
Abstract
Sowing is widely used for the restoration of species-rich grasslands but still there are knowledge gaps regarding the most suitable application of different seed mixtures. We tested the effect of seed mixtures application timing on the establishment of sown forbs and weed control. 36 experimental plots with nine sowing treatments were established in an abandoned cropland in Hungary. Grass-seeds, diverse forb seed mixture and the combination of the two were applied: diverse forb mixture was sown simultaneously or 1, 2 or 3 years after grass sowing, in plots sown previously with grass or in empty plots (fallows). All sowing treatments supported the rapid establishment of the sown species in large cover and hampered weed encroachment. Forbs performed better when sown into fallows than in grass-matrix and forbs establishment was worse in older fallows than in younger ones. Grasses expressed a strong priority effect, especially when forbs were sown at least two years later than grasses. We also investigated the relation between seed germinability, weather parameters and establishment success. Germination rate in the greenhouse could not predict the establishment success of forbs in the field and showed great differences between years, hence we recommend sowing target forbs in multiple years.
Similar content being viewed by others


Introduction
Halting degradation of terrestrial ecosystems by various restoration measures is in the focus of the United Nations Decade of Ecosystem Restoration (2021–2030). Countries all over the world should contribute by rehabilitating and restoring degraded ecosystems, including grasslands, to prevent or at least mitigate the catastrophic consequences of climate change and loss of biodiversity1.
Grasslands are among the most species-rich ecosystems in the word2 and can be substantial carbon sinks in the changing climate1,3. They also provide important ecosystem services such as water supply and regulation, soil protection, pollination, biomass production and cultural services4. The diversity of grasslands is positively related to their stability and resistance against disturbances and to the ecosystem services provided5,6,7. Halting their degradation, decreasing their fragmentation and increasing their area, as well as conserving their diversity are as important as afforestation principles8,9. By restoring species-rich grasslands not only plant diversity can be increased, but also habitat heterogeneity, which in turn facilitates restoration of other life forms10. These targets can be achieved both by restoration of degraded species-rich grassland ecosystems11 and by grassland recreation in abandoned croplands12,13.
The abandonment of croplands is a global phenomenon14,15 that is especially typical in the Northern Hemisphere16. In the European Union 11% of all croplands is considered to have high likeliness to be abandoned in the period of 2015–203017. Besides socio-economic changes, the main reasons of abandonment are the inadequate edaphic and climatic conditions for crop production15,18,19,20.
The low level of abiotic (i.e., high nutrient availability) and biotic (i.e., low competitive pressure) filtering in abandoned croplands allows the establishment of weedy vegetation from the weed-contaminated soil seed bank21,22,23. Weed encroachment provides ecosystem disservices that can also affect the neighbouring natural habitats and agricultural fields. In abandoned croplands active restoration methods are needed to suppress weeds, control vegetation development and achieve a successful grassland recovery24,25. For rehabilitating and restoring grassland ecosystems in a cost-effective way at the envisaged global scale, fine-tuning of already known restoration methods is necessary.
Seed sowing is a widely used method that offers predictable results which can be implemented at various scales. Sowing propagules of matrix grasses and forbs proved to be a feasible tool for restoring diverse grassland habitats and to suppress weeds24,26. The composition of seed mixtures differs considerably regarding the aim, budget, machinery and timeframe available for restoration27. Sowing the propagules of a single grass species or low-diversity grass seed mixtures is effective when the aim is the fast restoration of the grass matrix28,29. The closed sward of grass species suppresses weeds, decreases soil openness and therefore the soil erosion30,31. Sowing only a few grasses is a cost-effective measure, as seeds of a limited number of species (usually 1–8 grass species) should be easily purchased or collected27. The costs of acquiring diverse grass-forb seed mixtures are considerably higher27,32 and it is often difficult to obtain the seeds of a large set of species in sufficient quality and quantity. That is one of the reasons why diverse seed mixture sowing is seldom applied in large-scale restoration projects. Grass sowing is widely used approach in large-scale restoration projects, but it should be considered that the developed closed sward not only suppresses weeds but also hampers the establishment of late-successional target grassland forbs24,29,33. Application of diverse seed mixtures by overseeding the grass matrix34,35 or sowing into gaps with bare soil24,36 is used mainly to increase species richness of species-poor restored grasslands.
The order of species arrival is a major driver of the species composition of the developed grasslands. Previous studies found that a priority effect can be detected even when species are sown with only three weeks difference37,38. Studies suggest that species arriving first tend to dominate the recovering vegetation39,40. If grasses are sown first, the establishment of other functional groups is generally negatively affected. In contrast, in the case of forb sowing, priority effect is missing or weaker compared to grass sowing33,39,41. The order and timing of sowing different components of the seed mixtures have a long-term effect on the vegetation, that can be detected also in later successional stages33.
Understanding the mechanism of vegetation recovery and post-restoration assembly rules can enhance restoration success. In our study, we established a mesocosm experiment, where we manipulated the initial floristic composition of the experimental plots by sowing seed mixtures with various diversity and timing (in total nine treatments with four replicates, Fig. 1, Table 1). Plots received only grass seeds (Festuca pseudovina) (G plots), only diverse forb seed mixture (D plots) or the combination of both (G + D plots). Forb mixtures were sown into two types of plots: plots sown with grass seeds (G + D plots) or fallows (D plots). Forbs were sown 0, 1, 2 or 3 years after grass seed sowing (G + D0, G + D1, G + D2, G + D3 plots) or after leaving the plot fallow for 0, 1, 2 or 3 years (D0, D1, D2, D3 plots). This study design enabled us to test the effect of grass matrix age and fallow age on the establishment success of forbs under field conditions and give recommendations for upcoming restoration projects. In addition, we conducted a germination test under greenhouse conditions to detect differences in germination rate of the sown forbs between sowing years and linked our results to weather parameters of the seed ripening period. We also studied the relation between the seed mass and establishment success of the sown forbs under field conditions. Our main goal was to identify the most feasible timing of seed sowing and the composition of seed mixtures to support restoration practitioners in creating diverse grassland communities, enabling the establishment of target forb species and the suppression of weeds, thus supporting the development of species-rich communities.

Design of the experimental site. Abbreviations indicate treatment types: G + D—plots sown with grass and diverse forb seed mixture; D—plots sown only with diverse forb seed mixture; G—plots sown with grass (F. pseudovina) seeds. Numbers indicate the age of the grass-matrix or fallow when diverse forb seed mixture (D) was added. Bottom-right square indicates the location of the permanent 1 m × 1 m subplots within the plots.
We hypothesised that (i) Forbs can establish better when sown on fallow than when sown into the grass matrix; (ii) Both sown grasses and forbs hamper the establishment of weeds; (iii) Time since cropland abandonment (fallow age) or grass sowing (grass matrix age) negatively affect the establishment success of sown forbs. As weather conditions during the seed ripening period affect seed quality42 we expected differences in seed germinability of sown forbs between sowing years.
Results
Vegetation development
Out of the 20 sown forb species 19 were able to establish in at least one of the experimental plots. In both the G + D and D plots the cover of sown forbs increased from the first year to the second (Table 2, Supplementary Table S1, Supplementary Fig. S1). Both cover and species richness were significantly lower in G + D plots than in D plots in both years (Table 3, Fig. 2). The cover and species richness of weeds decreased generally from the first to the second year, with the exception G + D3 plots, where the cover was higher in the second year. Both scores were the lowest in G + D plots and highest in D plots. The cover and species richness of weeds was similar in D and G plots in the first study year, but became significantly different in the second study year, when both variables were significantly lower in G plots. The cover of F. pseudovina did not change over the years and was higher in G + D plots than in G plots, but only in the first study year.

Vegetation development in different treatments in the first and second study years. Lower-case letters indicate significant differences between treatments (comparison with emmeans, p ≤ 0.05). Boxplot lines represent median values. G + D—plots sown with grass and diverse forb seed mixture, D—plots sown only with diverse forb seed mixture, G—plots sown with grass (F. pseudovina) seeds.
Effect of grass matrix- and fallow age
Age of grass-matrix and fallow (Table 2, Fig. 3) had mostly negative effects on the studied variables. In G + D plots cover and species richness of sown forbs decreased with increasing time lag between grass and D seed mixture sowing. Species richness of sown forbs decreased markedly if sown in grass-matrixes older than one year (G + D2 and G + D3). This trend was more pronounced in the second study year (Supplementary Table S2, Supplementary Fig. S2). Increasing age of grass-matrix also resulted in lower cover and species richness of weeds, especially in the first study year, while to the second study year weed cover increased significantly in G + D3 plots. The cover of F. pseudovina increased with age of grass matrix, the difference being significant only in the second study year. In D plots both sown species cover and richness were the highest in D1 plots, when sowing happened in one-year-old fallows (Supplementary Table S2, Supplementary Fig. S2). In older fallows both cover and species richness of sown forbs decreased significantly. The trend was detectable in both study years, although not always statistically significant. Weed cover and richness was lowest in the D1 plots. The effect of fallow age on weed cover was more pronounced in the second study year.

Grass-matrix age and fallow-age effect on species groups pooled for the two study years. Lower-case letters indicate significant differences (comparison with emmeans, p ≤ 0.05). Boxplot lines represent median values. G + D—plots sown with grass and diverse forb seed mixture, D—plots sown only with diverse forb seed mixture, G—plots sown with grass (F. pseudovina) seeds. Numbers indicate the age of the grass-matrix or fallows when diverse forb seed mixture (D) was added.
Four-year vegetation development in early sown plots
We analysed the four-year long datasets of G + D0, G + D1, D0 and D1 plots. Sown forbs cover had a non-significant tendency to increasing over the years (Table 4, Supplementary Table S3, Supplementary Fig. S3), but their richness was similar across the years in both G + D and D plots. In the first three study years, although not always significantly, the cover of sown forbs was higher in G + D0 plots than in G + D1 plots; it was lower only in the fourth study year (Supplementary Table S4, Supplementary Fig. S4). There was no difference in sown forbs richness between G + D treatment plots. In D plots we detected a higher cover and species richness of sown forbs in almost all years in D1 plots than in D0 plots.
Weed cover and species richness decreased significantly with time in both G + D and D plots. In G + D plots weed cover was significantly higher in G + D0 than in G + D1 plots in the first two years. The weed species richness was similar in all study years between the G + D treatments. Weed species cover and richness were in general similar in D plots; only in the second study year were both variables higher in D0 plots than D1 plots.
The cover of Festuca pseudovina did not change with time in G + D plots and only differed in the third study year, when it was higher in G + D1 than G + D0 treatments.
Relation between seed germinability, weather parameters and establishment success
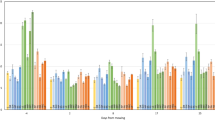
Germination rate of the sown forbs in greenhouse differed significantly between the years, being highest in 2015 (Supplementary Tables S5, S6). We did not find any effect of seed mass on germination rate of sown forbs in greenhouse (ß = − 1.296, SE = 1.524, z = − 0.8502, p = 0.3953). Temperature and precipitation conditions before seed collection had significant effect on germination rate (Table 5, Fig. 4). The germination rate of sown forbs was the highest when the temperature of the month preceding the seed collection was the lowest. A higher number of sown forbs were able to germinate after warmer weather conditions of the month preceding seed collection, although their success showed great variability. Germination rate was lower at higher average precipitation in the month preceding seed collection.

Germination rate of the sown forbs under greenhouse conditions in relation to 1 month average temperature and precipitation preceding seed collection.
Germination rate of the sown forbs under greenhouse conditions and their cover in field in the year following sowing in general were not related (Table 6).
Seed mass of the sown forbs had significant positive effect on their cover in field in the year following sowing; higher seed mass resulted in higher cover of the sown forbs (Table 7).
Discussion
Our results highlight the importance to introduce grasses and forbs at the same time to the target area to achieve the targeted high diversity of the restored grassland. Later introduction of forbs is usually less successful, but still costly, as the forbs cannot establish without the severe disturbance of entire removal of the existing vegetation.
In our study, both seed sowings, all the tested treatments applied alone or combined, resulted in the development of a high cover of grassland species within a short time period. However sown forb species were more successful in fallow plots, which supports our first hypothesis. In plots where F. pseudovina was sown prior to forbs both the cover and richness of sown forbs were lower, than in fallow plots, especially in the second study year. This indicates that competition among forb species is lower than competition between forbs and grasses. The sown grasses can hamper the development of other species groups in different stages of establishment, some of them already in the germination stage by their expressed allelopathic phytochemicals43,44,45. Furthermore, due to their high competitiveness that arises from their high allocation in belowground biomass (i.e., root system and clonal organs), efficient resource use ability and vegetative spreading ability, grasses can easily dominate grassland communities46,47. Besides, fast-growing grasses also increase competition for light48 and create microsite limitation24, that hampers the establishment of new-coming species.
In our system, F. pseudovina not only decreased the success of sown forbs but also that of weeds. F. pseudovina had especially negative effect on weeds in combination with forb species, supporting our second hypothesis, namely that both species groups hamper weed encroachment. As cover of weeds decreased over the years, they were replaced by sown forb species in plots where both sowing treatments (G + D) were applied; the grass cover was stable between the years, but that of sown forbs increased at the expense of weeds. This finding is in accordance with Werner et al.33 in general weeds are poor competitors compared to the sown species, both grasses and forbs, and are displaced by the latter species groups. The strong negative effect of the combination of the grass and diverse forb seed mixture is also supported by Humphries et al.22, who found that strong competition by other species is the most successful way to decrease weeds abundance.
As we hypothesized, time since grass sowing and cropland abandonment had a negative effect on the establishment success of sown forb species. Sown forb species were the most successful in the young grass-matrix or fallows, where competition by other species groups was still low and resources were not restricted. We also observed that changes in their cover mostly occurred from the first year to the second; the first year was important for forbs to establish and they increased their cover starting from the second year. The four-year long observations also support the importance of sowing forbs simultaneously with grasses or with just a one-year delay.
Grasses expressed a strong priority effect, so that the later introduced forb species were less successful. This is in accordance with other studies, where similarly to our study it was found, that grasses hamper the establishment of later-arriving species33,41 and prevent the development of species-rich grassland29. The previous studies found that even a short time difference in sowing grasses and forbs gives advantage for grasses, but in our study, we found that forbs can be still successful after one-year delay. A time lag longer than one year resulted in an irreversible advantage for F. pseudovina, as negative effects of grasses probably accumulated over the years (resource limitation, plant-soil feedback). To create and maintain a high diversity grassland, forbs, especially rare and weak competitors, should be sown as soon as possible, as even so there is high chance for some of them to perish over the years33,38. Besides, further interventions (harrowing, grazing) may also be needed to create suitable microhabitats for later-arriving species35,36.
Not only grass sowing, but also weed encroachment in fallow plots delayed sown forbs establishment. We observed that a two- or three-year-long fallow period resulted in a higher success of weeds, due to the lack of competitors and continuous seed-rain resulting the accumulation of a dense soil seed bank23,29,49,50, enabling them to dominate the unsown plots. We also detected a decreasing cover and richness of weeds after sowing the forb species. This is in accordance with other studies22,24,51, which found that restoration practices including propagule addition and follow-up management by mowing, can suppress weedy vegetation. It was however surprising that sowing in one-year-old fallows resulted in higher cover of sown forbs than sowing immediately after abandonment. This may be related to a higher overall seed germinability in that particular year, as seeds collected for sowing in one-year-old fallows (2015) also had the highest germination rates in the greenhouse. The difference was not observable in grass-matrix plots, suggesting that grass-competition had the strongest effect on the community development of those plots.
In general germination rate was not related to cover. One reason for this may be that under field conditions a high intraspecific competition was present between the individuals of species with high germination rate and negative frequency-dependence resulted in a decreased survival rate of individuals52. Another reason can be that in the field unfavourable post-germination conditions resulted in high mortality of seedlings, so the establishment of species with lower germination rate and gradual emergence may have been more successful42,53,54.
In our study the establishment success of sown forbs expressed by cover was positively related with seed mass; larger seed mass resulted in higher cover55. Larger-seeded species have an advantage at the early stages of their life cycle, as the resources for growing are provided for seedlings, so they can escape unfavourable conditions56. In natural or semi-natural communities, species with small seed would compensate with a higher seed number56,57,58,59 but our study design could not capture this trade-off, as we used equal seed numbers for all species. Seed dormancy can also contribute to the lower cover of small-seeded species, as small-seeded species more often have persistent seeds than species with larger seeds60.
Seed germinability was species-specific and was affected both by temperature and precipitation preceding seed collection. According to Fenner42, there is not an exact recipe based on which we can predict seed germinability. Weather parameters experienced by the seeds during their maturation can have contrasting effects on different species. High temperature in general decreases seed dormancy and increases seed germinability, but also results in thicker seed coat which in turn reduces seed germinability. In our study seed collection dates ranged from May to September, during which period temperature changed considerably. The highest germination rates were observed after seeds experienced lower temperatures during maturation; however, this was characteristic only for a few species, probably whose seed maturation ends in late spring or early summer. The majority of seeds germinated after being exposed to higher temperatures during their maturation, although higher temperature reduced overall germination rates. Lower germination rate does not necessarily equal with lower viability59; germination rate can be lower as a result of temperature-induced seed dormancy42. Similar to the effects of temperature, depending on the mechanism behind dormancy, drought can have similar contrasting and species-specific effects on seeds42. In our study the responses to precipitation preceding seed collection showed a high variation. Most of the seeds germinated better after drier conditions, presumably after experiencing summer drought, indicating the presence of autumn-germinating species and seeds. With increasing average precipitation, the germination rate slightly decreased but remained relatively high. Dormancy as well as other factors, like seed degradation and other unmeasured environmental parameters can be a cause of decreasing germination rate.
Based on our results, in the upcoming grassland restoration projects aiming to restore high-diversity grasslands we suggest the introduction of the target plant species simultaneously to the target area. This would reduce the necessity of severe disturbances (i.e., sward removal) later. However, because of the high variation in species germinability between years, reseeding may be still required. An overseeding with a one-year time lag should still result in high establishment success of the target forb species despite the presence of established grasses and still would minimize later costs.
Methods
Study site and experimental design
The study was conducted in an experimental site in Hajdúdorog (47.825718, 21.494500), East-Hungary. The climate of the region is continental, the mean annual temperature is 10.0 °C and the mean annual precipitation is 545 mm61. The experimental site is a private garden that is a former agricultural land. Agricultural management was stopped in 2014, prior to the start of the experiment. The last cultivated crop was potato. No remnants of natural- or semi-natural grasslands are present adjacent to the study site. We established 36 experimental plots of 4 m × 4 m in October 2014 (Fig. 1, Supplementary Fig. S5). Plots were arranged in a regular grid, with treatments assigned to plots in a regular pattern. One-meter-wide buffer zones were designated between plots. Plots were prepared by ploughing, rotary hoeing and raking, to remove vegetation remnants and roots and to create a fine-textured seed bed for seed sowing. Seed sowing took place yearly in October (21 Oct 2014, 22 Oct 2015, 14 Oct 2016, 11 Oct 2017). Plots received grass (G) seeds (Festuca pseudovina), diverse forb seed mixture (D mixture) or the combination of both (G + D). In all treatments with G, grasses were sown in 2014 (Year 0). Sowing D mixture had four levels: sowing in Year 0, 1, 2 or 3, i.e., sowing together with, or 1, 2 or 3 years after the sowing of grass seeds (Table 1). In October 2014 in total 24 plots were sown as follows: (i) in 16 plots only the seeds of F. pseudovina (G, G + D1, G + D2, G + D3 plots); (ii) in four plots F. pseudovina combined with D mixture (G + D0 plots) and (iii) in four plots only D seed mixture (D0 plots). The remaining 12 plots were left unsown in the first year to regenerate naturally and were managed the same way as the other plot types. The 12 empty (fallow) plots (D1, D2, D3) and 12 of the plots sown with F. pseudovina in 2014 (G + D1, G + D2, G + D3) were sown with D seed mixture in the following three years (2015, 2016 and 2017), each in four replicates per year (Table 1). In 2015, 2016 and 2017, prior to the D mixture sowing, G plots to be sown with D mixture were mown, while in unsown fallow plots mowing, rotary hoeing and raking was applied to remove the naturally established vegetation and to create the seed beds before D mixture sowing. In each case seeds were mixed with soil to prevent blowing by wind and to spread seeds more evenly in the plots. We managed the experimental plots by mowing twice in each year, in June and October, right after the vegetation surveys. Mowing was done by a handheld rotary mower, using a cutting height of 10 cm; the mown plant material was raked and removed from the plots immediately. Fertilizers and herbicides were not applied in any of the plots during the whole study period.
Diverse (D) forb seed mixture was composed of twenty forb species characteristic of the loess grasslands of the region (Supplementary Table S7). We collected the seeds from regional populations in the summer-autumn of each sowing year, at the peak of seed maturation period of each species. Seed collection followed the ENSCONET protocol62. Seeds were collected outside protected areas and none of the used species is under legal protection in Hungary. Therefore, no permissions were required for seed collection. Seeds were stored dry and cleaned manually. F. pseudovina was sown in a density of 20 kg/ha, which is the typical sowing density in dry grasslands in the region31. Forb species were represented in equal density in the D mixture with 1000 seeds each sown per the 16-m2-sized plots. The total amount of the grasses was 84% of the seeds sown in the G + D mixture, which corresponds to the proportion used in the restoration practice in the study region27.
Vegetation sampling
Between 2015 and 2019, percentage cover of all vascular plant species in plots where sowing previously occurred was recorded in June and October in four 1 m × 1 m-sized permanent subplots per plot (in total 144 plots). As species overlapped, total cover scores in a subplot can exceed a hundred-percent. Subplots were established inside each plot with 0.5-m-wide buffer zones from the plot edges and 1-m-wide buffers between the subplots. Survey of each plot started one year after sowing; i.e., G, D0 and G + D0 plots were sampled from 2015; D1 and G + D1 plots from 2016, D2 and G + D2 plots from 2017, and D3 and G + D3 plots from 2018, respectively.
To test whether there were differences between the germination rate of the collected seeds in the four years, a germination test of the sown forb species was conducted each year. This enabled us to compare the germination rate in the greenhouse and establishment capacity in the field for each sown forb species. Seed mass of each sown forb species was measured for three lots of 100 seeds with an analytical balance prior to seed sowing in each year in October. The measured seeds were sown in October (at the same time when the seeds were sown in the field experiment) in sterilized soil in plastic pots and kept in unheated greenhouse. Pots were watered every other day with tap water. The emerged seedlings were counted and removed weekly until no new seedlings were detected for three weeks.
Data processing and analysis
We classified species into three functional groups: (i) matrix grass (F. pseudovina), (ii) sown forb species (20 species, see Supplementary Table S7) and (iii) weeds. Weeds were classified based on Borhidi social behaviour types63; weeds (e.g. Capsella bursa-pastoris), ruderal competitors (e.g. Taraxacum officinale), adventive competitors (e.g. Ambrosia artemisiifolia) and introduced alien species (e.g. Anethum graveolens) were considered as weeds. Nomenclature follows Király64. We detected in total 40 other species (non-weed, non-sown) with an average cover of 16.13%. Given their low cover, we did not consider these species in the analyses of the functional groups. From cover data recorded in June and October of the same year, the higher one was used for each species. We retrieved average daily precipitation and temperature data from the database of the Hungarian Meteorological Service; from a 30-day-period before the seed collection dates of each forb species in each year.
In our analyses to study vegetation development between all treatment types we used the data from the first two years following the final sowing event of the given treatment. We included the cover data from G + D1 and G + D2 plots in the analysis of G plots, as before sowing D mixture into them, they served as G plots. To study the success of sown forbs in longer time period than two years, we used the four-year long data available from plots D0, G + D0, D1 and G + D1.
All data handling and statistical data analyses were carried out in R (v. 4.0.5, R Core Team)65. Throughout our analyses we mainly used generalised linear regression models (GLM) with the “glmmTMB” R-package66. Estimated marginal means (EMM) and factor-level comparisons were acquired with the R-package “emmeans”67. Initially plot ID was used as random factor to control for the non-independence of the replicates, but we excluded them from the final models, because of the very small random intercept variances, often leading to model convergence warnings or errors.
To test the effect of treatment and development time on species cover and richness, we used log-linked Gamma and quasi-Poisson generalized linear regression models (GLMs), respectively. In these models predictors were study year (as categorical factor, levels: 1, 2) and treatment type, also with control to their interaction. As response variables, we tested species cover and richness of sown forbs and weeds, and cover of Festuca pseudovina.
To test how species cover and richness changed across years in the separate treatments [i.e., with the age of grass-matrix (G + D) and fallow (D)], again log-linked Gamma and quasi-Poisson GLMs were fitted, respectively. In these models, the predictor was the age of grass-matrix/fallow (levels: 0–3) as categorical factor. Response variables were cover and richness of sown forbs and weeds, and cover of Festuca pseudovina.
Furthermore, we assessed the effect of development time on the difference between G + D0 and G + D1, and between D0 and D1 treatments (i.e., delayed sowing with one year), in species cover (log-linked Gamma GLM) and richness (quasi-Poisson GLM), by specifying study year (1–4, as categorical factor) and treatment type as predictors, and also controlling for their interaction. Hence we were able to assess for each study year whether or not there were significant differences between G + D0 and G + D1 plots, and between D0 and D1 plots.
We tested how average germination rate changed across survey years, by fitting a Beta GLMM, with germination rate as response, year (as categorical factor, levels: 2014–2017) as predictor, and species as random factor. We also tested the effect of seed mass on germination rate in a separate model, using only seed mass as predictor and species as random factor. In additional models, we assessed the effect of weather parameters [average temperature (°C) and rainfall (mm)] preceding seed collection on germination rate. We used separate models for temperature and rainfall to avoid multicollinearity. In the models we specified the orthogonal polynomials of the original weather variables to the power of 1 and 2, in order to be able to potentially detect non-linear effect of the utilized weather parameters. In all Beta models, prior to model fitting germination rate was transformed because Beta GLM(M)s generally cannot handle data containing zeros or ones:
where y and y′ are the original and transformed variables (germination proportion), respectively, and N is the number of values in the variable y (number of seeds, i.e. 100 for each species).
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Tölgyesi, C., Buisson, E., Helm, A., Temperton, V. M. & Török, P. Urgent need for updating a slogan of global climate actions from ‘tree planting’ to ‘restore native vegetation’. Restor. Ecol. 30, e13594. https://doi.org/10.1111/rec.13594 (2021).
Dengler, J., Janišová, M., Török, P. & Wellstein, C. Biodiversity of Palaearctic grasslands: A synthesis. Agric. Ecosyst. Environ. 182, 1–14 (2014).
Dass, P., Houlton, B. Z., Wang, Y. & Warlind, D. Grasslands may be more reliable carbon sinks than forests in California. Environ. Res. Lett. 13, 074027. https://doi.org/10.1088/1748-9326/aacb39 (2018).
Terrer, C. et al. A trade-off between plant and soil carbon storage under elevated CO2. Nature 591, 599–603 (2021).
Tilman, D., Reich, P. B. & Knops, J. M. H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 441, 629–632 (2006).
Stevens, C. J. Recent advances in understanding grasslands. F1000 Res. https://doi.org/10.12688/f1000research.15050.1 (2018).
Klaus, V. H. et al. Do biodiversity-ecosystem functioning experiments inform stakeholders how to simultaneously conserve biodiversity and increase ecosystem service provisioning in grasslands?. Biol. Conserv. 245, 108552. https://doi.org/10.1016/j.biocon.2020.108552 (2020).
Dudley, N. et al. Grasslands and savannahs in the UN decade on ecosystem restoration. Restor. Ecol. 28, 1313–1317 (2020).
Bardgett, R. D. et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2, 720–735 (2021).
Lengyel, S. et al. Restoration for variability: Emergence of the habitat diversity paradigm in terrestrial ecosystem restoration. Restor. Ecol. 28, 1087–1099 (2020).
Waldén, E. & Lindborg, R. Long term positive effect of grassland restoration on plant diversity: Success or not?. PLoS ONE 11, e0155836. https://doi.org/10.1371/journal.pone.0155836 (2016).
Lengyel, S. et al. Grassland restoration to conserve landscape-level biodiversity: A synthesis of early results from a large-scale project. Appl. Veg. Sci. 15, 264–276 (2012).
Sojneková, M. & Chytrý, M. From arable land to species-rich semi-natural grasslands: Succession in abandoned fields in a dry region of central Europe. Ecol. Eng. 77, 373–381 (2015).
Ellis, E. C. et al. Used planet: A global history. Proc. Natl. Acad. Sci. USA 110, 7978–7985 (2013).
Levers, C., Schneider, M., Prishchepov, A. V., Estel, S. & Kuemmerle, T. Spatial variation in determinants of agricultural land abandonment in Europe. Sci. Total Environ. 644, 95–111 (2018).
Winkler, K., Fuchs, R., Rounsevell, M. & Herold, M. Global land use changes are four times greater than previously estimated. Nat. Commun. 12, 2501. https://doi.org/10.1038/s41467-021-22702-2 (2021).
Perpiña Castillo, C. et al. Agricultural Land Abandonment in the EU within 2015–2030 (No: JRC113718) (Joint Research Centre (Seville site), 2018).
Müller, D., Leitão, P. J. & Sikor, T. Comparing the determinants of cropland abandonment in Albania and Romania using boosted regression trees. Agric. Syst. 117, 66–77 (2013).
Prishchepov, A. V., Müller, D., Dubinin, M., Baumann, M. & Radeloff, V. C. Determinants of agricultural land abandonment in post-Soviet European Russia. Land Use Policy 30, 873–884 (2013).
Prishchepov, A. V., Schierhorn, F. & Löw, F. Unraveling the diversity of trajectories and drivers of global agricultural land abandonment. Land 10, 97 (2021).
Bossuyt, B. & Honnay, O. Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J. Veg. Sci. 19, 875–884 (2008).
Humphries, T., Florentine, S., Dowling, K., Turville, C. & Sinclair, S. Weed management for landscape scale restoration of global temperate grasslands. Land Degrad. Dev. 32, 1090–1102 (2021).
Valkó, O. et al. Dynamics in vegetation and seed bank composition highlight the importance of post-restoration management in sown grasslands. Restor. Ecol. 29, e13192. https://doi.org/10.1111/rec.13192 (2021).
Valkó, O. et al. High-diversity sowing in establishment gaps: A promising new tool for enhancing grassland biodiversity. Tuexenia 36, 359–378 (2016).
Kövendi-Jakó, A. et al. Three years of vegetation development worth 30 years of secondary succession in urban-industrial grassland restoration. Appl. Veg. Sci. 22, 138–149 (2019).
Kiss, R. et al. Establishment gaps in species-poor grasslands: Artificial biodiversity hotspots to support the colonization of target species. Restor. Ecol. 29, e13135. https://doi.org/10.1111/rec.13135 (2021).
Török, P., Vida, E., Deák, B., Lengyel, S. & Tóthmérész, B. Grassland restoration on former croplands in Europe: An assessment of applicability of techniques and costs. Biodivers. Conserv. 20, 2311–2332 (2011).
Critchley, C. N. R., Fowbert, J. A., Sherwood, A. J. & Pywell, R. F. Vegetation development of sown grass margins in arable fields under a countrywide agri-environment scheme. Biol. Conserv. 132, 1–11 (2006).
Wagner, M., Walker, K. J. & Pywell, R. F. Seed bank dynamics in restored grassland following the sowing of high-and low-diversity seed mixtures. Restor. Ecol. 26, S189–S199 (2018).
Lepš, J. et al. Long-term effectiveness of sowing high and low diversity seed mixtures to enhance plant community development on ex-arable fields. Appl. Veg. Sci. 10, 97–110 (2007).
Török, P. et al. Restoring grassland biodiversity: Sowing low diversity seed mixtures can lead to rapid favourable changes. Biol. Conserv. 148, 806–812 (2010).
Schaub, S. et al. The costs of diversity: Higher prices for more diverse grassland seed mixtures. Environ. Res. Lett. 16, 094011. https://doi.org/10.1088/1748-9326/ac1a9c (2021).
Werner, C. M., Vaughn, K. J., Stuble, K. L., Wolf, K. & Young, T. P. Persistent asymmetrical priority effects in a California grassland restoration experiment. Ecol. Appl. 26, 1624–1632 (2016).
Williams, D. W., Jackson, L. L. & Smith, D. D. Effects of frequent mowing on survival and persistence of forbs seeded into a species-poor grassland. Restor. Ecol. 15, 24–33 (2007).
Klaus, V. H. et al. Enriching plant diversity in grasslands by large-scale experimental sward disturbance and seed addition along gradients of land-use intensity. J. Plant Ecol. 10, 581–591 (2017).
Kiss, R. et al. Zoochory on and off: A field experiment for trait-based analysis of establishment success of grassland species. J. Veg. Sci. 32, e13051. https://doi.org/10.1111/jvs.13051 (2021).
Weidlich, E. W. A. et al. Priority effects and ecological restoration. Restor. Ecol. 29, e13317. https://doi.org/10.1111/rec.13317 (2021).
Wilsey, B. Restoration in the face of changing climate: Importance of persistence, priority effects, and species diversity. Restor. Ecol. 29, e13132. https://doi.org/10.1111/rec.13132 (2021).
von Gillhaussen, P. et al. Priority effects of time of arrival of plant functional groups override sowing interval or density effects: A grassland experiment. PLoS ONE 9, e86906. https://doi.org/10.1371/journal.pone.0086906 (2014).
Eddy, K. C. & Van Auken, O. W. Priority effects allow Coreopsis tinctoria to avoid interspecific competition with a C4 grass. Am. Midl. Nat. 181, 104–114 (2019).
Delory, B. M., Weidlich, E. W., von Gillhaussen, P. & Temperton, V. M. When history matters: The overlooked role of priority effects in grassland overyielding. Funct. Ecol. 33, 2369–2380 (2019).
Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1, 75–84 (1991).
Ruprecht, E., Donath, T. W., Otte, A. & Eckstein, R. L. Chemical effects of a dominant grass on seed germination of four familial pairs of dry grassland species. Seed Sci. Res. 18, 239–248 (2008).
Partzsch, M., Faulhaber, M. & Meier, T. The effect of the dominant grass Festuca rupicola on the establishment of rare forbs in semi-dry grasslands. Folia Geobot. 53, 103–113 (2018).
Fenesi, A., Kelemen, K., Sándor, D. & Ruprecht, E. Influential neighbours: Seeds of dominant species affect the germination of common grassland species. J. Veg. Sci. 31, 1028–1038 (2020).
Garbowski, M. et al. Getting to the root of restoration: Considering root traits for improved restoration outcomes under drought and competition. Restor. Ecol. 28, 1384–1395 (2020).
Rehling, F., Sandner, T. M. & Matthies, D. Biomass partitioning in response to intraspecific competition depends on nutrients and species characteristics: A study of 43 plant species. J. Ecol. 109, 2219–2233 (2021).
Gross, K. L. & Mittelbach, G. G. Negative effects of fertilization on grassland species richness are stronger when tall clonal species are present. Folia Geobot. 52, 401–409 (2017).
Bakker, J. P. & Berendse, F. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends Ecol. Evol. 14, 63–68 (1999).
Kiss, R., Valkó, O., Tóthmérész, B. & Török, P. Seed bank research in Central-European grasslands: An overview. In Seed Banks: Types Roles and Research (ed. Murphy, J.) 1–34 (Nova Science Publishers, 2016).
Prach, K., Jongepierová, I. & Řehounková, K. Large-scale restoration of dry grasslands on ex-arable land using a regional seed mixture: Establishment of target species. Restor. Ecol. 21, 33–39 (2013).
Adler, P. B. et al. Competition and coexistence in plant communities: intraspecific competition is stronger than interspecific competition. Ecol. Lett. 21, 1319–1329 (2018).
Baskin, C. C. & Baskin, J. M. Seeds: Ecology, Biogeography, And Evolution of Dormancy and Germination (Academic Press, 1998).
Kövendi-Jakó, A. et al. Effect of seed storing duration and sowing year on the seedling establishment of grassland species in xeric environments. Restor. Ecol. 29, e13209. https://doi.org/10.1111/rec.13209 (2020).
Cevallos, D., Szitár, K., Halassy, M., Kövendi-Jakó, A. & Török, K. Larger seed mass predicts higher germination and emergence rates in sand grassland species with non-dormant seeds. Acta Bot. Hung. 64, 237–258 (2022).
Leishman, M. R., Wright, I. J., Moles, A. T. & Westoby, M. The evolutionary ecology of seed size. In Seeds: The Ecology of Regeneration in Plant Communities (ed. Fenner, M.) 31–57 (CAB International, 2000).
Westoby, M., Falster, D. S., Moles, A. T., Vesk, P. A. & Wright, I. J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Evol. Syst. 33, 125–215 (2002).
Moles, A. T. & Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 113, 91–105 (2006).
Scotton, M. Seed production in grassland species: Morpho-biological determinants in a species-rich semi-natural grassland. Grass Forage Sci. 73, 764–776 (2018).
Thompson, K., Bakker, J. P. & Bekker, R. M. The Soil Seed Banks of North West Europe: Methodology, Density and Longevity (Cambridge University Press, 1997).
Fick, S. E. & Hijmans, R. J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315 (2017).
ENSCONET (European Native Seed Conservation Network). ENSCONET Seed Collecting Manual for Wild Species. ENSCONET, Royal Botanic Gardens, Kew and Universidad Politécnica de Madrid (2009). http://www.kew.org/sites/default/files/ENSCONET_Collecting_protocol_English.pdf. Accessed 15 April 2014).
Borhidi, A. Social behaviour types, the naturalness and relative indicator values of the higher plants in the Hungarian flora. Acta Bot. Hung. 39, 97–181 (1995).
Király, G. (ed). Új magyar füvészkönyv. Magyarország hatásos növényei (New Hungarian Herbal. The Vascular Plants of Hungary. Identification Key) [in Hungarian]. (Aggtelek National Park Directorate, 2009).
R Core Team. R: A Language and Environment for Statistical Computing (4.0.5). Computer Software. R Foundation for Statistical Computing. https://www.R-project.org (2021).
Brooks, M. E. et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R. J. 9, 378–400 (2017).
Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means (Version 1.3.4) [R]. https://CRAN.R-project.org/package=emmeans (2019).
Acknowledgements
We are grateful to Szilvia Radócz, Sándor Borza, Ferenc Báthori and the Tóth family for their help in maintaining the experimental site. The help of Judit Sonkoly, Edina Tóth and Nóra Balogh in establishment of the experiment in 2014 is highly appreciated. The authors were supported by the Hungarian National Research, Development and Innovation Office (Grant Numbers: PD 137632—RK, KKP 144096—OV, FK 124404—OV, FK 135329—BD). AK was supported by the Bolyai János Scholarship of the Hungarian Academy of Science.
Author information
Authors and Affiliations
Contributions
O.V., B.D., R.K., conceived and designed the experiment; O.V., B.D., R.K., K.T., A.K., T.M., K.L., Á.T., L.G. collected the data; all authors take part in the maintenance of the experimental site, Z.R. and R.K. analysed the data; R.K. and O.V. wrote the manuscript; all authors checked and approved the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kiss, R., Deák, B., Tóth, K. et al. Co-seeding grasses and forbs supports restoration of species-rich grasslands and improves weed control in ex-arable land. Sci Rep 12, 21239 (2022). https://doi.org/10.1038/s41598-022-25837-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-25837-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
