
Abstract
Large gaps in reef distribution may hinder the dispersal of marine organisms, interrupting processes vital to the maintenance of biodiversity. Here we show the presence and location of extensive reef habitats on the continental shelf between the Amazon Reef System (ARS) and the Eastern Brazilian Reef System (ERS), two reef complexes off eastern South America. Formations located 20–50 m deep include both biogenic and geogenic structures. The presence of diverse reef assemblages suggests the widespread occurrence of rocky substrates below 50 m. These habitats represent an expansion of both the ARS and ERS and the closure of the only remaining large-scale gap (~ 1000 km) among West Atlantic reef environments. This indicates that the SW Atlantic harbors a single, yet heterogeneous, reef system that stretches for about 4000 km, and thus, represents one of the largest semi-continuous tropical marine ecosystems in the world.
Similar content being viewed by others


Introduction
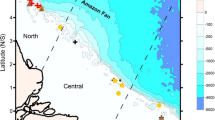
Large clusters of interconnected reefs extending for hundreds to thousands of square kilometers—referred to as reef systems1—are an important feature of tropical coasts. Along the South Atlantic Ocean two main reef systems have been recognized to date: the Eastern Brazilian Reef System (ERS), which comprises shallow-water (< 30 m depth) and mesophotic (30–150 m) reefs along Brazil's eastern continental shelf2,3,4; and the Amazon Reef System (ARS), encompassing mesophotic and rariphotic (30–220 m depth) ecosystems located on the Brazilian Equatorial Margin5,6,7,8,9,10 (Fig. 1). In both cases, reefs are associated with other habitats, such as rhodolith beds, sponge bottoms and seagrass meadows, and the reef systems are more properly a mosaic of interconnected benthic habitats8,10,11. Furthermore, the ARS may provide a bridge between the SW Atlantic and Caribbean, serving as an ecological corridor allowing the exchange of reef species between biogeographic provinces7,12,13,14.

West coast of the Atlantic Ocean showing the countries of Central and South America, large marine ecosystems, major reef systems, and main ocean currents. Areas of reefs and reef systems are out of scale. Shallow reef distribution modified from15 and Amazon reefs based on8. SAC Brazilian semi-arid coast, BC Brazil current, NBC North Brazil current, NECC North equatorial countercurrent, GC Guiana current, CC Caribbean current. Map made using QGIS 3.22 (https://qgis.org/). Country areas retrieved from GADM (https://gadm.org/).
Between the ARS and ERS (from coordinates 5° S 36° W to 2° S 44° W) is the Brazilian semi-arid coast (SAC)16. This area extends for about 1000 km in the SE-NW direction, and has distinct environmental conditions, such as a large interannual variability in rainfall, a predominance of small rivers, and a strong influence of wind on coastal landscapes16. Historically, the continental shelf along the SAC is considered poor in terms of biogenic reef occurrence17,18,19. The shallower sector of this continental shelf is subject to intense wind-driven longshore sediment drifts16, and large biogenic reefs are absent20. In nearshore areas, only fossil biogenic formations or small coral assemblages on intertidal rock outcrops have been reported, which are usually referred to as sandstone reefs20. However, the warm, clear, and oligotrophic waters make the deeper sectors of the SAC continental shelf suitable for the growth of calcifying organisms, and these areas do have a significant modern CaCO3 production21,22,23. Accordingly, the occurrence of biodiverse “coral banks” have been reported for these deeper sectors at least since the 1970s19, and some maps from that decade even mention deep-water “algal reefs”24. However, these reports were rather uncertain and mentioned structures were hardly ever described as reefs by subsequent authors, who mostly reported unconsolidated carbonate substrates such as rhodolith beds or carbonate sand and gravel22,25,26. Therefore the SAC is considered to be depauperate in the most recent reviews on Brazilian reefs3,4,17,18,27,28.
Nevertheless, in the past decade a growing number of studies have reported the occurrence of hard substrates in this region, located from the intertidal zone to tens of meters below the surface29,30,31,32,33,34. Biological and geological characterizations of these substrates are only beginning, but many sustain a diverse reef-associated biota (i.e., reef fishes and epilithic benthos), which play an important role in the economy and ecology of the region32,35,36,37. Furthermore, there are similarities between the SAC hard substrates and the biogenic reefs from the ARS and ERS2,33,34,38.
To date most of the studies along the SAC have been limited and it is not possible to delineate the extent and distribution of these hard-bottom environments, or to put them in a context among other Brazilian reef systems. A better understanding of the distribution of reefal habitats along the SAC is required to improve the uncertainty regarding connectivity between these tropical reef systems. Large gaps between suitable habitats could, for example, hinder the dispersal of marine organisms39,40. In this context, the North Brazil Current (NBC), which flows northwestward off shelf, has surface velocities around 60–100 cm s−141 (Fig. 1). But wind-driven shelf currents are slower, with an average velocity of about 20 cm s−142. At this speed, pelagic larvae would take around 60 days to cross the entire SAC, from the northern ERS tip to eastern ARS tip, which is longer than the planktonic larval duration of many reef species43,44. Accordingly, there seems to be a low demographic connectivity among populations of certain reef species along the equatorial SW Atlantic45. So rocky substrates along the SAC region may act as stepping-stones, aiding biological flux between ERS and ARS45. As such, an assessment of the types and locations of reef habitats along this area could help to clarify such a role.
Here we assess the occurrence and distribution of shallow-water and mesophotic hard-bottom habitats along the SAC continental shelf, by integrating new data with published and unpublished records. This integration of data provides evidence that ARS and ERS are spatially connected by numerous hard-bottom habitats distributed along the continental margin, and thus form a single extensive reef ecosystem extending from the southern Caribbean to the tropical southwestern Atlantic Ocean.
Results
Occurrence and distribution of Hard-bottom environments
There are records of hard substrates along most of the SAC continental margin. Eleven studies have reported a total of 192 sites of geo-referenced hard-bottom habitats, particularly between 20 and 50 m depth, including at least 59 named formations (Supplementary Table S1). Although the geological composition of these hard substrates was not determined in most of these studies, three articles reported CaCO3-cemented sandstones, most likely submerged beachrock, and one described a limestone covered by a well-developed coral assemblage (the Açu reefs), thus confirming the occurrence of biogenic reefs along the SAC (Supplementary Table S1). It is important to note that only primary studies are being reported here, and the nature of both the sandstone and limestone formations have been supported by subsequent studies29,30,31,38,46,47,48,49. Another two studies32,36 mentioned large coral assemblages on unspecified substrates, but only one32 quantified the benthic cover, reporting a dominance by seaweeds and sponges, with live corals and calcareous algae covering together 9.2% of the substrate. In the present study, many of these rocky structures could be identified and mapped by Landsat images and bathymetry. This mapping shows a semi-continuous ridge of consolidated substrates parallel to the coast extending along the entire SAC middle continental shelf (Fig. 2).

Distribution of euphotic and mesophotic hard-bottom habitats along the Brazilian semi-arid coast (Tropical SW Atlantic). Location data for benthic assemblages (including octocoral, scleractinian, and sponge grounds typically with reef fish assemblages) and fishing grounds obtained from the literature, scientific collections, and the monitoring of commercial fisheries (a list of data sources can be found in the supplementary materials S1). Mapped hard substrates depict known rocky-bottom environments, while benthic assemblages and fishing grounds indicate putative rocky substrates. Map made using QGIS 3.22 (https://qgis.org/). Bathymetric data retrieved from CPRM (https://www.cprm.gov.br/) and country area from GADM (https://gadm.org/).
In addition to these confirmed hard-bottom substrates, there are abundant records of epilithic communities along the SAC continental shelf (Supplementary Table S1). The literature and collection surveys resulted in 193 geo-referenced records of benthic assemblages: 99 records of sponge bottoms, with up to 47 species (mean 5.6 ± 7.0); 17 records of coral assemblages (Hexacorallia and Octocorallia), with up to 19 species (mean 3.4 ± 5.0); and 77 records of macroalgae, with up to 8 species (mean 1.7 ± 1.2). Many of these biotic communities co-occur with known rocky substrates, but some diverse assemblages were also found far from currently known hard substrates (Fig. 2), particularly along the continental shelf break where depth and distance from the coast prevent confirmation by satellite images or conventional scuba diving.
Despite the lack of detailed scientific descriptions, the outer SAC continental shelf is intensely fished for reef-fish species (Fig. 2). Three articles report at least 129 geo-referenced fishing grounds used primarily to the capture of reef species such as snappers (Lutjanidae) (Supplementary Table S1). Additionally, during the fishing campaigns monitored in the present study (Supplementary Table S2), 68.1% ± 12.9% of the biomass and 51.3% ± 6.8% of the species richness was composed of primarily reef-associated fish. Snappers (Lutjanus), black groupers (Mycteroperca bonaci), nurse sharks (Ginglymostoma cirratum) and stingrays (Hypanus) were present in more than 90% of the fishing grounds and accounted for 60.8% of the total captured biomass. Their abundance, along with the diverse benthic assemblages, is a strong indication of the occurrence of consolidated reefal substrates along the SAC outer continental shelf.
Ground-truthing
Our ground-truthing of the reported rocky substrates using Scuba diving indicates that these habitats usually rise a few centimeters to a few meters above the surrounding sandy substrate, forming plateaus with positive relief (Fig. 2 and map in Fig. 3). All visited reefal formations sustain a diverse benthic community, usually with a cover of calcifying organisms, such as reef-building scleractinian corals (mostly Montastraea cavernosa and Siderastrea spp.) (Fig. 3a,d) and calcareous algae (Fig. 3f). Reef fish assemblages were also present. Habitats located at greater depths, such as the Manoel Salvador (Fig. 3a) and the Uruaú Channel (Fig. 3d), have significant live coral cover. Whereas shallower formations sustain a noticeable layer of calcareous algae (note, for example, the purplish color of the substrate in Fig. 3b,c). These observations indicate a considerable biological contribution to these structures.

Reefal environments on the continental shelf off the Brazilian semi-arid coast. (a) “Manoel Salvador” reef (~ 30 m deep) covered by Montastraea cavernosa, (b) “Pedrinha” (~ 17 m) with sponges, ascidians, and calcareous algae, (c) reef fish (Holacanthus ciliaris) on the “Cabeço Seco” (~ 13 m) (d) “Canal do Uruaú” mesophotic reef (~ 34 m) with large Siderastrea spp. coral colonies, (e) “Pedra Grande da Majorlandia” (~ 16 m) covered by fleshy algae, and (f) “Pedra Preta” (~ 13 m) covered by fleshy and calcareous algae. Map made using QGIS 3.22 (https://qgis.org/). Bathymetric data retrieved from CPRM (https://www.cprm.gov.br/) and country area from GADM (https://gadm.org/).
Ground-truthing of four areas along the outer continental shelf by ROV also revealed complex habitats (Supplementary Video S1). Apparently, there are rocky ledges that do not rise distinctively above the surrounding sea floor, and often appeared covered by a sediment layer (Fig. 4). Due to this sand cover, the consolidated nature of these habitats is better seen in video than in still images. Scattered rocks, epilithic sponges (e.g., Xestospongia cf. muta, Agelas dispar, Aplysina fulva, Aplysina lacunosa, and Ircinia strobilina) and soft- and hard-bottom dwelling octocorals (such as Neospongodes atlantica) also appeared in these areas (Fig. 4). They also sustained typically reef-associated fish, including Acanthurus sp. (Acanthuridae), Bodianus sp. and Halichoeres sp. (Labridae), Holocentrus adscensionis (Holocentridae), Lutjanus jocu (Lutjanidae), Mycteroperca sp. and Epinephelus sp. (Epinephelidae) (Fig. 4a–d).

Putative rocky substrates on the outer continental shelf off the Brazilian semi-arid coast. Massive sponges and reef-associated fishes (families Acanthuridae, Lutjanidae, and Epinephelidae) observed by Remotely Operated Underwater Vehicle (ROV) suggesting the occurrence of widespread hard substrates at mesophotic depths. (a) Massive sponges (Agelas dispar) and reef fishes (Lutjanus jocu, Mycteroperca sp., Epinephelus sp. and Acanthurus sp.) at 41 m deep, (b) benthic assemblage dominated by octocorals (Neospongodes atlantica) apparently on a rocky substrate at 61 m, (c) a massive sponge (A. dispar), a columnar sponge (Aplysina sp.), and a school of Acanthurus sp. on a low relief rocky substrate covered by a sand veneer at 36 m, (d) Oval sponge (Ircinia strobilina) on a small rocky outcrop surrounded by soft sediments at 41 m. Map made using QGIS 3.22 (https://qgis.org/). Bathymetric data retrieved from CPRM (https://www.cprm.gov.br/) and country area from GADM (https://gadm.org/).
Due to their depths, the nature of these outer shelf habitats cannot be confirmed in the present study, therefore they are referred to as putative rocky substrates hereafter. Despite this uncertainty, confirmed and putative hard-bottom habitats occur relatively close to each other, forming a single reefal seascape that extends about 34,000 km2, and which is clearly aligned with ARS (to the west), possibly providing a bridge to ERS (to the east) along the equatorial margin of the SW Atlantic Ocean (Fig. 2).
Relationships among south Atlantic reef systems
The role of SAC formations as stepping-stones between ARS and ERS is supported by biogeographic patterns. The ordination analyses of two large datasets of marine species50,51 suggests a direct relationship between the ecological dissimilarities (in terms of species composition) and the geographic distances of ARS, SAC, ERS and other South American biogeographic regions (Fig. 5). Moreover, the superimposition of a minimum spanning tree on the ordination analyses52,53 provides an indication that biogeographic links between ARS and ERS consistently pass-through SAC (Fig. 5). This same connection pattern is observed both for the entire dataset, and for the majority (64%) of individual high-rank taxonomic groups in each dataset (Supplementary Figs. S1 and S2).

Ordination analyses (Sammon mapping) of South Atlantic reef systems and biogeographic regions based on the Sørensen dissimilarities among their marine biotas. (a) Analysis on a dataset with 2412 reef species, published by Ref.51, (b) analysis on a dataset with 8375 marine species, published by Ref.50. A minimum spanning tree was superimposed to the ordination graph in order to highlight putative connection pathways among regions52,53. Note that SAC is between ARS and ERS in both cases. ARS Amazon reef system, SAC Brazilian semi-arid coast reef system, ERS Eastern Brazilian reef system.
These connections among ARS, SAC and ERS indicate that these reef systems can be regarded as a single entity across the South Atlantic Ocean, at least at the broader scales of analysis. However, each reef system (or subsystem) has particularities that should not be disregarded in ecological studies (Table 1). Overall, mesophotic shelf-edge reefs seem to occur along most of the Brazilian continental margin, but as formations become shallower, their number, variety and biodiversity increase from north to south.
Discussion
The available data confirm that reefal seascapes associated with hard-bottoms are extensive and semi-continuous along 34.000 km2 of the middle-to-outer continental shelf of the SAC. This discovery of a spatial connection between ARS and ERS implies that this should be regarded as a single and extensive reef system, stretching for about 4000 km along the east coast of South America.
The mapping and physical characterization of this reef system should be improved in future studies. Due to the limited penetration of light into water, analyzes based on satellite data are normally restricted to underwater features located no more than 40 m deep1,29. Therefore, new reef areas along the SAC must be surveyed and validated, especially close to the shelf break. In addition, the shallower (< 10 m) and western sectors of the SAC continental shelf are generally more turbid33 and therefore more difficult to map with remote sensing techniques. The use of high spectral, spatial, and temporal resolution data, in addition to statistical methods and direct underwater assessments (via technical diving, ROVs and BRUVs) are essential to refine our initial estimates of the reef distribution.
The geological composition of SAC hard-bottom formations is uncertain, but, as we have shown here, include sandstones cemented by calcium carbonate (i.e. beachrocks)30 and limestones with well-developed coral communities (i.e., biogenic reefs)46. The only study that quantified the benthic cover on one of these formations32, a rocky plateau at 23 m depth, reported that live corals and calcareous algae covered 9.2% (4.6% each) of the substrate. This number is within the range commonly observed in Brazilian biogenic reefs54,55,56. However, maybe except for the mostly organic Açu reefs, it is generally not known how much these bioconstructors contribute to the framework of the SAC habitats. Therefore, ecosystems that develop on these hard substrates seem to range from deep and low-lying marginal biogenic reefs28,57,58 to non-framework building communities58, including coral grounds and carpets59,60 which can be regarded as an integral part of the reef system along the SAC. At the same time, the prevalence of biogenic versus geogenic formations should be reassessed throughout the entire South American reef system.
Outside the alignment of rocky-bottom habitats between 20 and 40 m deep, numerous localities along the SAC continental shelf support diverse epilithic biotas. For example, trawls made at mesophotic depths revealed a high species richness and biomass of reef benthic organisms, with biodiversity apparently increasing with depth61. The dominant taxon in these surveys was Porifera, which accounted for 90% of the total wet weight, especially due to the species Agelas dispar, Ircinia strobilina, Leucascus sp. and Monanchora arbuscula. Ascidians were the second most abundant group, with the species Stomozoa gigantea alone representing 5% of the total wet weight, followed by Cnidaria (3% of the biomass) and Bryozoa (2% of the biomass)61. Previous studies have shown that such assemblages are associated with the occurrence of geoforms, including reefs and associated habitats62. Additionally, similar biotic communities were found in the deeper zones of the eastern Brazilian continental shelf (along the ERS) and are also linked to the occurrence of mesophotic reef habitats63. Therefore, the presence of these assemblages is a strong indication of the occurrence of yet unmapped hard-bottom environments along the SAC, especially in the outer continental shelf.
Habitats deeper than 40 m explored using ROV data show a relatively flat seascape and hard substrates can only be inferred from the presence of epilithic biotas. Such flat seascapes are not typical of reef environments, which, by definition, have three-dimensional profiles. Nevertheless, ROV data is localized, and sandy and rocky flats are a known feature of reefs along the SAC32,38,46. In fact many formations along the ARS8,10 and ERS64 are flat and largely covered by marine sediment. Consequently, these relatively flat habitats seem to be another important component of the South American reef system, particularly at mesophotic depths10,31,64. Due to their apparent ubiquity across ARS, SAC and ERS, these mesophotic environments may form an extensive ecological corridor along most of the SW Atlantic coast.
Therefore, at a broad spatial scale, the SAC continental shelf harbors an extensive network of different types of consolidated substrates, which form a complex and heterogeneous reef system. Furthermore, much like the ARS and ERS2, this reef system forms a seascape with other habitats, such as rhodolith beds, seagrass and seaweed meadows, marine animal forests, and unconsolidated sediment deposits23,65. Data on the structure and functioning of these ecosystems is scarce, but the few available comparisons, both from within and outside the SAC, indicate important differences in terms of species composition and ecological dynamics11,66. The recognition of these particularities is essential for the adequate management and conservation of these habitats. In parallel, the SAC reef system and neighboring habitats are all equally under the energetic oceanographic conditions across the equatorial Brazilian shelf26,33,67. As such, some of these environments may even be subject to seasonal cycles of burial and exposure, due to the transport of unconsolidated sediments and possible migration of subaqueous sand ripples and dunes68,69. This may explain why some of the hard-bottom habitats are dominated by few stress-tolerant and weedy reef-building corals adapted to sedimentation and moderate turbidity, such as M. cavernosa and Siderastrea spp.2,27,32,36,60.
Due to their large spatial and depth distribution, the hard-bottom formations along the SAC connect ARS and ERS. Together, these reef systems may also link the Caribbean and Brazilian biogeographic provinces. Hence, recognizing the presence and extension of the SAC hard bottoms implies closing the only remaining broad scale gap among Western Atlantic reef habitats. This physical connectivity is mediated by the shelf currents and the NBC, both of which flow primarily from east to west (Fig. 1). However, the NBC also curves back to the east (reflects) between July and December around 6–8° N (as shown in Fig. 1) shedding eddies and rings that may flow eastward, possibly helping to interconnect the reef environments70,71. In this scenario, the SAC hard substrates effectively constitute stepping-stones between ARS and ERS, providing adequate substrates for at least some species, aiding larval dispersion, ontogenetic migration, and small-scale movements of adult animals, which are key processes connecting Western Atlantic reef systems45,72,73. It is noteworthy that both ARS and SAC reefs are typically composed of subsets of the Tropical Atlantic biota, as their environmental conditions seem to be challenging for many coral reef species confined to either the ERS or Caribbean. Nevertheless, the spatial connection provided by the SAC may help to explain the biogeographical affinities observed in previous studies among SW Atlantic reef species, including sponges74, corals75, anemones76, gastropods77, and reef fish78. Furthermore, a multi-taxa study with reef organisms reports a relatively low beta diversity and high nestedness among Brazilian biogeographical provinces, suggesting considerable interconnectivity along the South American reef system51.
The connectivity between ARS and ERS provided by the SAC may affect the resilience of the SW Atlantic reefs against environmental and anthropogenic stressors45. Such human pressures are threatening the biodiversity of both shallow and mesophotic habitats and, thus, urgently require conservation and mitigation actions79. To be successful, such actions need to consider the biogeography and dynamics of metapopulations among SW Atlantic reefs, especially given the wide spatial distribution of such habitats75,78. Further study is urgently required to characterize the ecological dynamics of these tropical reef habitats at multiple spatial and temporal scales in order to sustainably maintain the numerous ecosystem services provided.
Methods
Data on the occurrence of submerged hard-bottom substrates in this equatorial area was compiled from published and unpublished literature. Three types of reef-related data were compiled: (a) location of known hard-bottom habitats—i.e. rocky substrate topologically distinct from the surrounding sediment deposits, (b) occurrence of sponge, scleractinian or octocoral assemblages typical of hard-bottom habitats recorded by trawling or other indirect methods, and (c) coordinates of fishing grounds used to catch reef fishes, whose bottoms had not been characterized, but which may include reefs, marine animal forests, and rhodolith beds. The literature survey included publications in English and Portuguese, but only primary data sources were considered (i.e., only studies mentioning the coordinates of a particular location for the first time were included, and subsequent papers were disregarded). A relevant proportion of the data was only available through gray literature, so we focused on institutional repositories of Brazilian universities (Universidade Federal do Ceará, Universidade Estadual do Ceará, Universidade Federal do Rio Grande do Norte, and Universidade Federal de Pernambuco), governmental agencies (Portal de Periódicos da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), and all-purpose search engines (Google Scholar, Scielo, Science Direct, and Web of Science). Search terms included combinations of “reef”, “beachrock”, “continental shelf”, “Northeast Brazil”, “fisheries”, “fishing grounds”, “corals”, “octocorals”, and “sponges”, with their respective Portuguese translations (a list of the data sources is available as Supplementary Table S1).
This literature survey was complemented by new information produced during this study. Surveys of scientific collections’ databases (Universidade Federal do Ceará, Universidade Federal de Pernambuco, Museu Nacional do Rio de Janeiro/Universidade Federal do Rio de Janeiro, and Smithsonian National Museum of Natural History) were performed to assess the occurrence of sponges and corals (octocorals and scleractinians) along the SAC continental shelf, particularly near the shelf break (Supplementary Table S1). The location of fishing grounds was supplemented by data from a commercial fishing operation involving 35 fishing campaigns using bottom longlines along the SAC outer shelf (Supplementary Table S2). These commercial data were used to assess the occurrence and abundance of reef fish species (identified as such following78) closer to the shelf break. The fishing campaigns targeted exclusively mesopredators and predators, particularly of the Lutjanidae and Epinephelidae families, and elasmobranchs. Therefore, these data do not represent the entire fish diversity on the SAC outer continental shelf. Nevertheless, the eventual occurrence of typical reef-fish assemblages should reflect the distribution of hard-bottom environments63, thus meeting the objectives of the present study.
Based on these published and unpublished data, submerged rocky substrates were mapped using satellite imagery and bathymetric data. Landsat 8 (OLI sensor) and Landsat 5 (TM sensor) digital images were processed (morphological convolution filter, directional filters, mask, and color and band rendering) through bands 1 and 2 (0.45–0.52 µm), and RGB compositions (4-3-2 and 4-2-1) were produced to highlight the occurrence and distribution of hard substrates. Bathymetric contours were created to corroborate and complement satellite information, using bathymetric data from the Brazilian Navy (DHN), retrieved form board pages (no. 500, 600, and 700) and nautical charts (no. 21,700, 21,800, and 21,900). Owing to the clear oceanographic conditions along most of the tropical Brazilian coast, satellite data can be used effectively to map submerged reefs and other hard-bottom habitats29. However, due to limited light penetration in water, such analyses are normally restricted to underwater features located shallower than 40 m deep. Additionally, the shallower (< 10 m) and western sectors of the SAC continental shelf had higher sediment inputs and resuspension29, and thus were more difficult to map with satellite images.
Ground-truthing of compiled data was performed for selected reef formations by Scuba diving and Remotely Operated Underwater Vehicle (ROV) assessments (Supplementary Video S1). In both cases, punctual surveys were performed over the selected formations and underwater images were taken to describe the reef habitats. Due to the large extension of the SAC continental shelf, ground-truthing surveys focused on a representative set of habitats. Due to the rough sea conditions in this area (e.g., intense wind speed and seasonal swell waves), this selection was based primarily on reef accessibility (depth and distance from the coast). Data on the location and nature of hard-bottom habitats, as well as species occurrence records, were integrated into a Geographic Information System (GIS) to evaluate the distribution of the reef habitats along the SAC continental shelf and estimate their positions and distances relative to ARS and ERS.
To investigate possible interconnections among ARS, SAC and ERS we re-analyzed two recently published datasets on the distribution of marine species along the tropical SW Atlantic coast50,51. The first dataset contained literature data on 2412 typically reef species from eight coastal sectors along the Brazilian coast51. The second dataset included records taken from the Ocean Biodiversity Information System (OBIS) for 8375 species (reefal and non-reefal) distributed across six marine ecoregions encompassing the Brazilian, Guianan, and Caribbean coasts50. Both datasets were originally used to identify and describe connections among marine biogeographical regions along their studied coastlines, mainly through cluster analysis and the construction of dendrograms. In the present study, to highlight the interconnections among reef systems, we opted for an ordination analysis. A Sammon mapping80, which is similar to a Multidimensional Scaling putting emphasis on the neighborhood of points, was used to visualize the data. Additionally, we have superimposed a minimum spanning tree, constructed with Prim’s algorithm81, on the Sammon mapping to aid the visualization of connections. Variations of minimum spanning trees have been used in biogeography to investigate putative historical pathways of connection among marine areas52,53. Considering the mostly unidirectional flow of oceanic and shelf currents along ERS, SAC and ARS the minimum spanning tree should also be able to reflect the interconnection among reef systems in a parsimonious way. The ordination analyses and the algorithm for the minimum spanning trees were executed in R 4.1.2, with packages vegan and MASS.
Data availability
All data needed to evaluate the conclusions of this paper are presented in the paper and/or the Supplementary Materials. A list of geographic coordinates (lat/lon), additional images, and video shootings of the studied habitats are available from the corresponding authors upon reasonable request.
References
Roelfsema, C., Phinn, S., Jupiter, S., Comley, J. & Albert, S. Mapping coral reefs at reef to reef-system scales, 10s–1000s km2, using object-based image analysis. Int. J. Remote Sens. 34, 6367–6388 (2013).
Soares, M. O., Tavares, T. C. L. & Carneiro, P. Mesophotic ecosystems: Distribution, impacts and conservation in the South Atlantic. Divers. Distrib. 25(2), 255–268 (2019).
Leão, Z. M. A. N. et al. Brazilian coral reefs in a period of global change: A synthesis. Braz. J. Oceanogr. 64, 97–116 (2016).
Leão, Z. M. A. N., Kikuchi, R. K. P. & Oliveira, M. D. M. The coral reef province of Brazil. World Seas: An Environmental Evaluation Volume I: Europe, the Americas and West Africa vol. 1 (Elsevier Ltd., 2018).
Collette, B. B. & Rützler, K. Reef fishes over sponge bottoms off the mouth of the Amazon River. in Proceedings of Third International Coral Reef Symposium (ed. Taylor, D. L.) vol. 1 305–310 (Rosenstiel School of Marine and Atmospheric Science, 1977).
Cordeiro, R. T. S., Neves, B. M., Rosa-Filho, J. S. & Pérez, C. D. Mesophotic coral ecosystems occur offshore and north of the Amazon River. Bull. Mar. Sci. 91, 491–510 (2015).
Moura, R. L. et al. An extensive reef system at the Amazon River mouth. Sci. Adv. 2, e1501252 (2016).
Francini-Filho, R. B. et al. Perspectives on the Great Amazon Reef: Extension, biodiversity, and threats. Front Mar Sci 5, 1–5 (2018).
de Mahiques, M. M. et al. Insights on the evolution of the living Great Amazon Reef System, equatorial West Atlantic. Sci. Rep. 9, 1–8 (2019).
Vale, N. F. et al. Distribution, morphology and composition of mesophotic ‘reefs’ on the Amazon Continental Margin. Mar. Geol. 447, 106779 (2022).
Moura, R. L. et al. Tropical rhodolith beds are a major and belittled reef fish habitat. Sci. Rep. 11, 1–10 (2021).
Rocha, L. A. Patterns of distribution and processes of speciation in Brazilian reef fishes. J. Biogeogr. 30, 1161–1171 (2003).
Floeter, S. R. et al. Atlantic reef fish biogeography and evolution. J. Biogeogr. 31, 22–47 (2008).
Vale, N. F. et al. Structure and composition of rhodoliths from the Amazon River mouth, Brazil. J. S. Am. Earth Sci. 84, 149–159 (2018).
IMaRS/USF, IRD, UNEP/WCMC, The WorldFish Center & WRI. Global Coral Reefs composite dataset compiled from multiple sources for use in the Reefs at Risk Revisited project incorporating products from the Millennium Coral Reef Mapping Project. Preprint at (2011).
Soares, M. O. et al. Challenges and perspectives for the Brazilian semi-arid coast under global environmental changes. Perspect. Ecol. Conserv. 19, 267–278 (2021).
Castro, C. B. & Pires, D. O. Brazilian coral reefs: What we already know and what is still missing. Bull. Mar. Sci. 69, 357–371 (2001).
Leão, Z., Kikuchi, R. & Testa, V. Corals and coral reefs of Brazil. in Latin American Coral Reefs (ed. Cortés, J.) 9–52 (Elsevier Science Inc., 2003). https://doi.org/10.1016/B978-044451388-5/50003-5.
Laborel-Deguen, F., Castro, C. B., Nunes, F. D. & Pires, D. O. Recifes brasileiros: o legado de Laborel. (Museu Nacional, 2019).
Carneiro, P. et al. Marine hardbottom environments in the beaches of Ceará state, equatorial coast of Brazil. Arquivos de Ciências do Mar 54, 120–153 (2021).
Carneiro, P. B. M. et al. Structure, growth and CaCO3 production in a shallow rhodolith bed from a highly energetic siliciclastic-carbonate coast in the equatorial SW Atlantic Ocean. Mar. Environ. Res. 166, 105280 (2021).
Testa, V., Bosence, D. W. J. & Universita, C. Physical and biological controls on the formation of carbonate and siliciclastic bedforms on the north-east Brazilian shelf. Sedimentology 46, 279–301 (1999).
Carneiro, P. & Morais, J. O. de. Carbonate sediment production in the equatorial continental shelf of South America: Quantifying Halimeda incrassata (Chlorophyta) contributions. J. S. Am. Earth Sci. 72, 1–6 (2016).
Milliman, J. D. Role of Calcareous Algae in Atlantic Continental Margin Sedimentation. in Fossil algae: recent results and developments (ed. Flügel, E.) 232–247 (Springer, 1977). https://doi.org/10.1007/978-3-642-66516-5_26.
Knoppers, B., Ekau, W. & Figueiredo, A. G. The coast and shelf of east and northeast Brazil and material transport. Geo-Mar. Lett. 19, 171–178 (1999).
Vital, H. The north and northeast Brazilian tropical shelves. in Continental shelves of the world: their evolution during the lasta glacio-eustatic cycle (eds. Chiocci, F. L. & Chivas, A. R.) 35–46 (Geological Society, 2014).
Soares, M. de O. et al. Brazilian marine animal forests: A new world to discover in the southwestern Atlantic. Mar. Anim. For. 1–38. https://doi.org/10.1007/978-3-319-17001-5_51-1 (2016).
Soares, M. O. et al. Impacts of a changing environment on marginal coral reefs in the Tropical Southwestern Atlantic Ocean. Coast. Manag. 210, 105692 (2021).
Santos, C. L. A., Vital, H., Amaro, V. E. & de Kikuchi, R. K. P. Mapping of the submerged reefs in the coast of the Rio Grande do Norte, near Brazil: Macau to Maracajau. Revista Brasileira de Geofisica 25, 27–36 (2007).
Neto, I. C., Córdoba, V. C. & Vital, H. Morfologia, microfaciologia e diagênese de beachrocks costa-afora adjacentes à costa norte do Rio Grande do Norte, brasil. Geociências 32, 471–490 (2013).
Gomes, M. P. et al. The investigation of a mixed carbonate-siliciclastic shelf, NE Brazil: Side-scan sonar imagery, underwater photography, and surface-sediment data. Ital. J. Geosci. 134, 9–22 (2015).
Soares, M. O., Rossi, S., Martins, F. A. S. & Carneiro, P. The forgotten reefs: Benthic assemblage coverage on a sandstone reef (Tropical South-western Atlantic). J. Mar. Biol. Assoc. U.K. 97(8), 1585–1592. https://doi.org/10.1017/S0025315416000965 (2017).
Morais, J. O., Ximenes Neto, A. R., Pessoa, P. R. S. & Souza, L. P. Morphological and sedimentary patterns of a semi-arid shelf, Northeast Brazil. Geo-Ma. Lett. 40, 835–842. https://doi.org/10.1007/s00367-019-00587-x (2019).
Cordeiro, R. T., Neves, B. M., Kitahara, M. v., Arantes, R. C. & Perez, C. D. First assessment on Southwestern Atlantic equatorial deep-sea coral communities. Deep-Sea Res. Part I Oceanogr. Res. Papers 163, 103344 (2020).
Freitas, J. E. P. & Lotufo, T. M. C. Reef fish assemblage and zoogeographic affinities of a scarcely known region of the western equatorial Atlantic. J. Mar. Biol. Assoc. U.K. 95, 623–633 (2015).
Soares, M. O., Davis, M., Paiva, C. C. de & Carneiro, P. Mesophotic ecosystems: Coral and fish assemblages in a tropical marginal reef (northeastern Brazil). Mar. Biodivers. 1–6 (2016). https://doi.org/10.1007/s12526-016-0615-x.
Carneiro, P. B. M., Sátiro, I., COE, C. M. & Mendonça, K. V. Valoração ambiental do Parque Estadual Marinho da Pedra da Risca do Meio, Ceará, Brasil. Arquivo de Ciências do Mar 50, 25–41 (2017).
Gomes, M. P., Vital, H. & Droxler, A. W. Terraces, reefs, and valleys along the Brazil northeast outer shelf: Deglacial sea-level archives?. Geo-Mar. Lett. 40, 699–711. https://doi.org/10.1007/s00367-020-00666-4 (2020).
Cowen, R. K. & Sponaugle, S. Larval dispersal and marine population connectivity. Ann. Rev. Mar. Sci. 1, 443–466 (2009).
Raitsos, D. E. et al. Sensing coral reef connectivity pathways from space. Sci. Rep. 7, 1–10 (2017).
Silveira, I. C. A., Miranda, L. B. & Brown, W. S. On the origins of the North Brazil Current. J. Geophys. Res. 99, 22501–22512 (1994).
Dias, F. J. da S., Castro, B. M. & Lacerda, L. D. Tidal and low-frequency currents off the Jaguaribe River estuary (4° S, 37° 4′ W), northeastern Brazil. Ocean Dynamics 68, 967–985 (2018).
Wellington, G. M. & Victor, B. C. Planktonic larval duration of one hundred species of Pacific and Atlantic damselfishes (Pomacentridae). Mar. Biol. 101, 557–567 (1989).
Victor, B. C. Duration of the planktonic larval stage of one hundred species of Pacific and Atlantic wrasses (family Labridae). Mar. Biol. 90, 317–326 (1986).
Endo, C. A. K., Gherardi, D. F. M., Pezzi, L. P. & Lima, L. N. Low connectivity compromises the conservation of reef fishes by marine protected areas in the tropical South Atlantic. Sci. Rep. 9, 1–11 (2019).
Gomes, M. P. et al. Nature and condition of outer shelf habitats on the drowned Açu Reef, Northeast Brazil. in Seafloor Geomorphology as Benthic Habitat 571–585 (Elsevier, 2020). https://doi.org/10.1016/b978-0-12-814960-7.00034-8.
Neto, I. C., Córdoba, V. C. & Vital, H. Petrografia de beachrock em zona costa afora adjacente ao litoral norte do Rio Grande do Norte Brasil. Quat. Environ. Geosci. 2, 12–18 (2010).
Gomes, M. P., Vital, H., Bezerra, F. H. R., de Castro, D. L. & Macedo, J. W. de P. The interplay between structural inheritance and morphology in the Equatorial Continental Shelf of Brazil. Mar. Geol. 355, 150–161 (2014).
Rovira, D. P. T., Gomes, M. P. & Longo, G. O. Underwater valley at the continental shelf structures benthic and fish assemblages of biogenic reefs. Estuar. Coast. Shelf Sci. 224, 245–252 (2019).
Tosetto, E. G., Bertrand, A., Neumann-Leitão, S. & Nogueira Júnior, M. The Amazon River plume, a barrier to animal dispersal in the Western Tropical Atlantic. Sci. Rep. 12, 537 (2022).
Cord, I. et al. Brazilian marine biogeography: A multi-taxa approach for outlining sectorization. Mar. Biol. 169, 61 (2022).
Moalic, Y. et al. Biogeography revisited with network theory: Retracing the history of hydrothermal vent communities. Syst. Biol. 61, 127 (2012).
López-Pérez, A. et al. The coral communities of the Islas Marias archipelago, Mexico: Structure and biogeographic relevance to the Eastern Pacific. Mar. Ecol. 37, 679–690 (2016).
Cordeiro, C. A. M. M. et al. Conservation status of the southernmost reef of the Amazon Reef System: The Parcel de Manuel Luís. Coral Reefs 40, 165–185 (2021).
Segal, B. & Castro, C. B. Coral community structure and sedimentation at different distances from the coast of the Abrolhos Bank Brazil. Braz. J. Oceanogr. 59, 119–129 (2011).
Aued, A. W. et al. Large-scale patterns of benthic marine communities in the Brazilian Province. PLoS ONE 13, e0198452 (2018).
Soares, M. O. et al. Marginal Reefs in the Anthropocene: They Are Not Noah’s Ark. in Perspectives on the Marine Animal Forests of the World (eds. Rossi, S. & Bramanti, L.) 87–128 (Springer International Publishing, 2020). https://doi.org/10.1007/978-3-030-57054-5_4.
Perry, C. T. & Larcombe, P. Marginal and non-reef-building coral environments. Coral Reefs 22, 427–432 (2003).
Riegl, B. & Piller, W. E. Coral frameworks revisited - reefs and coral carpets in the northern Red Sea. Coral Reefs 18, 241–253 (1999).
Rodríguez-Martínez, R. E., Jordán-Garza, A. G., Maldonado, M. A. & Blanchon, P. Controls on coral-ground development along the Northern Mesoamerican Reef Tract. PLoS ONE 6, e28461 (2011).
Lotufo, T. M. et al. Sessile epifauna of Ceará´s shelf - high dominance of sponges. in 7th International Sponge Symposium - Biodiversity, Innovation, Sustainability 123–123 (Museu Nacional - UFRJ, 2006).
Fonseca, V. P., Pennino, M. G., de Nóbrega, M. F., Oliveira, J. E. L. & de Figueiredo Mendes, L. Identifying fish diversity hot-spots in data-poor situations. Mar. Environ. Res. 129, 365–373 (2017).
Olavo, G., Costa, P. A. S., Martins, A. S. & Ferreira, B. P. Shelf-edge reefs as priority areas for conservation of reef fish diversity in the tropical Atlantic. Aquat. Conserv. Mar. Freshwat. Ecosyst. 21, 199–209 (2011).
Eduardo, L. N. et al. Identifying key habitat and spatial patterns of fish biodiversity in the tropical Brazilian continental shelf. Cont. Shelf Res. 166, 108–118 (2018).
Carneiro, P. B. de M. et al. Structure, growth and CaCO3 production in a shallow rhodolith bed from a highly energetic siliciclastic-carbonate coast in the equatorial SW Atlantic Ocean. Mar. Environ. Res. 166, 105280 (2021).
Costa, A. C. P., Garcia, T. M., Paiva, B. P., Ximenes Neto, A. R. & Soares, M. de O. Seagrass and rhodolith beds are important seascapes for the development of fish eggs and larvae in tropical coastal areas. Mar. Environ. Res. 161, 105064 (2020).
Testa, V. & Bosence, D. W. J. Carbonate-siliciclastic sedimentation on a high-energy, ocean-facing, tropical ramp, NE Brazil. in Carbonate Ramps (eds. Wright, V. P. & Burchette, T. P.) 55–71 (The Geological Society, 1998).
Ximenes Neto, A. R., de Morais, J. O. & Ciarlini, C. Modern and relict sedimentary systems of the semi-arid continental shelf in NE Brazil. J. S. Am. Earth Sci. 84, 56–68 (2018).
Ximenes Neto, A. R., Morais, J. O. de, Paula, L. F. S. de & Pinheiro, L. de S. Transgressive deposits and morphological patterns in the equatorial Atlantic shallow shelf (Northeast Brazil). Region. Stud. Mar. Sci. 24, 212–224 (2018).
Sponaugle, S., Lee, T., Kourafalou, V. & Pinkard, D. Florida Current frontal eddies and the settlement of coral reef fishes. Limnol. Oceanogr. 50, 1033–1048 (2005).
Cruz, R. et al. Large-scale oceanic circulation and larval recruitment of the spiny lobster Panulirus argus (Latreille, 1804). Crustaceana 88, 298–323 (2015).
Luiz, O. J. et al. Ecological traits influencing range expansion across large oceanic dispersal barriers: Insights from tropical Atlantic reef fishes. Proc. R. Soc. B Biol. Sci. 279, 1033–1040 (2012).
Romero-Torres, M., Treml, E. A., Blanchon, P., Acosta, A. & Paz-García, D. A. The Eastern Tropical Pacific coral population connectivity and the role of the Eastern Pacific Barrier. Sci. Rep. 8, 1–13 (2018).
Leal, C. v. et al. Integrative taxonomy of Amazon Reefs’ Arenosclera spp.: A new clade in the Haplosclerida (Demospongiae). Front. Mar. Sci. 4, 291 (2017).
Peluso, L. et al. Contemporary and historical oceanographic processes explain genetic connectivity in a Southwestern Atlantic coral. Sci. Rep. 8, 1–12 (2018).
Targino, A. K. G. & Gomes, P. B. Distribution of sea anemones in the Southwest Atlantic: Biogeographical patterns and environmental drivers. Mar. Biodivers. 50, 1–17 (2020).
Barroso, C. X., Lotufo, T. M. da C. & Matthews-Cascon, H. Biogeography of Brazilian prosobranch gastropods and their Atlantic relationships. J. Biogeogr. 43, 2477–2488 (2016).
Pinheiro, H. T. et al. South-western Atlantic reef fishes: Zoogeographical patterns and ecological drivers reveal a secondary biodiversity centre in the Atlantic Ocean. Divers. Distrib. 24, 951–965 (2018).
Medeiros, A. P. M. et al. Deep reefs are not refugium for shallow-water fish communities in the southwestern Atlantic. Ecol. Evol. 11, 4413–4427 (2021).
Sammon, J. W. A nonlinear mapping for data structure analysis. IEEE Trans. Comput. C–18, 401–409 (1969).
Prim, R. C. Shortest connection networks and some generalizations. Bell Syst. Tech. J. 36, 1389–1401 (1957).
Acknowledgements
The authors would like to thank Dr. Eduardo Hajdu and Dr. Ulisses Pinheiro for their help in compiling data on Porifera occurrence.
Funding
Open Access funding enabled and organized by Projekt DEAL. Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). Alexander von Humboldt Foundation (AVH). Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP). Fundação de Amparo à Pesquisa do Estado do Piauí (FAPEPI).
Author information
Authors and Affiliations
Contributions
P.B.M.C. and M.O.S. conceived the study, performed the literature review and co-wrote the first draft of the paper. A.R.X.N., J.O.M., L.S.P., L.H.U.M. and M.D.A.B. acquired primary data on rocky substrates (i.e., sediment data, remote sensing and underwater images). B.J.Q., V.V.F., J.S.N., S.S., R.T.S.C., J.T.A. and T.M.G., acquired secondary data on rocky substrates (i.e., fisheries and scientific collection records). C.X.B., J.E.P.F., C.E.P.T, S.B., T.B.S., S.R., T.M.C.L., H.M.C. and C.V.F. interpreted the data and revised the text. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Supplementary Information 4.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Carneiro, P.B.M., Ximenes Neto, A.R., Jucá-Queiroz, B. et al. Interconnected marine habitats form a single continental-scale reef system in South America. Sci Rep 12, 17359 (2022). https://doi.org/10.1038/s41598-022-21341-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-21341-x
This article is cited by
-
Multiple nets better explain the diversity of marine fish larvae in equatorial environments
Marine Biodiversity (2024)
-
Genetic diversity patterns of lionfish in the Southwestern Atlantic Ocean reveal a rapidly expanding stepping-stone bioinvasion process
Scientific Reports (2023)
-
Hydroids (Cnidaria, Hydrozoa) from the Northern and North-eastern coast of Brazil: addressing knowledge gaps in neglected regions
Marine Biodiversity (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
