
Abstract
Based on known fossil evidence the extinct subfamily Architipulinae is considered to be the oldest evolutionary group of the Limoniidae, the largest family within the infraorder Tipulomorpha. The morphology of this subfamily, which includes 11 genera, has so far been based mainly on wing venation. New well-preserved representatives of the genus Cretolimonia Kalugina, 1986 were recovered from the Jurassic/Cretaceous boundary of Shevia and Daya, Transbaikalia, as well as from mid-Cretaceous amber from Kachin, Myanmar. This new material enriches our knowledge of the subfamily Architipulinae and of the genus Cretolimonia, and allows us to ascertain the detailed morphological structure of the female copulatory apparatus with spermathecae and the structure of the male hypopygium. The combination of detailed impression fossils with a specimen preserved three-dimensionally in resin has permitted study of the morphology of this Mesozoic fly genus almost to the level of modern genera. The paper includes descriptions of four new species of Cretolimonia: C. lukashevichae sp. nov., C. pseudojurassica sp. nov., C. dayana sp. nov. from sedimentary rocks, and C. mikolajczyki sp. nov. from Myanmar amber, supported with a key to all known species.
Similar content being viewed by others



Introduction
The Limoniidae (limoniid craneflies), with over 10,000 described extant species, is the largest family in the dipteran infraorder Tipulomorpha, and one of the largest among all Nematocera. It is currently divided into seven subfamilies, three of which are extinct (Architipulinae, Eotipulinae, Drinosinae), and four extant: Limnophilinae, Chioneinae, Dactylolabinae, and Limoniinae (Fig. 1). Based on the fossil data known to date Architipulinae is considered the oldest group of Limoniidae1,2,3,4,5,6,7, with the oldest representative Architipula youngi Krzemiński, 1992 dating from the Late Triassic of North America3. This species was used to calibrate the age of the Tipulomorpha clade in the phylogenetic tree of Diptera8. During the Jurassic, the family Limoniidae, including Architipulinae, underwent rapid radiation expressed in an abundant and diverse assemblage of genera and species3,7,9,10,11,12,13,14,15,16. In the Cretaceous, Architipulinae gradually became extinct, while other limoniid families had appeared in that time.

Geological ranges of subfamilies of the family Limoniidae.
The subfamily Architipulinae includes 11 fossil genera: Architipula Handlirsch, 19061, Protipula Handlirsch, 19061, Mesotipula Handlirsch, 192017, Paratipula Bode, 195318, Haplotipula Bode, 195318, Leptotipula Bode, 195318, Ozotipula Bode, 195318, Microtipula Bode, 195318, Cretolimonia Kalugina, 19862, Grimmenia Krzemiński and Zessin, 199019 and Metarchilimonia Blagoderov and Grimaldi, 20076. Up until now, knowledge of the morphology of the subfamily has been based mainly on wing impressions in sedimentary rocks. No representatives of Architipulinae were known from amber and no male genitalia had been found. Similarly, the second fossil subfamily, Eotipulinae, is known mainly from wing impressions in Jurassic and Cretaceous sedimentary rocks2,7, although a female has been described from Lower Cretaceous Spanish amber20. In contrast, in the third fossil subfamily, Drinosinae, the morphology of both sexes has been elucidated through material preserved in mid-Cretaceous Myanmar amber21.
The Architipulinae material described here shows an exceptional level of preservation, allowing us to augment the diagnosis of the genus Cretolimonia and enriching our knowledge of the whole subfamily Architipulinae. The hypopygium (a modified abdominal segment associated with the genitalia and having a clasping function in the males) is described for the first time on the basis of a perfectly preserved imprint in the rock, as well as the first representative of the subfamily from a fossil resin. Four new species of Cretolimonia are described, and a key to all known species is provided.
Results
Systematic palaeontology
-
Order Diptera Linnaeus, 1758
-
Infraorder Tipulomorpha Rodendorf, 196122
-
Family Limoniidae Speiser, 190923
-
Subfamily Architipulinae Handlirsch, 19061
-
Genus Cretolimonia Kalugina, 19862
Type species
Cretolimonia popovi Kalugina, 19862: figs. 87a, b: Gurvan-Ereny-Nuru (West Mongolia), Early Cretaceous. Genus description based on a wing fragment.
Species included
Table 1.
Amended diagnosis
The genus is distinguished from all Limoniidae by the characteristic wing venation pattern: vein Sc ends near Rs bifurcation, four radial veins present (R1, R3, R4 and R5), cross-vein r-r (R2) atrophied, and vein R3 very short, slightly curved; basal medial vein (Mb) well visible, long, all four medial veins present, d-cell closed, cross-vein m-cu located at the distal part of the d-cell base or sometimes in the middle (C. excelsa); ovipositor short, strongly curved dorsally; three small spermathecae present; male hypopygium with outer gonostylus strongly hooked; inner gonostylus lobed; aedeagus narrow and slightly curved, parameres large, triangular, very dilated at base.
Remarks
The genus Cretolimonia was included in the subfamily Architipulinae based on complex features which define this subfamily, i.e.: four radial and four medial veins reaching the wing margin, closed d-cell; cross vein m-cu located at the end of d-cell. The absence of cross-vein r-r (R2) distinguishes the genus Cretolimonia among other genera of the subfamily Architipulinae. So far, four species from the Middle Jurassic to mid-Cretaceous have been included in the genus (Table 1). For the purpose of the key, their wings are shown in Fig. 2A-D.

Wing venation of previously described species from genus Cretolimonia redrawn from original papers: (A) Cretolimonia popovi Kalugina, 1986; (B) Cretolimonia excelsa Gao et al., 2015; (C) Cretolimonia jurassica Lukashevich, 2009; (D) Cretolimonia pygmaea Lukashevich, 2009.
Key to species in genus Cretolimonia:
-
1. wing narrow, at least 3x longer than width .......... 2
-
- wing wide, about 2.5x longer than width .......... 7
-
2. Sc ends far beyond the Rs fork .......... 3
-
- Sc ends before or opposite the Rs fork .......... 4
-
3. M1 twice as long as upper edge of d-cell (Fig. 2A) .......... C. popovi Kalugina, 19862
-
- M1 only slightly longer than upper edge of d-cell (Fig. 2B) .......... C. excelsa Gao et al., 201523
-
4. Sc ends opposite Rs fork .......... C. dayana sp. nov.
-
- Sc ends in front of the Rs fork .......... 5
-
5. d-cell very small, no more than 1/10 of the wing length (Fig. 2C) .......... C. jurassica Lukashevich, 20097
-
- d-cell large, no more than 1/7 of the wing length .......... 6
-
6. R4 almost equal to the length of R2+3+4 .......... C. pseudojurassica sp. nov.
-
- R2+3+4 about 1/3 longer than R4 (Fig. 2D) .......... C. pygmea Lukashevich, 20097
-
7. petiola longer than M1; m-cu almost 2/3 along lower part of d-cell .......... C. lukashevichae sp. nov.
-
- petiola only about 1/3 as long as M1; m-cu just before the M3+4 bifurcation .......... C. mikolajczyki sp. nov.
-
Cretolimonia dayana Kopeć n. sp.

Cretolimonia dayana sp. nov.: (A) habitus of female, holotype No. 3063/1208; (B) female abdomen, No. 3063/399; (C) habitus of male, No. 3063/1079; (D) male hypopygium, No. 3063/1079.

Cretolimonia dayana sp. nov.: (A) wing venation, holotype No. 3063/1208; (B) antenna, holotype No. 3063/1208; (C) male hypopygium (reconstruction), No. 3063/1079; (D) female genitalia, No. 3063/399. Abbreviation: aed aedeagus, cer cercus, gx gonocoxite, hyp hypogenium, ing inner gonostylus, oug outer gonostylus, par parameres, ped pedicel, scp scapus, sp spermathecae.
Etymology
The name was established from the site where this species was discovered.
Material examined
Holotype No. 3063/1208, female (Fig. 3A,B). Additional material: 3063/399, female; 3063/1072, female; 3063/1079, male (Fig. 3C,D). Specimens come from Daya (Transbaikalia, Russia), Jurassic/Cretaceous boundary, housed in the Borissiak Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia (PIN).
Diagnosis
Wing narrow, about 3.3 times longer than its width; venation differs in the proportions of the individual veins from all species in this genus. Sc ends opposite the fork of Rs, d-cell long, narrow, expanded at distal part, no more than 1/9 of the wing length; m-cell lies at 2/3 length of the base of the d-cell; gonocoxites short, broad; outer gonostylus short, strongly chitinized, significantly hooked at end; inner gonostylus delicate, lobed; aedeagus long, narrow, slightly curved; parameres shorter than penis, broad at base.
Description
Wing length ca. 4.6 mm, width 1.4 mm (Fig. 4A); Head. Antennae with 16 segments, scapus tubular, half as long as its width, pedicel barrel-shaped, only slightly wider than scapus; flagellum with 14 flagellomeres, flagellomeres ovoid, gradually shortening, basal segment of flagellum expanded in lower part, last segment small and round, on all flagellomeres there are bristles shorter than width of segment on which they are located, on last segment 3 or 4 short bristles; clypeus 4-segmented, short, and the last segment almost equal in length to the penultimate one (Fig. 4B). Thorax. Wing narrow, about 3.3 times as long as its width; vein Sc ends opposite the Rs fork; cross-vein sc-r is about twice its length before the end of Sc; R1 ends opposite the R2+3+4 bifurcation at R3 and R4, and R2 (r-r) completely disappears; Rs about 1/4 longer than R2+3+4; R3 short, strongly sickle-shaped, slightly inclined, less than 1/3 length of R2+3+4, R4 about 1/4 shorter than Rs and equal in length to R2+3+4; four medial veins present, petiola ca. 1/4 longer than M1 and 1/3 longer than upper edge of d-cell; d-cell long, ca. 1/9 of wing length; cross-vein m-cu lies at 2/3 length of d-cell base; A2 vein not visible; legs long and delicate, tibial spurs absent. Abdomen. Male. Gonocoxites short, broad; outer gonostylus short, strongly chitinized, strongly hooked at the end; inner gonostylus delicate, lobed; penis long, narrow, slightly sigmoidally curved; parameres broad at base, shorter than aedeagus (3D,4C). Female. Ovipositor short, slightly curved dorsally; three small spermathecae (Fig. 3B, 4D).
Remarks
The male hypopygium is very well preserved in specimen No. 3063/1079 (Fig. 3D), being only slightly deformed by elongation of the left gonocoxite during fossilization. In previously described species of this genus the copulatory apparatus has not been preserved. The female ovipositor (Figs. 3B, 4D) is preserved almost perfectly in specimen No. 3063/399 with three small spermathecae present and well visible. The spermathecae are identical to those of most species in the subfamily Limnophilinae.
-
Cretolimonia lukashevichae Kopeć and Krzemiński n. sp.
-
Figure 5 
Holotypes of new Cretolimonia species: (A, C) Cretolimonia lukashevichae sp. nov. No. 3795/637, (A) habitus of holotype, (C) wing; (B, D) Cretolimonia pseudojurassica sp. nov. No. 3795/628, (B) habitus of holotype, (D) wing.
Figure 6 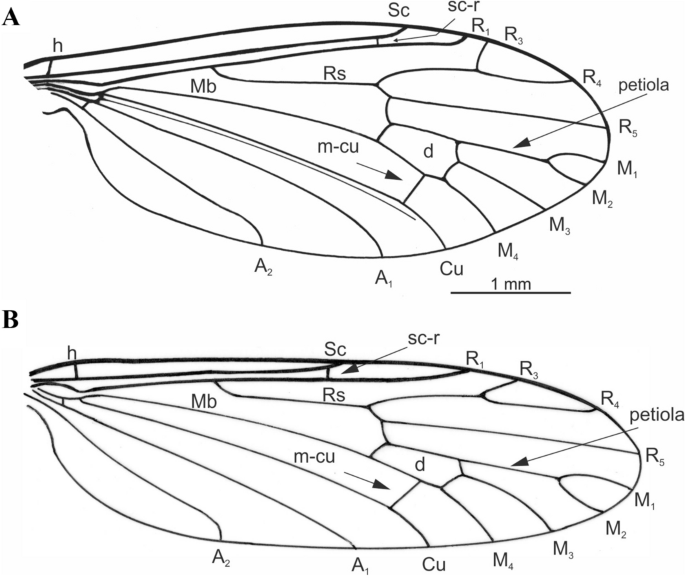
Holotypes of new Cretolimonia species: (A) Cretolimonia lukashevichae sp. nov., No. 3795/637; (B) Cretolimonia pseudojurassica sp. nov., No. 3795/623—wing venation.


Etymology
The species name is dedicated to Dr. Elena D. Lukashevich, who is involved in the study of fossil Diptera.
Material examined
Holotype No. 3795/637 sex unknown (Fig. 5A); paratype No. 3795/611 (sex unknown, hind wing preserved), Shevia (Transbaikalia, Russia), Jurassic/Cretaceous boundary; housed in the Borissiak Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia (PIN).
Diagnosis
Wing broad, 2.5 times as long as its width; Sc ends distinctly beyond fork of Rs; petiola more than half as long as M1 and 1/4 as long as upper margin of d-cell; m-cu lies nearly at 2/3 length of lower part of d-cell.
Description
Wing length 5.0 mm (right wing), width 2.0 mm, wing 2.5 times longer than its width (Fig. 5C, 6A); Sc ends distinctly beyond the Rs fork; cross-vein sc-r twice its length before the end of Sc; R1 ends before the R2+3+4 bifurcation at R3 and R4; R2 completely atrophied; Rs ca. 1/4 longer than vein R2+3+4; R3 short, almost vertical, constitutes ca. 1/3 length of R2+3+4; R4 ca. 1/4 shorter than R2+3+4; four medial veins present, petiola half longer than M1 and 1/4 longer than upper edge of d-cell; d-cell trapezoidal, relatively large, constituting 1/7 of wing length, cross-vein m-cu lies at 2/3 length of base of d-cell, far before bifurcation of M3+4 into M3 and M4; A2 vein almost straight.
Remarks
The holotype specimen shows two well-preserved wings and a fragment of the thorax. The left wing is longer than the right one, most probably deformed (elongated) during the fossilization process. Such a phenomenon has been observed in different groups of insects25,26. The right wing retained its normal structure.
Etymology
The species name emphasizes the similarity to another species in the genus, Cretolimonia jurassica Lukashevich, 20097.
Material examined
Holotype No. 3795/628 (Fig. 5B), sex unknown, only a single wing preserved. Additional material: 3795/633; 3795/638. Shevia (Transbaikalia, Russia), Jurassic/Cretaceous boundary. Housed in the Borissiak Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia (PIN).
Diagnosis
Wing narrow, nearly 3.3 times as long as its width; vein Sc ends distinctly before fork of Rs; R4 equal in length to R2+3+4; M1 slightly shorter than petiola and equal in length to upper margin of d-cell.
Description
Wing well-preserved with clearly visible veins, 5.1 mm long and 1.6 mm wide, almost 3.3 times as long as its width (Figs. 5D, 6B); Sc ends distinctly before bifurcation of Rs; cross-vein sc-r about its length before end of Sc; R1 ends before bifurcation of R2+3+4 into R3 and R4; R2 fully atrophied; Rs about 1/3 longer than R2+3+4; R3 short, almost parallel to R4, ca. 1/3 length of R2+3+4; R4 equal in length to R2+3+4; four medial veins present, petiola half longer than M1 and equal in length to upper edge of d-cell, d-cell trapezoidal, relatively large, constitutes ca. 1/6 length of wing; m-cu lies at 2/3 length of base of d-cell, far before bifurcation of M3+4 into M3 and M4; vein A2 slightly wavy.
Remarks
The wing venation resembles that of C. jurassica but significantly differs from that species in the structure of the d-cell and the proportions of the medial veins.
-
Cretolimonia mikolajczyki Kopeć, Krzemiński, Soszyńska-Maj sp. nov.
-
Figure 7 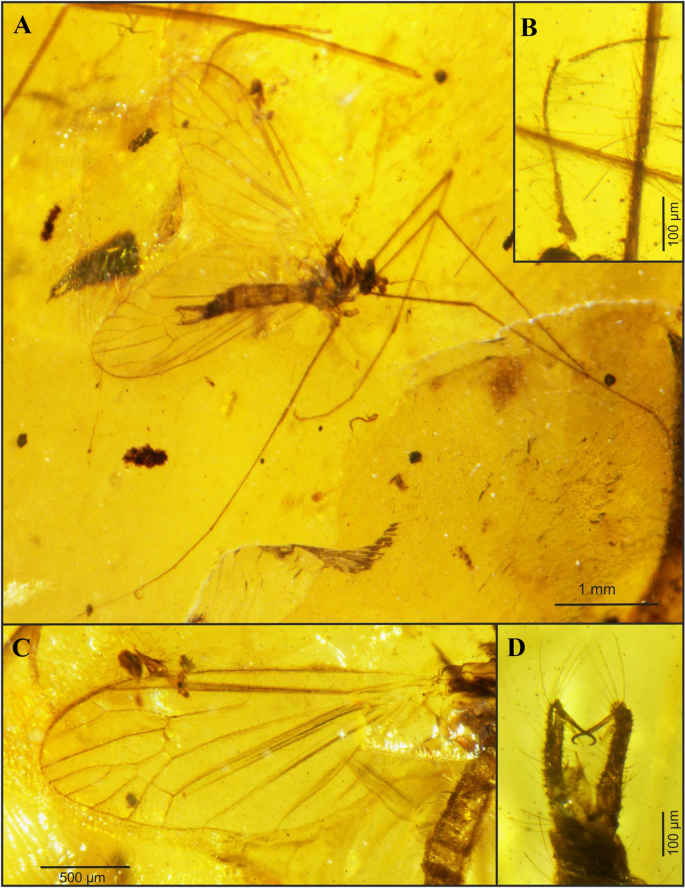
Cretolimonia mikolajczyki sp. nov., holotype No. MP/4082: (A) habitus of male; (B) antennae; (C) wing; (D) male hypopygium.
Figure 8 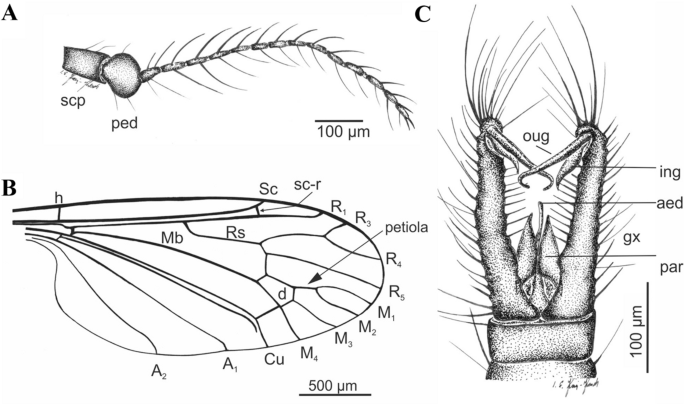
Cretolimonia mikolajczyki sp. nov. Holotype No. MP/4082: (A) antenna; (B) wing venation; (C) male hypopygium. Abbreviation: aed aedeagus, gx gonocoxite, ing inner gonostylus, oug outer gonostylus, par parameres, ped pedicel, scp scapus.


Etymology
We dedicate the species to the memory of a great and regrettably late colleague, the well-known Polish dipterologist Dr Waldemar Mikołajczyk.
Material examined
Holotype No. MP/4082, male (Fig. 7A); Kachin amber (Northern Myanmar); mid-Cretaceous, earliest Cenomanian. Housed in the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences (ISEA PAS).
Diagnosis
Wing broad, about 2.3 times as long as its width; wing venation differs in proportions of individual veins from all species in this genus; vein Sc ends opposite the fork of Rs, d-cell trapezoidal and distinctly expanded at the tip, constituting 1/10 of the wing length; petiola constitutes only about 1/3 of M1; cross-vein m-cu just before the fork M3+4 on M3 and M4; gonocoxites long, narrow, with numerous bristles, outer gonostylus long, narrow, strongly hooked at the tip; inner gonostylus delicate, lobed, elongated at the tip.
Description
Well-preserved specimen. Wing length 2.6 mm; width 1.2 mm; body length 2.8 mm. Head. Considerably wider than its length; antennae 16 segmented, scapus large, tubular half as long as its width, pedicel round, almost half as wide as scapus; flagellum 14 segmented, the first basal segment expanded in the lower part, the last small and rounded; bristles are present on all the flagellomeres, on the three basal segments bristles 2.5-3 times as long as the width of the segment, on the other flagellomeres bristles are shorter, 1.5-2 times as long as the width of the segment, on the last segment 3 or 4 short bristles (Fig. 7B, 8A). Palpi invisible. Thorax. Wing about 2.3 times as long as its width (Fig. 7C, 8B), additionally expanded in the anal field, vein Sc ends opposite bifurcation of Rs; cross-vein sc-r is about its length opposite the end of Sc; R1 ends opposite bifurcation of R2+3+4 into R3 and R4, R2 (r-r) fully atrophied; Rs about 1/4 longer than R2+3+4; R3 short, slanting, less than 1/3 length of R2+3+4; R4 only slightly shorter than R2+3+4; four medial veins present, and vein M1 2.5 times longer than petiola and twice as long as upper edge of d-cell; d-cell trapezoidal, small, constituting only 1/10 of wing length; cross-vein m-cu at end of d-cell, just before M3+4 bifurcates into M3 and M4; A2 vein strongly waved. Legs long, delicate, tibial spurs absent. Abdomen. Hypopygium (Fig. 7D, 8C): gonocoxites long, narrow, with numerous bristles, at end bearing a bunch of long bristles; outer gonostylus strongly chitinized, long, narrow, strongly hooked at end; inner gonostylus delicate, lobed, elongate at the end; penis long, narrow, slightly curved; parameres shorter than penis, broad at base, similar to parameres in Cretolimonia dayana sp. nov..
Remarks
Cretolimonia mikolajczyki sp. nov. is the first representative of the genus, and of the Architipulinae, found in fossil resin. Its excellent preservation permits detailed examination of morphological characters, especially the structure of the antennae and copulatory apparatus.
Discussion
Representatives of the infraorder Tipulomorpha were already present among the oldest fossil Diptera specimens from the early Middle Triassic (Anisian), ca. 245 Ma, from Arzviller (Vosges Mts., France)3,5,15,27,28. Archilimonia vogesiana Krzemiński and Krzemińska, 20035 and A. krzeminski Lukashevich and Ribeiro, 201916 belong to the fossil family Archilimoniidae. Archilimoniidae was included as a subfamily within the family Limoniidae, which we consider erroneous since the wing venation more closely resembles that of Pediciidae, especially in the radial field16. This being the case, the oldest known representative of the Limoniidae would be Architipula youngi Krzemiński, 19923 from the Late Triassic of North America (ca. 220 Ma)6 used for age calibration of Tipulomorpha8. It should be noted that the first 100 million years of dipteran, and hence tipulomorph, evolution is documented only by impression fossils, mainly wings28. The earliest examples of Limoniidae revealing three-dimensional structure have been recovered from Lower Cretaceous Lebanese amber, but such inclusions are very few in number29,30,31.
The specimens described here are of paramount importance for understanding the morphology of Architipulinae (Fig. 7A), and thus shed new light on the early evolution of flies of the suborder Tipulomorpha. The male genitalia of these flies are usually so severely deformed during fossilization that their precise structure and spatial arrangement cannot be determined. Although the preservation of Cretolimonia dayana sp. nov. is very good, being an impression fossil, it is only possible to obtain information in two dimensions. Therefore, finding a male of the same genus (C. mikolajczyki sp. nov.) in mid-Cretaceous amber presented a unique opportunity to fully reconstruct the genital anatomy, and thus to verify the structure of the hypopygium of C. dayana sp. nov. (Fig. 3B,4D). In sedimentary rocks, the genitalia of females are much better preserved, as they are usually strongly chitinized; in favourable circumstances even spermathecae are visible, and sometimes also genital plates26. However, so far, the number of spermathecae has not been known for the oldest known Limoniidae, the subfamilies Architipulinae or Eotipulinae. The well-preserved material from the Daya site allowed us to reconstruct the genitalia of the female C. dayana sp. nov. and to determine the number of spermathecae. The spermathecae are identical to those of most species in the family Limoniidae.
Most genera of Limoniidae have a full set of five radial veins, including the R2 vein, which takes the form of a cross-vein r-r. However, as far back as the Early Jurassic, a number of genera had appeared in which the cross-vein r-r (R2) had disappeared (i.e., atrophied) and one of these is the genus Cretolimonia. The disappearance of this cross-vein is observed also in some other Limoniidae, for example, in the subfamily Chioneinae, in the modern genera Gonomyia Meigen, 1818 and Rhabdomastix Skuse, 1890. There are, however, significant differences in other sectors of the wing in these genera; only three medial veins are always present, and the cross-vein m-cu is located in the anterior half of the d-cell, usually near the bifurcation of M3+4 into M3 and M4.
New material described here enabled us to characterize the morphology of the oldest (at least in the geological sense), extinct group of the Limoniidae; these features will be of great importance when introduced to the phylogenetical analysis of this large dipteran family.
Materials and methods
Geological context
This study was based on impression fossils and an amber inclusion (Fig. 9). The rock material comes from two sites, Shevia and Daya, in the Shelopuginsky District of the Chita region of Transbaikalia (Russia). Shevia (Dain Formation), dated Early Cretaceous, is located on the right bank of the Shevia River, 3 km below Shevia village, 2 km above the confluence of the Shevia and Shiviinsky Bumulei Rivers. The Daya site is located on the left bank of the Daya River above the Shevia Valley. It exposes sediments of the Glushkovo Formation, which are imprecisely dated but likely to lie close to the Jurassic/Cretaceous boundary32,33,34.

Map of localities yielding fossils of the genus Cretolimonia, yellow type—new localities.
The investigated amber inclusion derives from a former amber mine located near Danai (Tanai) Town (approximately at 26° 150″ N, 96° 340″ E) in the Hukawng Valley, state of Kachin in northern Myanmar. Radiometric U-Pb zircon dating of the volcaniclastic matrix of the amber produced an age of 98.79 ± 0.62 million years (earliest Cenomanian)29,35.
Specimen repository
All the specimens studied in the course of this work are deposited permanently in publicly owned collections in national museums. The Myanmar amber inclusion, the holotype of C. mikolajczyki sp. nov., No. MP/4082, is housed at the collection of the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences (ISEA PAS). It was acquired in 2016, before the armed conflict and the escalation of the ethnic strife in the area, in full compliance with the International Code of Zoological Nomenclature and Statement of the International Palaeoentomological Society36. The holotype of Cretolimonia dayana sp. nov. No. 3063/1208 and additional material 3063/399, 3063/1072; 3063/1079, holotype of C. lukashevichae sp. nov. No. 3795/637, paratype No. 3795/611 and holotypes of C. pseudojurassica sp. nov. No. 3795/623, along with additional material 3795/633; 3795/638 are housed in the collection of Borissiak Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia (PIN).
Methods
Specimens were studied using a Nikon SMZ25 stereomicroscope under reflected light, and photographs taken with a Nikon DSRi2 digital camera. The surfaces of rock specimens were wetted with 98% ethyl alcohol to improve the image contrast. Drawings were executed on the basis of the photographs with permanent referral to the specimens. The terminology of wing venation follows11,27. Geological ranges of subfamilies of the family Limoniidae are based on published data1,3,29,30,31,37.
References
Handlirsch, A. Die fossilen Insecten und die Phylogenie der rezent Formen (Engelmann, Leipzig, 1906–1908).
Kalugina, N. S. True flies. Muscida (=Diptera). Infraorders Tipulomorpha and Culicomorpha. Insects in the Early Cretaceous ecosystems of the West Mongolia. Trans. Joint Sov-Mong. Pal. Exped. 28, 112–125 (1986) (in Russian).
Krzemiński, W. Triassic and Lower Jurassic stage of Diptera evolution. Mitt. Schweiz. Entomol. Ges. 65, 39–59 (1992).
Shcherbakov, D. E., Lukashevich, E. D. & Blagoderov, V. A. Triassic Diptera and initial radiation of the order. Int. J. Dipterol. Res. 6, 75–115 (1995).
Krzemiński, W. & Krzemińska, E. Triassic Diptera: Descriptions, revisions and phylogenetic relations. Acta Zool. Cracov. 46(suppl. Fossil Insects), 153–184 (2003).
Blagoderov, V. A., Grimaldi, D. A. & Fraser, N. C. How time flies for flies: diverse Diptera from the Triassic of Virginia and early radiation of the order. Am. Mus. Novit. 3572, 1–39 (2007).
Lukashevich, E. D. Limoniidae (Diptera) in the Upper Jurassic of Shar Teg, Mongolia. Zoosymposia 3, 131–154 (2009).
Bertone, M. A., Courtney, G. W. & Wiegmann, B. M. Phylogenetics and temporal diversification of the earliest true flies (Insecta: Diptera) based on multiple nuclear genes. Syst. Entomol. 33, 668–687 (2008).
Krzemiński, W. & Kovalev, V. G. The taxonomic status of Architipula fragmentosa (Bode) and the family Eoasilidae (Diptera) from the Lower Jurassic. Syst. Entomol. 13, 55–56 (1988).
Krzemiński, W. The oldest Polyneura and their importance to the phylogeny of the group. Acta Zool. Crac. 35(1), 45–52 (1992).
Kopeć, K., Krzemiński, W., Skowron, K. & Coram, R. The genera Architipula Handlirsch, 1906 and Grimmenia Krzemiński and Zessin, 1990 (Diptera: Limoniidae) from the Lower Jurassic of England. Palaeontol. Electron. 20.1.15A, 1–7. https://doi.org/10.26879/637 (2017).
Kopeć, K., Ansorge, J., Soszyńska-Maj, A. & Krzemiński, W. Revision of the genus Mesotipula Handlirsch, 1920 (Diptera, Limoniidae, Architipulinae) from the Lower Jurassic of Northeast Germany. Hist. Biol. 32(4), 5000507. https://doi.org/10.1080/08912963.2018.1503257 (2018).
Kopeć, K., Soszyńska-Maj, A., Gehler, A., Ansorge, J. & Krzemiński, W. Mecoptera and Diptera from the early Toarcian (Early Jurassic) deposits of Wolfsburg—Groβe Kley (Lower Saxony, Germany). Earth Environ. Sci. Trans. R. Soc. Edinb. 107, 163–171 (2018).
Kopeć, K., Soszyńska-Maj, A., Lukashevich, E. & Krzemiński, W. Revision of the Mesozoic genus Mesotipula Handlirsch (Limoniidae, Diptera) from Asia extending its evolutionary history up to the Cretaceous. Cretac. Res. 114(1), 104504 (2020).
Kopeć, K. A new species of Mesotipula (Limoniidae, Diptera) from the Lower Jurassic of England. Ann. Zool. 68(1), 93–96 (2018).
Lukashevich, E. D. & Ribeiro, G. C. Mesozoic fossils and the phylogeny of Tipulomorpha (Insecta: Diptera). J. Syst. Palaeontol. 17(8), 635–652 (2019).
Handlirsch, A. Palaeontologie, 7. In Handbuch der Entomologie, III (ed. Schröde, S.) 117–304 (Jena: G. Fischer, 1920–1921).
Bode, A. Die Insektenfauna des ostniedersäschsischen Oberen Lias. Palaeontographica A 103(1–4), 1–375 (1953).
Krzemiński, W. & Zessin, W. The lower Jurassic Limoniidae from Grimmen (Diptera Nematocera). Dtsch. Entomol. Zschr. 37(1–3), 39–43 (1990).
Krzemiński, W. & Arillo, A. Alavia neli n. gen. and n. sp. the first Limoniidae (Diptera) from the Lower Cretaceous amber of Álava (Spain). Alavia neli n. Alavesia 1, 11–13 (2007).
Krzemiński, W., Kania, I., Krzemińska, E., Sevcik, J. & Soszyńśka-Maj, A. Fossils shed a new light on the diversity and disparity of the Family Limoniidae (Diptera, Nematocera). Insects 12(206), 206. https://doi.org/10.3390/insects12030206 (2021).
Rodendorf, B. B. The most ancient infraorders of Diptera from the Triassic of central Asia. Paleontol. J. 3, 90–100 (1961) (in Russian).
Speiser, P. 10. Diptera. 4. Orthorrhapha. In Wissenschaftliche Ergebnisse der Schwedischen Zoologischen Expedition nach dem Kilimandjaro, dem Meru und den umgebenden Massaisteppen Deutsch–Ostafrikas, 1905–1906. (ed. Sjöstedt, Y.) 31–65 (P. Palmquist, 1909).
Gao, J., Shih, C., Zhao, Y. & Ren, D. New species of Cretolimonia and Mesotipula (Diptera: Limoniidae) from the Middle Jurassic of Northeastern China. Acta Geol. Sin. Engl. 89, 1789–1796 (2015).
Ponomarenko, A. G. (ed.) The Cretaceous biocenotic crisis and insect evolution (Moscow Naturalist Society, 1988) (in Russian).
Krzemińska, E., Krzemiński, W. & Dahl, C. Monograph of Fossil Trichoceridae (Diptera) Over 180 Million Years of Evolution (ISEA, PAN, 2009).
Krzemiński, W., Krzemińska, E. & Papier, F. Grauvogelia arzvilleriana sp. n.—the oldest Diptera species (Lower/Middle Triassic of France). Acta Zool. Cracov. 37(2), 95–99 (1994).
Krzemiński, W. & Evenhuis, N. Review of Diptera palaeontological records. In Contributions to Manual of Palaearctic Diptera Vol. 1 (eds Papp, L. & Darvas, B.) 535–564 (Science Herald, 2000).
Kania, I., Krzemiński, W. & Krzemińska, E. The oldest representative of the genus Gonomyia (Diptera, Limoniidae) from Lebanese amber (Lower Cretaceous): New subgenus and species. Cretac. Res. 52, 516–521 (2015).
Kania, I., Krzemiński, W. & Azar, D. The oldest representative of Helius Lepeletier & Serville 1828 (Diptera: Limoniidae) from Lebanese amber (Early Cretaceous). Insect Syst. Evol. 44(2), 231–238 (2013).
Krzemiński, W., Kania, I. & Azar, D. The Early Cretaceous evidence of rapid evolution of the genus Helius Lepeletier and Serville, 1828 (Limoniidae, Diptera). Cretac. Res. 48, 96–101 (2014).
Rasnitsyn, A. P. Late Mesozoic Insects of Eastern Transbaikalia (Moscow Naturalist Society, 1990) (in Russian).
Ponomarenko, A. G. Mesozoic Insects and Ostracods of Asia (Moscow Naturalist Society, 1993) (in Russian).
Rasnitsyn, A. P. & Zherikhin, V. V. Appendix: Alphabetical list of selected insect fossil sites: Impression fossils. In History of Insects (eds Rasnitsyn, A. P. & Quicke, D. L. J.) 437–444 (Kluwer Academic Publishers, 2002).
Shi, G. et al. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 37, 155–163 (2012).
Szwedo, J., Wang, B., Soszyńska-Maj, A., Azar, D. & Ross, A. International Palaeoentomological Society Statement. Palaeoentomology 3, 221–222 (2020).
Podenas, S. & Poinar, G. O. New crane flies (Diptera: Limoniidae) from Burmese amber. Proc. Entomol. Soc. Wash. 111, 470–492 (2009).
Acknowledgements
The authors would like to show their gratitude to the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences (ISEA PAS) and Borissiak Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia (PIN) for providing the material for study.
Funding
The research was fully supported by a grant from the Polish National Science Center No. 2016/23/B/NZ8/00936.
Author information
Authors and Affiliations
Contributions
K.K. took the lead in writing the manuscript. K.K., W.K. were responsible for taxonomic decisions. K.K. was responsible for material preparation and photography. I.K.K. and A.S.M. performed graphic illustration. K.K., W.K., A.S.M. contributed to the interpretation of the results, correction of manuscript after reviews and general discussion. W.K. was the leader who supervised the work and financially supported the project. R.A.C. revised the text. A.S.M. was the corresponding author and coordinated the correction of the manuscript. All authors provided critical feedback and helped shape the research and manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kopeć, K., Soszyńska-Maj, A., Kania-Kłosok, I. et al. Morphology of the oldest fossil subfamily of Limoniidae (Diptera, Architipulinae) in the light of exceptionally preserved Mesozoic material. Sci Rep 11, 24137 (2021). https://doi.org/10.1038/s41598-021-03350-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-03350-4
This article is cited by
-
Cretaceous Antodicranomyia (Diptera: Limoniidae) and their paleohabitat
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
