic, audio-visual emotion displays in individuals with autism spectrum disorders. Dev. Sci. 16, 499–514 (2013).
Weiss, E. M., Rominger, C., Hofer, E., Fink, A. & Papousek, I. Less differentiated facial responses to naturalistic films of another person’s emotional expressions in adolescents and adults with High-Functioning Autism Spectrum Disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 89, 341–346 (2019).
Hermans, E. J., van Wingen, G., Bos, P. A., Putman, P. & van Honk, J. Reduced spontaneous facial mimicry in women with autistic traits. Biol. Psychol. 80, 348–353 (2009).
Tan, L., Ford, R., Morrison, A. & Stagg, S. ASD traits predict attenuated social mimicry in a subclinical sample. (2020).
Nordahl, H., Ødegaard, I. H., Hjemdal, O. & Wells, A. A test of the goodness of fit of the generic metacognitive model of psychopathology symptoms. BMC Psychiatry 19, 1–9 (2019).
Hartman, L. M. A metacognitive model of social anxiety: Implications for treatment. Clin. Psychol. Rev. 3, 435–456 (1983).
Müller-Pinzler, L. et al. Negativity-bias in forming beliefs about own abilities. Sci. Rep. 9, 1–15 (2019).
Sawyer, A. C. P., Williamson, P. & Young, R. Metacognitive processes in emotion recognition: Are they different in adults with Asperger’s disorder?. J. Autism Dev. Disord. 44, 1373–1382 (2014).
Wang, S. & Adolphs, R. Reduced specificity in emotion judgment in people with autism spectrum disorder. Neuropsychologia 99, 286–295 (2017).
DeBrabander, K. M., Pinkham, A. E., Ackerman, R. A., Jones, D. R. & Sasson, N. J. Cognitive and social cognitive self-assessment in autistic adults. J. Autism Dev. Disord. 51, 2354–2368 (2020).
Fleming, S. M. & Lau, H. C. How to measure metacognition. Front. Hum. Neurosci. 8, 1–9 (2014).
Zwick, J. C. & Wolkenstein, L. Facial emotion recognition, theory of mind and the role of facial mimicry in depression. J. Affect. Disord. 210, 90–99 (2017).
Westfall, J. PANGEA: Power ANalysis for GEneral Anova designs. (2015).
Altman, D. G. & Royston, P. The cost of dichotomising continuous variables. Br. Med. J. 332, 1080 (2006).
Wallhoff, F., Schuller, B., Hawellek, M. & Rigoll, G. Efficient Recognition of Authentic Dynamic Facial Expressions on the Feedtum Database. in 2006 IEEE International Conference on Multimedia and Expo 493–496. https://doi.org/10.1109/ICME.2006.262433 (2006).
Noldus. FaceReader: Tool for automatic analysis of facial expression. (2014).
Christiansen, M. Adobe after effects CC visual effects and compositing studio techniques. (2018).
Psychology Software Tools. E-Prime. (2016).
Nummenmaa, L. & Calvo, M. G. Dissociation between recognition and detection advantage for facial expressions: A meta-analysis. Emotion 15, 1–14 (2015).
Calvo, M. G., Gutiérrez-García, A., Fernández-Martín, A. & Nummenmaa, L. Recognition of facial expressions of emotion is related to their frequency in everyday life. J. Nonverbal. Behav. 38, 549–567 (2014).
Liebowitz, M. R. Social phobia. Mod. Probl. Pharmacopsy. 22, 141–173 (1987).
Baron-Cohen, S., Wheelwright, S., Skinner, R., Martin, J. & Clubley, E. The autism spectrum quotient (AQ): Evidence from asperger syndrome/high-functioning autism, males and females, scientists and mathematicians. J Autism Dev Disord 31, 5–17 (2001).
Fridlund, A. J. & Cacioppo, J. T. Guidelines for human electromyographic research. Psychophysiology 23, 567–589 (1986).
Biopac Systems Inc. AcqKnowledge 4.0 software guide. Retrieved April 20, 2023 from https://www.biopac.com/wp-content/uploads/acqknowledge-4-software-guide.pdf. (2009).
Sjak-Shie, E. E. PhysioData Toolbox (Version 0.5) [Computer software]. Retrieved from https://PhysioDataToolbox.leidenuniv.nl (2019).
Dignath, D., Berger, A., Spruit, I. M. & van Steenbergen, H. Temporal dynamics of error-related corrugator supercilii and zygomaticus major activity: Evidence for implicit emotion regulation following errors. Int. J. Psychophysiol. 146, 208–216 (2019).
van Buuren, S. & Groothuis-Oudshoorn, K. mice: Multivariate imputation by chained equations in R. J. Stat. Softw. 45, 1–67 (2011).
Mennin, D. S. et al. Screening for social anxiety disorder in the clinical setting: Using the Liebowitz Social Anxiety Scale. J. Anxiety Disord. 16, 661–673 (2002).
Rytwinski, N. K. et al. Screening for social anxiety disorder with the self-report version of the liebowitz social anxiety scale. Depress Anxiety 26, 34–38 (2009).
Fresco, D. M. et al. The Liebowitz Social Anxiety Scale: A comparison of the psychometric properties of self-report and clinician-administered formats. Psychol. Med. 31, 1025–1035 (2001).
Ruzich, E. et al. Measuring autistic traits in the general population: A systematic review of the Autism-Spectrum Quotient (AQ) in a nonclinical population sample of 6900 typical adult males and females. Mol. Autism 6, (2015).
Keating, C. T., Fraser, D. S., Sowden, S. & Cook, J. L. Differences between autistic and non-autistic adults in the recognition of anger from facial motion remain after controlling for alexithymia. J. Autism Dev. Disord. https://doi.org/10.1007/s10803-021-05083-9 (2021).
R Core Team. R: A language and environment for statistical computing. https://www.R-project.org/ (2020).
Kuznetsova, A., Brockhoff, P. B. & Christensen, R. H. B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 82, 1–26 (2017).
Hothorn, T., Bretz, F. & Westfall, P. Simultaneous inference in general parametric models. Biom. J. 50, 346–363 (2008).
Lüdecke, D. sjPlot: Data visualization for statistics in social science. https://CRAN.R-project.org/package=sjPlot.
Wickham, H. ggplot2: Elegant graphics for data analysis (Springer-Verlag, 2016).
Staugaard, S. R. Threatening faces and social anxiety: A literature review. Clin. Psychol. Rev. 30, 669–690 (2010).
Heimberg, R. G., Brozovich, F. A. & Rapee, R. M. A cognitive behavioral model of social anxiety disorder: update and extension. in Social Anxiety (Second Edition) (eds. Hofmann, S. G. & DiBartolo, P. M.) 395–422 (Academic Press, 2010). https://doi.org/10.1016/B978-0-12-375096-9.00015-8.
Dijk, C., Voncken, M. J. & de Jong, P. J. I blush, therefore I will be judged negatively: influence of false blush feedback on anticipated others’ judgments and facial coloration in high and low blushing-fearfuls. Behav. Res. Ther. 47, 541–547 (2009).
Kashdan, T. B. & Savostyanova, A. A. Capturing the biases of socially anxious people by addressing partner effects and situational parameters. Behav. Ther. 42, 211–233 (2011).
Voncken, M. J. & Bögels, S. M. Social performance deficits in social anxiety disorder: Reality during conversation and biased perception during speech. J. Anxiety Disord. 22, 1384–1392 (2008).
Gkika, S., Wittkowski, A. & Wells, A. Social cognition and metacognition in social anxiety: A systematic review. Clin. Psychol. Psychother. 25, 10–30 (2018).
Dannahy, L. & Stopa, L. Post-event processing in social anxiety. Behav. Res. Ther. 45, 1207–1219 (2007).
Helbig-Lang, S., von Auer, M., Neubauer, K., Murray, E. & Gerlach, A. L. Post-event processing in social anxiety disorder after real-life social situations—An ambulatory assessment study. Behav. Res. Ther. 84, 27–34 (2016).
Nordahl, H. & Wells, A. Metacognitive therapy for social anxiety disorder: An A-B replication series across social anxiety subtypes. Front. Psychol. 9, 1–7 (2018).
Green, C. & Guo, K. Factors contributing to individual differences in facial expression categorisation. Cogn. Emot. 32, 37–48 (2018).
Mathersul, D., McDonald, S. & Rushby, J. A. Automatic facial responses to briefly presented emotional stimuli in autism spectrum disorder. Biol. Psychol. 94, 397–407 (2013).
Oberman, L. M., Winkielman, P. & Ramachandran, V. S. Slow echo: Facial EMG evidence for the delay of spontaneous, but not voluntary, emotional mimicry in children with autism spectrum disorders. Dev. Sci. 12, 510–520 (2009).
Stel, M., van den Heuvel, C. & Smeets, R. C. Facial feedback mechanisms in autistic spectrum disorders. J. Autism. Dev. Disord. 38, 1250–1258 (2008).
Enticott, P. G. et al. Emotion recognition of static and dynamic faces in autism spectrum disorder. Cogn. Emot. 28, 1110–1118 (2014).
Garfinkel, S. N. et al. Discrepancies between dimensions of interoception in autism: Implications for emotion and anxiety. Biol. Psychol. 114, 117–126 (2016).
Ketelaars, M. P., In’T Velt, A., Mol, A., Swaab, H. & van Rijn, S. Emotion recognition and alexithymia in high functioning females with autism spectrum disorder. Res. Autism. Spectr. Disord. 21, 51–60 (2016).
Shah, P., Hall, R., Catmur, C. & Bird, G. Alexithymia, not autism, is associated with impaired interoception. Cortex 81, 215–220 (2016).
Folz, J., Fiacchino, D., Nikolić, M., van Steenbergen, H. & Kret, M. E. Reading your emotions in my physiology? reliable emotion interpretations in absence of a robust physiological resonance. Affect Sci. 3, 480–497 (2022).
Hess, U. & Fischer, A. Emotional mimicry as social regulation. Pers. Soc. Psychol. Rev. 17, 142–157 (2013).
Israelashvili, J., Aviezer, H. & Hassin, R. When emotions run high: A critical role for context in the unfolding of dynamic. Real-Life Facial Affect. Emotion 19, 558–562 (2019).
Schilbach, L. et al. Toward a second-person neuroscience. Behav. Brain Sci. 36, 393–414 (2013).
Peper, C. E., van der Wal, S. J. & Begeer, S. Autism in action: Reduced bodily connectedness during social interactions?. Front. Psychol. 7, 1–6 (2016).
Bolis, D., Balsters, J., Wenderoth, N., Becchio, C. & Schilbach, L. Beyond Autism: Introducing the dialectical misattunement hypothesis and a Bayesian account of intersubjectivity. Psychopathology 50, 355–372 (2018).
Wallace, G. L. et al. Diminished sensitivity to sad facial expressions in high functioning autism spectrum disorders is associated with symptomatology and adaptive functioning. J. Autism Dev. Disord. 41, 1475–1486 (2011).
Boraston, Z., Blakemore, S. J., Chilvers, R. & Skuse, D. Impaired sadness recognition is linked to social interaction deficit in autism. Neuropsychologia 45, 1501–1510 (2007).
Dimberg, U. & Lundquist, L. O. Gender differences in facial reactions to facial expressions. Biol. Psychol. 30, 151–159 (1990).
Caballo, V. E. et al. Differences in social anxiety between men and women across 18 countries. Pers. Individ. Dif. 64, 35–40 (2014).
Kastendieck, T., Mauersberger, H., Blaison, C., Ghalib, J. & Hess, U. Laughing at funerals and frowning at weddings: Top-down influences of context-driven social judgments on emotional mimicry. Acta Psychol. (Amst) 212, (2021).
Arnold, A. J., Winkielman, P. & Dobkins, K. Interoception and social connection. Front. Psychol. 10, 1–6 (2019).
Smith, R., Parr, T. & Friston, K. J. Simulating emotions: An active inference model of emotional state inference and emotion concept learning. Front. Psychol. 10, 1–24 (2019).
Funding
This research was funded by the Dutch National Science Foundation (Nederlandse Organisatie voor Wetenschappelijk Onderzoek, NWO) [grant number 016.VIDI.185.036].
Author information
Authors and Affiliations
Contributions
J.F.: conceptualization, methodology, software, formal analysis, data curation, writing—original draft, visualization, project administration. M.N.: methodology, writing—review and editing, supervision. R.A.: conceptualization, methodology, writing—original draft, Iivestigation. H.v.S.: methodology, formal analysis, writing—review and editing. M.E.K.: conceptualization, methodology, resources, writing—review and editing, supervision, funding acquisition.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Folz, J., Akdağ, R., Nikolić, M. et al. Facial mimicry and metacognitive judgments in emotion recognition are distinctly modulated by social anxiety and autistic traits. Sci Rep 13, 9730 (2023). https://doi.org/10.1038/s41598-023-35773-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-35773-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
