
Abstract
Despite being one of the most important crops in the recent prehistory of Eurasia, the arrival and exploitation of millets in the westernmost part of Europe are still largely underexplored. Here and for the first time, we report multipronged biomolecular evidence of millet consumption along the Atlantic façade of northern Iberia through a combination of radiocarbon dating, stable isotopes, and dental calculus analyses on the human individuals found in the burial site of El Espinoso cave (Asturias, Spain). The high-resolution chronological framework established for individuals placed the burials between 1235 and 1099 cal. BC. The discovery of high δ13C values on their bone collagen and the identification of polyhedral starch grains within their dental plaque underline the relevance of C4 plants in their diet and highlights the timing of the systematic consumption of millets in the Late Bronze Age. Our data support previous regional archaeobotanical evidence and establish a more precise chronology of the dispersal of millets into northern Iberia during the Bronze Age, becoming an essential crop until the arrival of maize from America after AD 1492. This study emphasizes the importance of multidisciplinary methods to ascertain the origin and development of agricultural practices during recent prehistory.
Similar content being viewed by others

Introduction
Millets are small-seed spring/summer domestic grasses that played a key role in human diet and culture across Europe by the first-millennium cal. BC1. Two millet species have been traditionally cultivated in Europe during recent prehistory: Setaria italica (foxtail millet) and Panicum miliaceum (broomcorn millet)2. These plants have a short lifecycle3, allowing for annual crop rotation with other cereals, like wheat or barley4. Millets are crops with great ecological adaptability to different altitudes, soils, and climates5 and a rich nutritional value6. They are used to feed both people and animals7, and as ingredient in diverse alcohol fermentation techniques8. Unfortunately, despite their relevance among farming societies during the Iron Age, little is known about when and how millets were introduced into western Europe, specifically in northern Atlantic Iberia9.
The earliest evidence for domesticated broomcorn and foxtail millets comes from north-eastern China and eastern Inner Mongolia, dating to around 6000–5500 cal. BC10,11,12, although broomcorn millet has slightly earlier chronologies than foxtail millet in these areas. From East and Central Asia, the spread of millets would have occurred in the late third millennium cal. BC across the Inner Asian Mountain Corridor13 and the Eurasian Steppes14. In West Asia and East Europe, the spread of millet cultivation took place during the second millennium cal. BC, producing the first episode of food globalization which laid the foundations of an interconnected Eurasian continent15. In these areas, mobile pastoralists might have constituted key agents in the diffusion of these crops16 based on the coexistence of livestock and plant cultivation practices across Europe17. Direct radiocarbon dates obtained from several broomcorn millet grains, recovered at different Central and Eastern European sites, confirm that millet cultivation arrived in these regions during the second half of the second millennium cal. BC18,19,20, a considerably younger chronology than what was previously thought21. Instead, for western regions such as Britain, the latest direct radiocarbon dates point to a slightly later arrival around the first half of the first millennium cal. BC22.
Millets have been traditionally seen as minor crops, with a secondary role in past human economies, used as food for people of low social status and/or as fodder for animals23. Their small-seed morphology made them hard to find in old excavations, but the implementation of intensive collection programs and the systematic use of flotation systems during recent decades24 have favored the recovery of millet seeds, significantly enriching the archaeobotanical record worldwide. Ethnographic and experimental studies have also improved knowledge about millet cultivation, including harvesting, threshing, sieving, milling, storage, and cooking activities25,26. Furthermore, comparisons of nutritional components of millets show a similar, and even higher, dietary intake in terms of proteins, minerals, and vitamins than other cereals crops27. Recently, ancient starches and phytoliths retrieved in dental calculus and ground stone tool surfaces have provided new direct and indirect evidence for the prehistoric consumption of millets, given their potential to be differentiated at the genus or species level28,29,30 and between wild or domestic species31,32. Simultaneously, isotopic evidence for C4 plant consumption has been reported in different European regions, questioning the relevance of millets in their diets during the Bronze Age33. Thus, the current evidence about millet consumption is continuously growing, and millets might have been more relevant than previously thought in the recent prehistory.
In northern Iberia, the Cantabrian region has yielded an extraordinary Bronze Age funerary record where caves were selected as burial locations by early farmers34. Despite the richness of these assemblages, biomolecular approaches have rarely been applied to explore evidence related to the origin and development of its regional agriculture, such as the timing of the arrival of millets and how the human populations exploited them. Until the 1990s, millet cultivation was considered to be an Iron Age innovation associated with the hillfort culture35. The currently available carpological data support the presence of broomcorn and foxtail millets in this area, at least, since the Late Bronze Age, with some limited evidence from the Middle Bronze Age9. However, no direct dates on those seeds confirm their exact chronology. Similarly, stable isotope analysis of bone collagen from Bronze Age individuals failed to detect isotopic signals related to C4 plant consumption. Moreover, dental calculus analysis has never been applied to this spatio-temporal range that would allow identifying millet species' starch grains.
With the aim to identify the millet consumption in the Atlantic coast of northern Iberia and to ascertain when and how these crops were systematically exploited during the Bronze Age of this region, this paper combines direct radiocarbon dating on human remains, stable isotope analysis (δ13C, δ15N and δ34S) on human and animal bone collagen, and the study of starch grains entrapped in the dental calculus of humans found at El Espinoso burial cave.
El Espinoso cave
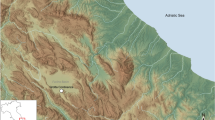
El Espinoso is located on the central coast of the Cantabrian Region (northern Spain), in the easternmost sector of Asturias province (Fig. 1), which is characterized by an Atlantic climate. The cave opens out from a 20-m-high limestone cliff, which dominates a closed valley, at only 200 m from the present Atlantic shoreline. The cave entrance is oriented towards SW, and to access it is necessary to climb a four-meter-high part of the cliff. At the end of the cavity, a collective burial was discovered, where the human bones were scattered around the cave floor, highly fragmented and with neither anatomical connections nor associated grave goods. Data from three humans recovered from the regional sites of La Llana, Los Cinchos and La Fragua caves, belonging to the Early and Middle Bronze Age, were used as a comparison with those found at El Espinoso (See Supplementary Texts 1 to 4 for more information about these sites). Recently, provisional genetic results from El Espinoso’s individuals have shown a Steppe ancestry of almost 30%36, constituting one of the Bronze Age sites with the greatest proportion of this ancestry in Iberia. Genetic evidence demonstrates that human migrations introduced the Steppe ancestry in Iberia during the Chalcolithic/Bronze Age transition37. This trend was remarkably stronger in northern Iberia, giving clues by the place where these populations arrived. The incoming ancestry is even more significant in El Espinoso, suggesting a possible second flow of people in the Late Bronze Age. These population dynamics may have caused social and economic changes in northern Iberia, including the potential arrival of new crops.

Location of El Espinoso cave in the Atlantic façade of southwestern Europe (A) and the Cantabrian Region (northern Iberia) marked in black (B). (C) Aerial photography of the site location. Maps of (A) and (B) were generated using ArcGis Pro software (v2.4.0 https://arcgis.com/). The original base maps were extracted from the SRTM Data web site (https://srtm.csi.cgiar.org/srtmdata/). Aerial photography (C) was generated using Google Earth Pro software (v7.3.4.8642 https://www.google.com/intl/es/earth/).
Results
Radiocarbon dating and Bayesian modelling
Fifteen individuals of the total of 20 were directly dated. All the dates fall in the last third of the second millennium cal. BC, corresponding to the Late Bronze Age (Supplementary Table 1)38. The El Espinoso sequence showed a convergence greater than 99%, and the model and overall agreement indices were 107.9% and 100.2%, respectively. Only OxA-38660 date had a poor agreement of 52.9%. A model of the burial phase at the site was built with start and end boundaries (Fig. 2). This model was replicated up to three times to prove its reliability, and no more outliers were found. The model provides a start date between 1235 and 1129 cal. BC and end date between 1214 and 1099 cal. BC (at 95.4% probability). Thus, the formation of the assemblage spans between 1235 and 1099 cal. BC and the maximum duration of the burial phase is 115 years. KDE plot shows three events along the funerary sequence of the site with greater importance of the central peak when the summed probability distribution suggests that most individuals were buried. Thus, the El Espinoso cave was used for funerary practices for a relatively short time (a few generations, at most). These data constitute a well-defined, almost synchronic event when the deceased individuals of a specific population were repeatedly introduced into the cave.

(A) Radiocarbon dates from El Espinoso cave modelled in OxCal v4.4.2 against IntCal20 curve. (B) KDE plot of the likelihood distributions of El Espinoso burial phase.
Stable isotope analysis
The stable isotope values for the human and animal remains are reported in Supplementary Table 2 and plotted in Fig. 3. Collagen extraction was successfully undertaken in all the samples with %Col > 5. Their %C, %N, %S, C:N, C:S, and N:S elemental ratio values indicate good collagen quality39,40,41,42.

Scatterplot of human and animal bone collagen δ13C, δ15N and δ34S values of El Espinoso cave and δ13C, δ15N and δ34S values of human bone collagen from La Llana, Los Cinchos, and La Fragua caves.
From El Espinoso cave, the humans analyzed (n = 14) had δ13C values ranging between − 17.8 and − 14‰ (m = − 15.6‰), δ15N values between 9.1 and 10.4‰ (m = 9.5‰) and δ34S values between 13.5 and 16.9‰ (m = 14.9‰).The two faunal specimens available were analyzed as a local baseline from the site: a cow specimen reported a δ13C signature of − 22.1‰, a δ15N value of 4.1‰ and a δ34S signature of 16.2‰, and a sheep had a δ13C value of − 20.6‰, a δ15N signature of 3.7‰ and a δ34S value of 13.5‰. Humans from La Llana, Los Cinchos, and La Fragua sites reported δ13C values of − 20.9‰, − 21.1‰ and − 20.4‰ and δ15N values of 8.6‰, 9.2‰ and 9.2‰ and δ34S values of 14.9‰, 14.5‰ and 14.8‰, respectively.
Dental calculus analysis
Seventy-four starch grains were found in the dental calculus of 16 individuals (Supplementary Table 3). Based on their morphometric characteristics, two different types of grains were identified and assigned to the Triticeae and Paniceae tribes of the Poaceae family (grasses). Both morphotypes were documented in suitable proportions. Despite the small size of the calculus samples, most of the starch grains had an excellent preservation state. Microremains, like the ones identified in the analyzed dental calculus, were not retrieved in the dust traps placed in the laboratory, supporting the archaeological nature of the microfossils identified in the calculus samples. Sediment samples were analyzed as well and showed a great diversity of microremains, including phytoliths and fungal spores, much more abundant than in the dental calculus samples. Starch grains were not recovered from sediment samples.
Morphotype I
The first morphotype of starch grains represents 60.8% of the total, and it is characterized by a bimodal distribution typical of most grasses of the Triticeae tribe (Supplementary Table 3) (Fig. 4E–H). Such distribution is equally frequent in the samples. Large grains (≤ 15 μm), defined as type A, are round to oval in 2D shape, lenticular in 3D, with central or slightly sunken hilum and few lamellae. The largest grains are similar to those of cereals like wheat and barley. Smaller grains (< 10 μm), defined as type B, are almost spherical and have a central hilum43,44. They potentially also belong to the Poaceae family, although similar grains can be found in many other plants. The mean dimension of these starch grains is 13.8 μm.

(A–L) Starch grains embedded in El Espinoso's dental calculus in light microscope and polarised light microscope. (A–H) Polyhedral starch grains with central hilum and fissures associated with the Paniceae tribe; (I–L) Round to oval starch grains "bimodal distribution" consistent with the Triticeae tribe. (Scale bar, 20 μm).
Morphotype II
39.2% of the starch grains come from this morphotype. They have a 3D polyhedral shape, with a central hilum and fissures radiating from it. The extinction cross is visible, with straight arms, while lamellae are not identified (Supplementary Table 3) (Fig. 4A–D). This type of grain is abundant in the samples and is consistent with those of the Paniceae tribe, likely representing millets. The mean dimension of these starch grains is 14.5 μm.
A variety of other plant structures were found as well (Supplementary Fig. 8). These microremains of probable non-dietary origin include fungal spores, charcoals, fibres, wooden tissues, and several conifer pollen granules.
Discussion
Stable isotope evidence of millet consumption
The El Espinoso animal samples had δ15N signatures 3–4‰ lower than the human ones, which constitutes the expected relationship between consumers and prey45,46. Nitrogen isotopes from other Bronze Age contexts in northern Iberia, such as Cova do Santo47 and La Castañera48, show similar trends between humans and animals (Fig. 5). The animals had δ13C values typical for a terrestrial European C3 ecosystem49, while the carbon results from El Espinoso human individuals showed that they were significantly enriched in 13C, suggesting an intake of C4 plants or marine foods50,51. As the length of the aquatic food chain is longer than the terrestrial one, nitrogen isotope values can be used to discard the hypothesis that the individuals ate marine foods, which would reflect higher nitrogen values, especially fish or mammals46, which would be reflected in higher δ15N values. Shellfish can overlap signals with C4 plants, but despite the proximity of the site to the sea, there is no archaeological evidence of marine resource consumption (shells or fish remains) during the regional Bronze Age, neither in the cave deposits nor in the few documented open-air sites38,52. However, future archaeomalacological and ichthyofaunal studies will help to verify this.

δ13C and δ15N values of bone collagen analysed from northern Atlantic Iberian Bronze Age sites, including humans and animals from El Espinoso (Late Bronze Age); and La Llana, Los Cinchos and La Fragua (Early/Middle Bronze Age) in this study. Cova do Santo, La Castañera, La Garma A, Santimamiñe (Early/Middle Bronze Age) and Ondarre (Late Bronze Age) from previously published data.
In addition, to reject the possible influence of fish on the El Espinoso human diet due to the higher δ13C values, sulfur isotope ratios were also analyzed. The freshwater and marine resources consumption produce elevated δ34S values53. At El Espinoso, humans but also faunal specimens, who follow different diets, showed the dispersion of the same values ranging between 13.5 and 16.9‰. On the other hand, La Llana, La Fragua and Los Cinchos individuals reflected similar sulfur values, but they did not provide enrichment of carbon values. Therefore, the δ34S values recorded were more likely to be associated with the environment where they were living rather than with the influence of fish in the diet. Indeed, humans and animals of El Espinoso were predominantly living near the coast, where the sea spray effect can reach more than 30 km inland, affecting the sulfur signatures of coastal environments54. In this sense, the sulfur isotope values from humans and animals of Los Avellanos I and II, dated in the Late Neolithic/Chalcolithic and without typical carbon isotope values of marine diets, showed the same higher sulfur isotope signals55. Consequently, the most likely hypothesis to explain the δ13C enrichment in El Espinoso individuals could be a significant consumption of C4 plants, likely millets. Isotopic measurements of modern millets have provided δ13C values from − 10 to − 12‰ and δ15N values from 3 to 4‰56, as well as isotopic measurements of Iron Age millets have reported δ13C and δ15N values around − 10‰ and 4‰57. Millet has a protein content of approximately 10%; thus, a significant intake of these crops is required to detect a substantial change in collagen isotope ratios58.
In contrast to El Espinoso, the isotopic data from the humans of La Llana, Los Cinchos, and La Fragua sites reported lower δ13C values, showing that the diet was predominantly terrestrial, based on C3 plants and animal protein. This dietary pattern was also observed in other human bone collagen isotopic studies from the Bronze Age sites, such as Cova do Santo47, La Castañera48, La Garma A59, Santimamiñe60 and Ondarre61 (Fig. 5). These archaeological sites are dated to the Early and Middle Bronze Age, except for one individual from Ondarre, which belongs to the Late Bronze Age. Still, surprisingly, it showed an opposite carbon value to the El Espinoso group. In all of the groups mentioned above, isotopic signals of C4 plant consumption were not detected, which suggests that millets were probably not eaten yet in northern Iberia during these periods. In this sense, dentine sequential analysis of carbon and nitrogen isotopes from seven human teeth documented in Los Canes cave (Asturias), dated in the Late Bronze Age, showed isotopic signals of C4 plant consumption during the childhood, while two more teeth dated in the Early Bronze Age indicated a C3 exclusive diet62. With the current isotopic evidence, if C4 plant resources were consumed before the Late Bronze Age, the quantities were not high enough to be registered in the long-term bone collagen record. This would suggest infrequent or occasional cultivation of millets during the Middle Bronze Age, becoming a staple crop only in later periods, probably from Late Bronze Age, as the isotopic results from El Espinoso cave demonstrate. Until now, the earliest bone collagen isotopic evidence of C4 plant consumption in northern Iberia is documented at La Hoya village, an inland site 150 km from the current Atlantic sea in Alava and dated in the Iron Age37. El Espinoso individuals are more than 700 years older than La Hoya, setting back the isotopic evidence of intensive millet consumption in northern Iberia to the Late Bronze Age.
According to the evidence, millets were introduced into Iberia across the Pyrenees from Central Europe during the Middle/Late Bronze Age transition, reaching earlier and playing a more important role in the northern regions of Iberia than the southern ones63. A longitudinal gradient for the dispersal from Eastern to Western Europe during recent prehistory is proposed, with the Atlantic coast being one of the last regions to adopt millets. However, the radiocarbon evidence indicates a relatively quick spread of these crops through Europe20. Concerning the published stable isotope analyses from different European regions, some studies have found isotopic evidence of millet consumption around 5500 cal. BC in East Asia64,65, between 2500 and 2000 cal. BC in Central Asia17,66, between 1600 and 1400 cal. BC in the Caucasus region67, between 1600 and 1200 cal. BC in northern and central Italy68,69, between 1400 and 1200 cal. BC in France70 and between 1300 and 1000 cal. BC in northern Spain.
Regarding the carbon values across the humans’ age distribution of El Espinoso, three out of four subadults showed the higher carbon values of the sample, suggesting the existence of inequalities in C4 plant consumption by age. These minor differences can be explained by the higher importance of millet in the children's diet. This pattern was previously observed at La Hoya and Los Canes62,71. Here millets were introduced to infants and young children during the weaning process, probably in the form of porridge57. Considering the stable isotope data from different millet consumers of Europe, in some cases, all the individuals of the same group ate large amounts of millet, while in others, the millet consumers lived in a community where most people did not consume this crop. This fact suggests dietary differences related to social, cultural, or economic status33. However, the current isotopic evidence indicates that intensive consumption of millets outside China was sporadic during recent prehistory until the first millennium cal. BC, when the results tend to be more homogeneous from the Eurasian steppes to the Atlantic fringe.
Dental calculus evidence of millet consumption
The richest archaeological evidence of millet grains within dental calculus comes from Asia, where these crops were primarily domesticated. Different studies have revealed starch grains and phytoliths from other millet species72,73,74,75,76. Based on the phytolith and starch grain morphology, these crops show great potential for identifying different genera or species, opening new possibilities for their detection in archaeological contexts31. There is little evidence of preserved millet grains in dental calculus from Europe, with even fewer examples in Iberia. This lack of knowledge can be explained by the absence of studies on the recent prehistory of this geographical area. Among a great diversity of taxa represented, some starches of the Paniceae tribe were recovered within the dental calculus of the Chalcolithic individuals from El Mirador cave (Burgos, Spain), based on their average size and morphological features77. In Italy, 79 starch grains were identified among individuals from the Chalcolithic/Bronze Age site of Grotta dello Scoglietto (Tuscany)28.
Polyhedral starch grains (Morphotype II) found in the El Espinoso individuals may be attributed to different C4 plants. Experimental starch grains from Panicum miliaceum (Supplementary Fig. 5) or Setaria italica (Supplementary Fig. 6) conducted in this work share these sizes and morphological features, although slight differences can be identified in the size classes and fissure patterns. Setaria italica can have occasional oval grains and a centric hilum traversed by fissures which vary in form. In contrast, starch grains from Panicum miliaceum have a mostly polyhedral shape with a centric hilum where fissures are less common32. Experimental starch grains size differs slightly between both species, being marginally smaller than the grains of Panicum miliaceum. Wild millet species, such as Setaria viridis or Setaria verticillata are also smaller, with a more characteristic spherical morphology and short fissures31 (Supplementary Table 4) (Supplementary Fig. 9). The morphological features and the mean dimensions of the archaeological starch grains suggest that Setaria italica was the crop consumed by this human group. However, both species might have been consumed in a mixed way since millet polycropping has been suggested archaeologically and documented ethnographically7,63. The presence of millet starches in El Espinoso individuals, although in small quantities, can be considered conclusive, confirming the millet consumption suggested by the stable isotope values. However, phytoliths typical of millet species have not been documented. The low presence of starch grains and the absence of phytoliths could be explained by the small size of the calculus samples. Only small amounts of dental calculus could be sampled from most individuals, even less than 1 mg (Supplementary Table 3). This can be considered satisfactory since it has allowed microfossils extraction in such small quantities, establishing size limits for further studies.
Starch grains with an oval morphology and "bimodal distribution" (Morphotype I), typical of grass grains, as those of the Triticeae tribe (Supplementary Fig. 7), were also identified in reasonable quantities as millet starch grains. Previous experimental analyses in species of the Triticeae tribe have revealed that wild starch grains (Aegilops genus) show a larger size distribution than domestic ones (Triticum and Hordeum genus)78,79. Additionally, B-type grains are more abundant than A-type in wheat and barley species. Our experimental measurements have shown the great variety of starch grain sizes for the different Triticeae domestic species (Supplementary Table 4) (Supplementary Fig. 9). However, the morphology and size obtained for the archaeological Morphotype I starch grains are consistent with the wheat and barley species. This evidence complements the isotopic data that indicates the great importance of millet in the human diet at El Espinoso and the relevance of other C3 plants. As the millets allowed for two growing seasons per year, other crops such as wheat or barley could also have been cultivated along with them annually. These findings reveal diversified agriculture during this time, with the millet being complemented by meat, dairy products, and other cereals. Concerning the other plant residues entrapped within the tartar, they may have been related to a wide range of non-dietary activities such as oral hygiene with toothpicks, the para-masticatory use of the mouth in various tasks, inhalation of airborne paleoenvironmental debris due to the exposure to hearths, and craft activities such as cordage, textile, basketry or net-making80.
Macro-botanical evidence of millets in the archaeological record of northern Iberia
Millets are poorly represented in the Bronze Age archaeological record. The first carpological evidence of millet exploitation in Iberia dates to the Middle Bronze Age. However, they were not systematically exploited until the Late Bronze Age and Iron Age1. No carpological evidence of the wild native species that are relatives of the domesticated millets (Panicum repens, Setaria viridis, or Setaria verticillata), also known as “forgotten millets”, has been found during the recent prehistory of northern Iberia81. Today they are widely distributed across central and south Europe82 but almost unknown in Atlantic Europe83. The only C4 plants found in Iberia during recent prehistory are domestic millets, particularly broomcorn and foxtail millets (Panicum miliaceum and Setaria italica). They are the C4 crops that are frequently identified across Europe33, constituting the primary candidates cultivated in large proportions by northern Iberian farmers. In the Vizcaya province, macroremains of foxtail millet were discovered in Kobaederra (Level 1) and Arenaza (Layer 9), chronologically assigned to the Chalcolithic and Early Bronze Age, respectively84. However, those seeds have not been directly dated; thus, we cannot exclude the possibility that they might be intrusive. Alternatively, there is an absence of Panicum milliaceum in the prehistoric carpological records of the Cantabrian Region. Also, in northern Portugal, several millet grains were found in the Chalcolithic sequence of Crasto de Palheiros85, but stratigraphic issues were reported in those layers. In the Middle Bronze Age in northern Portugal, the carpological remains of broomcorn millet were identified in the sites of Sola IIb86 and Terraço das Laranjeiras87. Later, during the Late Bronze Age, a higher presence of millet (exclusively P. miliaceum) is recorded across the region, in the neighboring areas of Galicia province (A Fontela, Penalba, and Penarrubia) and northern Portugal (Castelo de Matos, São Julião, Santinha, and Senhora da Guia)9. Likewise, seeds from both millet species have been identified in different sites in the Ebro basin (northern Mediterranean Iberia) during the Middle and Late Bronze Age, such as at Punta Farisa, Masada de Ratón, El Vilot and Vincament88,89.
Consequently, the available evidence confirms that millet cultivation in northern Iberia seems to have started during the Middle/Late Bronze Age transition. However, it was not until the Late Bronze Age that millets became a significant and regular crop. The transition from the Middle to Late Bronze Age marks the shift of millets from a minor to a major role in the agriculture of the European farming societies90. To better understand the origin and development of millet cultivation, direct dating of millet grains will be needed20 as the morphology and size of these tiny seeds facilitate their vertical movements downwards through the stratigraphical sequences and must be called into question for future research23.
Ethnographic insights of millets in northern Spain
Ethnographic studies can provide valuable information that can be used as a complement to the archaeological evidence about the cultivation and consumption of millets in the past. This is the case in northern Spain, where Setaria italica (foxtail millet) and Panicum miliaceum (broomcorn millet) were traditionally cultivated. Ethnobotanical data have shown a higher presence of S. italica or both species sown together in the Asturias province. Such place-names as "Panizales, Panicera, Paniceres," etc., are familiar in the Asturias and refer to the Spanish name of foxtail millet91. At the same time, the most common crop found in Galicia and northern Portugal is P. miliaceum7. This coincides with the archaeobotanical remains recovered from prehistoric sites, with foxtail millet found in the Cantabrian Region and the broomcorn millet in the northwestern area of Iberia. Ethnographic approaches in Galicia during the twentieth century have provided insights into the utilization of millet grains for human consumption. Simultaneously, both grains and plants are used as feed or fodder for animals35. In recent times in the wider northwestern regions of Iberia, millet has been used to make porridge, stews, puddings, or bread for human consumption7. However, there are some differences observed among regions. In Asturias, S. italica is used exclusively for foddering, while in Galicia and northern Portugal, besides animal consumption, P. miliaceum, is also used as human food. Both crops have been cultivated until the present, and their importance was exceptionally high during the Roman and Medieval periods, as testified by ancient sources92 and stable isotope analyses93. However, the arrival of maize (Zea mays) from America in the seventeenth century displaced their cultivation to a marginal role. Also, ethnographic studies in other European regions reported the use of millets to produce an alcoholic drink94.
Conclusions
In this study, we infer direct and precise data about the systematic use of millets during the Late Bronze Age in northern Iberia by combining different methodologies approaches on human remains including: (1) radiocarbon dating; (2) stable isotope (δ13C, δ15N and δ34S); and (3) dental calculus analysis. El Espinoso cave was used as a burial location during the last third of the second millennium cal. BC. The Bayesian model places the span of the burial phase between 0 and 115 years, supporting a practically synchronous use of the cave for a few decades. δ13C and δ15N isotope values show a diet with high consumption of C4 plants, likely millets, and contrast with the values obtained in different Early and Middle Bronze Age funerary sites for the same region, suggesting that the systematic consumption of millets in this area started from the Late Bronze Age. The study of the microresidues entrapped within the dental calculus of El Espinoso individuals confirms the presence of starch grains with a polyhedral morphology typical of the Paniceae tribe. Besides, other round to oval starch grains characteristic of the Triticeae tribe have also been identified, demonstrating a mixed agricultural economy for the Late Bronze Age of northern Iberia. The multidisciplinary research undertaken on the human bone assemblage of El Espinoso cave has generated a corpus of high-quality biomolecular data about millet consumption and constitutes one of the oldest and most direct evidence of the exploitation of millets in northern Atlantic Iberia.
Methods
In this research, a multidisciplinary approach was conducted to reassess the relevance of millets in the prehistoric human diet. Radiocarbon dating was carried out to establish a precise chronology for the humans analyzed here. The dates were calibrated in OxCal v4.495 using the IntCal20 calibration curve96. All results are presented at a 95.4% probability. The results were modelled using Bayesian statistics. Stable isotope analyses (δ13C, δ15N, δ34S) of humans and animals were performed to ascertain these individuals' carbon, nitrogen, and sulfur signatures. Bone collagen extraction was conducted in the EvoAdapta Group (University of Cantabria) according to the procedures proposed by Richards and Hedges97. Carbon, nitrogen and sulfur isotope analysis of the collagen samples was undertaken by Elemental Analysis—Isotope Ratio Mass Spectrometry (EA-IRMS) in Iso-analytical laboratory (Crewe, UK). Quality indicators widely used were employed39,40,41,42. Dental calculus analysis was achieved to discern what types of plants were involved in the diet of this human group. The dental calculus matrix was removed from the teeth following the protocol by Sabin and Fellow-Yates98. Decontamination and the extraction procedures for micro-debris were carried out following standard protocols as described by Cristiani et al.43,99 and Fiorin et al.100, and were conducted in dedicated clean spaces under strict environmental monitoring of the DANTE—Diet and ANcient TEchnology laboratory of Sapienza University of Rome (Italy). The analysis of the microfossils was carried out using a Zeiss Imager2 polarised microscope (100×–630×) at the DANTE laboratory and a Leica DVM6 M digital microscope at the EvoAdapta Group. Morphological and size comparisons were carried out with the modern plant reference collections housed at these institutions (See Supplementary Text 5 for more detailed descriptions of methods).
Data availability
All the data reported in this article are provided in the Figures and Tables of the Supplementary Information and Manuscript files. Radiocarbon dates are listed in the Supplementary Table 1, as well as the Bayesian modelling which are included in Fig. 2. Stable isotope analyses on bone collagen (δ13C, δ15N and δ34S) and the quality indicators of the samples were provided in Supplementary Table 2 and plotted in Fig. 3. The archaeological microremains extracted from the dental calculus are described in Supplementary Table 3 and shown in Fig. 4. and Supplementary Fig. 8. Experimental starch grains from the species involved in this study are included in Supplementary Figs. 5, 6 and 7. Statistical analysis of the length measurement of these starch grains is reported in Supplementary Table 4 and Supplementary Fig. 9. All the other chronological and regional data supporting the findings and interpretations of this study are available in existing publications referenced in the text and Supplementary Information. The archaeological remains studied and sampled in this research are curated in the laboratory of EvoAdapta Group (University of Cantabria, Spain). These materials will be returned to Museo Arqueológico de Asturias (Oviedo, Spain) and Museo de Prehistoria y Arqueología de Cantabria (Santander, Spain).
References
Buxó, R. & Piqué, R. Arqueobotánica: Los Usos de las Plantas en la Península Ibérica. (Grupo Planeta GBS, 2008).
Miller, N. F., Spengler, R. N. & Frachetti, M. Millet cultivation across Eurasia: Origins, spread, and the influence of seasonal climate. Holocene 26, 1566–1575 (2016).
James, T. K., Rahman, A., McGill, C. R. & Trivedi, P. D. Biology and survival of broomcorn millet (Panicum miliaceum) seed. N. Z. Plant Prot. 64, 142–148 (2011).
Kirleis, W., Dal Corso, M. & Filipović, D. Millet and What Else?: The Wider Context of the Adoption of Millet Cultivation in Europe. vol. 14 (Sidestone Press, 2022).
Sherratt, A. Water, soil and seasonality in early cereal cultivation. World Archaeol. 11, 313–330 (1980).
Rachie, K. O. The Millets: Importance, Utilization and Outlook. 74 (International Crops Research Institute for the Semi-Arid Tropics, 1975).
Moreno-Larrazabal, A., Teira-Brión, A., Sopelana-Salcedo, I., Arranz-Otaegui, A. & Zapata, L. Ethnobotany of millet cultivation in the north of the Iberian Peninsula. Veg. Hist. Archaeobot. 24, 541–554 (2015).
Liu, L. et al. The origins of specialized pottery and diverse alcohol fermentation techniques in Early Neolithic China. Proc. Natl. Acad. Sci. USA 116, 12767–12774 (2019).
Tereso, J. P. et al. Agriculture in NW Iberia during the Bronze Age: A review of archaeobotanical data. J. Archaeol. Sci. Rep. 10, 44–58 (2016).
Liu, C., Kong, Z. & Lang, S. D. A discussion on agricultural and botanical remains and the human ecology of Dadiwan site, in Chinese. Zhongyuan Wenwu 4, 25–29 (2004).
Zhao, Z. New archaeobotanic data for the study of the origins of agriculture in China. Curr. Anthropol. 52, S295–S306 (2011).
Crawford, G. W., Xuexiang, C., Fengshi, L. & Jianhua, W. A Preliminary analysis on plant remains of the Yuezhuang site in Changqing District, Jinan City, Shandong Province. Jianghan Archaeol. 2, 107–113 (2013).
Frachetti, M. D. Multiregional emergence of mobile pastoralism and nonuniform institutional complexity across Eurasia. Curr. Anthropol. 53, 2–38. https://doi.org/10.1086/663692 (2012).
Ventresca Miller, A. R. & Makarewicz, C. A. Intensification in pastoralist cereal use coincides with the expansion of trans-regional networks in the Eurasian Steppe. Sci. Rep. 9, 8363 (2019).
Jones, M. et al. Food globalisation in prehistory: The agrarian foundations of an interconnected continent. J. Br. Acad. 4, 73–87 (2016).
Spengler, R. et al. Early agriculture and crop transmission among Bronze Age mobile pastoralists of Central Eurasia. Proc. Biol. Sci. 281, 20133382 (2014).
Hermes, T. R. et al. Early integration of pastoralism and millet cultivation in Bronze Age Eurasia. Proc. Biol. Sci. 286, 20191273 (2019).
Frachetti, M. D., Spengler, R. N., Fritz, G. J. & Maryashev, A. N. Earliest direct evidence for broomcorn millet and wheat in the central Eurasian steppe region. Antiquity 84, 993–1010 (2010).
Motuzaite-Matuzeviciute, G., Richard, A. S., Hunt, H. V., Liu, X. & Jones, M. K. The early chronology of broomcorn millet (Panicum Miliaceum) in Europe. Antiquity 338, 1073–1085 (2013).
Filipović, D. et al. New AMS 14C dates track the arrival and spread of broomcorn millet cultivation and agricultural change in prehistoric Europe. Sci. Rep. 10, 13698 (2020).
Hunt, H. V. et al. Millets across Eurasia: Chronology and context of early records of the genera Panicum and Setaria from archaeological sites in the Old World. Veg. Hist. Archaeobot. 17, 5–18 (2008).
Brudenell, M., Fosberry, R., Phillips, T. & Kwiatkowska, M. Early cultivation of broomcorn millet in southern Britain: Evidence from the Late Bronze Age settlement site of Old Catton, Norfolk. Antiquity 2022, 1–6 (2022).
Weber, S. A. & Fuller, D. Q. Millets and their role in early agriculture. Pragdhara 18, 69–90 (2007).
Shelton, C. P. & White, C. E. The hand-pump flotation system: A new method for archaeobotanical recovery. J. Field Archaeol. 35, 316–326 (2010).
Barboff, M. Le millet au Portugal. In Millet– Hirse–Millet. Actes du Congres d’Aizenay (ed. Hörandner, E.) 113–122 (Grazer Beitra¨ge zur 731 europa¨ischen Ethnologie, 1995).
Reddy, S. N. If the Threshing Floor Could Talk: Integration of Agriculture and Pastoralism during the Late Harappan in Gujarat, India. J. Anthropol. Archaeol. 16, 162–187 (1997).
Dayakar Rao, B. et al. Nutritional and health benefits of millets. In ICAR_Indian Institute of Millets Research (IIMR), Rajendranagar, Hyderabad 112 (2017).
Mariotti-Lippi, M., Pisaneschi, L., Sarti, L., Lari, M. & Moggi-Cecchi, J. Insights into the Copper-Bronze Age diet in Central Italy: Plant microremains in dental calculus from Grotta dello Scoglietto (Southern Tuscany, Italy). J. Archaeol. Sci. Rep. 15, 30–39 (2017).
Lu, H. et al. Phytoliths analysis for the discrimination of Foxtail millet (Setaria italica) and Common millet (Panicum miliaceum). PLoS ONE 4, e4448 (2009).
Lucarini, G., Radini, A., Barton, H. & Barker, G. The exploitation of wild plants in Neolithic North Africa. Use-wear and residue analysis on ground stone tools from the Farafra Oasis, Egypt. Quat. Int. 410, 77–92 (2016).
Madella, M., Lancelotti, C. & García-Granero, J. J. Millet microremains—an alternative approach to understand cultivation and use of critical crops in Prehistory. Archaeol. Anthropol. Sci. 8, 17–28 (2016).
Yang, X. et al. From the modern to the archaeological: Starch grains from millets and their wild relatives in China. J. Archaeol. Sci. 39, 247–254 (2012).
Lightfoot, E., Liu, X. & Jones, M. K. Why move starchy cereals? A review of the isotopic evidence for prehistoric millet consumption across Eurasia. World Archaeol. 45, 574–623 (2013).
Armendariz, A. In Las cuevas sepulcrales del País Vasco. (Tesis Doctoral Inédita, Universidad del País Vasco-Euskal Herriko Unibertsitatea, 1992).
Vazquez-Varela, J. M. El cultivo del mijo, (Panicum miliaceum, L.), en la cultura castreña del noroeste de la peninsula iberica. Cuad. Estud. Gallegos 1, 65–73 (1994).
Patterson, N. et al. Large-scale migration into Britain during the Middle to Late Bronze Age. Nature 601, 588–594 (2021).
Olalde, I. et al. The genomic history of the Iberian Peninsula over the past 8000 years. Science 363, 1230–1234 (2019).
Arias, P. & Armendariz, A. Aproximación a la Edad del Bronce en la región cantábrica. A Idade do Bronce en Galicia: novas perspectivas. Cadernos Semin. Sargadelos 77, 47–80 (1998).
DeNiro, M. J. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317, 806–809 (1985).
Ambrose, S. H. Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17, 431–451 (1990).
van Klinken, G. J. Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Archaeol. Sci. 26, 687–695 (1999).
Nehlich, O. & Richards, M. P. Establishing collagen quality criteria for sulphur isotope analysis of archaeological bone collagen. Archaeol. Anthropol. Sci. 1, 59–75 (2009).
Cristiani, E., Radini, A., Edinborough, M. & Borić, D. Dental calculus reveals Mesolithic foragers in the Balkans consumed domesticated plant foods. Proc. Natl. Acad. Sci. USA 113, 10298–10303 (2016).
Henry, A. G. & Piperno, D. R. Using plant microfossils from dental calculus to recover human diet: A case study from Tell al-Raqā’I, Syria. J. Archaeol. Sci. 35, 1943–1950 (2008).
Minagawa, M. & Wada, E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. Acta 48, 1135–1140 (1984).
Hedges, R. E. M. & Reynard, L. M. Nitrogen isotopes and the trophic level of humans in archaeology. J. Archaeol. Sci. 34, 1240–1251 (2007).
López-Costas, O., Müldner, G. & Martínez-Cortizas, A. Diet and lifestyle in Bronze Age Northwest Spain: The collective burial of Cova do Santo. J. Archaeol. Sci. 55, 209–218 (2015).
Jones, J. R. et al. Investigating prehistoric diet and lifeways of early farmers in central northern Spain (3000–1500 CAL BC) using stable isotope techniques. Archaeol. Anthropol. Sci. 11, 3979–3994 (2019).
DeNiro, M. J. & Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 42, 495–506 (1978).
O’Leary, M. H. Carbon isotope fractionation in plants. Phytochemistry 20, 553–567 (1981).
Chisholm, B. S., Nelson, D. E. & Schwarcz, H. P. Stable-carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science 216, 1131–1132 (1982).
de Blas Cortina, M. Á. De la caverna al lugar fortificado: Una mirada a la edad del bronce en el territorio Astur-Cántabro. Quad. Prehist. Arqueol. Castelló 29, 105–134 (2011).
Nehlich, O. The application of sulphur isotope analyses in archaeological research: A review. Earth-Sci. Rev. 142, 1–17 (2015).
Richards, M. P., Fuller, B. T. & Hedges, R. E. M. Sulphur isotopic variation in ancient bone collagen from Europe: Implications for human palaeodiet, residence mobility, and modern pollutant studies. Earth Planet. Sci. Lett. 191, 185–190 (2001).
González-Rabanal, B. et al. Diet, mobility and death of Late Neolithic and Chalcolithic groups of the Cantabrian Region (northern Spain). A multidisciplinary approach towards studying the Los Avellanos I and II burial caves. J. Archaeol. Sci. Rep. 34, 1–13 (2020).
McGovern, P. E. et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 101, 17593–17598 (2004).
Fernández-Crespo, T., Ordoño, J., Bogaard, A., Llanos, A. & Schulting, R. A snapshot of subsistence in Iron Age Iberia: The case of La Hoya village. J. Archaeol. Sci. Rep. 28, 1–10 (2019).
Hedges, R. E. M. On bone collagen?apatite-carbonate isotopic relationships. Int. J. Osteoarchaeol. 13, 66–79 (2003).
Arias, P. Determinaciones de isótopos estables en restos humanos de la región Cantábrica. Aportación al estudio de la dieta de las poblaciones del Mesolítico y el Neolítico. Munibe Antropol.-Arkeol. 57, 359–374 (2005).
Palencia-Madrid, L. et al. Ancient mitochondrial lineages support the prehistoric maternal root of Basques in Northern Iberian Peninsula. Eur. J. Hum. Genet. 25, 631–636 (2017).
Fernández-Crespo, T., Mujika, J. A. & Ordoño, J. Aproximación al patrón alimentario de los inhumados en la cista de la Edad del Bronce de Ondarre (Aralar, Guipúzcoa) a través del análisis de isótopos estables de carbono y nitrógeno sobre colágeno óseo. Trab. Prehist. 73, 325–334 (2016).
Higuero Pliego, A. In Análisis Isotópico de Carbono y Nitrógeno en Secuencias de Dentina y de Estroncio en Esmalte Procedente de Restos Humanos Prehistóricos de la Cueva de Los Canes (Cabrales, Asturias). (Tesis Doctoral Inédita, Universidad de Cantabria, 2020).
Teira-Brión, A. Traditional millet cultivation in the Iberian Peninsula: Ethnoarchaeological reflections through the lens of social relations and economic concerns. In (eds. Kirleis, W. et al.) Millet and What Else?: The Wider Context of the Adoption of Millet Cultivation in Europe vol. 14 263–278 (Sidestone Press, 2022).
Pechenkina, E. A., Ambrose, S. H., Xiaolin, M. & Benfer, R. A. Reconstructing northern Chinese Neolithic subsistence practices by isotopic analysis. J. Archaeol. Sci. 32, 1176–1189 (2005).
Hu, Y. et al. Palaeodietary study of Sanxingcun Site, Jintan, Jiangsu. Chin. Sci. Bull. 52, 660–664 (2007).
Motuzaite-Matuzeviciute, G., Ananyevskaya, E., Sakalauskaite, J., Soltobaev, O. & Tabaldiev, K. The integration of millet into the diet of Central Asian populations in the third millennium BC. Antiquity 96, 560–574 (2022).
Herrscher, E. et al. The origins of millet cultivation in the Caucasus: Archaeological and archaeometric approaches. Préhistoires Méditerr. 2018, 6 (2018).
Tafuri, M. A., Craig, O. E. & Canci, A. Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Am. J. Phys. Anthropol. 139, 146–153 (2009).
Varalli, A., Moggi-Cecchi, J., Moroni, A. & Goude, G. Dietary variability during bronze age in central Italy: First results. Int. J. Osteoarchaeol. 26, 431–446 (2016).
Goude, G., Rey, L., Toulemonde, F., Cervel, M. & Rottier, S. Dietary changes and millet consumption in northern France at the end of Prehistory: Evidence from archaeobotanical and stable isotope data. Environ. Archaeol. 22, 268–282 (2017).
Fernández-Crespo, T., Ordoño, J., Llanos, A. & Schulting, R. J. Make a desert and call it peace: Massacre at the Iberian Iron Age village of La Hoya. Antiquity 94, 1245–1262 (2020).
Lu, H. et al. Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10,000 years ago. Proc. Natl. Acad. Sci. USA 106, 7367–7372 (2009).
Li, M. et al. Starch grains from dental calculus reveal ancient plant foodstuffs at Chenqimogou site, Gansu Province. Sci. China Earth Sci. 53, 694–699 (2010).
Zhang, J., Lu, H., Wu, N., Yang, X. & Diao, X. Phytolith analysis for differentiating between foxtail millet (Setaria italica) and Green Foxtail (Setaria viridis). PLoS ONE 6, e19726 (2011).
Ge, Y. et al. Phytolith analysis for the identification of barnyard millet (Echinochloa sp.) and its implications. Archaeol. Anthropol. Sci. 10, 61–73 (2018).
Tao, D., Zhang, G., Zhou, Y. & Zhao, H. Investigating wheat consumption based on multiple evidences: Stable isotope analysis on human bone and starch grain analysis on dental calculus of humans from the Laodaojing cemetery, Central Plains, China. Int. J. Osteoarchaeol. 30, 594–606 (2020).
Bucchi, A., Burguet-Coca, A., Expósito, I., Aceituno-Bocanegra, F. J. & Lozano, M. Comparisons between methods for analyzing dental calculus samples from El Mirador cave (Sierra de Atapuerca, Spain). Archaeol. Anthropol. Sci. 11, 6305–6314 (2019).
Cristiani, E. et al. Wild cereal grain consumption among Early Holocene foragers of the Balkans predates the arrival of agriculture. Elife 10, 1–37 (2021).
Bocanegra, F. J. A. & Sáez, J. A. L. Caracterización morfológica de almidones de los géneros Triticum y Hordeum en la Península Ibérica. Trabprehist 69, 332–348 (2012).
Hardy, K., Buckley, S. & Copeland, L. Pleistocene dental calculus: Recovering information on Paleolithic food items, medicines, paleoenvironment and microbes. Evol. Anthropol. 27, 234–246 (2018).
López-Dóriga, I. In The use of plants during the Mesolithic and the Neolithic in the Atlantic coast of the Iberian peninsula. (Tesis Doctoral Inédita, Universidad de Cantabria, 2016).
Nava, A. et al. Multipronged dental analyses reveal dietary differences in last foragers and first farmers at Grotta Continenza, central Italy (15,500–7000 BP). Sci. Rep. 11, 1–14 (2021).
Pyankov, V. I., Ziegler, H., Akhani, H., Deigele, C. & Lüttge, U. European plants with C4 photosynthesis: Geographical and taxonomic distribution and relations to climate parameters. Bot. J. Linn. Soc. 163, 283–304 (2010).
Zapata, L. In La explotación de los recursos vegetales y el origen de la agricultura en el País Vasco. (Tesis Doctora Inédita, Universidad del País Vasco, 2002).
Figueiral, I., de-Jesus-Sanches, M. & Cardoso, J. L. Crasto de Palheiros (Murça, NE Portugal, 3rd - 1st millennium BC): From archaeological remains to ordinary life. Estudos Quat. 17, 13–28 (2017).
Bettencourt, A. M. S. O povoado da Idade do Bronze da Sola, Braga, norte de Portugal. Cadernos Arqueol. 9, 29–44 (2000).
Jesus, A., Tereso, J. P. & Gaspar, R. Interpretative trajectories towards the understanding of negative features using Terraço das Laranjeiras Bronze Age site as a case study. J. Archaeol. Sci. Rep. 30, 1–14 (2020).
Alonso-Martínez, N. Registro arqueobotánico de Cataluña occidental durante el II y I milenio a.n.e.. Complutum 11, 221–238 (2000).
Tarongi-Chavarri, M. Análisis comparativo de los estudios carpológicos de la Depresión del Ebro durante el III y I milenio a. C. Un estado de la cuestión. Rev. d’arqueologia Ponent 27, 41–59 (2017).
Stika, H.-P. & Heiss, A. G. Plant cultivation in the Bronze Age. In The Oxford Handbook of the European Bronze Age (eds. Fokkens, H. & Harding, A.) 348–369 (2013).
González-y-Fernández-Valles, J. M. Temas de toponimia asturiana. Archivum 21, 121–140 (1971).
de Carvallo, L. A. In Antiguedades y Cosas Memorables del Principado de Asturias. (Julian de Paredes, 1695).
MacKinnon, A. T., Passalacqua, N. V. & Bartelink, E. J. Exploring diet and status in the Medieval and Modern periods of Asturias, Spain, using stable isotopes from bone collagen. Archaeol. Anthropol. Sci. 11, 3837–3855. https://doi.org/10.1007/s12520-019-00819-2 (2019).
Renfrew, J. M. Palaeoethnobotany: The Prehistoric Food Plants of the Near East and Europe (Columbia University Press, 1973).
Bronk Ramsey, C. Bayesian Analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Reimer, P. J. et al. The IntCal20 Northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Richards, M. P. & Hedges, R. E. M. Stable isotope evidence for similarities in the types of marine foods used by late mesolithic humans at sites along the Atlantic Coast of Europe. J. Archaeol. Sci. 26, 717–722 (1999).
Sabin, S. & James, A. In Dental Calculus Field-Sampling Protocol (Sabin version) v2 (protocols.io.bqecmtaw). (2020). https://doi.org/10.17504/protocols.io.bqecmtaw.
Cristiani, E. et al. Dental calculus and isotopes provide direct evidence of fish and plant consumption in Mesolithic Mediterranean. Sci. Rep. 8, 8147 (2018).
Fiorin, E. et al. Combining dental calculus with isotope analysis in the Alps: New evidence from the Roman and medieval cemeteries of Lamon. Italy. Quat. Int. https://doi.org/10.1016/j.quaint.2021.11.022 (2021).
Acknowledgements
This study is part of B.G.R.’s Doctoral dissertation, supervised by A.B.M.A. and M.R.G.M. This research is funded by the research projects of the Spanish Economy, Industry and Competitiveness Ministry HAR2016-75605-R to M.R.G.M. and HAR2017- 84997-P to A.B.M.A. Part of this research, including the stable isotope analyses, was funded by the ERC Consolidator Grant (SUBSILIENCE ref. 818299), awarded to A.B.M.A. Analysis of dental calculus was carried out in the framework of the ERC Starting Grant (HIDDEN FOODS ref. 639286), awarded to E.C. The authors would like to thank C. García de Castro and B. López for giving information and allowing the sampling of Los Cinchos cave. We are grateful to L. Agudo Perez for her invaluable help during the collagen extraction protocols. The authors wish to acknowledge Dr Lawrence G. Straus (University of New Mexico) for editing the English version of this manuscript. Finally, we also thanks to two anonymous reviewers for their constructive comments that improved the manuscript.
Author information
Authors and Affiliations
Contributions
B.G.R. and A.B.M.A. designed the research. B.G.R. performed the research. M.R.G.M. excavated the site of El Espinoso cave. B.G.R., A.B.M.A., E.C. and A.Z. performed the data analysis. B.G.R., A.B.M.A., E.C., A.Z. and M.R.G.M. wrote and provided comments on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
González-Rabanal, B., Marín-Arroyo, A.B., Cristiani, E. et al. The arrival of millets to the Atlantic coast of northern Iberia. Sci Rep 12, 18589 (2022). https://doi.org/10.1038/s41598-022-23227-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-23227-4
This article is cited by
-
Biomolecular evidence for changing millet reliance in Late Bronze Age central Germany
Scientific Reports (2024)
-
Direct evidence of plant consumption in Neolithic Eastern Sudan from dental calculus analysis
Scientific Reports (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
