
Abstract
The first comprehensive species distribution models for orchid, its fungal symbionts and pollinator are presented. To evaluate impact of global warming on these organisms three different projections and four various climate change scenarios were analysed. The niche modelling was based on presence-only records of Limodorum abortivum, two species of Russula and three insects pollinating orchid (Anthophora affinis, Bombus terrestris, Rhodanthidium septemdentatum). Two sets of orchid predictions were examined—the first one included only climatic data and the second one was based on climate data and data on future distribution of orchid fungal symbionts. Overall, a poleward range shift is predicted to occur as a result of climate change and apparently global warming will be favorable for L. abortivum and its potential geographical range will expand. However, due to the negative effect of global warming on fungal symbionts of L. abortivum, the actual extension of the suitable niches of the orchid will be much limited. Considering future possibility of cross-pollination, the availability of A. affinis for L. abortivum will decrease and this bee will be available in the worst case scenarios only for 21% of orchid populations. On the other hand, the overlap of orchid and the buff-tailed bumblebee will increase and as much as 86.5% of plant populations will be located within B. terrestris potential range. Also the availability of R. septemdentatum will be higher than currently observed in almost all analysed climate change projections. This study showed the importance of inclusion of ecological factors in species distribution models as the climate data itself are not enough to estimate the future distribution of plant species. Moreover, the availability of pollen vectors which is crucial for long-term survival of orchid populations should be analysed in context of climate changes.
Similar content being viewed by others


Introduction
Climate is the most important factor determining species distribution1. To understand how the conditions of our planet changed over time various research teams developed climate models (general circulation models, GCMs) describing past2, present3 and future4 climatic Earth conditions. These GCMs became important tool in biogeographic5,6,7,8,9, phylogenetic10,11 and ecological12,13,14,15 research. Species distribution models based on GCMs are also used in nature conservation planning and preventing invasive species expansion16,17,18,19. Considering possible negative impact of climate change modification of temperature and precipitation patterns together with extreme weather events can alter geographical ranges of species20, their ecological interactions21, and the timing of biological events (phenology), which could fundamentally transform ecosystems and food webs22.
Clearly, any environmental change is potentially more dangerous for specialized organisms characterized by narrow ecological tolerance and depending on numerous interspecific interactions. Due to the complex life cycle Orchidaceae are among the most threatened plants in the global scale23. According to the International Union for the Conservation of Nature (IUCN) almost half of the extinct orchid species are terrestrial herbaceous perennials24. All Orchidaceae develop mycorrhizal symbioses with fungi in natural habitats that affect not only their seed germination, but also protocorm growth, and adult nutrition25. Orchids interact with a more limited set of mycorrhizal fungi as compared to other mycorrhizal plants25. Although many initial mycoheterotrophs develop into autotrophic mature plants, some orchids continue to obtain carbon from mycorrhizal fungi throughout their lifecycle (full mycoheterotrophy) or utilize both photosynthesis and mycoheterotrophy at maturity (partial mycoheterotrophy, mixotrophy)26,27. Generally, orchid flowers are adapted to promote cross-pollination and most species depend on pollen vectors for reproduction28,29,30,31. Autogamy is not common in Orchidaceae and this mode of reproduction evolved only in about 5–20% of the family representatives32,33,34.
One of the very interesting orchid species which is strongly associated with fungi not only during seed germination, but also as mature plant, and which is adapted to both cross-pollination and autogamy is Limodorum abortivum, also known as the Violet Limodore. This terrestrial plant does not produce any basal leaves and the upper, cauline ones are modified into sheathing bracts. Whitish-violet flowers of L. abortivum are arranged in a long racemose inflorescence. The Violet Limodore is a myco-heterotrophic plant and so far three species of Russula (R. brevipes, R. chloroides, R. delica) were found to be L. abortivum fungal symbionts compensating insufficient plant CO2 fixation35. Flowers of L. abortivum are considered to be mostly cleistogamous36, but some pollination events were recorded. So far three pollen vectors of the Violet Limodore were identified—two bee species (Anthophora biciliata and Rhodanthidium septemdentatum) and the buff-tailed bumblebee (Bombus terrestris)37. Limodorum abortivum is native to mainland Europe, western Asia and the Mediterranean area. According to the IUCN it is a species of Least Concern38 but in numerous geographical areas it is considered to be endangered (Critically Endangered39,40, Endangered41, Vulnerable42 or Near Threatened43,44).
The aim of this study was to evaluate the importance of fungal symbionts presence in the future distribution of L. abortivum and to estimate fluctuations in the availability of orchid pollen vector under various climate change scenarios using ecological niche modelling.
Methods
List of localities
The database of records of L. abortivum, its fungal symbionts and pollinators was compiled based on data included in the public catalogue—Global Biodiversity Information Facility (GBIF). The initial datasets (Supplementary Annex 1) were verified and only records correctly georeferenced with a precision of at least 1 km were analysed further. Because previous studies45,46 indicated that usage of a restricted area in ENM analysis is more reliable than modelling on a global scale, the area studied was limited to 23.42S-65.64ºN, 13.96ºW-57.41ºE.
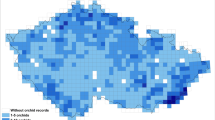
Spatial thinning was conducted using SDMtoolbox 2.3 for ArcGIS47,48 to reduce the spatial bias of sampling49,50,51. While geographic and environmental thinning are conceptually equivalent, as they use a distance measure to determine a filter size52, they can lead to a disproportionate sampling in the environmental space. For that reason the additional factor, topography, was included in the spatial thinning process. The topographic heterogeneity of the study area was calculated53 and divided into five classes. Localities were spatially filtered at 5.0, 10.0, 15.0, 20.0 and 25.0 km in areas of high, medium high, medium, medium low and low heterogeneity, respectively. The final database of localities included 1074 records of L. abortivum, 19 of A. affinis, 1440 of B. terrestris, 103 of Rhodanthidium septemdentatum, 388 of R. chloroides, and 606 of R. delica (Fig. 1, Supplementary Annex 2).

Ecological niche modelling
Two models were used to estimate the effect of global warming on L. abortivum. The first was based exclusively on bioclimatic variables (bioclims) and the second included the modelled distributions of the fungal symbionts (bioclims + fungi).
MaxEnt v. 3.3.254,55 which is considered to be one of the best-performing niche modeling algorithm based on presence-only data56,57 was used to produce the ecological niche model of the species studied. The algorithm selection was also motivated by the scarce information on distribution of studied organisms and lack of possibility of creating reliable pseudo-absence data. Due to this obstacle none of the existing presence-absence modelling methods could be used58. For the analyses bioclimatic variables (bioclims) in 30 arc-seconds of interpolated climate surface downloaded from WorldClim v. 2.13 were used. Pearsons’ correlation coefficient was calculated using SDMtoolbox v. 2.3 for ArcGIS47,48 and highly correlated bioclimatic variables (above 0.9; Supplementary Annex 3) were removed from the analyses (Table 1).
To evaluate the impact of global warming on the distribution of species studied predictions of the extent of the climatic niches of L. abortivum, its symbionts and pollinator in 2080–2100 were made. The models were compiled using climate projections for four Shared Socio-economic Pathways (SSPs): SSP1-2.6, SSP2-4.5, SSP3-7.0 and SSP5-8.5. These trajectories were established to describe probable major global developments that would lead in the future to different challenges for mitigation and adaptation to climate change59. The SSPs are based on narratives describing alternative socio-economic developments (sustainable development, regional rivalry, inequality, fossil-fueled development, middle-of-the-road development) with global warming in 2100 ranging from of 3.1ºC to 5.1ºC above pre-industrial levels60,61. Three different simulations of future climate developed by Coupled Model Intercomparison Project Phase 6 (CNRM-CM6-1), Goddard Institute for Space Studies (GISS-E2-1), and Institute for Numerical Mathematics (INM-CM5) were used. While some Global Climate Models (GCMs) were proved to work better in specific geographical regions62, the area in this study included whole Europe and north Africa, hence it was not possible to select one, best-performing simulation. For that reason the three aforementioned GCMs which significantly differ in the simulated maximum temperature and precipitation within study area were used. This approach allows to present the broadest spectrum of possible changes in the distribution of studied organisms.
In all analyses the maximum number of iterations was set to 10,000 and convergence threshold to 0.00001. The neutral regularization multiplier value and auto features were utilized. The random test partition and background subset for each run was applied using 30% of the samples as test points. The run was performed as a bootstrap with 100 replicates, and the output was set to logistic. All analyses of GIS data were carried out using ArcGis v. 10.8 (Esri, Redlands, CA, USA). To prevent extrapolations outside the environmental range of the training data the “fade by clamping” function in MaxEnt was used63,64,65. The evaluation of the created models was made using the area under the curve (AUC)66 and True Skill Statistic (TSS)67. Both metrics are frequently used measure of model performance as they are independent of prevalence68.
SDMtoolbox v. 2.3 for ArcGIS47,48 was used to calculate changes in the distribution of suitable niches of L. abortivum caused by global warming. For this operation created models (for present-time and future) were converted into binary rasters in Goode homolosine projection. The max kappa value was used as presence threshold and used to compare the extent and location of suitable niches of studied species between present-time and future models. To calculate max kappa value a phyloclim package for R was used69.
Results
Models evaluation and limiting factors
The model performance indexes (AUC and TSS) are presented in Table 2 together with Max Kappa value which was used as a presence threshold. The AUC values of 0.875–0.971 indicate high reliability of the models, however, the TSS for Anthophora affinis and Bombus terrestris received lower scores and these models should be taken with caution.
The results of the jackknife test of variable importance in both sets of models created for L. abortivum showed that variable with highest gain when used in isolation is bio1, which therefore appears to have the most useful information by itself. The same variable appears to have the most information that isn't present in the other variables as it reduces the gain the most when it is omitted is bio1 (Fig. 2). Other important bioclimatic factors influencing distribution of L. abortivum are bio9, bio4, and bio7. As indicated in the jackknife test, the presence of the two Russula species is more important than bio3, bio14, bio2, bio8 and bio15.

The results of the jackknife test of variable importance. (A) bioclims only based models, (B) models created based on combined bioclims and fungi data). Values shown are averages over replicate runs.
Impact of climate change on orchid symbionts
The future of Russula chloroides is uncertain (Table 3, Supplementary Annex 4). While in almost all climate change scenarios projected by CNRM and GISS this fungi will face serious suitable niche loss (13–61%), the INM prediction is more favorable and predicts even 95% expansion of the current fungus range, especially in the northern part of its geographical range.
Russula delica will lose suitable niches in all predicted climate change scenarios (Table 3, Supplementary Annex 4). According to CNRM projections the potential range of this fungi will be reduced for 39–68%. Similar result was obtained in GISS projection (39–54%) but in the INM forecast the loss will be lower (9–26%). The most significant niche loss will be observed in southern ad central part of the fungus range.
Impact of climate change on orchid
According to models created based on bioclimatic data only, the potential range of L. abortivum will expand in most predicted climate change scenarios except of SSP5-8.5 (CNRM and GISS projections) and SSP3-7.0 (GISS projection) (Table 3, Supplementary Annex 5). However, due to the predicted loss of suitable niches by its fungal symbionts, the models based on both bioclimatic data and distribution models of fungi, the extent of range expansion will be lower in all climate change scenarios. Moreover, according to the GISS projections the orchid will lose 3–38% of currently suitable niches (Table 3). The most significant niche loss will be observed in the southern part of the orchid range and expansion to the north and north-east direction is predicted (Fig. 3).

Changes in the orchid distribution according to the combined (bioclims + fungi) models. (a) SSP1-2.6 scenario, (b) SSP2-4.5 scenario, (c) SSP3-7.0 scenario, (d) SSP5-8.5 scenario. Legend: -1—range expansion, 0—no occurrence, 1—present, 2—range contraction. Maps generated by the author in ArcGIS47,48.
Impact of climate change on orchid pollinators
As mentioned before, models of Anthophora affinis and Bombus terrestris received lower scores of TSS than expected (but relatively high AUC scores), however, the models of these insects created for the present time are consistent with the known geographical ranges of species studies and should be discussed as reliable ones. Anthophora affinis will lose suitable niches in the southern part of its geographical range according to all conducted analyses (Table 3, Supplementary Annex 6). The expansion of current range is expected to occur mostly in Central Europe. In CNRM forecast the range will contract by 14–63%, in GISS 12–30% and in INM by 15–38%. The currently observed overlap of the potential range of Anthophora affinis and L. abortivum is about 67% but in almost all predictions it will decrease as a result of climate change (Table 4, Fig. 4).

Presence of pollinator within L. abortivum range. Areas of overlap between pollinator and orchid range marked with green, areas suitable only for an orchid occurrence marked in sand yellow. (a) present time, (b) SSP1-2.6 scenario, (c) SSP2-4.5 scenario, (d) SSP3-7.0 scenario, (e) SSP5-8.5 scenario. Maps generated by the author in ArcGIS47,48.
The future of Bombus terrestris is uncertain. According to CNRM and INM projections the climate changes will be generally favorable for this pollinator which will expand its geographical range in almost all analysed climate change scenarios (Table 3, Supplementary Annex 6), except of CNRM SSP5-8.5 and SSP3-7.0 predictions On the other hand, the GISS projection predicts the significant loss (19–22%) of suitable niches in all scenarios of global warming (Table 3). Overall, the contraction is expected to occur in south and south-eastern part of the insect geographical range. The new niches should be available for the bumblebee in northern and north-western edges of the current range. The overlap of orchid and B. terrestris ranges will be much higher than currently observed (Table 4, Fig. 4) and this pollinator will be available for almost all populations of L. abortivum.
Climate change will be very favorable for Rhodanthidium septemdentatum according to CNRM and INM projections (except of INM SSP1-2.6 scenario). In these predictions the insect will much expand its potential range (Table 3, Supplementary Annex 6). However, as the analyses conducted using GISS projections indicated future loss of suitable niches of orchid pollinator (33–71%). This bee is currently available for approximately 23% of orchid populations. The overlap of orchid and R. septemdentatum potential ranges will be similar to or higher than currently observed in almost all CNRM and INM projections. On the contrary, the availability of pollen vector will be reduced in GISS simulations (Table 4, Fig. 4).
Discussion
Importance of ecological factors in species distribution models
This study showed the importance of ecological factor in species distribution models. Even though according to the jackknife test, the geographical range of L. abortivum is shaped mainly by climatic factors, the incorporation of symbiotic fungi models in the analyses significantly modified the predicted distribution of orchid under future climate change.
While all Orchidaceae require mycorrhizal symbioses during the seed germination as their seeds lack endosperms and contain limited storage reserves25, some species were found to keep receiving carbon from mycorrhizal fungi even after initiation of photosynthesis70,71. As indicated in previous research72 the photosynthetic rates of the Violet Limodore are very low and the carbon assimilation activity is insufficient to support the energy requirements of adult plants.
So far little is known on the impact of global warming on plant endophytes, primarily because most symbionts remain unidentified73. While molecular studies allowed to isolate and genetically characterize fungi from orchid roots, leaves and stems74,75,76,77, the actual composition of orchid fungal symbionts is still poorly explored and most of taxa found in plant tissues have been assigned only to family or genus rank78,79,80,81. The lack of species-level identification of orchid symbionts excludes possibility of any reliable broad-scale analyses of the importance of fungi on the Orchidaceae distribution.
As any research aimed at estimating possible impact of climate change on complex relationships between plant, fungi and insects, this study has some limitations. So far the analyses of L. abortivum symbionts were geographically limited and further investigations are required to evaluate the actual variation of fungal symbionts of this orchid. It is possible that not only Russulaceae can compensate insufficient plant CO2 fixation. Moreover, Girlanda et al.35 identified three species of Russulaceae as symbionts of L. abortivum. Two of them were included in ENM analyses for this study. The third one, Russula brevipes is widespread in North America and only recently was reported outside its native range. It is not sure if this fungus will actually spread in Eurasia to serve as a long-term symbiont for L. abortivum. Also, due to the small number of records of this fungus in the non-native areas, at this point it is not possible to evaluate its current or future potential range in Eurasia.
Noteworthy, Russula representatives are ectomycorrhizal fungi and their occurrence is strictly related with the presence of host plants. R. delica seems to have broad spectrum of potential hosts and it was reported growing with both conifers (Pseudotsuga menziesii) and broadleaved trees (Corylus, Fagus, Quercus, Tilia)82. On the other hand, R. chloroides favours habitats with oak trees, although it was sometimes reported growing under other broadleaf trees (Carpinus, Crataegus) and conifer trees (Abies alba)83,84.
Uncertainty of climate models
As global warming became one of the most important threats to biodiversity and ecosystems functioning, numerous mathematical models of future climate changes scenarios has been produced to describe possible impact of human on Earth temperature and precipitations85. Clearly, the usefulness of any climate model is tested by the conformity of its output to a given set of known conditions. For that reason, the actual trustworthiness of any models presenting future climate is impossible to assess. In this study three different simulations of climatic conditions were used to produce the most reliable prediction of changes in the distribution of suitable niches of species studied.
Noteworthy, earlier models created for L. abortivum86 based on formerly recognized climate change scenarios (A1b, A2a and B2a) indicated loss of orchid suitable niches. In the present study habitat loss was predicted only in GISS simulation of future climate change while both CNRM and INM projections forecasted expansion of orchid geographical range. While most areas currently occupied by the orchid will not be suitable for this species in the future, all created models predicted migration of L. abortivum to the higher latitudes and loss of suitable niches in the southern part of species geographical range. The same scheme of poleward range shift as a response to the global warming was already predicted to occur in other plant87 and animal88,89 species. Evans and Jacquemyn90 suggested that terrestrial Orchidaceae with a wide distribution will be more capable of shifting their distributions under global warming than species with a restricted geographical range. While obviously species with broader environmental tolerance have higher survival chances than more specialized taxa, the fragility of ecological interactions can further affect persistence of widely distributed organisms. Studies on future distribution of European orchids are rather scarce and while some taxa (Nigritella nigra91, Pseudorchis albida92) are predicted to lose their suitable niches across their ranges, others are expected to experience a poleward range shift (some Orchis93, Ophrys insectifera94, some Epipactis90). Noteworthy, none of the previous research considered the importance of mycorrhizal fungi on Orchidaceae distribution and most of the published analyses ignored also the future availability of orchid pollen vectors.
While it is generally accepted that climatic factors play the primary role in shaping biodiversity at broad scales, there are numerous other abiotic variables which can affect species occurrence. Scientists still explore the importance of topography95, physico-chemical properties of soil96,97,98, and the substrate thickness99 on the plant distribution100. Unfortunately, at this moment there are no tools which could be used to predict changes of these variables in response to climate change and hence these factors cannot be included in the niche modelling.
Another obstacle in broad scale modelling of species distribution is lack of data and unequal sampling throughout the geographical range of the species. The availability of precise location data remains limited, especially when dealing with rare or poorly recognized species49,101,102. Moreover, species records are often constrained by the variety of their sources and spatial biases caused by unequal sampling efforts103 and by uneven field accessibility104. The adequacy of sample bias correction methods remains uncertain105 and field validation is still considered to be the best standard practice to assess models’ reliability106. Unfortunately, field validation is sometimes impossible to use, especially in geographically extensive scale studies105. In this study some geographical regions also seemed to be inadequately sampled and for that reason the spatial filtering on various scales of topographic heterogeneity was conducted to reduce sampling bias. The general consistence of the models created for the present time with the known geographical range of studied species suggest reliability of conducted analyses, also for species which received lower scores of TSS tests (but relatively high AUC test scores).
Future availability of pollinators
Flowers of the Violet Limodore are most often cleistogamous (pollination takes place inside the flower bud), but insect pollination is more beneficial for the long-term survival of L. abortivum populations by increasing their genetic diversity. So far very few studies on the impact of global warming on orchid distribution incorporated also the analyses of future availability of pollen vectors for plant populations107,108,109. This study indicated that the climate change can be favorable for orchid not only by direct expansion of the niches suitable for the plant occurrence, but also in increasing pollinators availability as a result of insects range shifts.
In this study two species of solitary bees were included in the analyses. According to created models A. affinis will lose numerous suitable niches while R. septemdentatum will expand its geographical range. However, as indicated in previous studies110, for solitary bees, responses to climate change will be related to biological processes occurring prior to emergence. Among possible consequences of global warming accelerated development111, reduced energy reserves112,113, increased mortality114, advanced emergence115, and reduced post-emergence lifespan116 are expected to occur.
While according to available data the main flight period of both studied bee species, A. biciliata and R. septemdentatum, is rather short and is susceptible for climate change, the buff-tailed bumblebee is relatively resistant to climate warming. According to the published study results117 advances in mid-date of the main flight period over the 35-year period was 13 days for B. terrestris and the duration of the main flight period was reduced by about 7 days. The models presented in this study do not consider possibility of incompatibility of orchid flowering time and its pollinator phenology. This dangerous phenomena which can disturb pollen transfer and plant reproductive success was already detected in European and Australian orchids118,119. The possible mis-match of orchid flowering time and insect activity period can further limit the possibility of cross-pollination of L. abortivum.
Conclusions
To conclude, the incorporation of ecological relationships, e.g. fungal symbionts, pollen vectors, is crucial to produce more accurate distribution models of plant species. Moreover, due to the discrepancies between projections of future climatic conditions, various predictions and climate change scenarios should be analysed to uncover all possible changes in the studied species potential geographical range. The maps presented in this study can be useful for establishing conservation actions on L. abortivum. The priority in creating new conservation areas should be given to the regions which will be suitable for the occurrence of the orchid, its symbionts and pollinators in the future. Without doubt, more effort should be made to identify orchid symbionts and incorporate geographical distribution records of fungi into public databases to allow more efficient geographical analyses of plant-fungi relationships.
Data availability
All relevant data are presented in the manuscript and supplementary files.
References
Mantyka-Pringle, C. S., Martin, T. G. & Rhodes, J. R. Interactions between climate and habitat loss effects on biodiversity: A systematic review and meta-analysis. Glob. Change Biol. 18, 1239–1252. https://doi.org/10.1111/j.1365-2486.2011.02593.x (2012).
Brown, J. L., Hill, D. J., Dolan, A. M., Carnaval, A. C. & Haywood, A. M. PaleoClim, high spatial resolution paleoclimate surfaces for global land areas. Sci. Data 5, 180254. https://doi.org/10.1038/sdata.2018.254 (2018).
Fick, S. & Hijmans, R. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. https://doi.org/10.1002/joc.5086 (2017).
Voldoire, A. et al. Evaluation of CMIP6 DECK experiments with CNRM-CM6-1. J. Adv. Model. Earth Syst. 11, 2177–2213. https://doi.org/10.1029/2019MS001683 (2019).
Wollan, A., Bakkestuen, V., Kauserud, H., Gulden, G. & Halvorsen, R. Modelling and predicting fungal distribution patterns using herbarium data. J. Biogeogr. 35, 2298–2310. https://doi.org/10.1111/j.1365-2699.2008.01965.x (2008).
Pierrat, B. et al. Large-scale distribution analysis of Antarctic echinoids using ecological niche modelling. Mar. Ecol. Prog. Ser. 463, 215–230. https://doi.org/10.3354/meps09842 (2012).
Brun, P. et al. Ecological niches of open ocean phytoplankton taxa. Limnol. Oceanogr. 60, 1020–1038. https://doi.org/10.1002/lno.10074 (2015).
Quesada-Quiros, M., Acosta-Vargas, L., Arias-Aguilar, D. & Rodriguez-Gonzalez, A. Ecological niche modeling based on three climate change scenarios for five species of plants in high areas of Costa Rica. Revista Forestal Mesoamerica Kuru-Rfmk 14, 1–12. https://doi.org/10.18845/rfmk.v14i34.2991 (2017).
Sharifian, S., Kamrani, E. & Saeedi, H. Global future distributions of mangrove crabs in response to climate change. Wetlands https://doi.org/10.1007/s13157-021-01503-9 (2021).
Wang, P., Liu, Y., Chang, Y., Wang, N. & Zhang, Z. The role of niche divergence and geographic arrangement in the speciation of Eared Pheasants (Crossoptilon, Hodgson 1938). Mol. Phylogenet Evol. 113, 1–8. https://doi.org/10.1016/j.ympev.2017.05.003 (2017).
Calixto-Rojas, M., Lira-Noriega, A., Rubio-Godoy, M., de Leon, G. & Pinacho-Pinacho, C. Phylogenetic relationships and ecological niche conservatism in killifish (Profundulidae) in Mesoamerica. J. Fish Biol. 99, 396–410. https://doi.org/10.1111/jfb.14727 (2021).
Pavlek, M. & Mammola, S. Niche-based processes explaining the distributions of closely related subterranean spiders. J. Biogeogr. 48, 118–133. https://doi.org/10.1111/jbi.13987 (2021).
Fourcade, Y. Fine-tuning niche models matters in invasion ecology. A lesson from the land planarian Obama nungara. Ecol. Modell. https://doi.org/10.1016/j.ecolmodel.2021.109686 (2021).
Bobrowski, M., Weidinger, J., Schwab, N. & Schickhoff, U. Searching for ecology in species distribution models in the Himalayas. Ecol. Modell. https://doi.org/10.1016/j.ecolmodel.2021.109693 (2021).
Prieto-Torres, D., Rosas, L., Figueroa, D. & Arizmendi, M. Most Mexican hummingbirds lose under climate and land-use change: Long-term conservation implications. Perspect. Ecol. Conserv. 19, 487–499. https://doi.org/10.1016/j.pecon.2021.07.001 (2021).
Meyer, A. L., Pie, M. R. & Passos, F. C. Assessing the exposure of lion tamarins (Leontopithecus spp.) to future climate change. Am. J. Primatol. 76, 551–562. https://doi.org/10.1002/ajp.22247 (2014).
Firn, J. et al. Priority threat management of invasive animals to protect biodiversity under climate change. Glob. Chang. Biol. 21, 3917–3930. https://doi.org/10.1111/gcb.13034 (2015).
Magalhães-Júnior, A. J. C., Moura, G. J. B., Ribeiro, L. B. & Azevedo-Júnior, S. M. Potential distribution and conservation of the Colobosauroides carvalhoi Soares and Caramaschi, 1998: A rare and endemic lizard of Northeast Brazil. Braz. J. Biol. 77, 686–695. https://doi.org/10.1590/1519-6984.15815 (2017).
Li, R. Protecting rare and endangered species under climate change on the Qinghai Plateau China. Ecol. Evol. 9, 427–436. https://doi.org/10.1002/ece3.4761 (2019).
Lovejoy, T. Climate change and biodiversity. Rev. Sci. Tech. 27, 331–338 (2008).
Freimuth, J., Bossdorf, O., Scheepens, J. F. & Willems, F. M. Climate warming changes synchrony of plants and pollinators. bioRxiv 41, 425984. https://doi.org/10.1101/2021.01.10.425984 (2021).
Zhang, L., Takahashi, D., Hartvig, M. & Andersen, K. H. Food-web dynamics under climate change. Proc. Biol. Sci. https://doi.org/10.1098/rspb.2017.1772 (2017).
Cribb, P. J., Kell, S. P., Dixon, K. W. & Barrett, R. L. in Orchid conservation (eds K. W. Dixon, S. P. Kell, R. L. Barrett, & P. J. Cribb) 1–24 (Natural History Publications, 2003).
Swarts, N. D. & Dixon, K. W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 104, 543–556. https://doi.org/10.1093/aob/mcp025 (2009).
Li, T., Yang, W., Wu, S., Selosse, M. A. & Gao, J. Progress and prospects of mycorrhizal fungal diversity in orchids. Front. Plant Sci. 12, 646325. https://doi.org/10.3389/fpls.2021.646325 (2021).
Delaux, P.-M. et al. Algal ancestor of land plants was preadapted for symbiosis. Proc. Natl. Acad. Sci. 112, 13390–13395. https://doi.org/10.1073/pnas.1515426112 (2015).
Miura, C. et al. The mycoheterotrophic symbiosis between orchids and mycorrhizal fungi possesses major components shared with mutualistic plant-mycorrhizal symbioses. Mol. Plant Microbe Interact 31, 1032–1047. https://doi.org/10.1094/MPMI-01-18-0029-R (2018).
Jersakova, J., Johnson, S. & Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 81, 219–235. https://doi.org/10.1017/S1464793105006986 (2006).
Vereecken, N. J. & Schiestl, F. P. The evolution of imperfect floral mimicry. Proc. Natl. Acad. Sci. U. S. A. 105, 7484–7488. https://doi.org/10.1073/pnas.0800194105 (2008).
Micheneau, C. et al. Orthoptera, a new order of pollinator. Ann. Bot. 105, 355–364. https://doi.org/10.1093/aob/mcp299 (2010).
Schiestl, F. P. & Schluter, P. M. Floral isolation, specialized pollination, and pollinator behavior in orchids. Annu. Rev. Entomol. 54, 425–446. https://doi.org/10.1146/annurev.ento.54.110807.090603 (2009).
Tremblay, R., Ackerman, J., Zimmerman, J. & Calvo, R. Variation in sexual reproduction in orchids and its evolutionary consequences: A spasmodic journey to diversification. Biol. J. Lin. Soc. 84, 1–54. https://doi.org/10.1111/j.1095-8312.2004.00400.x (2005).
Wang, Q. et al. A novel case of autogamy and cleistogamy in Dendrobium wangliangii: A rare orchid distributed in the dry-hot valley. Ecol. Evol. 9, 12906–12914. https://doi.org/10.1002/ece3.5772 (2019).
Liu, K. W. et al. Pollination: Self-fertilization strategy in an orchid. Nature 441, 945–946. https://doi.org/10.1038/441945a (2006).
Girlanda, M. et al. Inefficient photosynthesis in the Mediterranean orchid Limodorum abortivum is mirrored by specific association to ectomycorrhizal Russulaceae. Mol. Ecol. 15, 491–504. https://doi.org/10.1111/j.1365-294X.2005.02770.x (2006).
Wood, J. Limodorum abortivum (Orchidaceae). Kew Mag. 10, 161–166 (1993).
Claessens, J. & Kleynen, J. The pollination of European Orchids Part 3: Limodorum and Epipactis. J. Hardy Orchid Soc. 11, 64–72 (2014).
Rankou, H. Limodorum abortivum (Europe assessment). IUCN Red List Threat. Species 2011, 2022 (2011).
Holub, J. & Procházká, F. Red list of vascular plants of the Czech Republic - 2000. Preslia 72, 187–230 (2000).
Ludwig, G. & Schnittler, M. Rote Liste gefährdeter Pflanzen Deutschlands. Schriftenreihe für Vegetationskunde 28, 709–739 (1996).
Witkowski, Z. J., Król, W. & Solarz, W. (2003) Carpathian list of endangered species. 68 (WWF and Institute of Nature Conservation, Polish Academy of Sciences, London).
Petrova, A. & Vladimirov, V. Red list of Bulgarian vascular plants. Phytologia Balcanica 15, 63–94 (2009).
Király, G. Red list of the vascular flora of Hungary (Saját kiadás, 2007).
Moser, D., Gygax, A., Bäumler, B., Wyler, N. & Raoul, P. Rote Liste der gefährdeten Arten der Schweiz: Farn- und Blütenpflanzen (BUWAL, 2002).
Mendes, P., Velazco, S., de Andrade, A. & De Marco, P. Dealing with overprediction in species distribution models: How adding distance constraints can improve model accuracy. Ecol. Modell. https://doi.org/10.1016/j.ecolmodel.2020.109180 (2020).
Barve, N. et al. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 222, 1810–1819. https://doi.org/10.1016/j.ecolmodel.2011.02.011 (2011).
Brown, J. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 5, 694–700. https://doi.org/10.1111/2041-210X.12200 (2014).
Brown, J. L., Bennett, J. R. & French, C. M. SDMtoolbox 20: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 5, e4095. https://doi.org/10.7717/peerj.4095 (2017).
Kramer-Schadt, S. et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 19, 1366–1379. https://doi.org/10.1111/ddi.12096 (2013).
Veloz, S. Spatially autocorrelated sampling falsely inflates measures of accuracy for presence-only niche models. J. Biogeogr. 36, 2290–2299. https://doi.org/10.1111/j.1365-2699.2009.02174.x (2009).
Tourne, D. C. M. et al. Strategies to optimize modeling habitat suitability of Bertholletia excelsa in the Pan-Amazonia. Ecol. Evol. 9, 12623–12638. https://doi.org/10.1002/ece3.5726 (2019).
Castellanos, A., Huntley, J., Voelker, G. & Lawing, A. Environmental filtering improves ecological niche models across multiple scales. Methods Ecol. Evol. 10, 481–492. https://doi.org/10.1111/2041-210X.13142 (2019).
Luoto, M. & Heikkinen, R. K. Disregarding topographical heterogeneity biases species turnover assessments based on bioclimatic models. Glob. Change Biol. 14, 483–494. https://doi.org/10.1111/j.1365-2486.2007.01527.x (2008).
Babar, S., Amarnath, G., Reddy, C., Jentsch, A. & Sudhakar, S. Species distribution models: Ecological explanation and prediction of an endemic and endangered plant species (Pterocarpus santalinus L.f.). Current Sci. 102, 1157–1165 (2012).
Elith, J. et al. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 17, 43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x (2011).
Elith, J. et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29, 129–151. https://doi.org/10.1111/j.2006.0906-7590.04596.x (2006).
Grimmett, L., Whitsed, R. & Horta, A. Presence-only species distribution models are sensitive to sample prevalence: Evaluating models using spatial prediction stability and accuracy metrics. Ecol. Modell. 431, 109194. https://doi.org/10.1016/j.ecolmodel.2020.109194 (2020).
Guillera-Arroita, G. et al. Maxent is not a presence–absence method: A comment on Thibaud. Methods Ecol. Evol. 5, 1192–1197. https://doi.org/10.1111/2041-210X.12252 (2014).
Riahi, K. et al. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Global Environ. Change-Human Policy Dimens. 42, 153–168. https://doi.org/10.1016/j.gloenvcha.2016.05.009 (2017).
van Vuuren, D. P. et al. Energy, land-use and greenhouse gas emissions trajectories under a green growth paradigm. Glob. Environ. Chang. 42, 237–250. https://doi.org/10.1016/j.gloenvcha.2016.05.008 (2017).
van Vuuren, D. et al. The shared socio-economic pathways: Trajectories for human development and global environmental change. Global Environ. Change-Human Policy Dimens. 42, 148–152. https://doi.org/10.1016/j.gloenvcha.2016.10.009 (2017).
Parding, K. M. et al. GCMeval – An interactive tool for evaluation and selection of climate model ensembles. Clim. Serv. 18, 100167. https://doi.org/10.1016/j.cliser.2020.100167 (2020).
Owens, H. et al. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol. Model. 263, 10–18. https://doi.org/10.1016/j.ecolmodel.2013.04.011 (2013).
Tobena, M., Prieto, R., Machete, M. & Silva, M. Modeling the potential distribution and richness of cetaceans in the Azores from fisheries observer program data. Front. Mar. Sci. https://doi.org/10.3389/fmars.2016.00202 (2016).
Phillips, S., Anderson, R. & Schapire, R. Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026 (2006).
Mason, S. & Graham, N. Areas beneath the relative operating characteristics (ROC) and relative operating levels (ROL) curves: Statistical significance and interpretation. Q. J. R. Meteorol. Soc. 128, 2145–2166. https://doi.org/10.1256/003590002320603584 (2002).
Čengić, M. et al. On the importance of predictor choice, modelling technique, and number of pseudo-absences for bioclimatic envelope model performance. Ecol. Evol. 10, 12307–12317. https://doi.org/10.1002/ece3.6859 (2020).
Shabani, F., Kumar, L. & Ahmadi, M. A comparison of absolute performance of different correlative and mechanistic species distribution models in an independent area. Ecol. Evol. 6, 5973–5986. https://doi.org/10.1002/ece3.2332 (2016).
Phyloclim: Integrating phylogenetics and climatic niche modeling (R package version 0.9–4 2013).
Stöckel, M., Těšitelová, T., Jersáková, J., Bidartondo, M. I. & Gebauer, G. Carbon and nitrogen gain during the growth of orchid seedlings in nature. New Phytol. 202, 606–615. https://doi.org/10.1111/nph.12688 (2014).
Tsukaya, H. How leaves of mycoheterotrophic plants evolved – from the view point of a developmental biologist. New Phytol. 217, 1401–1406. https://doi.org/10.1111/nph.14994 (2018).
Shevtsov, S. et al. The complete plastid genome sequence and the photosynthetic activity of the putative mycoheterotrophic orchid Limodorum abortivum. Israel J. Plant Sci. 66, 69–88. https://doi.org/10.1163/22238980-00001075 (2019).
Favre-Godal, Q., Gourguillon, L., Lordel-Madeleine, S., Gindro, K. & Choisy, P. Orchids and their mycorrhizal fungi: An insufficiently explored relationship. Mycorrhiza 30, 5–22. https://doi.org/10.1007/s00572-020-00934-2 (2020).
Ma, X., Kang, J., Nontachaiyapoom, S., Wen, T. & Hyde, K. Non-mycorrhizal endophytic fungi from orchids. Curr. Sci. 109, 72–87 (2015).
Sarsaiya, S., Shi, J. & Chen, J. A comprehensive review on fungal endophytes and its dynamics on Orchidaceae plants: Current research, challenges, and future possibilities. Bioengineered 10, 316–334. https://doi.org/10.1080/21655979.2019.1644854 (2019).
Adeleke, B. S. & Babalola, O. O. The plant endosphere-hidden treasures: A review of fungal endophytes. Biotechnol. Genet. Eng. Rev. 37, 154–177. https://doi.org/10.1080/02648725.2021.1991714 (2021).
Khalil, A. M. A., Abdelaziz, A. M., Khaleil, M. M. & Hashem, A. H. Fungal endophytes from leaves of Avicennia marina growing in semi-arid environment as a promising source for bioactive compounds. Lett. Appl. Microbiol. 72, 263–274. https://doi.org/10.1111/lam.13414 (2021).
Kristiansen, K. A., Freudenstein, J. V., Rasmussen, F. N. & Rasmussen, H. N. Molecular identification of mycorrhizal fungi in Neuwiedia veratrifolia (Orchidaceae). Mol. Phylogenet. Evol. 33, 251–258. https://doi.org/10.1016/j.ympev.2004.05.015 (2004).
Jacquemyn, H., Brys, R., Cammue, B. P., Honnay, O. & Lievens, B. Mycorrhizal associations and reproductive isolation in three closely related Orchis species. Ann. Bot. 107, 347–356. https://doi.org/10.1093/aob/mcq248 (2011).
Chen, J., Wang, H. & Guo, S. X. Isolation and identification of endophytic and mycorrhizal fungi from seeds and roots of Dendrobium (Orchidaceae). Mycorrhiza 22, 297–307. https://doi.org/10.1007/s00572-011-0404-0 (2012).
Fernández di Pardo, A. et al. Mycorrhizal fungi isolated from native terrestrial orchids of pristine regions in Cordoba (Argentina). Rev. Biol. Trop. 63, 275–283. https://doi.org/10.15517/rbt.v63i1.14226 (2015).
Legon, N. W., Henrici, A. (RBG Kew, 2022).
Adamčík, S., Ripková, S. & Zaliberová, M. Diversity of Russulaceae in the Vihorlatské vrchy Mts. (Slovakia). Czech Mycol. 58, 43–66 (2006).
Rog, I., Rosenstock, N. P., Körner, C. & Klein, T. Share the wealth: Trees with greater ectomycorrhizal species overlap share more carbon. Mol. Ecol. 29, 2321–2333. https://doi.org/10.1111/mec.15351 (2020).
Zhang, S. & Chen, J. Uncertainty in projection of climate extremes: A comparison of CMIP5 and CMIP6. J. Meteorol. Res. 35, 646–662. https://doi.org/10.1007/s13351-021-1012-3 (2021).
Kolanowska, M. et al. Global warming not so harmful for all plants - response of holomycotrophic orchid species for the future climate change. Sci. Rep. 7, 12704. https://doi.org/10.1038/s41598-017-13088-7 (2017).
Cavanaugh, K. C. et al. Poleward expansion of mangroves is a threshold response to decreased frequency of extreme cold events. Proc. Natl. Acad. Sci. U. S. A. 111, 723–727. https://doi.org/10.1073/pnas.1315800111 (2014).
Pélissié, M., Johansson, F. & Hyseni, C. Pushed northward by climate change: Range shifts with a chance of co-occurrence reshuffling in the forecast for northern European odonates. Environ. Entomol. 51, 910–921. https://doi.org/10.1093/ee/nvac056 (2022).
Hickling, R., Roy, D. B., Hill, J. K., Fox, R. & Thomas, C. D. The distributions of a wide range of taxonomic groups are expanding polewards. Glob. Change Biol. 12, 450–455. https://doi.org/10.1111/j.1365-2486.2006.01116.x (2006).
Evans, A. & Jacquemyn, H. Range size and niche breadth as predictors of climate-induced habitat change in Epipactis (Orchidaceae). Front. Ecol. Evol. 10, 78963 (2022).
Kolanowska, M., Rewicz, A. & Nowak, S. Significant habitat loss of the black vanilla orchid (Nigritella nigra sl, Orchidaceae) and shifts in its pollinators availability as results of global warming. Global Ecol. Conserv. 27, e01560. https://doi.org/10.1016/j.gecco.2021.e01560 (2021).
Kolanowska, M., Nowak, S. & Rewicz, A. Will Greenland be the last refuge for the continental European small-white orchid?Niche modeling of future distribution of Pseudorchis albida. Front. Environ. Sci. https://doi.org/10.3389/fevo.2022.894616 (2022).
Evans, A., Janssens, S. & Jacquemyn, H. Impact of climate change on the distribution of four closely related Orchis (Orchidaceae) species. Divers. Basel https://doi.org/10.3390/d12080312 (2020).
Charitonidou, M., Kougioumoutzis, K., Karypidou, M. C. & Halley, J. M. Fly to a safer north: Distributional shifts of the orchid Ophrys insectifera L. due to climate change. Biology 11, 497 (2022).
Wang, Q. et al. Effects of topography on structuring species assemblages in a subtropical forest. J. Plant Ecol. 10, 440–449. https://doi.org/10.1093/jpe/rtw047 (2017).
Iwara, A. I., Gani, B. S., Njar, G. N. & Deekor, T. N. Influence of soil physico-chemical properties on the distribution of woody tree/shrub species in south-southern Nigeria. J. Agric. Sci. 2, 69–75. https://doi.org/10.1080/09766898.2011.11884669 (2011).
John, R. et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. 104, 864–869. https://doi.org/10.1073/pnas.0604666104 (2007).
Chauvier, Y. et al. Influence of climate, soil, and land cover on plant species distribution in the European Alps. Ecol. Monogr. 91, e01433. https://doi.org/10.1002/ecm.1433 (2021).
Nardini, A., Andri, S. & Crasso, M. Influence of substrate depth and vegetation type on temperature and water runoff mitigation by extensive green roofs: Shrubs versus herbaceous plants. Urban Ecosyst. 15, 697–708. https://doi.org/10.1007/s11252-011-0220-5 (2012).
Zhang, Q.-P., Wang, J. & Wang, Q. Effects of abiotic factors on plant diversity and species distribution of alpine meadow plants. Ecol. Inf. 61, 101210. https://doi.org/10.1016/j.ecoinf.2021.101210 (2021).
Lobo, J. M. & Tognelli, M. F. Exploring the effects of quantity and location of pseudo-absences and sampling biases on the performance of distribution models with limited point occurrence data. J. Nat. Conserv. 19, 1–7. https://doi.org/10.1016/j.jnc.2010.03.002 (2011).
Inman, R., Franklin, J., Esque, T. & Nussear, K. Comparing sample bias correction methods for species distribution modeling using virtual species. Ecosphere 12, 03422. https://doi.org/10.1002/ecs2.3422 (2021).
Bird, T. J. et al. Statistical solutions for error and bias in global citizen science datasets. Biol. Cons. 173, 144–154. https://doi.org/10.1016/j.biocon.2013.07.037 (2014).
Stolar, J. & Nielsen, S. Accounting for spatially biased sampling effort in presence-only species distribution modelling. Divers. Distrib. 21, 595–608. https://doi.org/10.1111/ddi.12279 (2015).
Dubos, N. et al. Assessing the effect of sample bias correction in species distribution models. Ecol. Indic. 145, 109487. https://doi.org/10.1016/j.ecolind.2022.109487 (2022).
Araújo, M. B., Pearson, R. G., Thuiller, W. & Erhard, M. Validation of species–climate impact models under climate change. Glob. Change Biol. 11, 1504–1513. https://doi.org/10.1111/j.1365-2486.2005.01000.x (2005).
Kolanowska, M. & Jakubska-Busse, A. Is the lady’s-slipper orchid (Cypripedium calceolus) likely to shortly become extinct in Europe?-Insights based on ecological niche modelling. PLoS ONE 15, e0228420. https://doi.org/10.1371/journal.pone.0228420 (2020).
Kolanowska, M. The future of a montane orchid species and the impact of climate change on the distribution of its pollinators and magnet species. Global Ecol. Conserv. https://doi.org/10.1016/j.gecco.2021.e01939 (2021).
Kolanowska, M., Michalska, E. & Konowalik, K. The impact of global warming on the niches and pollinator availability of sexually deceptive orchid with a single pollen vector. Sci. Total Environ. https://doi.org/10.1016/j.scitotenv.2021.148850 (2021).
Slominski, A. H. & Burkle, L. A. Solitary bee life history traits and sex mediate responses to manipulated seasonal temperatures and season length. Front. Ecol. Evol. 7, 314 (2019).
O’Neill, K. M., Delphia, C. M. & O’Neill, R. P. Oocyte size, egg index, and body lipid content in relation to body size in the solitary bee Megachile rotundata. PeerJ 2, 314. https://doi.org/10.7717/peerj.314 (2014).
Sgolastra, F. et al. The long summer: Pre-wintering temperatures affect metabolic expenditure and winter survival in a solitary bee. J. Insect. Physiol. 57, 1651–1659. https://doi.org/10.1016/j.jinsphys.2011.08.017 (2011).
Schenk, M., Mitesser, O., Hovestadt, T. & Holzschuh, A. Overwintering temperature and body condition shift emergence dates of spring-emerging solitary bees. PeerJ 6, e4721. https://doi.org/10.7717/peerj.4721 (2018).
Bosch, J. & Kemp, W. P. Effect of wintering duration and temperature on survival and emergence time in males of the orchard pollinator Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 32, 711–716. https://doi.org/10.1603/0046-225X-32.4.711 (2003).
Forrest, J. R. K. & Thomson, J. D. An examination of synchrony between insect emergence and flowering in Rocky Mountain meadows. Ecol. Monogr. 81, 469–491. https://doi.org/10.1890/10-1885.1 (2011).
Bosch, J., Sgolastra, F. & Kemp, W. P. Timing of eclosion affects diapause development, fat body consumption and longevity in Osmia lignaria, a univoltine, adult-wintering solitary bee. J. Insect. Physiol. 56, 1949–1957. https://doi.org/10.1016/j.jinsphys.2010.08.017 (2010).
Pawlikowski, T. et al. Rising temperatures advance the main flight period of Bombus bumblebees in agricultural landscapes of the Central European Plain. Apidologie 51, 652–663. https://doi.org/10.1007/s13592-020-00750-9 (2020).
Phillips, R. D. et al. Mismatch in the distribution of floral ecotypes and pollinators: Insights into the evolution of sexually deceptive orchids. J. Evol. Biol. 28, 601–612. https://doi.org/10.1111/jeb.12593 (2015).
Hutchings, M. J., Robbirt, K. M., Roberts, D. L. & Davy, A. J. Vulnerability of a specialized pollination mechanism to climate change revealed by a 356-year analysis. Bot. J. Linn. Soc. 186, 498–509. https://doi.org/10.1093/botlinnean/box086 (2018).
Acknowledgements
In the graphical abstract photographs authored by Björn S. (L. abortivum), Hg6996 (R. delica), discoverthewild.co.uk (Russila choroides), Max Kasparek (R. septemdentatum), Easter Ecological Science Center (Anthophora sp.) and medianauka.pl (B. terrestris) were used and modified.
Author information
Authors and Affiliations
Contributions
M.K.: Conceptualization, Methodology, Data curation, Software, Data curation, Investigation, Visualization, Writing- Original draft preparation, Writing- Reviewing and Editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kolanowska, M. Loss of fungal symbionts and changes in pollinator availability caused by climate change will affect the distribution and survival chances of myco-heterotrophic orchid species. Sci Rep 13, 6848 (2023). https://doi.org/10.1038/s41598-023-33856-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-33856-y
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
