
Abstract
Under the archaeological canine surrogacy approach (CSA) it is assumed that because dogs were reliant on humans for food, they had similar diets to the people with whom they lived. As a result, the stable isotope ratios of their tissues (bone collagen and apatite, tooth enamel and dentine collagen) will be close to those of the humans with whom they cohabited. Therefore, in the absence of human tissue, dog tissue isotopes can be used to help reconstruct past human diets. Here δ13C and δ15N ratios on previously published dog and human bone collagen from fourteenth-seventeenth century AD ancestral Iroquoian village archaeological sites and ossuaries in southern Ontario are used with MixSIAR, a Bayesian dietary mixing model, to determine if the dog stable isotope ratios are good proxies for human isotope ratios in dietary modeling for this context. The modeling results indicate that human dietary protein came primarily from maize and high trophic level fish and dogs from maize, terrestrial animals, low trophic level fish, and human feces. While isotopes from dog tissues can be used as general analogs for human tissue isotopes under CSA, greater insights into dog diets can be achieved with Bayesian dietary mixing models.
Similar content being viewed by others

Introduction
Multiple lines of archaeological evidence are used to reconstruct the diets of past people. These can include macrobotanical, microbotanical, and animal bone assemblages; lipids and proteins extracted from pottery fabric and charred encrusted cooking residues; stable isotope analyses of cooking residues, specific compounds extracted from residues and pottery fabrics, and human tissue (generally bone collagen and apatite and/or tooth dentine collagen and enamel). The last of these can be among the most important because human tissues provide direct evidence of resource consumption: “you are what you eat”1. However, in many regions this is not possible because of the lack of suitable remains, objections from descendant communities to destructive analyses of human tissue, and laws that require the repatriation of human remains to descendant communities. One way that archaeologists have responded to the lack of human remains is to analyze dog tissue. This is done under the assumption that dogs relied on humans for much their food and, therefore, stable isotopic ratios of dog tissue may be close to the tissues of human with whom they lived, depending on environmental, bio-behavioral, and cultural factors (2, p. 362). This has been labeled the canine surrogacy approach (CSA;2) and has been applied in several studies in northeastern North America (e.g.,3,4,5,6) as well as other regions of the continent (e.g.,7,8). These studies have either used dogs as direct analogs for human diets or as indirect analogs to assess the extent to which maize, a C4 plant in an otherwise C3 plant biome, was a significant food resource2.
Recently, Glencross and colleagues9 used new δ13C and δ15N ratios on large series of dog bone collagen from Iroquoian village sites in southern Ontario to test their applicability as direct analogs for reconstruction of fourteenth–seventeenth century AD human diets by comparing them to previously published ratios on human bone collagen from the same village site or associated ossuary. They conclude that δ13C ratios are good proxies for human ratios. While dog δ15N ratios are < 2‰ lower than human ratios, they “generally support the premise that dogs can serve as dietary proxies for humans, any approximations would likely underestimate the actual human values” (9, p. 12). However, these results do not directly address the issue of whether dog and human diets were similar enough that stable isotope ratios of dog bone are suitable direct analogs for humans. While the values are similar, dogs are not people and have different consumption patterns (e.g., caecotrophy), the possibilities of which Glencross and colleagues (9, pp. 7, 12) mention and is recognized in the broader CSA literature2 While this analysis resulted in important insights into human and dog diets in the region, it is possible to move beyond the typical applications of CSA by using Bayesian dietary mixing models to gain more detailed dietary estimates for dogs as isotopic analogs for associated humans.
Used extensively in other disciplines (e.g.,10,11,12,13) and in archaeological human and dog dietary studies globally (e.g.,8,14,15,16,17,18) but rarely in northeastern North American archaeology19,20,21, Bayesian dietary mixing models can be used with multiple tracers (e.g., stable isotope ratios) of consumers and potential resources (sources), taking into account trophic enrichment factors (TEF), and other sources of prior information, to estimate dietary fractions of specific resources and resource groups. Here I use MixSIAR22, which uses a Markov Chain Monte Carlo simulation to produce models of probable food source fractions in consumers’ diets. Included in the simulations are isotopic uncertainties of sources, consumers, and TEFs. This results in probabilities of resource fractions in consumer diets, allowing for more informed assessments of diets than those based on single isotopes or two isotope scatterplots as are typically used in CSA. I compare modeled protein sources in the diets of humans and dogs by century and specific village sites and, where appropriate, associated ossuaries. In the models I use large datasets of source and human and dog isotopic ratios on bone collagen extracted from the literature; collagen isotopes reflect primarily the protein fraction of diets23.
Contemporary free-ranging dog diets
Globally, most contemporary dogs are not household pets24. Rather, they are associated with households but range freely within and beyond settlements, relying primarily on human-sourced food for sustenance. This includes direct feeding by humans and opportunistic scavenging from garbage dumps and other human-derived waste, and sheltering in human structures (e.g.,24, p. 147). Based on a series of experiments with free-ranging dogs in India, Sarkar and colleagues (25, p. 43) found that foraging free-ranging dogs preferred protein-rich over carbohydrate-rich foods and followed a “rule of thumb” that “if it smells like meat, eat it.” Butler and colleagues26 report on the diets of free-ranging dogs in remote agricultural communities of northwestern Zimbabwe. Dogs are associated with specific households but are unrestrained and roam widely. The primary component of human diets in the region is sadza a porridge composed primarily of maize (Zea mays ssp. mays) but which may include other grains such as millet (Pennisetum americanum), sorghum (Sorghum bicolor), or finger millet (Eleusine coracana), and is also fed to dogs. Uneaten food and other waste are disposed of in open pits external to houses, which are accessible to dogs. Most households lack plumbing or latrines, and people defecate in the open away from their houses, shallowly burying their feces, and so are accessible by dogs. Resource frequency occurrence in 945 analyzed dog scats (26, p. 6) were: sadza (87.9%), mammal remains (81.3%), vegetables and fruit (69.8%), and human feces (56.2%). Observation of 689 meals recorded consumption of animal remains in 48.8%, sadza in 22.1%, and human feces in 20.5% (26, p. 8). In total, 13% of dog diets were directly fed by humans, while the remainder was obtained by foraging resources ultimately derived from human activities. Similar results have been found for free-ranging dogs in other regions (e.g.,27,28), with a primary dietary emphasis on animal carcasses, human feces, and grains.
Evidence for Iroquoian dog and human diets
Very little evidence for Iroquoian dog behavior and consumption patterns is provided in the 17th-century southern Ontario ethnohistorical record. Relevant to current purposes is a brief account by Sagard (29, p. 226) suggestive of contemporary free-ranging dog dietary behavior: “their most usual fare is nothing but the refuse they find in the streets and on the roads. They also very frequently put their pointed nose into the savages’ pot of sagamité.” Sagamité, like sadza in Zimbabwe, was a maize-based stew or porridge into which was added fish or terrestrial animal tissue and/or other crops such as common bean (Phaseolus vulgaris), squash (Cucurbita pepo), and sunflower (Helianthus annuus)30,31.
Maize is generally the most ubiquitous and abundant food-related macrobotanical remain recovered from ancestral Iroquoian archaeological sites (e.g.32). Isotopic analysis of fourteenth-seventeenth century ancestral Iroquoian bone and teeth indicate the importance of maize in diets; maize accounting for > 50% to > 70% of consumed resources (e.g.20,33). Freshwater fish remains are common on ancestral Iroquoian archaeological sites in southern Ontario (e.g.34), and isotopic analyses of human tissue confirms the importance of fish in human diets20,33,35. While mammal and bird bone are common on ancestral Iroquoian sites in southern Ontario (e.g.,36,37), isotopic analyses suggest terrestrial animal resources did not contribute substantially to human diets20.
Results
The CSA literature indicates dog collagen δ15N ratios are typically 2–3‰ lower than human ratios, although the differences between the two species vary widely (2, pp. 354–356); Glencross and colleagues (9, p. 12) indicate a mean difference of < 2.0‰ in δ15N ratios between Iroquoian dogs and humans. Nonparametric Mann–Whitney tests of dog and human δ15N ratios by century have significant differences in sample medians and the nonparametric Epps-Singleton test indicates the sample distributions are significantly different (Table 1). This suggests that the isotope ratios potentially reflect different diets. While caceotrophy has been suggested as a possible cause of the lower δ15N ratios in dogs2, it is also possible based on what is known about free-range dog dietary behavior that differences may result from consumption of different resources than those comprising human diets or the same resources but in different proportions. The MixSIAR results indicate that both are likely causes. Results of the models are presented in Tables 2, 3, Figs. 2, 3, 4, 5, and Supplementary Data S1 Tables 1, 2, 3, 4, 5, 6, online. Model diagnostics are also provided in Supplementary Data S1, online.
Century models
Site locations are provided in Fig. 1. Consistent with previous modeling20, source dietary fraction estimates for humans indicate maize and high and medium δ15N fish were the primary contributors of dietary protein in each century (Fig. 2, Tables 2, S1 Table 1); high and medium δ15N fish representing high trophic-level fish. Maize dietary protein medians range from 0.461 to 0.508 (Table 2). There is a strong negative correlation (> 0.70) between high and medium δ15N fish in each of the models, indicating that the human models cannot cleanly distinguish between these resources (38, p. 294). In these cases, the resources may be under- or overestimated relative to one another38. The medians for these sources range from 0.181 to 0.308 and 0.117 to 0.132, respectively (Table 2). Terrestrial resource medians range from 0.054 to 0.077 (Table 2).

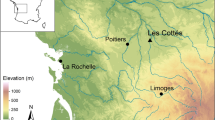
General locations of sites with dog and/or human bone collagen isotope data used in the models. 1. Christian Island, 2. Maurice Ossuary, 3. Warminster/Cahiague, 4. Ossossané Village/Ossuary, 5. Ball, 6. Carson, 7. Kelly-Campbell, 8. Milne, 9. Uxbridge Ossuary, 10. Mantle, 11. Hidden Springs, 12. Teston, 13. New, 14. Fairty Ossuary, 15. Kleinburg Ossuary, 16. Robb, 17. Staines Ossuary, 18. Seed-Barker, 19. Crawford Lake, 20. Rife, 21. Pipeline, 22. Bogle 2, 23. Hamilton, 24. Cleveland, 25. Fonger, 26. Walker, 27. Thorold, 28. Slack-Caswell.

Model 1 dietary fractions of by century. Whiskers are the 2.5% and 97.5 quantiles, the box represents the 25%, 50% (median) and 75% quantiles.
The human results contrast with Model 1 for dogs, which uses the same sources as the human models (Fig. 2, Table 2, S1 Table 1, online). As in the human models, in all dog models, maize is estimated to be the highest contributor of dietary protein in dog diets, and estimated fractions for dogs are close to those for humans. However, unlike the human models, terrestrial animals are consistently estimated to be the second largest contributor to dog dietary protein with low δ15N fish being the third largest contributor; low δ15N fish representing lower trophic-level fish than high and medium δ15N fish. In all the models there are strong negative correlations between terrestrial animals and low δ15N fish indicating they may be under- or overestimated relative to one another. Regardless, dietary fraction medians for these two sources together range from 0.285 to 0.363 and 0.50 to 0.167, respectively, which contrasts with combined human medians 0.037 to 0.050 and 0.054 to 0.077, respectively. High and medium δ15N fish are estimated to have contributed substantially lower dietary protein fractions to dogs (medians = 0.013–0.041) than in the human models. Assuming dogs only ate what humans ate, then, the models indicate substantially different dietary protein fractions for all resources other than maize, with dogs more reliant on terrestrial animal and low δ15N fish tissue and humans more reliant on high and medium δ15N fish tissue.
Because caceotrophy is a dog behavioral trait, dog Model 2 included isotope and concentration estimates for human feces (Table 3, S1 Table 2, online). Maize and terrestrial animals continue to be the primary sources of dietary protein. Human feces are estimated to be the fourth highest contributor of protein to dog diets after maize, terrestrial animals, and low δ15N fish. Estimates for low δ15N fish continue to be higher than medium and high δ15N fish individually and together. There are strong negative correlations between low δ15N fish and terrestrial animals in models and in the 16th-century model between maize and human feces, indicating these source pairs may be under- or overestimated relative to one another.
Glencross et al. (9, p. 7) suggest that micromammal (mice shrews, voles) consumption may account, in part, for the high δ13C ratios in dogs, with dogs hunting and consuming rodents that fed on maize stores. To assess this possibility, dog Model 3 includes micromammals with high δ13C ratios (> − 17.00‰) taken from37. This model has little effect on overall estimated resource fractions, with the estimate medians for micromammals ranging from 0.010 to 0.027 (Fig. 3, Table 1, S1 Table 3, online).

Model 3 dietary fractions of by century.
Site specific models
Models for humans and dogs from the same village sites (Ball and Kelly-Campbell) or ossuaries for humans and village sites for dogs (Fairty Ossuary-Robb village; Kleinberg Ossuary-Seed-Barker village; Ossossané ossuary-Ossossané village) following Glencross and colleagues9 were run using the same sources. Results are presented in Figs. 4 and 5, Table 3, and S1 Tables 4, 5, 6, online. In each of the human models, maize is the highest estimated source of dietary protein, followed by high and medium δ15N fish (Fig. 4, Table 3, S1 Table 4, online).

Model 1 dietary fractions of by sites.

Model 3 dietary fractions of by sites.
In dog Model 1 maize is estimated to be the highest contributor of dietary protein followed by terrestrial prey and low δ15N fish (Fig. 4, Table 3, S1 Table 4, online). In each model the latter two sources are highly negatively correlated and so may be under- or overestimated to one another. This ranking continues in Model 2 (Table 3, S1 Table 5, online), with human feces ranking as the fourth highest estimated source of dietary protein. In the Ossossané model maize and human feces are strongly negatively correlated and so may be under- or overestimated relatively to one another. Model 3 results in human feces as the fourth-ranked estimated source of dietary protein for each site (Fig. 5, Table 3, S1 Table 6, online). Dietary fraction estimate medians for micromammals range from 0.029 to 0.053.
In sum, while there is variation in dietary fraction between centuries and sites, the primary modeled sources of protein in human diets were maize and high and medium δ15N fish. The primary sources of protein in dog diets were maize and terrestrial animals followed by low δ15N fish and human feces.
Discussion and conclusions
The primary assumption of CSA is that stable isotope ratios obtained on dog tissues reflect diets that are reasonably close to the diets of humans with whom the dogs lived. Glencross and associates9 recently performed the first CSA analysis for ancestral Iroquoian sites in southern Ontario by comparing δ15N and δ13C ratios of dog tissues with those of human tissues from related contexts. They conclude that despite small differences in isotopic ratios between dogs and humans, that dog tissue δ13C and δ15N ratios are reasonable analogs for ancestral fourteenth through seventeenth century human ratios.
Few CSA studies in North America have moved beyond direct comparison of dog and human δ13C and δ15N ratios; with few exceptions (e.g.,8), Bayesian dietary mixing models have not been performed. Here I have used dog and human δ13C and δ15N ratios extracted from the literature, including those recently published by Glencross and associates9, in Bayesian dietary mixing models to obtain estimates of the sources of protein for ancestral Iroquoians and their dogs. The results indicate variation in δ13C and δ15N ratios between humans and dogs reflect dietary differences. Human dietary protein derived primarily from maize and high and medium δ15N fish. Dog dietary protein, on the other hand, derived primarily from maize and terrestrial animals, with contributions from low δ15N fish and human feces.
Maize was the primary source of calories for ancestral Iroquoian populations in southern Ontario as reflected in isotopic analyses of human tissues20,33,35 and sixteenth and seventeenth-century AD ethnohistorical accounts29,39,40. Ethnohistorical accounts indicate that a common dish was sagamité a stew/soup with a maize base to which other foods were added. The comment by Sagard29 on dogs eating directly from human-held pots of sagamité indicates that the animals were accustomed to eating food prepared for human consumption. This suggests that dogs were directly fed portions of their diets by their associated humans as is typical for free-ranging dogs. It is also possible that dogs also obtained maize from food discarded by people as in contemporary rural Zimbabwe communities.
Free-ranging dogs obtain large proportions of food from opportunistic scavenging, including the carcasses of animals away from settlements and tissue scraps from human-processed carcasses. This was apparently the case for Iroquoian dogs given that terrestrial animals were consistently the second ranked estimated source of dietary protein. Given that terrestrial animals were not a primary source of dietary protein for ancestral Iroquoian individuals it is likely that dogs obtained this source through scavenging carcasses away from villages. While fish were important in both human and dog diets, humans consumed primarily high trophic-level fish (high and medium δ15N fish), while dogs consumed lower trophic-level fish (low δ15N fish). This is consistent with ethnohistorical and ethnographic records for northeastern North America, in which humans fed large proportions of harvested fish to dogs with a preference for human consumption of large fatty fish rather than small, leaner fish, which were fed to dogs41. Although observations for this pattern are not evident in the Iroquoian ethnohistorical record, it is a possible explanation for the differences in the model estimates. Another source of protein in dog diets was human feces. On average humans produce 29 g of fecal solids each day (42, p. 1854). Given that fourteenth−sixteenth century AD Iroquoian villages in southern Ontario housed hundreds to over 1000 individuals43, human feces were a potentially large source of dietary protein for dogs. Depending on disposal patterns, free-ranging dogs would have had ample opportunities to consume human feces. This was evidently the case given that the fourteenth-sixteenth-century models estimated median fractions range from 0.042 to 0.097 and the site-specific models estimated median fractions ranging from 0.085 to 0.124.
In conclusion, results of the Bayesian dietary mixing models indicate that dog diets were not the same as ancestral Iroquoian diets. This is consistent with what is known about contemporary free-ranging dog dietary behaviors; dogs are reliant on human-sourced food, but do not have the same diets as humans. Despite its limitations, CSA can provide important insights into human diets as has been demonstrated in numerous applications. Of particular note in eastern North America is the use of CSA to assess the presence of maize in dog diets as a potential indicator of the extent of its consumption by humans3,6,44. However, as the current analysis demonstrates, moving beyond standard approaches to CSA through the application of Bayesian dietary mixing models has the potential to provide more detailed assessments of dog diets in given environmental and cultural settings. To accomplish this stable isotope ratios must be available for a range of potential food sources as in the current study.
Methods and materials
Bayesian dietary mixing models were done with MixSIAR version 3.1.1122 in R version 4.1.2 within R Studio version 2022.12.0 + 35 using the default MixSIAR parameters. An example model code is provided in Supplemental Data S2, online. Isotopic data for humans and dogs are δ13C and δ15N ratios measured on bone collagen obtained from the literature (S2 Tables 1 and 2, online). Collagen δ13C ratios primarily reflect the protein fraction of diets, while bone apatite and tooth enamel reflect the whole diet23. Therefore, dietary sources that were likely to have contributed substantively to human and dog dietary protein were used in the models. Complementary dog and human bone apatite and tooth enamel isotope ratio datasets are not available to model whole diets. Models were run for humans and dogs grouped by century and by specific site pairs. Site pairs were those used in9.
Samples sizes by century were: fourteenth century humans = 23, dogs = 13; fifteenth century humans = 22, dogs = 19; 16th century humans = 20, dogs = 21; seventeenth century humans = 16, dogs = 41. Human and dog samples from specific sites/site pairs were modeled separately. Sample sizes for site pairs were: Fairty Ossuary humans = 8, Robb Village dogs = 9; Kleinberg Ossuary humans = 12, Seed-Barker Village dogs = 5; Ball Village humans = 6, dogs = 12; Kelly-Campbell Village humans = 6, dogs = 8; Ossossoané Ossuary humans = 5, Ossossoané Village dogs = 11.
C and N concentrations for representative resources (S2 Table 7, online) were calculated with the following formulas:
Data for these calculations were obtained from the USDA FoodData Central database: https://fdc.nal.usda.gov/. C and N percentages for feces were calculated from26.
A large dataset of δ13C and δ15N ratios obtained on collagen from terrestrial animal and fish bone recovered at Iroquoian sites in southern Ontario previously compiled from the literature20, was used as sources in the Bayesian models (S2 Tables 3 and 4, online). Isotopic data for micromammals (mice, shrews, voles) with high δ13C ratios (> − 17‰) from Iroquoian sites in southern Ontario (S2 Table 5, online) were obtained from37. Following20 fish were separated into high, medium, and low δ15N statistically significant clusters reflecting trophic levels. High δ15N fish contain offshore species, while medium and low δ15N fish contain in-shore species (20, p. 7). These clusters were in the current modeling.
Given that free-ranging dogs are known to eat human feces, δ13C and δ15N estimates for human feces were used in dog models 2 and 3. Isotope ratios for human feces were estimated based on a controlled human dietary study47. This included a diet with animal tissue derived from fish48. Mean offsets between the fish and vegetable diet and feces were + 1.1 for δ13C and + 0.4 for δ15N (47, p. 393). To obtain δ13C and δ15N estimates for human feces, the TEF (5 for δ13C, 3 for δ15N) was subtracted from and then offsets from47 were added to each of the human ratios (S2 Table 1, online). Maize δ13C and δ15N were taken from49. Source means and concentration values are provided in S2 Table 6, online. Collagen (source) to collagen (consumer) TEFs used in the models were + 1.1‰ ± 0.2‰ for δ13C and + 3.8‰ ± 0.2‰ for δ15N; for maize and human feces the TEFs were 5‰ ± 0.1‰ for δ13C and + 3.0‰ ± 0.1‰ for δ15N21,50 (S2 Table 8, online). Mann–Whitney and Epps-Singleton statistical tests were done in PAST version 4.1151.
Models for humans included maize, terrestrial prey, and low, medium, and high δ15N fish. Three models were run for dogs. Model 1 included the same five sources as in the human models, Model 2 added human feces, and Model 3 added high δ13C micromammals per37.
Data availability
All data generated or analyzed during this study are included in this published article (and its Supplementary Information files).
References
Kohn, M. J. You are what you eat. Science 283, 335–336. https://doi.org/10.1126/science.283.5400.335 (1999).
Guiry, E. J. Dogs as analogs in stable isotope-based human paleodietary reconstructions: A review and considerations for future use. J. Archaeol. Method Theory 19, 351–376. https://doi.org/10.1007/s10816-011-9118-z (2012).
Allitt, S., Stewart, R. M. & Messner, T. The utility of dog bone (Canis familiaris) in stable isotope studies for investigating the presence of prehistoric maize (Zea mays ssp. mays): A preliminary study. N. Am. Archaeol. 29, 343–367. https://doi.org/10.2190/NA.29.3-4.h (2008).
Guiry, E. J. & Grimes, V. Domestic dog (Canis familiaris) diets among coastal Late Archaic groups of northeastern North America: A case study for the canine surrogacy approach. J. Anthropol. Archaeol. 32, 732–745. https://doi.org/10.1016/j.jaa.2013.04.005 (2013).
Tankersley, K. B. & Koster, J. M. Sources of stable isotope variation in archaeological dog remains. N. Am. Archaeol. 30, 361–375. https://doi.org/10.2190/NA.30.4.b (2009).
Edwards, R. W. IV., Jeske, R. J. & Coltrain, J. B. Preliminary evidence for the efficacy of the Canine Surrogacy Approach in the Great Lakes. J. Archaeol. Sci. Rep. 13, 516–525. https://doi.org/10.1016/j.jasrep.2017.05.008 (2017).
Ames, K. M. et al. Stable isotope and ancient DNA analysis of dog remains from Cathlapotle (45CL1), a contact-era site on the Lower Columbia River. J. Archaeol. Sci. 57, 268–282. https://doi.org/10.1016/j.jas.2015.02.038 (2015).
Waterman, A. J. et al. Stable and radiogenic isotope analysis of faunal remains from a western Iowa Oneota complex site: An investigation of diet and mobility variation in the Late Prehistoric period of the upper Midwest. Midcont. J. Archaeol. 46, 223–254. https://doi.org/10.5406/23274271.46.3.02 (2021).
Glencross, B., Warrick, G., Smith, T. & Prowse, T. L. Estimating ancient Huron-Wendat diet in southern Ontario using stable isotopes from dogs. J. Archaeol. Sci. Rep. 41, 103324. https://doi.org/10.1016/j.jasrep.2021.103324 (2022).
Chiaradia, A., Forero, M. Z. G., McInnes, J. C. & Ramírez, F. Searching for the true diet of marine predators: Incorporating Bayesian priors into stable isotope mixing models. PLoS ONE 9, e92665. https://doi.org/10.1371/journal.pone.0092665 (2014).
Domingo, L. et al. Late Quaternary climatic and environmental conditions of northern Spain: An isotopic approach based on the mammalian record from La Paloma cave. Palaeogeogr. Palaeoclimatol. Palaeoecol. 440, 417–430. https://doi.org/10.1016/j.palaeo.2015.09.017 (2014).
García-Seoane, R., Viana, I. G. & Bode, A. Using MixSIAR to quantify mixed contributions of primary producers from amino acid δ15N of marine consumers. Mar. Environ. Res. 183, 105792. https://doi.org/10.1016/j.marenvres.2022.105792 (2023).
Hopkins, J. B. III. & Ferguson, J. M. Estimating the diets of animals using stable isotopes and a comprehensive Bayesian mixing model. PLoS ONE 7, e28478. https://doi.org/10.1371/journal.pone.0028478 (2012).
Hillis, D., McKechnie, I., Guiry, E., St Claire, D. E. & Darimont, C. T. Ancient dog diets on the Pacific Northwest Coast: Zooarchaeological and stable isotope modelling evidence from Tseshaht territory and beyond. Sci. Rep. 10, 1–12. https://doi.org/10.1038/s41598-020-71574-x (2020).
Jovanović, J. et al. Last hunters–first farmers: New insight into subsistence strategies in the Central Balkans through multi-isotopic analysis. Archaeol. Anthropol. Sci. 11, 3279–3298. https://doi.org/10.1007/s12520-018-0744-1 (2019).
Laffoon, J. E., Hoogland, M. L. P., Davies, G. R. & Hofman, C. L. A multi-isotope investigation of human and dog mobility and diet in the pre-colonial Antilles. Environ. Archaeol. 24(132–148), 2019. https://doi.org/10.1080/14614103.2017.1322831 (2019).
Moscardi, B. F. et al. Diet composition and prey choice in prehistoric human individuals from Northwest Patagonia: An application of species distribution and isotope mixing models. Am. J. Biol. Anthropol. 179, 568–584. https://doi.org/10.1002/ajpa.24626 (2022).
Tsutaya, T., Naito, Y. I., Ishida, H. & Yoneda, M. Carbon and nitrogen isotope analyses of human and dog diet in the Okhotsk culture: Perspectives from the Moyoro site, Japan. Anthropol. Sci. 2, 89–99. https://doi.org/10.1537/ase.140604 (2014).
Hart, J. P., Feranec, R. S., Abel, T. J. & Vavrasek, J. L. Freshwater reservoir offsets on radiocarbon-dated dog bone from the headwaters of the St. Lawrence River, USA. PeerJ 7, e7174. https://doi.org/10.7717/peerj.7174 (2019).
Feranec, R. S. & Hart, J. P. Fish and maize: Bayesian mixing models of fourteenth-through seventeenth-century AD ancestral Wendat diets, Ontario, Canada. Sci. Rep. 9, 16658. https://doi.org/10.1038/s41598-019-53076-7 (2019).
Ledogar, S. H., Feranec, R. S. & Zuhlke, J. M. Isotopic evidence for broad diet including anadromous fish during the mid-Holocene in northeastern North America. J. Archaeol. Sci. Rep. 19, 505–512. https://doi.org/10.1016/j.jasrep.2018.03.029 (2018).
Stock, B. C. et al. Analyzing mixing systems using a new generation of Bayesian tracer mixing models. PeerJ 6, e5096. https://doi.org/10.7717/peerj.5096 (2018).
Sealy, J., Armstrong, R. & Schrire, C. Beyond lifetime averages: tracing life histories through isotopic analysis of different calcified tissues from archaeological human skeletons. Antiquity 69, 290–300. https://doi.org/10.1017/S0003598X00064693 (1995).
Boitani, L., Ciucci, P. & Ortolani, A. Behaviour and social ecology of free-ranging dogs. In The Behavioural Biology of Dogs (ed. Jensen, P.) 147–165 (CAB International, 2019).
Sarkar, R., Sau, S. & Bhadra, A. Scavengers can be choosers: A study on food preference in free-ranging dogs. Appl. Anim. Behav. Sci. 216, 38–44. https://doi.org/10.1016/j.applanim.2019.04.012 (2019).
Butler, J. R. A., Brown, W. Y. & Du Toit, J. T. Anthropogenic food subsidy to a commensal carnivore: the value and supply of human faeces in the diet of free-ranging dogs. Animals 8, 67. https://doi.org/10.3390/ani8050067 (2018).
Atickem, A., Bekele, A. & Williams, S. D. Competition between domestic dogs and Ethiopian wolf (Canis simensis) in the Bale Mountains National Park, Ethiopia. Afr. J. Ecol. 48, 401–407. https://doi.org/10.1111/j.1365-2028.2009.01126.x (2010).
Vanak, A. T. & Gompper, M. E. Dogs Canis familiaris as carnivores: Their role and function in intraguild competition. Mammal Rev. 39, 265–283. https://doi.org/10.1111/j.1365-2907.2009.00148.x (2009).
Wrong, G. M. (ed.) (Greenwood Press, 1968).
Trigger, B. G. Children of Aataentsic: A History of the Huron People to 1660 (McGill-Queen’s Press, 1976).
Waugh, F. W. Iroquis [sic] Foods and Food Preparation. Canada Department of Mines, Geological Survey, Memoir 86, Anthropological Series No. 12. (Government Printing Office, 1916).
Monckton, S. G. Huron Paleoethnobotany. Ontario Archaeological Reports No. 1. (Ontario Heritage Foundation, 1992).
Pfeiffer, S., Sealy, J. C., Williamson, R. F., Needs-Howarth, S. & Lesage, L. Maize, fish, and deer: Investigating dietary staples among ancestral Huron-Wendat villages, as documented from tooth samples. Am. Antiq. 81, 515–532. https://doi.org/10.1017/S0002731600003978 (2016).
Hawkins, A. L., Needs-Howarth, S., Orchard, T. J. & Guiry, E. J. Beyond the local fishing hole: a preliminary study of pan-regional fishing in southern Ontario (ca. 1000 CE to 1750 CE). J. Archaeol. Sci. Rep. 24, 856–868. https://doi.org/10.1016/j.jasrep.2019.03.007 (2019).
Katzenberg, M. A., Schwarcz, H. P., Knyf, M. & Melbye, F. J. Stable isotope evidence for maize horticulture and paleodiet in southern Ontario, Canada. Am. Antiq. 60, 335–350. https://doi.org/10.2307/282144 (1995).
Booth, L. A stable isotope analysis of faunal remains from special deposits on Ontario Iroquoian tradition sites. PhD. Dissertation, The University of Western Ontario, London; https://ir.lib.uwo.ca/etd/2644/ (2014).
Guiry, E., Orchard, T. J., Needs-Howarth, S. & Szpak, P. Isotopic evidence for garden hunting and resource depression in the Late Woodland of northeastern North America. Am. Antiq. 86, 90–110. https://doi.org/10.1017/aaq.2020.86 (2021).
Parnell, A. C. et al. Bayesian stable isotope mixing models. Environmetrics 24, 387–399. https://doi.org/10.1002/env.2221 (2013).
Heidenreich, C. E. Huronia: A History and Geography of the Huron Indians, 1600–1650 (McClelland & Stewart, 1971).
Tooker, E. An ethnography of the Huron Indians, 1615–1649. Smithsonian Institution, Bureau of American Ethnology Bulletin 190. (US Government Printing Office, 1964).
Lovis, W. A. & Hart, J. P. Fishing for dog food: Ethnographic and ethnohistoric insights on the freshwater reservoir in northeastern North America. Radiocarbon 57, 557–570. https://doi.org/10.2458/azu_rc.57.18352 (2015).
Rose, C., Parker, A., Jefferson, B. & Cartmell, E. The characterization of feces and urine: A review of the literature to inform advanced treatment technology. Crit. Rev. Environ. Sci. Technol. 45, 1827–1879. https://doi.org/10.1080/10643389.2014.1000761 (2015).
Birch, J. & Williamson, R. F. The Mantle Site: An Archaeological History of an Ancestral Wendat Community (AltaMira Press, 2012).
Edwards, K., Walde, D. A. & Katzenberg, M. A. Searching for evidence of maize consumption at Cluny: Stable carbon and nitrogen isotope analysis of dog and bison bone collagen. Can. J. Archaeol. 40, 319–331 (2016).
Hart, J. P., Lovis, W. A., Schulenberg, J. K. & Urquhart, G. R. Paleodietary implications from stable carbon isotope analysis of experimental cooking residues. J. Archaeol. Sci. 34, 804–813. https://doi.org/10.1016/j.jas.2006.08.006 (2007).
Jones, D.B. Factors for Converting Percentages of Nitrogen in Foods and Feeds into Percentages of Protein. Circular 183. (US Department of Agriculture, 1941).
Kuhnle, G. G. C., Joosen, A. M. C. P., Kneale, C. J. & O’Connell, T. C. Carbon and nitrogen isotopic ratios of urine and faeces as novel nutritional biomarkers of meat and fish intake. Eur. J. Nutr. 52, 389–395. https://doi.org/10.1007/s00394-012-0328-2 (2013).
Joosen, A. M. C. P. et al. Effect of dietary meat and fish on endogenous nitrosation, inflammation and genotoxicity of faecal water. Mutagenesis 25, 243–247. https://doi.org/10.1093/mutage/gep070 (2010).
Hart, J. P. & Feranec, R. S. Using Maize δ15N values to assess soil fertility in fifteenth-and sixteenth-century AD Iroquoian agricultural fields. PLoS ONE 15, e0230952. https://doi.org/10.1371/journal.pone.0230952 (2001).
Bocherens, H. et al. Reconstruction of the Gravettian food-web at Předmostí I using multi-isotopic tracking (13C, 15N, 34S) of bone collagen. Quat. Int. 359, 211–228. https://doi.org/10.1016/j.quaint.2014.09.044 (2015).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 9 (2001).
Acknowledgements
I thank Susan Winchell-Sweeney for producing Fig. 1 and Robert Feranec for discussions on Bayesian dietary mixing models and for comments and suggestions on an earlier draft of this paper.
Ethics declarations
Competing interests
The author declares no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hart, J.P. Human and dog Bayesian dietary mixing models using bone collagen stable isotope ratios from ancestral Iroquoian sites in southern Ontario. Sci Rep 13, 7177 (2023). https://doi.org/10.1038/s41598-023-34216-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-34216-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
