
Abstract
Human α2-macroglobulin (hα2M) is a large homotetrameric protein involved in the broad inhibition of endopeptidases. Following cleavage within a bait region, hα2M undergoes stepwise transitions from its native, expanded, highly flexible, active conformation to an induced, compact, triggered conformation. As a consequence, the peptidase is entrapped by an irreversible Venus flytrap mechanism. Given the importance of hα2M, biochemical studies galore over more than seven decades have attempted to ascertain its role, typically using authentic hα2M purified from frozen and non-frozen fresh blood plasma, and even outdated plasma. However, hα2M is sensitive once isolated and purified, and becomes heterogeneous during storage and/or freezing, raising concerns about the functional competence of frozen plasma-derived hα2M. We therefore used a combination of native and sodium dodecylsulfate polyacrylamide gel electrophoresis, affinity and ion-exchange chromatography, multi-angle laser light scattering after size-exclusion chromatography, free cysteine quantification, and peptidase inhibition assays with endopeptidases of two catalytic classes and three protein substrates, to characterize the biochemical and biophysical properties of hα2M purified ad hoc either from fresh plasma or frozen fresh plasma after thawing. We found no differences in the molecular or functional properties of the preparations, indicating that protective components in plasma maintain native hα2M in a functionally competent state despite freezing.
Similar content being viewed by others

Introduction
The α2-macroglobulin (α2M) family of large multi-domain proteins is found in animals and some colonizing bacteria1,2,3,4,5,6,7,8,9,10,11. The family is typified by human tetrameric α2M (hα2M), which is present at high concentrations in blood plasma. This protein appears to be essential because no total deficiency has been described, indicating that such a deficiency would be embryonically lethal12,13,14. The hα2M protein has many functions, including the transport of growth factors, cytokines, and hormones; the binding of misfolded and inactivated proteins as a chaperone; and the binding of metals5,9,15,16,17. In addition, the best-characterized function of hα2M is its broad-spectrum capacity to inhibit endopeptidases of the four major catalytic classes regardless of their specificity13. In this manner, hα2M regulates proteolysis in complex physiological contexts such as nutrition, haemostasis, signalling and tissue remodelling2,8,10,11, and mediates innate defence against external peptidases during envenomation and microbial infection8,10,18,19,20. Remarkably, the protein is an acute-phase protein in rodents21 but not in humans22.
Secreted hα2M is a glycosylated 1451-residue protein with 11 domains, which forms a ~ 675-kDa homotetramer comprising a dimer of disulfide-linked dimers. This large protein operates as a suicidal trap using a Venus flytrap mechanism, which is based on protomers adopting either an expanded or compact conformation23. In its native active state, hα2M is a large, highly flexible structure with the four protomers in the expanded conformation, forming wide openings of up to 70 × 50 Å that allow disparate plasma components to enter its large central lumen (~ 600 nm3). Peptidases that cleave within a flexible multi-target bait region within the hα2M bait region domain24 cause a massive conformational rearrangement of the tetramer, involving several intermediates with protomers in either the expanded or compact state. This eventually leads to an irreversibly induced or triggered state25, in which all four protomers are compact and the capacity to inhibit peptidases is lost23. This structure has a much smaller central lumen (~ 300 nm3) and features 12 narrow openings of up to 30 × 40 Å, through which the trapped prey peptidase cannot escape. Moreover, the native and triggered tetramers differ in size and shape23,26,27, which increases the mobility of the triggered form in native polyacrylamide gel electrophoresis (PAGE)28. Macromolecules that enter the native tetramer without cleaving the bait can leave by diffusion. Accordingly, hα2M functions as a molecular sieve, screening circulating proteins and selectively catching peptidases.
Once within hα2M, the trapped peptidase is still able to interact with small substrates and inhibitors that diffuse into the particle through the aforementioned narrow openings. Finally, the C-terminal receptor-binding domain of each protomer, which is cryptic in the native tetramer23,27, becomes exposed on the hα2M surface and is recognized by cell surface receptors such as the low-density lipoprotein receptor-related protein. This triggers receptor-mediated endocytosis and destruction of the hα2M–peptidase complex in the lysosomes29.
This sequence of events has been established by structural and biochemical studies over more than seven decades. The starting point of most of these studies was authentic hα2M isolated from human plasma because it is difficult to produce functional recombinant hα2M capable of peptidase inhibition30,31,32, with some notable exceptions33,34,35,36. Once isolated and purified, native hα2M tends to be instable in vitro37,38,39. These findings cast doubt on the suitability of thawed frozen fresh plasma (FR) as a source of native hα2M instead of non-frozen fresh plasma (NF), because the former might yield a heterogeneous, artefactual and damaged population of hα2M particles40. Here, we address this question by using a combination of biophysical and functional assays to compare the structure and functionality of hα2M prepared from FR or NF plasma.
Results and discussion
The overall conformation of native hα2M is unaltered by plasma freezing
The α2Ms belong to the wider thioester-containing protein family, which includes not only endopeptidase inhibitors but also complement proteins C3, C4 and C5; the cell surface antigen CD109; and thioester-containing proteins from animals10,19,41. A hallmark of these proteins is a C–X–E–Q motif (C972–Q975 in hα2M), which forms a relatively stable β-cysteinyl-γ-glutamyl thioester bond in the native state. Native hα2M protomer structures in the expanded conformation were determined by cryo-electron microscopy (cryo-EM), revealing that stability results from the location of the thioester bond in a hydrophobic cavity and its protection by the nearby receptor-binding domain in partially different conformations23,27,42.
Under physiological conditions, thioester hydrolysis occurs after the conformational rearrangement triggered by cleavage of the bait region, which exposes the thioester bond23,26,27. The latter may then be targeted by surface lysine residues of the trapped peptidase, which gives rise to an ε-(γ-glutamyl)-lysine that is covalently linked to hα2M, releasing a free cysteine (C972)43,44. However, this covalent entrapment is not essential for peptidase inhibition by hα2M45,46.
In the absence of proteolytic activation, high concentrations of small reactive nucleophiles such as hydroxylamine or methylamine (MA) can cleave the hidden thioester bond43,44,47. This causes a conformational rearrangement similar to (but slower to form than) the peptidase-induced structure, which is revealed by its greater mobility in native PAGE and distinct elution profile during size-exclusion chromatography (SEC). This reaction likewise gives rise to the free cysteine but the hα2M tetramer can no longer bind and inhibit peptidases, despite possessing an intact bait region47.
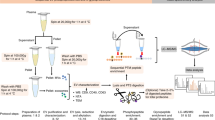
We compared the suitability of NF plasma and FR plasma, which is used for clinical transfusions as frozen fresh plasma preparations48, for the purification of hα2M using an established protocol based on polyethylene glycol (PEG) precipitation followed by zinc-affinity chromatography, ion-exchange chromatography (IEC) and SEC23,31,49,50. We analysed the zinc-affinity chromatography and IEC eluates by sodium dodecylsulfate PAGE (SDS-PAGE) (Fig. 1a) and compared the IEC profiles (Fig. 1b), revealing that the two hα2M preparations were equivalent. Next, we analysed the native and MA-treated samples by native PAGE and observed the aforementioned change in electrophoretic mobility, which was again indistinguishable between the two plasma sources (Fig. 1c). The MA-treated sample showed greater mobility than the native form in each case, as anticipated. Multi-angle laser light scattering after SEC (SEC–MALLS) showed the difference in migration between MA-treated and native samples, but the FR and NF preparations were again indistinguishable (Fig. 1d), also in terms of molecular mass before and after MA treatment (Fig. 1e). Overall, these results indicate that freezing blood plasma does not affect the conformation of hα2M, at least to the extent detectable using biophysical methods, and that hα2M from both sources appears well folded.

Purification and biophysical analysis of hα2M from non-frozen fresh plasma (NF) or frozen fresh plasma (FR). (a) SDS-PAGE analysis of elutes from zinc affinity chromatography (lanes 1 and 3) and IEC (lanes 2 and 4). (b) Comparison of the IEC profiles of native hα2M purified from NF (solid black line) and FR (dashed orange line). (c) Native PAGE analysis of native (lanes 1 and 3) and MA-treated induced (lanes 2 and 4) hα2M from NF (lanes 1 and 2) and FR plasma (lanes 3 and 4). (d) SEC–MALLS analysis of native (black and orange lines) and MA-treated induced (grey and blue lines) protein purified from FR (dashed lines) and NF (solid lines) plasma. (e) The molecular masses determined by SEC–MALLS were 671 kDa for the native tetrameric species and 675–678 kDa for the MA-treated species, which match the theoretical value (643.2 kDa) plus glycosylation. (f) Concentration of free thiol groups in hα2M from non-frozen (NF, white bars) or thawed frozen fresh (FR, grey bars) samples according to Ellmann’s reaction51. The protomeric hα2M concentration used was 157 μΜ. Data are presented as means ± SD (n = 3). Statistical significance was determined using a two-sided Student’s t-test (n.s., p > 0.1). Panels a-e are representative of more than three independent experimental replicates.
Freezing blood plasma does not affect hα2M thioester bonds
To test the state of the thioester bonds in the native and MA-treated samples from FR and NF plasma, we quantified the free cysteines using Ellman’s reaction51 (Fig. 1f). The stoichiometry detected in the native preparations (62 μM and 67 μΜ) was equivalent and corresponded to a baseline reaction with cysteines other than C972, putatively from plasma molecules bound by hα2M (see Conclusions). In contrast, the MA-treated samples revealed an abrupt increase in the free thiol contents (205 μM and 209 μΜ). These concentrations corresponded to > 90% of the protomer concentration once the baseline was subtracted (i.e., close to one thiol group per protomer), which indicates nearly complete cleavage of the thioester bond by MA and the subsequent liberation of the C972 Sγ-atoms44. The values from the FR and NR samples were equivalent within experimental error.
Plasma freezing does not impair the ability of native hα2M to inhibit peptidases
The Venus flytrap mechanism of inhibition is initiated by cleavage within the promiscuous bait region, which is exposed in the central lumen of native hα2M23,26,52,53. The efficiency of inhibition of hα2M is very high, so there is often a stoichiometric relationship between the number of bait regions cleaved and the molecules of peptidase inhibited. The limit is determined by the cavity size of the inner lumen of the induced tetramer, which can accommodate up to two peptidase molecules the size of trypsin23,26, and the overall shape of the peptidases. Sequestered peptidases remain able to cleave substrates small enough to access the lumen of the compact hα2M (and thus the active site of the bound peptidase) through any of the narrow openings.
We compared the stoichiometry of inhibition in the FR and NF protein preparations by titrating hα2M against two model endopeptidases from different catalytic classes at various molar ratios. We evaluated the serine peptidase trypsin from bovine pancreas and the metallopeptidase thermolysin from Bacillus thermoproteolyticus by testing their activity against a fluorogenic casein derivative (Fig. 2a,b). Both endopeptidases showed equivalent residual activities after preincubation with native hα2M from the two preparations at inhibitor tetramer:peptidase molar ratios of 2:1, 1:1 and 1:2. This reflected the ability of cleaved substrate fragments, still labelled with the fluorophore, to access the trapped peptidase, leading to the emission of fluorescent signal as previously reported54. At the highest molar ratio (1:4), the inhibitor was saturated and the excess peptidase therefore led to a sudden increase of proteolytic activity (Fig. 2a,b). Again, the FR and NF preparations were indistinguishable, as in the preceding experiments.

Inhibitory activity in vitro of hα2M from non-frozen fresh plasma (NF) or frozen fresh plasma (FR). Inhibition of (a) trypsin and (b) thermolysin activity against the fluorescent BODIPY FL-casein substrate. Percentages of remaining activity are represented as means ± SD (n = 3). Statistical significance was determined using a two-sided Student’s t-test (n.s., p > 0,1). (c–f) SDS-PAGE inhibition assay at different tetrameric hα2M:peptidase ratios but constant peptidase concentrations, showing the inhibition of trypsin activity against (c) α-casein and (e) fibrinogen, and the inhibition of thermolysin activity against (d) α-casein and (f) fibrinogen, for which only the Aα chain is cleaved. Lanes S and SP correspond to the substrate alone and the substrate incubated with and processed by the peptidase, respectively.
Finally, we analysed the ability of hα2M to inhibit trypsin and thermolysin cleavage of α-casein (32 kDa) and fibrinogen (340 kDa, divided into the three chains Aα, Bβ and γ), which are good substrates for both peptidases, by SDS-PAGE (Fig. 2c–f). We found that hα2M inhibited both peptidases at molar ratios between 1:1 and 1:2, and the FR and NF samples were once again indistinguishable. Overall, these results indicate that freezing plasma does not affect the inhibitory activity of hα2M, suggesting it remains in its functional native conformation.
Repeated freezing and thawing cycles of purified hα2M induce sample heterogeneity
It has been reported that, once purified, native hα2M is more susceptible to freeze and thaw39. To assess this in the complex context of plasma, we analysed the biophysical and functional properties of pure native hα2M after several fast (Fig. 3a) or slow (Fig. 3c,d) freeze-and-thaw cycles (FTs). In agreement with39, fast freezing in liquid nitrogen in up to three FTs apparently did not significantly impair the protein and high-molecular-weight species, which are indicative of conformational changes as shown in Fig. 1 of39, appeared only after the third cycle (Fig. 3a). Contrary to39, however, we did not detect induced species, which we attribute to the presence of sodium azide in these published studies. This reagent is a chemical preservative and potent nucleophile55, which can attack the thioester bond and induce rearrangement in a similar fashion to MA. In contrast, slow FTs already caused biophysical heterogeneity after the first cycle (Fig. 3c). This correlated with an associated slight loss of inhibitory capacity against thermolysin (Fig. 3d).

Effect of long-term storage and freezing/thawing of native hα2M. (a, c) Native PAGE analysis of native hα2M from FR samples stored for (a) 1 month or 2 years or (c) 3 months at – 30 °C. Native hα2M samples were further subjected to seven fast (a) or slow (b) freeze–thaw cycles (FTx). This procedure causes the emergence of high-molecular-weight (HMW) species, as previously reported for protein stored for several months at 4 °C39. MA-treated hα2M was used as a control for induced hα2M. (b, d) Inhibition of thermolysin activity against the fluorescent substrate BODIPY FL-casein by (b) hα2M from FR samples stored for 1 month or 2 years at – 30 °C and (d) hα2M samples subjected to freeze–thaw cycles. The percentages of remaining activity are represented as means ± SD (b: n = 3; d: n = 6). MA-treated hα2M is representative of the inactive inhibitor.
Finally, we tested whether long-term storage at -30ºC impaired native hα2M, as the source of several past studies had been outdated plasma, which corresponds to frozen fresh plasma after three years. We assessed the above biophysical and functional properties of hα2M purified from FR stored for 2 years at – 30 °C (Fig. 3a,b). No significant differences could be detected when compared with the results after one month storage.
Conclusions
The structure and biochemistry of hα2M, an essential multimeric and multi-domain protein with many physiological functions and orthologs across animal species, has been investigated for decades. However, structural analysis has been hampered by the conformational variability of the native state23,27,28. In addition, early reports showed that native preparations featured serine peptidase activity56. This suggested the presence of induced, peptidase laden hα2M particles, which were also identified by cryo-EM, supporting clinical studies that detected peptidase-complexed hα2M in the plasma of healthy individuals13. These structural studies identified several reaction intermediates between the fully native and fully induced conformations in theoretically native purified hα2M from FR and NF plasma23,27, disputing claims that native hα2M purified from NF plasma is homogeneous40.
The conformational variability of native hα2M in our cryo-EM study, which was based on the use of FR plasma23, raised concerns about its functionality40. This reflected the reported fact that purified hα2M is sensitive to storage, lyophilisation and freezing37,38,39 due to a stress-induced conformational rearrangement that renders the proteins inactive or dysfunctional57. To address this issue, we compared the biophysical and functional properties of hα2M from FR and NF, which was freshly prepared for immediate use following purification as in our structural studies23, and was not stored, frozen or lyophilized. Moreover, treatment of hα2M from FR with trypsin had resulted in a homogeneously induced sample in cryo-EM23.
Our results unambiguously showed that the two protein preparations were identical in terms of biochemical, biophysical and functional properties. This indicates that blood plasma shields hα2M against stress caused by freeze-thawing. In plasma, hα2M is surrounded by other macromolecules at high concentrations of 60–80 mg/mL58. This environment has a cryoprotective effect based on macromolecular crowding, which enhances the native structure and stability of proteins59. Specifically, plasma from healthy donors has a glucose content of 0.7–1.4 g/L60, which is consistent with the protective effect of sucrose on purified hα2M39. Moreover, plasma contains large amounts of serum albumin (60% of the total protein content), which plays an important role in modulating osmotic pressure. Its bovine counterpart is widely used as an additive for the preservation of tissues, cells and proteins such as lactate dehydrogenase61 and catalase62 during freezing.
The equivalence of our results with FR and NF plasma is also consistent with the fact that the plasma used in our studies over the last decade23,26,31,50 was obtained, fresh or frozen, from the only authorized blood bank in Catalonia and was not outdated. It conforms to the quality criteria of therapeutic fresh frozen plasma48, which is administered immediately after thawing to treat coagulation disorders63. The relevance of this reagent is highlighted by its inclusion on the WHO list of essential medicines64. Moreover, early clinical studies reported that fresh frozen plasma compensates for the loss of hα2M that occurs during early stages of pancreatitis65. These data together confirm that hα2M from FR plasma is functionally competent.
In summary, the storage of native hα2M in FR plasma before purification does not alter its properties. Indeed, FR plasma has been used in several laboratories worldwide for decades to prepare native hα2M for biochemical and structural studies23,26,31,32,50,66,67,68,69,70,71. Often, the FR plasma was outdated, which we here also found to be functionally competent. FR plasma therefore appears suitable for native hα2M preparation, as previously discussed38, and can be envisaged as a source of hα2M on demand without needing to follow the time-consuming procedures from blood banks or to rely on donations each time the protein is required.
Methods
Preparation and purification of native and MA-treated hα2M
All experiments were performed according to the applying guidelines and regulations and were approved by the Committee for Ethics in Research with Medicines and the Commission of Research Projects of the Vall d’Hebron University Hospital (Barcelona). NF plasma was obtained < 24 h after collection from anonymous healthy human donors (LST-BIOBANC, Banc de Sang i Teixits, National Government of Catalonia), so that no consent from the subjects was required. Each sample was anonymized and checked to be free of hepatitis viruses (HVB and HVC), Treponema pallidum ssp. pallidum (the syphilis pathogen) and human immunodeficiency virus (HIV). This plasma is suitable for transfusion into patients, is routinely stored at − 30 °C in the blood bank, and has a shelf-life of 3 years, thus conforming to the clinical standards of therapeutic fresh frozen plasma. Half of the sample was used immediately for purification without freezing (NF plasma) and the rest was frozen at − 20 °C within 1 h, stored at this temperature for 16 h and then used for purification (FR plasma) following the procedure of previous cryo-EM studies23. Subsequent sequential precipitation steps with 4–12% PEG 4000 were carried out at 4 °C as previously described50. The final precipitate containing hα2M was reconstituted in 20 mM sodium phosphate buffer (pH 6.8) supplemented with 5 mM phenylmethylsulfonyl fluoride. All subsequent purification steps were performed at 4 °C. The hα2M was first captured on a zinc-chelating resin (G-Biosciences) in an open column (Bio-Rad) and was then washed with 50 mM sodium phosphate buffer (pH 7.2) containing 250 mM sodium chloride and 10 mM imidazole before elution in 50 mM sodium phosphate buffer (pH 7.2) containing 250 mM sodium chloride and 100 mM ethylenediaminetetraacetate (EDTA). The eluate was exchanged to buffer A (20 mM sodium phosphate, pH 7.4) on a PD-10 column (Cytiva) followed by IEC on a TSKgel DEAE-2SW column (TOSOH Bioscience) equilibrated with buffer A. Fractions were eluted in 30 mL buffer B (20 mM sodium phosphate, 1 M sodium chloride, pH 7.4) applied as a 2–50% gradient vs. buffer A, and the central peak fractions were pooled and concentrated. Final purification was carried out by SEC on a Superose 6 10/300 column (GE Healthcare Life Sciences) in buffer C (20 mM Tris–HCl, 150 mM sodium chloride, pH 7.4). MA-treated hα2M (MA-hα2M) was obtained by reacting native hα2M, buffer-exchanged to 100 mM Tris–HCl (pH 8) after the IEC purification step, with 200 mM methylamine hydrochloride overnight at 4 °C, followed by SEC as described for the native counterpart.
Sample freezing and thawing
Purified native hα2M was subjacted to fast or slow freeze–thaw (FT) cycles. Fast FT was performed by freezing in liquid nitrogen and thawing in a water bath at room temperature, while slow FT was performed by directly placing the samples at – 20 °C (freezing) and on ice (thawing).
Proteolytic inhibition assays
Purified native hα2M was used to study the inhibition of bovine pancreatic trypsin and Bacillus thermoproteolyticus thermolysin (both from Sigma-Aldrich) after incubation for 10 min at room temperature. Reactions were carried out in buffer C using tetrameric hα2M:peptidase ratios of 2:1, 1:1, 1:2 and 1:4. Reaction products were used directly to monitor residual peptidase activity against fluorogenic and natural substrates at 37 °C. For the fluorogenic substrates, the activity of 50 nM peptidase was tested in a reaction volume of 100 μL in a Synergy H1 microplate fluorimeter (Biotek) using the EnzCheck Assay Kit, which includes 5 μg/mL BODIPY FL-casein (λex = 505 nm and λem = 513 nm; Invitrogen). As natural substrates, we used α-casein from bovine milk (35 kDa) and fibrinogen from human plasma (340 kDa) at a concentration of 0.5 mg/mL (both from Sigma-Aldrich) with trypsin and thermolysin at 100 nM and 5 nM, respectively. Reactions were monitored for 10 min (thermolysin) or 90 min (trypsin), and cleavage was assessed by 10–14% Tricine-SDS-PAGE after stopping the reactions with small-molecule inhibitors (0.7 mM Pefabloc SC from Roche Life Sciences for trypsin and 20 mM EDTA for thermolysin) and subsequent heating for 5 min at 95 °C.
SEC–MALLS
Samples were assessed in a Dawn Helios II device (Wyatt Technologies) coupled to a Superose 6 10/300 Increase SEC column (Cytiva) at the joint IBMB/IRB Crystallography Platform, Barcelona Science Park (Catalonia). The column was equilibrated in buffer C at 25 °C to analyse native and induced hα2M from FR and NF plasma. Data were processed and analysed using ASTRA 7 software (Wyatt Technologies) and a dn/dc value typical for proteins (0.185 mL/g).
Determination of free sulfhydryl groups
Preparations of native hα2M (after IEC) and MA-hα2M were exchanged to buffer C at a final concentration of 25 mg/mL, and free sulfhydryl groups were determined by reaction with Ellman’s reagent (5,5′-dithiobis-2-nitrobenzoic acid; Sigma)51 for 15 min. The change in A412 was monitored using a Power-Wave XS microplate spectrophotometer (Biotek). The concentration of free-thiol groups was calculated based on the molar extinction coefficient of Ellman’s reagent (14,150 M−1 cm−1;72), as previously done for hα2M samples by others71. The absorbance signal was measured in 96-well plates containing 200 μL of Ellman’s assay samples plus 20 μL of cysteine or test samples in triplicate.
Miscellaneous
Protein identity and purity were assessed by 10–14% Tris–Glycine SDS-PAGE stained with Coomassie-brilliant blue using Unstained Protein Molecular Weight Marker (10–200 kDa; Thermo Fisher Scientific) and BlueStar Plus Prestained Protein Marker (10–240 kDa; NIPPON Genetics) as molecular-mass markers. Native protein samples were also analysed by native NuPAGE 3–8% Tris–Acetate Mini Protein Gels (Invitrogen) stained with Coomassie-brilliant blue, and NativeMark Unstained Protein Standard (20-to-1200 kDa; Invitrogen) was used as molecular-mass marker.
Ultrafiltration steps were performed with Vivaspin 15, Vivaspin 2 and Vivaspin 500 filter devices with cut-off values ranging from 50 to 100 kDa (Sartorius Stedim Biotech). Protein concentrations were estimated by measuring A280 values in a NanoDrop spectrophotometer and applying the corresponding theoretical extinction coefficients. Concentrations were also measured using the BCA Protein Assay Kit (Thermo Fisher Scientific) with bovine serum albumin as a standard.
Data availability
All data and reagents are freely available from Dr. Gomis-Rüth upon reasonable request.
References
Cohn, E. J. et al. Preparation and properties of serum and plasma proteins. IV. A system for the separation into fractions of the protein and lipoprotein components of biological tissues and fluids. J. Am. Chem. Soc. 68, 459–475 (1946).
Barrett, A. J. & Starkey, P. M. The interaction of α2-macroglobulin with proteinases. Characteristics and specificity of the reaction, and a hypothesis concerning its molecular mechanism. Biochem. J. 133, 709–724 (1973).
Budd, A., Blandin, S., Levashina, E. A. & Gibson, T. J. Bacterial α2-macroglobulins: Colonization factors acquired by horizontal gene transfer from the metazoan genome?. Genome Biol. 5, R38. https://doi.org/10.1186/gb-2004-5-6-r38 (2004).
Doan, N. & Gettins, P. G. W. α-Macroglobulins are present in some Gram-negative bacteria: Characterization of the α2-macroglobulin from Escherichia coli. J. Biol. Chem. 283, 28747–28756. https://doi.org/10.1074/jbc.M803127200 (2008).
Ignjatovic, V., Mertyn, E. & Monagle, P. The coagulation system in children: Developmental and pathophysiological considerations. Semin. Thromb. Hemost. 37, 723–729. https://doi.org/10.1055/s-0031-1297162 (2011).
Rehman, A. A., Ahsan, H. & Khan, F. H. α2-Macroglobulin: A physiological guardian. J. Cell. Physiol. 228, 1665–1675. https://doi.org/10.1002/jcp.24266 (2013).
Wong, S. G. & Dessen, A. Structure of a bacterial α2-macroglobulin reveals mimicry of eukaryotic innate immunity. Nat. Commun. 5, 4917. https://doi.org/10.1038/ncomms5917 (2014).
Goulas, T. et al. Structural and functional insight into pan-endopeptidase inhibition by α2-macroglobulins. Biol. Chem. 398, 975–994. https://doi.org/10.1515/hsz-2016-0329 (2017).
Seddighi, S., Varma, V. & Thambisetty, M. α2-Macroglobulin in Alzheimer’s disease: New roles for an old chaperone. Biomark. Med. 12, 311–314. https://doi.org/10.2217/bmm-2018-0027 (2018).
Garcia-Ferrer, I., Marrero, A., Gomis-Rüth, F. X. & Goulas, T. α2-Macroglobulins: Structure and function. Subcell. Biochem. 83, 149–183. https://doi.org/10.1007/978-3-319-46503-6_6 (2017).
Starkey, P. M. & Barrett, A. J. Inhibition by α-macroglobulin and other serum proteins. Biochem. J. 131, 823–831 (1973).
Roberts, R. C. Protease inhibitors of human plasma. Alpha2-macroglobulin. J. Med. 16, 129–224 (1985).
Zucker, S., Lysik, R. M., Zarrabi, M. H., Fiore, J. J. & Strickland, D. K. Proteinase-α2-macroglobulin complexes are not increased in plasma of patients with cancer. Int. J. Cancer 48, 399–403. https://doi.org/10.1002/ijc.2910480316 (1991).
Borth, W. α2-Macroglobulin, a multifunctional binding protein with targeting characteristics. FASEB J. 6, 3345–3353 (1992).
Gonias, S. L. et al. α2-macroglobulin and the α2-macroglobulin receptor/LRP. A growth regulatory axis. Ann. N. Y. Acad. Sci. 737, 273–290 (1994).
Chu, C. T. & Pizzo, S. V. Interactions between cytokines and α2-macroglobulin. Immunol. Today 12, 249 (1991).
Liu, N.-M. et al. Transcuprein is a macroglobulin regulated by copper and iron availability. J. Nutr. Biochem. 18, 597–608. https://doi.org/10.1016/j.jnutbio.2006.11.005 (2007).
Armstrong, P. B. Proteases and protease inhibitors: A balance of activities in host-pathogen interaction. Immunobiology 211, 263–281. https://doi.org/10.1016/j.imbio.2006.01.002 (2006).
Armstrong, P. B. & Quigley, J. P. α2-macroglobulin: An evolutionarily conserved arm of the innate immune system. Dev. Comp. Immunol. 23, 375–390. https://doi.org/10.1016/s0145-305x(99)00018-x (1999).
Dubin, G., Koziel, J., Pyrc, K., Wladyka, B. & Potempa, J. Bacterial proteases in disease—role in intracellular survival, evasion of coagulation/fibrinolysis innate defenses, toxicoses and viral infections. Curr. Pharm. Des. 19, 1090–1113 (2013).
Jennings, G., Cruickshank, A. M., Shenkin, A., Wight, D. G. & Elia, M. Effect of aseptic abscesses in protein-deficient rats on the relationship between interleukin-6 and the acute-phase protein, alpha2-macroglobulin. Clin. Sci. (Lond.) 83, 731–735. https://doi.org/10.1042/cs0830731 (1992).
Saso, L., Silvestrini, B., Guglielmotti, A., Lahita, R. & Cheng, C. Y. Abnormal glycosylation of α2-macroglobulin, a non-acute-phase protein in patients with autoimmune diseases. Inflammation 17, 465–479. https://doi.org/10.1007/BF00916586 (1993).
Luque, D. et al. Cryo-EM structures show the mechanistic basis of pan-peptidase inhibition by human α2-macroglobulin. Proc. Natl. Acad. Sci. USA 119, e2200102119. https://doi.org/10.1073/pnas.2200102119 (2022).
Sottrup-Jensen, L. et al. Primary structure of the ‘bait’ region for proteinases in a2-macroglobulin. Nature of the complex. FEBS Lett. 127, 167–173. https://doi.org/10.1016/0014-5793(81)80197-4 (1981).
Haverback, B. J., Dyce, B., Bundy, H. F., Wirtschafter, S. K. & Edmondson, H. A. Protein binding of pancreatic proteolytic enzymes. J. Clin. Invest. 41, 972–980. https://doi.org/10.1172/JCI104576 (1962).
Marrero, A. et al. The crystal structure of human α2-macroglobulin reveals a unique molecular cage. Angew. Chem. Int. Ed. 51, 3340–3344 (2012).
Huang, X. et al. Cryo-EM structures reveal the dynamic transformation of human alpha-2-macroglobulin working as a protease inhibitor. Sci. China-Life Sci. 6, 5. https://doi.org/10.1007/s11427-11022-12139-11422 (2022).
Saunders, R., Dyce, B. J., Vannier, W. E. & Haverback, B. J. The separation of alpha-2 macroglobulin into five components with differing electrophoretic and enzyme-binding properties. J. Clin. Invest. 50, 2376–2383. https://doi.org/10.1172/JCI106736 (1971).
Andersen, O. M. et al. Specific binding of α-macroglobulin to complement-type repeat CR4 of the low-density lipoprotein receptor-related protein. Biochemistry 39, 10627–10633 (2000).
Suda, S. A., Dolmer, K. & Gettins, P. G. W. Critical role of asparagine 1065 of human α2-macroglobulin in formation and reactivity of the thiol ester. J. Biol. Chem. 272, 31107–31112. https://doi.org/10.1074/jbc.272.49.31107 (1997).
Marino-Puertas, L., Del Amo-Maestro, L., Taules, M., Gomis-Rüth, F. X. & Goulas, T. Recombinant production of human α2-macroglobulin variants and interaction studies with recombinant G-related α2-macroglobulin binding protein and latent transforming growth factor-β2. Sci. Rep. 9, 9186. https://doi.org/10.1038/s41598-019-45712-z (2019).
Arandjelovic, S., van Sant, C. L. & Gonias, S. L. Limited mutations in full-length tetrameric human α2-macroglobulin abrogate binding of platelet-derived growth factor-BB and transforming growth factor-β1. J. Biol. Chem. 281, 17061–17068. https://doi.org/10.1074/jbc.M602217200 (2006).
Boel, E. et al. Expression of human α2-macroglobulin cDNA in baby hamster kidney fibroblasts: Secretion of high levels of active α2-macroglobulin. Biochemistry 29, 4081–4087. https://doi.org/10.1021/bi00469a009 (1990).
Van Rompaey, L., Van den Berghe, H. & Marynen, P. Synthesis of a Cys949Tyr α2-macroglobulin thiol ester mutant: Co-transfection with wild-type α2-macroglobulin in an episomal expression system. Biochem. J. 312(Pt 1), 183–190. https://doi.org/10.1042/bj3120183 (1995).
Ikai, A. et al. A recombinant bait region mutant of human α2-macroglobulin exhibiting an altered proteinase-inhibiting spectrum. Cytotechnology 31, 53–60. https://doi.org/10.1023/A:1008011919876 (1999).
Harwood, S. L. et al. Development of selective protease inhibitors via engineering of the bait region of human α2-macroglobulin. J. Biol. Chem. 297, 100879. https://doi.org/10.1016/j.jbc.2021.100879 (2021).
Jones, J. M., Creeth, J. M. & Kekwick, R. A. Thio reduction of human α2-macroglobulin. The subunit structure. Biochem. J. 127, 187–197. https://doi.org/10.1042/bj1270187 (1972).
Salvesen, G. & Enghild, J. J. [7] α-Macroglobulins: Detection and characterization. Meth. Enzymol. 223, 121–141. https://doi.org/10.1016/0076-6879(93)23041-K (1993).
Wyatt, A. R., Kumita, J. R., Farrawell, N. E., Dobson, C. M. & Wilson, M. R. Alpha-2-macroglobulin Is acutely sensitive to freezing and lyophilization: implications for structural and functional studies. PLoS ONE 10, e0130036. https://doi.org/10.1371/journal.pone.0130036 (2015).
Harwood, S. L., Andersen, G. R. & Enghild, J. J. Recent cryoEM structures of human A2M may not be representative of the native protein. Proc. Natl. Acad. Sci. U. S. A. 119, e2210218119. https://doi.org/10.1073/pnas.2210218119 (2022).
Theopold, U. & Schmid, M. Thioester-containing proteins: At the crossroads of immune effector mechanisms. Virulence 8, 1468–1470. https://doi.org/10.1080/21505594.2017.1355662 (2017).
Luque, D. et al. Reply to Harwood et al.: Alternative functional conformations of native human α2-macroglobulin. Proc. Natl. Acad. Sci. U. S. A. 119, e2211048119. https://doi.org/10.1073/pnas.2211048119 (2022).
Sottrup-Jensen, L., Petersen, T. E. & Magnusson, S. A thiol-ester in α2-macroglobulin cleaved during proteinase complex formation. FEBS Lett. 121, 275–279. https://doi.org/10.1016/0014-5793(80)80361-9 (1980).
Sottrup-Jensen, L. Role of internal thiol esters in the α-macroglobulin-proteinase binding mechanism. Ann. N. Y. Acad. Sci. 737, 172–187 (1994).
Enghild, J. J., Salvesen, G., Thøgersen, I. B. & Pizzo, S. V. Proteinase binding and inhibition by the monomeric α-macroglobulin rat α1-inhibitor-3. J. Biol. Chem. 264, 11428–11435 (1989).
Travis, J. & Salvesen, G. S. Human plasma proteinase inhibitors. Annu. Rev. Biochem. 52, 655–709. https://doi.org/10.1146/annurev.bi.52.070183.003255 (1983).
Steinbuch, M., Pejaudier, L., Quentin, M. & Martin, V. Molecular alteration of α2-macroglobulin by aliphatic amines. Biochim. Biophys. Acta Prot. Struct. 154, 228–231. https://doi.org/10.1016/0005-2795(68)90277-8 (1968).
De Backer, D. et al. Guidelines for the use of fresh frozen plasma. Acta Clin. Belg. 63, 381–390. https://doi.org/10.1179/acb.2008.079 (2008).
Marrero, A., Mallorquí-Fernández, G., Guevara, T., García-Castellanos, R. & Gomis-Rüth, F. X. Unbound and acylated structures of the MecR1 extracellular antibiotic-sensor domain provide insights into the signal-transduction system that triggers methicillin resistance. J. Mol. Biol. 361, 506–521 (2006).
Goulas, T., Garcia-Ferrer, I., García-Piqué, S., Sottrup-Jensen, L. & Gomis-Rüth, F. X. Crystallization and preliminary X-ray diffraction analysis of eukaryotic α2-macroglobulin family members modified by methylamine, proteases and glycosidases. Mol. Oral Microbiol. 29, 354–364. https://doi.org/10.1111/omi.12069 (2014).
Ellman, G. L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 82, 70–77. https://doi.org/10.1016/0003-9861(59)90090-6 (1959).
Sottrup-Jensen, L. α-Macroglobulins: Structure, shape, and mechanism of proteinase complex formation. J. Biol. Chem. 264, 11539–11542 (1989).
Sottrup-Jensen, L., Sand, O., Kristensen, L. & Fey, G. H. The α-macroglobulin bait region. Sequence diversity and localization of cleavage sites for proteinases in five mammalian α-macroglobulins. J. Biol. Chem. 264, 15781–15789 (1989).
Thompson, V. F., Saldana, S., Cong, J. & Goll, D. E. A BODIPY fluorescent microplate assay for measuring activity of calpains and other proteases. Anal. Biochem. 279, 170–178. https://doi.org/10.1006/abio.1999.4475 (2000).
Tashrifi, Z., Khanaposhtani, M. M., Larijani, B. & Mahdavi, M. Sodium azide: An inorganic nitrogen source for the synthesis of organic N-compounds. ChemistrySelect 6, 13382–13723. https://doi.org/10.1002/slct.202103271 (2021).
Dyce, B. J., Wong, T., Adham, N., Mehl, J. & Haverback, B. J. Human plasma kallikrein esterase associated with alpha-2-macroglobulin binding protein. Clin. Res. 15, 101–101 (1967).
Bhatnagar, B. S., Bogner, R. H. & Pikal, M. J. Protein stability during freezing: Separation of stresses and mechanisms of protein stabilization. Pharm. Dev. Technol. 12, 505–523. https://doi.org/10.1080/10837450701481157 (2007).
Leeman, M., Choi, J., Hansson, S., Storm, M. U. & Nilsson, L. Proteins and antibodies in serum, plasma, and whole blood-size characterization using asymmetrical flow field-flow fractionation (AF4). Anal. Bioanal. Chem. 410, 4867–4873. https://doi.org/10.1007/s00216-018-1127-2 (2018).
Stagg, L., Zhang, S. Q., Cheung, M. S. & Wittung-Stafshede, P. Molecular crowding enhances native structure and stability of α/β protein flavodoxin. Proc. Natl. Acad. Sci. USA 104, 18976–18981. https://doi.org/10.1073/pnas.0705127104 (2007).
Bolli, G. B. & Fanelli, C. G. Physiology of glucose counterregulation to hypoglycemia. Endocrinol. Metab. Clin. North Am. 28, 467–493. https://doi.org/10.1016/s0889-8529(05)70083-9 (1999).
Honjoh, K. I. et al. Cryoprotective activities of group 3 late embryogenesis abundant proteins from Chlorella vulgaris C-27. Biosci. Biotechnol. Biochem. 64, 1656–1663. https://doi.org/10.1271/bbb.64.1656 (2000).
Hara, M., Terashima, S. & Kubois, T. Characterization and cryoprotective activity of cold-responsive dehydrin from Citrus unshiu. J. Plant Physiol. 158, 1333–1339. https://doi.org/10.1078/0176-1617-00600 (2001).
O’Shaughnessy, D. F. et al. Guidelines for the use of fresh-frozen plasma, cryoprecipitate and cryosupernatant. Br. J. Haematol. 126, 11–28. https://doi.org/10.1111/j.1365-2141.2004.04972.x (2004).
Samukange, W. T., Gardarsdottir, H., Leufkens, H. G. M. & Mantel-Teeuwisse, A. K. Selection of blood, blood components, and blood products as essential medicines in 105 low- and middle-income countries. Transfus. Med. Rev. 34, 94–100. https://doi.org/10.1016/j.tmrv.2019.10.005 (2020).
Leese, T. FFP in pancreatitis. Br. J. Surg. 74, 75. https://doi.org/10.1002/bjs.1800740129 (1987).
Imber, M. J. & Pizzo, S. V. Clearance and binding of two electrophoretic “fast” forms of human α2-macroglobulin. J. Biol. Chem. 256, 8134–8139 (1981).
Björk, I. & Fish, W. W. Evidence for similar conformational changes in α2-macroglobulin on reaction with primary amines or proteolytic enzymes. Biochem. J. 207, 347–356 (1982).
Enghild, J. J., Salvesen, G., Brew, K. & Nagase, H. Interaction of human rheumatoid synovial collagenase (matrix metalloproteinase 1) and stromelysin (matrix metalloproteinase 3) with human α2-macroglobulin and chicken ovostatin. Binding kinetics and identification of matrix metalloproteinase cleavage sites. J. Biol. Chem. 264, 8779–8785 (1989).
Dubin, A., Potempa, J. & Travis, J. Isolation of nine human plasma proteinase inhibitors by sequential affinity chromatography. Prep. Biochem. 20, 63–74. https://doi.org/10.1080/00327489008050177 (1990).
Björk, I., Larsson, L. J., Lindblom, T. & Raub, E. Stoichiometry of reactions of α2-macroglobulin with trypsin and chymotrypsin. Biochem. J. 217, 303–308. https://doi.org/10.1042/bj2170303 (1984).
Steiner, J. P., Migliorini, M. & Strickland, D. K. Characterization of the reaction of plasmin with α2-macroglobulin: Effect of antifibrinolytic agents. Biochemistry 26, 8487–8495. https://doi.org/10.1021/bi00399a068 (1987).
Riddles, P. W., Blakeley, R. L. & Zerner, B. Reassessment of Ellman’s reagent. Meth. Enzymol. 91, 49–60. https://doi.org/10.1016/s0076-6879(83)91010-8 (1983).
Acknowledgements
We are grateful to Mònica Buxaderas and Joan Pous from the joint IBMB/IRB Automated Crystallography Platform for assistance with SEC-MALLS. This study was supported in part by grants from Spanish and Catalan public and private bodies (Grant/fellowship references PID2019-107725RG-I00, PID2021-128682OA-I00, BES-2016-076877 and PID2022-137827OB-I00 from the State Agency of Research [MICIN/AEI/https://doi.org/10.13039/501100011033; may include “Next Generation EU/PRTR” and/or “cofunded by the EU” and/or “ERDF—A way of making Europe” and/or “ESF Investing in your future”]; 2017SGR3, 2021SGR00432 and Fundació La Marató de TV3 201815). The authors thank Richard M. Twyman for editing the manuscript.
Author information
Authors and Affiliations
Contributions
F.X.G.R. and T.G. conceived and supervised the project; S.R.M. purified proteins and performed in vitro studies with the help of T.G.; and F.X.G.R. and T.G. wrote the manuscript with contributions from S.R.M.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mendes, S.R., Gomis-Rüth, F.X. & Goulas, T. Frozen fresh blood plasma preserves the functionality of native human α2-macroglobulin. Sci Rep 13, 4579 (2023). https://doi.org/10.1038/s41598-023-31800-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-31800-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
