
Abstract
Thanks to detailed studies of inclusions in Spanish and Kachin amber, it was also possible to study the morphology of insects belonging to the genus Gonomyia. A new material under investigation made it possible to establish two new nominative for science subgenera within the genus Gonomyia has been designated with unique set of characters of antenna, wing venation and genitalia. Two new species within two new subgenera have been described and documented by drawings and photographs, there are Gonomyia (Iberiana) penalveri subgen. et sp. nov. and Gonomyia (Cretagonomyia) burmitica subgen. et sp. nov. The new discovery is the first record of the genus Gonomyia (Diptera: Limoniidae) in Cretaceous Spanish amber and the second in Kachin amber. The new discovery adds to the knowledge of the crane fies’ diversity and evolution, especially its first stage in the Cretaceous.
Similar content being viewed by others


Introduction
The Cretaceous was the time represented either archaic or recent forms and in many respects an intermediate period between the Mesozoic and the Cenozoic eras. To understand the functioning of modern ecosystems, we must go back to the Cretaceous period and learn about the relationship between organisms living at that time. Such possibilities are offered by studies on Cretaceous Spanish and Kachin amber. The large variety and number of insect inclusions preserved in Kachin amber, especially the flies, is unique and allows us to recreate some of the details of ancient ecosystems.
The genus Gonomyia Meigen1 (Limoniidae, Chioneinae) is represented in recent fauna by over 600 species and subspecies belonging to nine subgenera2 as Gonomyia, Gonomyina Alexander3; Idiocerodes Savchenko4, Leiponeura Skuse5, Megalipophleps Alexander6, Neolipophleps Alexander7, Paralipophleps Alexander7, Prolipophleps Savchenko4 and Teuchogonomyia, Alexander8. Within them the most diversified are subgenera Leiponeura and Gonomyia, with 327 and 190 species and subspecies2. In fossil record four subgenera are represented as Gonomyia, Paralipophleps, Electrogonomyia Alexander9 and Azaria Kania, Krzemiński and Krzemińska10, the last two being known only from the fossil record. Nine species of Gonomyia known from fossil record remain unplaced in any subgenus. The oldest species of Gonomyia—Gonomyia (Azaria) libanensis Kania, Krzemiński and Krzemińska10 is known from Cretaceous Lebanese amber dated on Barremian11, the oldest Chioneinae are known since Lower Cretaceous10. Additionaly, one species of Gonomyia was mentioned from Upper Cretaceous Kachin amber12. Two other species of this genus were described from Eocene Baltic amber and classified into two different subgenera—Electrogonomyia and Gonomyia9. Four species of Gonomyia are known from Eocene/Oligocene of England, two from Oligocene of Germany, and two from Eocene and Miocene of Italy (imprints in sediments), two others from Dominican amber—Gonomyia (Paralipophleps) asymmetrica13 and one representative of Gonomyia unplaced to any species14. In total, 14 species of Gonomyia have been described from fossil record (Table 1).
Analysis of new fossil materials allowed to provide new information about Cretaceous (the oldest) stage of evolution of Gonomyia. There is no evidence that this group of insects existed on Earth earlier. It is the first evidence of the presence of this genus’ representatives—recorded as an inclusions in Cretaceous Spanish amber and the second from Kachin amber from Hukawng Valley, Myitkyina (Fig. 1C–F).

Maps with the position of Spanish and Kachin amber deposits localities. (A) Geographical setting of the studied section of Spanish amber deposits; (B) Geological setting with location of the studied section of Spanish amber deposits (after Barrón et al.21, modified); (C) Spanish and Kachin amber deposits localities; (D) Spanish and Kachin amber deposits localities in the Cretaceous; (E) Location of recent amber mining area in the Hukawng Valley, Myitkina Province, Myanmar; (A) Geological setting with location of the studied section of Kachin amber deposits. (E, F) Compiled from data provided by Kania et al.22.
Results
Systematic palaeontology
Order: Diptera Linnaeus23.
Infraorder: Tipulomorpha Rohdendorf24.
Family: Limoniidae Rondani25.
Subfamily: Chioneinae Rondani25.
Genus: Gonomyia Meigen1.
Type species: Limnobia tenella Meigen1, by monotypy. [Originally proposed in synonimy; available by established use prior to 1961.]
Subgenus: Iberiana subgen. nov.
Type species: Gonomyia (Iberiana) penalveri subgen. et sp. nov.
Diagnosis. Antenna 10-segmented, shorter than palpus; last palpomere very elongate, as long as the previous three; Sc with its tip beyond half length of Rs; very elongate and almost straight Rs; short R3, approximately 0.3 × the length of R4, basal section of R5 separate Rs at almost right angle; the position of crossvein m-cu behind the bifurcation of Mb into M1+2 and M3+4 wherein closed; very small, only 1.5 × as long as wide, almost rhomboidal d-cell; A2 short, straight.
Etymology. The specific name is derived from Iberia. Gender: feminine.
Description. By monotypy, the description of the subgenus is the same as for the species.
Remark. Due to the morphology of the wing venation and morphology of antenna, it was possible to designate a new monotypic subgenus based on the female characteristics. Diversification of the number of segments of antennae due to sexual dimorphism is not observed in modern species belonging to the genus, and such variability has not been indicated in the fossil material. The monotypic subgenus Azaria known from the Cretaceous period characterize by d-cell open by atrophy of the basal section of vein M3 and by the position of cross-vein m-cu far beyond the bifurcation of Mb into M1+2 and M3+4. In Iberiana subgen. nov. cross-vein m-cu occur far behind the fork of Mb like in subgenus Azaria, but d-cell in Iberiana subgen. nov. is closed, vein A2 is short and straight, while in subgenus Azaria this vein is elongate and waved. The number of antennal segments in Iberiana subgen. nov. is unique.
Gonomyia (Iberiana) penalveri subgen. et sp. nov

Gonomyia (Iberiana) penalveri subgen. et sp. nov. (Limoniidae), inclusions in Spanish amber; body, lateral view.

Gonomyia (Iberiana) penalveri subgen, et sp. nov. (Limoniidae), inclusions in Spanish amber: (A) head and thorax, lateral view; (B) antennae, lateral view; (C) palpi lateral view.

Gonomyia (Iberiana) penalveri subgen. et sp. nov. (Limoniidae), inclusions in Spanish amber: (A) wing; (B) enlarged view of part of wing; (C) base of wing; (D) ovipositor.

Gonomyia (Iberiana) penalveri subgen. et sp. nov. (Limoniidae), inclusions in Spanish amber: (A) antenna; (B) palpus; (C) wing; (D) ovipositor.
Diagnosis. As for the subgenus.
Etymology. The specific name is dedicated to eminent geologist and paleontologist Enrique Peñalver from the Museo Geominero, Madrid, Spain.
Material examined. Holotype No. MCNA 8818 (female), housed in the Museo de Ciencias Naturales de Álava, (Vitoria-Gasteiz, Álava, Spain).
Horizon and locality. Lower Cretaceous (Upper Albian); Peñacerrada I amber site (Peñacerrada I, Moraza), eastern area of the Basque-Cantabrian Basin, Burgos, northern Spain.
Description. Body (Fig. 2) 2.63 mm long, darkbrown, wings without color pattern, pterostigma absent.
Head (Figs. 2, 3A–C): antenna (Figs. 2, 3A,B, 5A) 0.64 mm long (1/0.08; 2/0.06; 3/0.10; 4/0.09; 5/0.05; 6/0.07; 7/0.05; 8/0.04; 9/0.04; 10/0.06); longer than head, shorter than palpus; scape and pedicel rather short, shorter than first flagellomere; scape cylindrical, narrow, pedicel widened distally, massive, only slightly longer than wider, flagellomeres elongate, oval, flagellomeres 1–5 approximately 3 × as long as wide, flagellomeres 6–8 at most 2 × as long as wide, flagellomeres became more slender to the apex of antenna; last flagellomere shortenest, without elongate setae at apex, first flagellomere with a few elongate setae, approximately as long as segment bearing them, each other flagellomeres with two elongate setae usually longer than segments bearing them; palpus (Figs. 2, 3A,C, 5B) elongate, 0.94 mm long (1/0.19; 2/0.12; 3/0.17; 4/0.46), palpomeres 1–3 elongate, narrow, first palpomere 5 × as long as wide, only slighlty longer than second and third, third palpomere slightly windened in the middle second and third palpomeres 3.5 × as long as wide, last palpomere very elongate and tiny, 2.5 × as long as third one; palpomeres 1–3 with a few elongate setae, only slightly shorter than segments bearing them, last palpomere with several not very elongate setae, approximately equal in length to width of this palpomere.
Thorax (Figs. 2, 3A): scutellum widened in the middle; wing (Figs. 2, 4A–C, 5C) 2.75 mm long, 0.86 mm wide; tip of R1 just before fork of R2+3+4; sc-r at approximately three of its length from the tip of Sc; Rs 0.65 mm long, longer than half the length of R5; R1 elongate, ending just beyond d-cell, in approximately 0.25 × the length of R2+3+4; R4 longer than R2+3+4, achieves about 0.6 × the length of Rs; d-cell 0.20 mm long, 1.5 × as long as wide, 0.25 × the length of M3; tip of M4 just beyond fork of R2+3+4 on R3 and R4; crossvein m-cu almost as long as d-cell; tip of A2 just before m-cu; tip of A1 just before fork of Rb; haltere (Figs. 2, 3A): stem narrow, elongate, slightly longer than knob.
Abdomen (Figs. 2, 4D): female terminalia – ovipositor (Figs. 4D, 5D) 0.72 mm long, rather short and wide, massive in comparison to the rest of the body, tenth tergite elongate; cercus elongate, tiny, pointed, hypogynal valvae massive, but acutely pointed, tip reaches to middle of cercus.
Subgenus: Cretagonomyia subgen. nov.
Type species: Gonomyia (Cretagonomyia) burmitica subgen. et sp. nov.
Diagnosis. Antenna 13-segmented, longer than palpus; scape and pedicel elongate, comparative length, approximately 2.5 × as long as wide, scape cylindrical, pedicel widened in the middle, first flagellomere elongate, narrow, approximately 4 × as long as wide; palpus shorter than antenna, last palpomere shorter than two penultimate; Sc with its tip just before fork of M3+4 on M3 and M4; elongate and almost straight Rs; R3 not very short, approximately 1.5 × the length of R2+3+4 and 0.75 × the length of R4; basal section of R5 separate Rs at an acute angle; vein M3 1.5 × as long as R2+3+4; morphology of gonostyles very simple, gonostyles not strongly sclerotized, outer gonostylus narrow, elongate, pointed, longer than inner gonostylus, inner gonostylus as narrow as outer, pointed; A2 short, arched.
Etymology. The specific name is derived from “creta” (Latin) = Cretaceous and “Gonomyia” from nominative genus. Gender: feminine.
Description. By monotypy, the description of the subgenus is the same as for the species.
Gonomyia (Cretagonomyia) burmitica subgen. et sp. nov

Gonomyia (Cretagonomyia) burmitica subgen. et sp. nov. (Limoniidae), inclusions in Kachin amber: (A) body, latero-ventral view; (B) antenna and palpi; (C) wing; (D) hypopygium.

Gonomyia (Cretagonomyia) burmitica subgen. et sp. nov. (Limoniidae), inclusions in Kachin amber: (A) antenna; (B) palpus; (C) hypopygium; (D) wing.
Diagnosis. As for subgenus.
Etymology. The specific name is derived from mineralogical name of the resin containing inclusions—burmite.
Material examined. Holotype No. BA02-050 (male), housed in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China.
Horizon and locality. Lowermost Cenomanian, Hukawng Valley, northern Myanmar. The mining is done at a hill named Noije Bum, near Tanai Village (26°21′33.41ʺN, 96°43′11.88ʺE).
Description. Body (Fig. 6A) 3.02 mm long, brown, wings without color pattern, pterostigma absent.
Head (Fig. 6A): antenna (Figs. 6A,B, 7A) 1.08 mm long (1/0.12; 2/0.10; 3/0.13; 4/0.05; 5/0.05; 6/0.05; 7/0.05; 8/0.05; 9/0.05; 10/0.05; 11/0.05; 12/0.05; 13/0.05); longer than head, longer than palpus; scape cylindrical, narrow, pedicel widened in the middle, massive, approximately 2.5 × longer than wider, flagellomeres elongate, cylindrical, approximately 4 × as long as wide, became more slender to the apex of antenna; last flagellomere as long as penultimate one, without elongate setae at apex, first flagellomere with a few elongate setae, approximately as long as or shorter than segment bearing them, each other flagellomeres excluding the last one with two elongate setae, one of these setae approximately twice as long as segments bearing them, the second one as long or shorter than segments bearing them; palpus (Figs. 6B, 7B) rather short, 0.32 mm long (1/0.10; 2/0.06; 3/0.06; 4/0.10), palpomeres narrow, first palpomere approximately 2.5 × as long as wide, only slightly longer than second and third, third palpomere slightly windened in the middle, second and third palpomeres 2 × as long as wide, last palpomere rather short, only slightly longer than penultimate one, 1.4 × as long as third one; palpomeres with a few not very elongate setae, shorter than segments bearing them.
Thorax (Fig. 6A): scutellum widened in the middle; wing (Figs. 6A,C, 7C) 3.72 mm long, 1.12 mm wide; sc-r at approximately three of its length from the tip of Sc; Rs not very elongate, 0.53 mm long, shorter than half the length of R5, R1 elongate, ending far beyond d-cell, in approximately 0.2 × the length of R3; R3 elongate, longer than half the length of R4; R4 approximately twice as long as R2+3+4, longer than Rs; vein M3 approximately 2.5 × the length of d-cell; d-cell 0.43 mm long, 2 × as long as wide; tip of M4 just before fork of R2+3+4 on R3 and R4; crossvein m-cu shorter than d-cell; tip of A2 just before m-cu and fork of Mb; tip of A1 far before fork of Rb.
Haltere (Fig. 6A,C): stem narrow, elongate, slightly longer than knob.
Abdomen (Fig. 6A,D, 7D): male terminalia—hypopygium 0.44 mm long, rather short and narrow; gonocoxite narrow, approximately 2.5 × as long as wide; gonostyles narrow, slightly sclerotized, outer gonostylus slightly longer than inner gonostylus, arrange more than 0.3 × the length of gonocoxite, pointed; inner gonostyle widened at base, pointed; aedeagus the average length, parameres short, strongly sclerotized; gonocoxite with not numerous setae, rather short setae.
Discussion
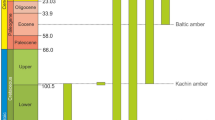
Though the craneflies of the genus Gonomyia are known from many places of different age, and they were preserved as an inclusions in Cretaceous Lebanese or Kachin amber, Eocene Baltic amber and Miocene Dominican amber9,10,12,13,14, or as an imprints are known from area of today’s Europe, from Miocene locality of Italy, Oligocene of Germany or Eocene/Oligocene of England15,16,17,18,19,20 the knowledge about fossil representatives of Gonomyia is still insufficient. In recent fauna the genus is diversified and differentiated, while from fossil record only 14 species were known so far, most of them unplaced to any subgenus, two described only as Gonomyia sp. (Table 1). From the Creatceous period, from ante-Barremian Lebanese amber is known the oldest representative of Gonomyia10, and one from Lowermost Cenomanian Kachin amber classified by Podenas and Poinar12 to subgenus Gonomyia. The discovery of new materials give us new information very important for understanding the earliest stage of evolution of the genus Gonomyia. Finding representatives of the genus first in Lebanese amber, and then in Spanish or Kachin amber may indicate that the genus was already widespread on Earth in the Cretaceous. Moreover, comparison of known and new materials show that the genus was probably diversified in this period. Known from the Lebanese amber species belongs to subgenus Azaria, a lineage that is not represented either in the modern fauna or in the fossil record of other periods in the history of the Earth (Fig. 8). Characteristic position of crossvein m-cu behind the bifurcation of Mb into M1+2 and M3+4 occur in Azaria. Regardless of whether or not it is plesiomorphic character, this characteristic still persists in the present in taxa of several subgenera (e.g. Gonomyia and Teuchogonomyia). Such features as long vein R2+3+4, crossvein r–r (R2) atrophied, relatively short vein R3 or narrow d-cell allowed to classify the newly described herein species to the genus Gonomyia. Moreover, the study new peculiar materials allowed to establish two new subgenera with differentiated morphology of wing venation and number of flagellomeres. The subgenus Iberiana subgen. nov. is characterized for example by ten number of antenna, or small and almost rhomboidal d-cell, while the gonostyles in subgenus Cretagonomyia are very simple, only slightly sclerotized without any processes. But, both of them characterize by the occurence of crossvein m-cu in the position shifted beyond fork of Mb, but in contrast to the subgenus Azaria d-cell in these two subgenera is closed. This location of m-cu indicate the ancestral position of these subgenera within the genus Gonomyia, same as for the subgenus Azaria10.

Chronostratigraphic distribution of the Gonomyia Meigen1 (Diptera: Limoniidae), fossil species.
One more species have been described from the Cretaceous period12. Based on inclusion of female body in Cretaceous Kachin amber Podenas and Poinar12 described one new species which classified to the subgenus Gonomyia and there is no doubt that these two different species represents two different subgenera. The differences between G. (I.) penalveri subgen. et sp. nov. are well visible not only in morphology of antenna or wing venation, but also in the structures of ovipositor. Ovipositor of species described by Podenas and Poinar12 is uniformly light brown with tenth tergite elongate, comparatively short cercus with upturned tip and hypogynial valve are long, acutely pointed, tip of this structure reaches to middle of cercus. In G. (I.) penalveri sp. nov. dark brown ovipositor is equipped with elongate tenth tergite, cercus is long and tin pointed and hypogynial valvae are long, acutely pointed, of its tip also reaches to middle of cercus, but ovipositor is rather short and massive in comparison to the rest of the body. Some significant differences are also visible in morphology of antenna, palpus and wing venation. In G. (I.) penalveri sp. nov. antenna is 10-segmented with two elongate setae on each flagellomere, usually longer than segment bearing them, while in G. (G.) sp. antenna is 16-segmented (what is characteristic for subgenus Gonomyia) with setae shorter than length of flagellomeres. In G. (I.) penalveri sp. nov. palpomeres are elongate, approximately 3 × as long as wide, while in G. (G.) sp. at most 2 × as long as wide. In G. (I.) penalveri sp. nov. three basal palpomeres are elongate, 3 × to 5 × (first palpomere) as long as wide, last palpomere is equal in length preceding palpomeres taken together, in G. (G.) sp. three basal palpomeres are short, approximately equal in length, their length only slightly exceeds their width, terminal palpomere is elongate, cylindrical, about as long as two preceding palpomeres taken together. In newly described species d-cell is rather romboidal, in G. (G.) sp. is almost rectangular, vein R2+3+4 in G. (I.) penalveri sp. nov. is elongate, arrange 2.5 × the length of R3 but is shorter than R4, R3 is short, and reach only 0.3 × the length of R4, in G. (G.) sp. vein R2+3+4 is rather short, corresponds to 1.5 × the length of R3, and half the length of R4. Tip of R1 is situated just before fork of R2+3+4 in G. (I.) penalveri sp. nov., in G. (G.) sp. just beyond fork of Rs. Basal section of R5 separate Rs almost at right angles in G. (I.) penalveri sp. nov., while in G. (G.) sp. is elongate and connected to Rs at an acute angle.
In recent fauna the genus Gonomyia is reach in species and widespread from cold to tropical zones (except for Antarctica). Although the genus is the most diversified in species in the Neotropic (202 species and subspecies), in this region is represented by four subgenera—Gonomyia, Leiponeura, Neolipophleps and Paralipophleps, while in Nearctic occur only 40 species and subspecies within six subgenera—Gonomyia, Idiocerodes, Leiponeura, Neolipophleps, Paralipophleps or Teuchogonomyia2.
In the ancient ages of the Earth’s history, the genus Gonomyia was probably also rich in species and widespread, as evidence by the presence of representatives in many different age fossil resins and in sediments from various places around the world, even if they are single species. The Cretaceous period in the history of the Earth is of great importance due to changes in fauna and flora, but also climate change. The resin from which the Spanish amber was produced in subtropical forests with seasonal wet-dry climate dominated by conifers and inhabited by ferns, gymnosperms and early angiosperms26. Kachin amber deposits as considered Grimaldi et al.27 were formed in tropical environment within an average temperature range of 32–55 °C, Lebanese amber formed in a tropical or subtropical, moderate to hot and very wet, dense forests28. In recent fauna representatives of Gonomyia are rather connected with flowing and standing waters. For example larvae of Gonomyia (Gonomyia) abscondita Lackschewitz29 are often found near the shores of flowing and standing waters, the species occur near streams and water margins on neutral acidic soil, frequently in wet woodland or scrub. Imago of this species usually occur in springs and headwater streams26. The larvae of the other species of the same subgenus as Gonomyia (Gonomyia) conoviensis Barnes30 are probably semi-aquatic, associated with sheltered wooded streams in upland areas, as well as other habitats in coastal locations, often occured with small streams in upland woodland situations. Larvae of Gonomyia (Gonomyia) hippocampi Stubbs and Geiger31 can be found in marginal situations along flowing and standing waters. Also other subgenera within Gonomyia are associated with the aquatic, wetland environment as the representatives of subgenus Leiponeura were found in the middle reaches of the river, their larvae occur also in saturated earth that ranges from coarse and sand to fine silit, but are much more frequent and aboundant in distinctly sandy situations. And although representatives of the subgenus Prolipophleps can be a specialist indicator of calcareous soil or bedrock, they are found in riverine sediments, mixed broadleaved forests along stream2.
The discovery of representatives of Gonomyia in these three Cretaceous resins testifies that this genus must have been diversified and widespread in the past. Moreover, these insects were adapted to tropical and subtropical palaeohabitats already in the Cretaceous period, as evidenced by the origination environment of Lebanese amber, Kachin and Spanish amber.
Material and methods
The study material—specimen No. MCNA 8818 comes from Cretaceous amber deposit of Peñacerrada I (Álava amber), is dated to Upper Albian (105 Ma21), are located in the northern slope of Sierra de Cantabria, in the southern limit of the Basque-Cantabrian Basin (northern Spain)26. Amber occurs in lutitic layers of deltaic origin with abundant coal. The specimen is housed in the Museo de Ciencias Naturales de Álava, (Vitoria-Gasteiz, Álava, Spain) (Fig. 1A–D). The specimens were embedded in epoxy resin (EPO-TEK 301) as described Corral et al.32 and Nascimbene and Silverstein33, which allowed physical protection and optimal study in ventral, lateral and dorsal views. The piece of amber has meassures of 8 × 6 × 1.5 mm and is embebbed in a resin piece of 10 × 8 × 1.5 mm.
The specimen No. BA02-050 (male) was found as an inclusion in the Cretaceous Kachin amber which deposits comes from the Hukawng Valley in the northern Myanmar, Myitkyina and Upper Chindwin districts (Myanmar)34,35,36,37 and are dated on 98.79 ± 0.62 Ma according to Shi et al.38 (data received based on research of zircons from the amberbearing bed), therefore, the amber is likely to be of early Cenomanian age39.
The specimen is housed in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China.
The specimens were examined with a Nikon (SMZ25) stereomicroscope Nikon SMZ 1500 equipped with a Nikon DS–Fi1 camera. The measurements were taken with NIS–Elements D 3.0 software. The length of the discal cell—measurements were given from its posterior edge to the point of connection of vein m-m with vein M3. The measurements were given only for undamaged structures. Drawings were completed by tracing the specimen and photographs, were made by Iwona Kania-Kłosok. Map (Fig. 1C) was built using the map Maps-For-Free (https://maps-for-free.com), map (Fig. 1D) was built using the plate tectonic reconstruction (https://www.odsn.de), both were modified with the software programs Corel Draw and Corel Photopaint X7. Abbreviations in accordance with: A1—first anal vein; A2—second anal vein; ant—antennae; Cu—cubital vein; d—discal cell; gx—gonocoxite; ing—inner gonostylus; M1–M4—first to fourth medial vein; m-cu—medio-cubital crossvein; Mb—medial-basal vein; oug—outer gonostylus; ovip—ovipositor; p—paramere; pl—palpi; ped—pedicel; r–r (R2)—second radial vein; R3–R5—third to fifth radial veins; Rb—radial-basal vein; Rs—radial sector vein; Sc—subcostal vein; sc-r—subcostal crossvein; scp—scape; I–IV—palpomeres first to fourth; Fig. 8: red circle—deposits of amber inclusion; yellow circle—deposits of imprint. The wing venation nomenclature follows that of McAlpine40.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Meigen, J. W. Systematische Beschreibung der Bekannten Europaischen Zweiflügeligen Insecten. (Erster Theil. F.W. Forstmann, 1818).
Oosterbroek, P. CCW: Catalogue of the Crane-flies of the World (Diptera, Tipuloidea: Pediciidae, Limoniidae, Cylindrotomidae, Tipulidae) (version May 2021). https://ccw.naturalis.nl/stats.php, http://nlbif.eti.uva.nl/ccw/index.php. Accessed 15 March 2022.
Alexander, C. P. Undescribed species of Tipulidae from the State of Sao Paulo, Brasil (Diptera). Livro Homenagem 1, 1–10 (1946).
Savchenko, E. N. New subgenera and species of the genus Gonomyia (Diptera, Limoniidae). Zool. Zhurnal 51(9), 1329–1341 (1972) (In Russian with English summary).
Skuse, F. A. A. Diptera of Australia, Part 7. The Tipulidae brevipalpi. Proc. Linn. Soc. New South Wales 2, 757–892 (1890).
Alexander, C. P. New or insufficiently known Australasian crane flies I (Tipulidae, Diptera). Stud. Entomol. 14, 267–318 (1971).
Alexander, C. P. Notes on the tropical American species of Tipulidae (Diptera). II. The primitive Eriopterini: Sigmatomera, Trentepohlia, Gnophomyia, Neognophomyia, Gonomyia and allies. Rev. Entomol. 18, 65–100 (1947).
Alexander, C. P. New or little-known Tipulidae from eastern Asia (Diptera). LXII. Philippine J. Sci. 96, 29–72 (1968).
Alexander, C. P. Crane-flies of the Baltic amber (Diptera). Bernstein-Forschungen 2, 1–135 (1931).
Kania, I., Krzemiński, W. & Krzemińska, E. The oldest representative of the genus Gonomyia (Diptera, Limoniidae) from Lebanese amber (Lower Cretaceous): New subgenus and species. Cretac. Res. 52, 516–521. https://doi.org/10.1016/j.cretres.2014.05.011 (2015).
Maksoud, S. & Azar, D. Lebanese amber: Latest updates. Palaeoentomology 003(2), 125–155. https://doi.org/10.1017/S0952836902001152 (2020).
Podenas, S. & Poinar, G. O. Jr. New crane flies (Diptera: Limoniidae) from Burmese amber. Proc. Entomol. Soc. Wash. 111(2), 470–492 (2009).
Podenas, S. & Poinar, G. O. New crane flies (Diptera: Limoniidae) from Dominican Amber. Proc. Entomol. Soc. Wash. 101, 595–610 (1999).
Krzemiński, W. The oldest Polyneura and their importance to the phylogeny of the group. Acta Zool. Cracov. 35(1), 45–52 (1992).
Krzemiński, W. & Gentilini, G. New information on Limoniidae from Monte Castellaro, Italy (Upper Miocene). Acta Zoologica Cracov. 35(1), 87–95 (1992).
von Heyden, L. Fossile Dipteren aus der Braunkohle von Rott im Siebengebirge. Palaeontographica 17, 237–266 (1870).
Statz, G. New Dipteren (Nematocera) aus dem Oberoligocän von Rott. III. Familie Limnobiidae (Stelzmücken). IV. Familie: Tipulidae (Schnaken). V. Familie: Culicidae (Stechmücken). Palaeontographica 95, 93–120 (1944).
Cockerell, T. D. A. Fossil Arthropods in the British Museum: VI Oligocene insects from Gurnet Bay Isle of Wight. Ann. Mag. Nat. Hist. Ser. 9(7), 453–480 (1921).
Cockerell, T. D. A. & Haines, F. H. Fossil Tipulidae from the Oligocene of the Isle of Wight (part). Entomologist 54, 81–84 (1921).
Krzemiński, W. & Krzemińska, E. Tipulomorpha (Diptera) of the Middle Eocene deposits from Pesciara di Bolca near Verona (Italy). Acta Zool. Cracov. 33(22), 495–499 (1990).
Barrón, E. et al. Palynology of Aptian and upper Albian (Lower Cretaceous) amber-bearing outcrops of the southern margin of the Basque-Cantabrian basin (northern Spain). Cretac. Res. 52, 292–312. https://doi.org/10.1016/j.cretres.2014.10.003 (2015).
Kania, I., Wang, B. & Szwedo, J. Dicranoptycha Osten Sacken, 1860 (Diptera, Limoniidae) from the earliest Cenomanian Burmese amber. Cretac. Res. 52, 522–530. https://doi.org/10.1016/j.cretres.2014.03.002 (2015).
Linnaeus, C. Systema nature per regna tria naturae, secundum classes, ordines, genera, species, cum caracteribus, differentiis, synonymi, locis. Tomus I. Editio decima, reformata. L. Salvii, (Holmiae, 1758).
Rohdendorf, B. B. Historical development of dipterous insects. Trudy Paleontol. Inst. AN SSSR 100, 1–311 (1964). [In Russian, English translation in Rohdendorf, 1974]
Rondani, C. Sopra una specie di insetto dittero. Memoria I. Nuovi Ann. Sci. Nat. Rend. 6, 284 (1841).
Peñalver, E. & Delclòs, X. Spanish Amber. In Biodiversity of fossils in amber from the major world deposits (ed. Penney, D.) 236–270 (Siri Scientific Press, 2010).
Grimaldi, D. A., Engel, M. S. & Nascimbene, P. C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Am. Mus. Novit. 3361, 1–71 (2002).
Azar, D., Gèze, R. & Acra, F. Lebanese amber. in Biodiversity of fossils in amber from the major world deposits. (ed. Peney, D.) 271–298 (Sri Scientific Press Manchester, 2010).
Lackschewitz, P. Neue Eriopterinen (Dipt. Nematoc.) aus dem Deutschen Entomologischen Institut. Nat. Maandblad 24, 9–14 (1935).
Barnes, H. F. Gonomyia bispinosa Barnes; change of name. Entomol. Monthly Mag. 60, 140 (1924).
Stubbs, A. E. & Geiger, W. Gonomyia (s. str.) hippocampi sp. n., a new european Eriopterinae (Diptera, Limoniidae). Mitteil. Schw. Entomol. Gesellsch. 66, 435–437 (1993).
Corral, J. C., LópezdelValle, R. & Alonso, J. E. ámbar cretácico de Álava (Cuenca Vasco-Cantábrica, norte de Espana). Su colecta y preparación. Estud. Museo Cienc. Nat. Álava 14, 7–21 (1999).
Nascimbene, P. & Silverstein, H. The preparation of fragile Cretaceous ambers for conservation and study of organismal inclusions. In Studies on Fossils in Amber, with Particular Reference to the Cretaceous of New Jersey (ed. Grimaldi, D.) 93–102 (Backhuys Publishers, 2000).
Zherikhin, V. V. & Ross, A. J. A review of the history, geology and age of Burmese amber (Burmite). Bull. Br. Mus. Nat. Hist. Geol. Ser. 56(1), 3–10 (2000).
Cruikshank, R. D. & Ko, K. Geology of an amber locality in the Hukawng Valley, Northern Myanmar. J. Asian Earth Sci. 21, 441–455. https://doi.org/10.1016/S1367-9120(02)00044-5 (2003).
Ross, A., Mellish, C., York, P. & Crighton, B. Chapter 12. Burmese Amber. In Biodiversity of Fossils in Amber from the Major World Deposits (ed. Penney, D.) 208–235 (Siri Scientific Press, 2010).
Yu, T. T. et al. An ammonite trapped in Burmese amber. PNAS 116, 11345–11350 (2019).
Shi, G. H. et al. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 37, 155–163. https://doi.org/10.1016/j.cretres.2012.03.005 (2012).
Smith, R. D. & Ross, A. Amberground pholadid bivalve borings and inclusions in Burmese amber: implications for proximity of resinproducing forests to brackish waters, and the age of the amber. Earth Environ. Sci. Trans. R. Soc. Edinb. 107, 239–247. https://doi.org/10.1017/S1755691017000287 (2017).
McAlpine, J. F. B. et al. Manual of nearctic diptera. Agric. Can. Res. Branch Monogr. 1, 27–674 (1981).
Acknowledgements
We would like to say thank you Editor Rongging Zhang and Reviewers Jorge Mederos López, and Anonymus Reviewer of the manuscript for valuable comments and remarks. We would like to acknowledge Rector University of Rzeszów for financial support. This project was realized within the project CRE: Global bioevent of massive resin production at the initial diversification of modern forest ecosystems, funded by the Spanish AEI/FEDER, UE Grant CGL2017-84419 and the project of the National Science Centre, Poland, Grant UMO-2016/23/B/NZ8/00936.
Funding
Open Access funding enabled and organized by University of Rzeszów.
Author information
Authors and Affiliations
Contributions
I.K.-K. conceived and designed the study, lead and performed the data analysis, interpretations and analysis, writing and corrections of the manuscript, graphics, measurements. A.A. obtaining material, map graphics, information about horizon and locality of Spanish amber, corrections of the manuscript. M.T. enviromental analysis, measurements, corrections of the manuscript. Q.Z. obtaining material, corrections of the manuscript. W.K. obtaining material, analysis and corrections of the manuscript. All authors reviewed manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kania-Kłosok, I., Arillo, A., Tuchowski, M. et al. Early stage of evolution of Gonomyia (Diptera: Limoniidae), new significant discoveries in the Cretaceous Iberian and Kachin amber. Sci Rep 12, 21118 (2022). https://doi.org/10.1038/s41598-022-25803-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-25803-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
