
Abstract
An in vitro Hohenheim gas test was conducted to analyze the fermentation end-products from 17 cultivars of eight polyphenol containing forage species. The polyphenol composition and proanthocyanidin (PA) structural features of all the cultivars were analyzed with UPLC-MS/MS in leaves of vegetative or generative plants. The samples were incubated with and without polyethylene glycol (PEG, a tannin-binding agent) to separate the tannin-effect on methane (CH4, ml/200 mg DM) production from that of forage quality. Sulla and big trefoil, two particularly PA rich species, were found to have the highest CH4 reduction potential of up to 47% when compared to the samples without PEG. However, concomitant reduction in gas production (GP, ml/200 mg DM) of up to 44% was also observed. An increase in both GP and CH4 production under PEG treatments, confirms the role of tannins in CH4 reduction. Moreover, PA structural features and concentration were found to be an important source of variation for CH4 production from PA containing species. Despite having low polyphenol concentrations, chicory and plantain were found to reduce CH4 production without reducing GP. Additionally, interspecies variability was found to be higher than intraspecies variability, and these results were consistent across growth stages, indicating the findings’ representativeness.
Similar content being viewed by others

Introduction
One of the main environmental issues from ruminant livestock production are enteric methane (CH4) emissions, which contribute to up to 40% of the total livestock sector emissions worldwide1. The radiative forcing capacity of CH4 is 63 times higher than that of CO2 over the first 20 years, and 27 times higher over the 100-year horizon. That makes CH4 the second most important greenhouse gas after CO2, regarding anthropogenic climate change2. On the other hand, the very shorter atmospheric lifetime of CH4 (around 12 years), results in a much higher potential to reduce the radiative forcing in the short term, compared to CO2 and N2O. Thus, it can provide a valuable opportunity to limit the extent of anthropogenic climate change until long term solutions are in place3. Although, CH4 emissions from ruminants cannot be completely eliminated without losing the ability of ruminants to digest cellulose, their reduction is also desirable from an economic point of view, as energy losses in the form of CH4 can amount up to approximately 2 to 12% of gross energy intake in ruminants4.
The inclusion of forages rich in plant specialized metabolites (PSMs) (e.g. tannins and saponins) in the animal diet can potentially reduce CH4 emissions in the ruminants5. Particularly tannin-rich forages (TRFs) are considered a promising solution due to their multifunctionality and their beneficial effects on animal health and productivity. Tannin-rich forages can exert beneficial and anti-nutritional effects on ruminants, however their potential depends on tannin type and concentration supplied in the feed6. Generally, tannins are known to bind with feed constituents such as protein, fibre, and carbohydrates. They can potentially improve protein utilization of the forages by making protein-tannin complexes in the rumen (pH = 6–7), and transporting them to abomasum (pH < 3.5) and small intestine (pH < 7) where they will eventually dissociate due to post ruminal pH shifts and release proteins. This makes the protein available for gastric or pancreatic digestion, which can reduce feed energy losses in form of CH4 emissions7. This is especially relevant for ruminant production systems in temperate regions where despite having high protein content, the conventional forages, such as lucerne (Medicago sativa L.) and white clover (Trifolium repens L.), have been associated with bloating and inefficient N utilisation8,9. The high extent of protein degradation in the rumen increases high urinary N excretion, which could potentially lead to increased ammonia (NH3) volatilization from the cattle manure and concomitantly negatively affect the environment6.
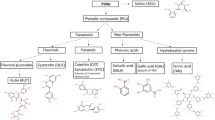
Tannins can be broadly classified as proanthocyanidins (PAs; syn. condensed tannins), hydrolysable tannins (HTs) and phlorotannins, and the latter are mainly found in marine organisms. Hydrolysable tannins can be further subdivided into gallic acid derivatives, ellagitannins and gallotannins10, while PAs exist in the form of polymers and oligomers of flavan-3-ol subunits, most often catechin or epicatechin together known as procyanidins (PCs), and gallocatechin and epigallocatechin (prodelphinidins, PD)11. Procyanidins and PDs differ from each other in terms of their hydroxylation pattern of their flavan-3-ol B-ring, with PD subunits containing three hydroxyl groups and PC subunits containing two hydroxyl groups in the B-ring6. The structural traits are important, particularly the percentage of PD subunits from total PAs (PD%) and the polymer size have been shown to effect the mode and extent of the desired bioactive properties from PAs12. Both forages rich in PAs, such as sainfoin (Onobrychis viciifolia L.) and birdsfoot trefoil (Lotus corniculatus L.), as well as forages rich in HTs, such as salad burnet (Sanguisorba minor), have emerged as promising species that can improve N utilisation in the ruminants while simultaneously curtailing CH4 emissions13,14. When supplied in low to moderate concentrations, tannins (both PAs and HTs) in forages can potentially improve animal performance, reduce NH3 volatilization from manure by redirecting N excretion from urine to feces, lower CH4 emissions and decrease bloating incidences in cattle7,15. Birdsfoot trefoil, one of the promising TRFs, has been found to increase milk yield and milk protein content compared to perennial ryegrass in Friesian cows, and improved wool growth in sheep16.
Several in vivo and in vitro studies have analyzed the antimethanogenic potential of TRFs; however, a high variability in the results from these studies makes it difficult to harness their benefit in the livestock systems17. This is likely due to the abovementioned high structural diversity of tannins, which makes it difficult to assess their bioactivity, and understand their effect on animal physiology and nutrition. It has been widely recognized that PA structural characteristics are important determinants of bioactivity, however, in the majority of animal nutrition studies the structural characteristics or the composition of PAs and HTs are rarely considered. These structural characteristics are known to vary not only across different plant species but also within their cultivars and have been found to be affected by plant’s phenological stage18. However, the variation within the cultivar for each species was found to be less than the variation across the species19. To date, no study has identified the antimethanogenic effect of several cultivars each from multiple forage species grown under identical environmental conditions and across multiple harvests to determine not only the interspecies variability but also the reproducibility of these findings within species and across harvests, which is ultimately a main prerequisite for their use in practice.
Consequently, the study aimed to assess the in vitro CH4 reduction potential while minimizing the aforementioned variations and accounting the often-neglected PA structural features. To our knowledge, this is the first study to analyze the CH4 mitigation potential of eight agronomically important temperate forage plants grown under identical environmental conditions and harvested at the same phenological stage (Fig. 1). Main objectives of this study were, (i) to analyze the inter- and intraspecies variation of the CH4 mitigation potential of multiple forage species, (ii) to assess the tannin-effect on the CH4 reduction from the TRFs, and (iii) determine the influence of PA structural features, such as PD% and PA polymer size, on the CH4 production.

A greenhouse experiment was conducted with 17 cultivars from eight forage species. The cultivars were harvested twice at different phenological stages (Flowering and vegetative stage). Harvested plants were analysed for their polyphenolic composition, PA structural features, and antimethanogenic potential.
Results
Forage quality and tannin composition
The forage chemical composition of the leaves varied extensively across the species. While general trends existed, and herbs had lower CP and higher NDF concentrations compared to legumes, there was a large variation across different functional groups. In general, highest CP concentration was found in big trefoil and birdsfoot trefoil cultivars (224–272 g/kg DM) when plants were harvested at the flowering stage (first harvest) (Table 1), while sainfoin cultivars had the lowest CP concentrations (157–180 g/kg DM) of all legumes and generally the highest NDF concentrations (518–548 g/kg DM). In the vegetative stage at the second harvest, however, two sainfoin cultivars (CPI 63750 and Esky) had the highest CP concentrations of all species. However, generally the differences in CP concentration decreased with the vegetative stage and no clear differences were observed between legumes and herbs at this stage. Additionally, differences between cultivars of same species were mainly observed for CP concentration for the species ribwort plantain, salad burnet and chicory in the first harvest.
Significant differences among cultivars were observed for total polyphenol (TP), total flavonols (TF, sum of myricetin, quercetin and kaempferol concentration), and total tannin (TT) concentration (P < 0.001). The lowest TP concentration was found in chicory, ribwort plantain, and lucerne. Lucerne was found to be void of polyphenols, and was used as tannin-free negative control in the experiment. Highest TF concentrations were found in sainfoin cultivars with a range from 1.3 to 3.8 g/kg DM, while ribwort plantain cultivars generally contained no flavonols19. Salad burnet was the only HT containing forage in our study and had the highest TP concentrations ranging from 42 to 50 g/kg DM. Proanthocyanidin containing species were sainfoin, sulla, big trefoil and birdsfoot trefoil with concentrations ranging from 1 to 25 g/kg DM across both harvests. Proanthocyanidin structural features, PD% and mean degree of polymerisation (mDP), generally differed significantly (P < 0.001) across species, while differences across the cultivars of a single species were restricted to birdsfoot trefoil (Table 2)19. In birdsfoot trefoil, Rocco was consistently found to have significantly higher mDP values (12–14) and PD% (48–52%) than Bull (mDP: 9–10, PD%: 30–33.3%) and Lotar (mDP: 9–11, PD%: 33–39%)19.
Differences in gas and methane production were observed across species and their cultivars
The in vitro gas production (GP) from the forages differed significantly across the cultivars (P < 0.001) with the GP ranging from 25 to 54 ml/200 mg DM across both harvests (Fig. 2, −PEG). At the species level, chicory had the highest GP (44 to 54 ml/200 mg DM), while sulla had the lowest GP (25 to 32 ml/200 mg DM). The variability within the cultivars of any species for GP was only observed among the cultivars of sainfoin and ribwort plantain, with the plantain cultivars showing the largest differences, as Svatojansky had produced 28% less gas compared to PLA60 in the first harvest. The effect of harvest (P = 0.63) on GP was not significant; however, a significant interaction effect of cultivar and harvest (P < 0.001) on GP was observed. This was apparent for ribwort plantain, where the cultivar Svatojansky produced 40% more gas in the second harvest compared to the first harvest and as a result, its GP then exceeded that of the cultivar PLA60. For salad burnet on the other hand, the GP was reduced in the second harvest compared to the first harvest by approx. 26%, yet this difference was homogeneous across both the cultivars.

Gas production from 17 cultivars of 8 species under different treatments for two harvests. −PEG indicates the samples incubated without polyethylene glycol (PEG) and + PEG refers to the samples incubated with PEG. Different uppercase letters represent significant differences (P < 0.05) between the cultivars for each species, and different lowercase letters represent the significant differences within the treatments for each cultivar.
The CH4 production was also influenced by the plant species and their cultivars (P < 0.001) and the effect of harvest (P = 0.06) was not significant (Fig. 3, −PEG treatment). The lowest CH4 production was observed for the ribwort plantain cultivar Svatojansky (5.6 ml/200 mg DM) and the sulla cultivar Sudda (5.6 ml/200 mg DM), followed by the big trefoil cultivar Lot 29 (6.8 ml/200 mg DM). Despite containing PAs, CH4 production from birdsfoot trefoil (9–11 ml/200 mg DM) was the highest and was similar to lucerne (10.8 ml/200 mg DM). In general, the CH4 production from the cultivars in the first harvest was lower compared to the second harvest and there was a significant cultivar-harvest interaction (P < 0.001). Within species variation in CH4 production was observed in the cultivars from birdsfoot trefoil, sainfoin and ribwort plantain. As seen for GP, the variability in CH4 production between ribwort plantain cultivars was larger in the first harvest, with Svatojansky producing 35% less CH4 than PLA60. In the second harvest, the variation between the cultivars decreased and Svatojansky produced 5% more CH4 than PLA60. In birdsfoot trefoil, Rocco generally produced lowest CH4 and had on average 10% lower CH4 formation compared to other birdsfoot trefoil cultivars (Bull and Lotar). Largely, GP and CH4 production from the cultivars were stable across the harvests.

Methane production from 17 cultivars of 8 species under different treatments for two harvests. −PEG indicates the samples incubated without polyethylene glycol (PEG) and + PEG refers to the samples incubated with PEG. Different uppercase letters represent significant differences (P < 0.05) between the cultivars for each species, and different lowercase letters represent the significant differences within the treatments for each cultivar.
The cultivars were observed to differ significantly in terms of CH4 percentage in total gas ( MP, (P < 0.001)). The MP ranged from 17 to 25% in the first harvest and from 20 to 26% in the second harvest. The effect of harvest was not significant (P = 0.06) on MP, however the interaction between harvest and cultivar was found to be significant (P < 0.001). Within species variability in MP was again limited to cultivars from sainfoin, ribwort plantain, and birdsfoot trefoil (Fig. 4; −PEG treatment). Generally, the MP from tannin containing forages (20–26%) was found to be higher compared to ribwort plantain and chicory (17–21%). The lowest CH4 produced per unit total gas production was found in ribwort plantain followed by chicory.

Methane percentage in total gas from 17 cultivars of 8 species under different treatments for two harvests. −PEG indicates the samples incubated without polyethylene glycol (PEG) and + PEG refers to the samples incubated with PEG. Different uppercase letters represent significant differences (P < 0.05) between the cultivars for each species, and different lowercase letters represent the significant differences within the treatments for each cultivar.
In general, there was a concomitant reduction of GP and CH4 production from all the tannin containing species when compared to lucerne. It was observed that the decline in CH4 production (7–47%) with TRFs (sulla, big trefoil, salad burnet and sainfoin), was higher than the GP reduction (5–43%) when compared to lucerne (Supplementary Table S2). Contrary to that, chicory cultivars were found to increase GP (7–21%) and reduce CH4 production (5–25%) with respect to lucerne in both the harvests.
Addition of PEG, significantly increased gas and methane production from tannin rich forages
The addition of PEG resulted in increments of GP (P < 0.001) and decrements in CH4 production (P < 0.001) during the in vitro fermentation (Figs. 2 and 3, + PEG). These changes were, however, limited to the TRFs. Ribwort plantain and chicory cultivars did not change their fermentation patterns with the addition of PEG. As a result, significant interaction effect of cultivar and PEG treatment existed for GP (P < 0.001) and CH4 production (P < 0.001). The GP in the + PEG treatment was found to range from 33 to 53 ml/200 mg DM, while the CH4 ranged from 5 to 12 ml/200 mg DM. In the first harvest, ribwort plantain cultivar Svatojansky produced the lowest CH4 (5 ml/200 mg DM) under the + PEG treatment, the salad burnet cultivar, Sang 10 had the lowest CH4 production (8.9 ml/200 mg DM) under the + PEG treatment in the second harvest. The highest change in GP and CH4 production caused by the PEG addition was found with sulla cultivar, Sudda, where the + PEG treatment increased GP and CH4 by 46 and 45 percent respectively, compared to the control (−PEG). Conversely, the lowest effect from the PEG addition in tannin containing forages, was observed on birdsfoot trefoil cultivars. While the effect of PEG treatment on the MP was significant (P < 0.001, Fig. 4), there was no significant interaction between treatment and harvest for the MP. There was, however, a significant effect of three-way interaction between cultivar, treatment, and harvest (P < 0.001) observed for MP. Still, the major variations in MP were observed for birdsfoot trefoil and sainfoin in the first harvest, while in the second harvest, a significant treatment effect was observed on the MP from birdsfoot trefoil, big trefoil, salad burnet and plantain.
Covariates generally explain the variability of fermentation parameters
A model comparison was done by substituting fixed factors, cultivar and harvest, sequentially with measured covariates, to determine whether the measured covariates explain the formation of fermentation end products (CH4, GP and MP) equally well, compared to the fixed factors. Yet, replacing the fixed factors with the covariates was not able to improve the quality of the model in terms of the AIC value for neither CH4 (AIC = 312) nor GP (AIC = 1749; Table 3). Similarly, for MP the AIC was -646. The marginal R2 for CH4 and GP of the final models were generally high with 0.81 and 0.68, respectively. This was despite them being lower than the respective marginal R2 values of the reference model, with 0.89 and 0.88 for CH4 and GP, respectively. With regards to covariates, the covariates which significantly affected the fermentation products differed among CH4, GP and MP. Accordingly, the GP was significantly increased by the TF concentration, and decreased by HHDP, PD%, and mDP, while the CH4 production was decreased by PA concentration, CP, and mDP. The MP was significantly decreased by the PD%.
Discussion
The inclusion of forages with diverse functional groups (e.g. legumes) in mixtures generally improves the yield stability in the grassland-based livestock production systems20,21. Forages rich in tannins and other polyphenols provide even more benefits as they have been shown to decrease CH4 and N emissions from ruminant based production systems22,23. Although, several studies have analyzed TRFs for their effect on animal performance and CH4 reduction potential, the results till date have been contrasting, making it difficult to adapt the forages on the large scale. The reason for these contrasting results is assumed to be at least partially on the environmental conditions during plant growth, as these can affect tannin synthesis, as well as the results from colorimetric tannin assays that do not identify tannin structural characteristics or other compounds with potential interactions17,24. Simultaneously, CH4 emissions can vary highly across different animal species as well as the measurement methods. Both in vivo and in vitro techniques have been found to have their own set of advantages and disadvantages, leading to inconsistent results across different studies. In vivo techniques can accurately identify the impact of TRFs in an animal, yet they require large amounts of feed and are highly cost intensive thus, limiting the number of replicates or treatments. Additionally, they are still affected by the pre-diet and breed of the experimental animals hence, variability still exists across several in vivo studies25. On the other hand, in vitro studies are inexpensive and can be used to test several feedstuffs simultaneously with sufficient number of replicates, yet they fail to take into account the changes in rumen processes such as shifts in ruminal pH as well as ruminal buffering capacity during the fermentation of the feed26,27. However, in vitro studies are crucial in screening the promising feed additives on ruminal fermentation and predicting their effects in vivo. In the present study, the comparative assessment of 17 cultivars from eight agronomically promising temperate forage species, across two harvests, provides a novel insight of how the bioactive properties of these forages vary across the different species, and how stable are their antimethanogenic activity across different species and their cultivars under these harvests.
PA structural characteristics were found to be more stable than forage chemical composition across harvests
The CP levels in legumes are known to generally be in the range of 180 to 300 g/kg of DM and this mostly exceeds animal requirements (130–170 g/kg of DM)28. As expected, in the present study, CP concentrations in the forages were higher in the second harvest where the plants were harvested at the vegetative stage compared to the first harvest (flowering stage). This can be explained by the much higher leaf shares in second harvest, as leaves are richer in CP than stems29. Higher CP content increases the digestibility of the forage, as it is easily degraded by microbes30. Conversely, high NDF and ADF content in the forages is known to be an indicator of lower forage digestibility28. A meta-analysis by Maccarana, et al.31 found that NDF content in the feed is negatively correlated to GP in the rumen. It can reduce the availability of rapidly fermentable carbohydrates leading to lower microbial activity in the rumen and hence a decrease in GP32. The flavonoid content of the cultivars was also determined, due to potential matrix effects between tannins and flavonoids, as well as because of known anti-inflammatory and antimicrobial properties that are beneficial for ruminant immunological system (Olagaray and Bradford 2019). Assessing the effect of tannins on ruminants based only on their concentration has given rise to ambiguous results in the past as structural traits of tannins have been found to play an important role in determining their bioactivity11. Consequently, PA structural features were also analyzed in addition to concentration as they have been hypothesised to play an important role in determining their bioactivity, yet are rarely analysed in the animal studies. In the present study, these features were found to be stable across the harvests, and within the cultivars of the same species, the differences were comparatively lower than the variability across the species.
Methane and gas production from TRFs were consistently lower than lucerne in both the harvests
Numerous studies have demonstrated that TRFs reduce enteric fermentation in ruminants, resulting in lower GP and CH4 production33,34,35. In the first harvest, across all the species with varying forage quality and plant metabolite composition, the largest reductions in CH4 and GP were observed from TRFs, which were sulla, big trefoil, sainfoin and salad burnet, in comparison with lucerne. This could be attributed to the high TT concentration in these forages compared to lucerne. This is in line with the previous studies, where TRFs reduced the fermentation in the ruminants resulting in lower GP34. Tannins have the propensity to bind with protein and carbohydrates (both structural and non-structural carbohydrate), preventing fermentation and degradation of the molecules, thus lowering GP35. A study by Jayanegara et al.36 found that HTs are more potent compared to PAs. In the present study, however, this was not confirmed, as both the GP and CH4 production of PI 308861 (salad burnet) were found to be higher than those of Sudda (sulla) and Lot 29 (big trefoil). This was despite the higher TT and TP concentrations in salad burnet, which reaffirms the results from studies, indicating that the extent of CH4 reduction from TRFs does not only depend on tannin concentration but also on the tannin source and structural features37,38. At the animal scale, the differences between PAs and HTs bioactivity is also likely to be dependent on their fate in biological systems. Proanthocyanidins exist in the form of large polymers, enabling them to bind to the feed components strongly and they do not get easily degraded in the rumen. On the other hand, HTs have been shown to undergo hydrolysis, hence degrading to form low molecular weight phenolics in the rumen, and can thereby, lose part of their bioactivity39,40. However, the enzymatic or auto-oxidation of the initial HTs or their hydrolysis products could also produce large bio-oligomers or polymers with tannin-like functions in the rumen or small intestine39. Consequently, this makes it difficult to assess the fate of tannins in biological systems from in vitro studies alone. A general trend of higher GP and CH4 production, when plants were harvested at the vegetative stage (second harvest), was recorded compared to when they were harvested at the flowering stage (first harvest) which could be a result of lower tannin content in the plants in the second harvest. Furthermore, lower content of instantly fermentable fractions, such as water soluble carbohydrates in the leaves at the flowering stage can result into lower GP41. Interestingly, the species with lower TP and TT content, such as ribwort plantain and chicory, were also found to reduce CH4, but it was not accompanied by reduction in GP. This could result from PSMs other than tannins, found in these forages. A study by Navarrete, et al.42, found that chicory is rich in inulin and sesquiterpene lactones and, ribwort plantain leaves are known to contain acteoside, aucubin, and catapol.
Additionally, it is important to look at the CH4 percentage (MP) in the total gas, as this can be used as an indicator of the potential CH4 emissions per unit of organic matter (OM) degraded. Due to the concomitant reductions in GP and CH4 from TRFs, their MP ranged between 21 and 26% and hence was comparable to that of lucerne (24%) (Supplementary, Table S3). Chicory and ribwort plantain (17–21%) had the lowest MP compared to the other tested species in this study. The reduction in the CH4 from these species was not accompanied by the reduction in GP indicating their higher potential for CH4 abatement in ruminants without negatively affecting ruminal fermentation.
PEG treatment affirmed the antimethanogenic activity of TRFs, however no influence on methane percentage in total gas was observed
The inclusion of PEG as a tannin inactivating agent, enabled the quantification of the specific tannin-effect from different species on the ruminal fermentation in vitro. Polyethylene glycol can inactivate tannins by displacing proteins from tannin-protein complexes, and binding with tannins to form tannin-PEG complexes. Increased GP and CH4 production from TRFs under + PEG treatment could be a result of high availability of protein after being dissociated from tannin-protein complexes. The increased availability of N compounds that can no longer bind to tannins with the addition of PEG leads to more substrate for microbial degradation and higher microbial growth. This in turn results into increased ruminal fermentation and higher GP38,43. Simultaneously, as the tannins are inactive they can no longer influence the methanogens in the rumen which could affect CH4 production from these species. As expected, no differences in GP and CH4 production were observed from the samples of ribwort plantain, lucerne and chicory, due to the absence of tannins in these species. Generally, the variation in the extent of GP and CH4 reduction from across the different TRFs under −PEG and + PEG treatments was in line with the variation in tannin concentration and composition. Accordingly, stronger PEG-treatment effect was observed in cultivars with higher tannin concentrations, which is in line with the studies by Basha et al.43 and Jayanegara et al.35.
Although, it is evident that tannins reduce CH4 emissions, the mode of action by which they reduce CH4 in ruminants is still not well understood. A study by Hassanat and Benchaar33 found that the PA (acacia and quebracho) and HT (chestnut and valonea) extracts had a more pronounced effect on rumen methanogenesis compared to the substrate degradation33. Tannins are found to modify the rumen microflora by either directly inhibiting methanogenic population or indirectly affecting methanogen-protozoa symbiosis by inhibiting protozoal population in rumen7,17. In a study by O'Donovan and Brooker44, PAs from acacia were able to change the morphology of Streptococcus bovis, Butyrivibrio fibrisolvens and Prevotella ruminicola, and, hence, affected their activity. Additionally, HTs and PAs from different plant sources were also found to decrease CH4 production by suppressing the methanogenic archaea and protozoal populations in vitro45.
PA structural features were able to explain the variability in methane production from PA containing species but model was not improved
As mentioned above, not only the differences between HT and PA are relevant, but also their structural characteristics affect their potential bioactivity. Both mDP and PD% are among the most relevant features of PAs, determining their protein precipitation activity, which in turn is linked to anthelmintic and antimethanogenic bioactivity of PAs11,46. Hence, in the present study, a stepwise regression was performed to analyze which forage parameters were best suited to explain the total gas and CH4 production. By identifying covariates as potential predictors, fermentation patterns and CH4 reduction potential could be estimated from plant traits. This would enable extrapolations to any other species in terms of their bioactivity and would potentially help in predicting their influence on ruminants. However, the updated model (covariate-based model) with forage composition parameters as covariates did not improve the base model in terms of AIC value. Nevertheless, the conditional R2 for CH4 and GP remained the same and even increased for MP. This indicates that the overall updated model, including both fixed and random effects, was able to explain the variance of CH4 and GP in the present study for PA containing species. However, in order to predict the fermentation parameters, cultivar remains the better estimate compared to the sum of all covariables, indicating that due to the large number of factors with interconnected effects, the tannin source remains more relevant compared to the traits alone. This could also arise from the fact that in addition to PAs, plants may produce other bioactive PSMs such as terpenes, organic acids, saponins and others which can also influence the CH4 production from the substrate6,47 which were not quantified in this study. Furthermore, the influence of matrix effects between tannins and other PSMs on tannin bioactivity remains unexplored. Still, the model quality was sufficient to identify parameters which had a strong influence on the in vitro fermentation end-products. Particularly in case of CH4, the variables CP, PA concentration and mDP, were found to exhibit a negative relationship with CH4 production. This is in accordance with the study by Hatew et al.48, which observed that the polymer size (mDP) was an important determinant of in vitro CH4 and GP production from the substrate. With regards to MP, however, both PA and PD% were found to have limited influence as indicated by their near-zero coefficient and low marginal R2 (0.05). One solution to improve the predictive capabilities of CH4 reductions from tannin traits compared to forage quality traits might be the inclusion of tannin extracts in in vitro incubation instead of the whole plant sample.
The tested forage species are promising alternatives as sustainable feedstock in livestock production systems
The results in the present study underpin that TRFs have high potential for their use in livestock production as their CP content across both harvests was comparable to lucerne. This was further demonstrated by the GP production in the presence of PEG, which did not vary significantly from lucerne. The inclusion of PEG treatment further established the role of tannins in the antimethanogenic activity of TRFs. Hydrolysable tannins have been observed to affect methanogens in the rumen, however, the mode of action of PA containing forages has been found to be variable24. A significant treatment effect was observed when sulla and big trefoil were employed as a substrate, indicating their higher antimethanogenic activity which resulted from their high PA content and potent PA structural characteristics. Sulla is a highly palatable forage and its supplementation can reportedly reduce the dependency of the livestock production systems on proprietary anthelmintics49. The supplementation of fresh fed big trefoil was found to reduce CH4 emissions by 26% (g CH4 kg−1 DM intake) compared to lucerne in sheep50. However, the nutritional value of sulla and big trefoil need to be evaluated before providing them as a substantial feed component. Reportedly, owing to the high tannin content, sulla and big trefoil can compromise both, N digestibility and microbial activity in rumen which leads to lower GP, in addition to CH4 reduction51. Furthermore, forages containing PAs with high mDP and high PD% (big trefoil, sulla and sainfoin) are able to reduce protein degradation more effectively than PC-rich forages such as birdsfoot trefoil38. Additionally, PA concentration in the leaves of birdsfoot trefoil in our study, was found to be too low (approx. 0.1%) to exhibit detectable antimethanogenic activity. The concomitant reduction in GP and CH4 from TRFs in the present study, makes it essential to research on how these forages can be incorporated in animal feed and in which proportion, to exploit their beneficial effects. A study by Orlandi, et al.52 found that even though total-tract N digestibility by PA inclusion decreased linearly with the increased concentration, there was a linear increase in N retention and the efficiency of N utilization was improved with the inclusion of tannin extracts of Acacia mearnsii until the concentration of 18 g/kg DM, implying that the positive impact of duodenal flux of amino acid supply can outweigh the reduction in protein digestibility when tannins are supplied at optimum levels52. Although more research is required to determine the relationship between PAs and post ruminal amino acid availability, it is well established that low to moderate PA concentrations leads to increased rumen undegraded protein and post ruminal amino acid flux which improves the N utilization efficiency in ruminants6.
Furthermore, the mechanism by which tannins exert their antimethanogenic activity needs to be investigated. There is a possibility that rumen microbiome can get adapted to these bioactive compounds which could influence their effect in the long term and, thus, their CH4 reduction potential6,53. However, the research on this topic is still limited, and further research is warranted to understand long-term feeding effects of tannin-containing legumes rumen microbiome and their antimethanogenic activity. The stability of PA structural features across the different harvests is a promising ground to include PA structural features for future structure–activity relationship studies of tannins.
Additionally, this study also showed that underutilized forbs, such as chicory and ribwort plantain, have a high CH4 reduction potential despite being void of tannins. The CH4 reduction from these species did not result in a negative impact on GP when compared to the other tested species in this study. This is in line with an in vivo study by Niderkorn et al.54, which found that sheep produced 23% lower CH4 per kg DM intake with pure chicory compared to when fed with pure perennial ryegrass (Lolium perenne L., cv. AberAvon) which no significant differences in the DM digestibility. The bioactive compounds, aucubin and acteoside, in these forages are found to reduce net NH3 production during ruminal fermentation, and increase potential total GP42. Both chicory and ribwort plantain are promising alternatives to produce high quality feed to overcome feed-deficits during dry seasons due to their drought tolerance, and their inclusion in grass-clover mixtures can enhance above- and below-ground primary production in temporary grasslands55,56. Furthermore, they are known to benefit animal health due to their antimicrobial and anthelmintic properties which could be attributed to the presence of the PSMs other than polyphenols which were not analyzed in this study23,57.
Conclusions
In summary, low intraspecies variation compared to interspecies variation in antimethanogenic potential of these species is a promising basis for breeding and selection opportunities. Our study found that the intraspecies variability was much lower compared to the interspecies variability in terms of polyphenolic composition as well as PA structural features, and these features were generally stable across the harvests. Furthermore, these variations were translated to the antimethanogenic activity of the species. The reproducibility of the bioactive effects of TRFs across both the harvests provides a promising outlook of their potential to reduce CH4 in the livestock production systems. However, as the measurements were conducted under controlled conditions, it is crucial to assess their potential under field conditions. Additionally, inclusion level of these species in ruminant diet should also be assessed to prevent their negative effects on digestibility and animal performance. Inclusion of PEG treatment was able to establish the role of tannins in the antimethanogenic activity of TRFs. Furthermore, it was illustrated that the antimethanogenic property of the PA containing forages is at least in part a consequence of PA structural characteristics, as the highest CH4 reduction was observed from sulla containing highest PD% and mDP value. Moreover, the stability of PA structural characteristics across the harvests and less variation within the cultivars of the same species than across the species, provides an opportunity for breeding and selection programmes based on PA structural characteristics. Hence, follow up studies to screen the species with high antimethanogenic property for their variation in PA structural features and bioactivity are suggested, which can in turn improve the predictability of structure–activity relationship of tannins. Assessing the effect of PA structural features in addition to PA concentration on the bioactivity of these forages could be a useful criterion to understand the diverse effects of different plant sources containing similar PA concentration, as these differences have been shown to produce quantifiable effects.
Methods
Experimental design
A greenhouse experiment was conducted with 17 different forage cultivars from 8 species (Table 4). The seeds for the plants were obtained from the seedbank of IPK Leibniz Plant Genetics and Crop Plant Research, Gatersleben. The plants were harvested twice at different developmental stages. In the first harvest, plants were harvested at the flowering stage and in the second harvest, which was done after a 6-week regrowth interval, plants were harvested at the vegetative stage. The plants were grown in flower pots of 24 × 24 × 40 cm3 (LxWxH) each and 5 plants were sown in each pot. Each cultivar had 4 pots as replicates. The pots were arranged in a complete randomized design and reflective foils were installed to ensure even lighting. For details see Verma et al.19. The plants were consequently separated into leaves, stems and flowers. The samples were freeze dried and stored at − 80 °C for further analysis. The leaf samples were subsequently ball milled, and stored at − 80 °C to preserve the structural integrity of the polyphenols in the sample. The average monthly temperature in the greenhouse during the plant growth ranged from 24 to 27 °C (March to August). The experimental research on plants complies with the relevant institutional, national, and international guidelines and legislation.
Chemical composition of forages
Nitrogen concentration of the forages was determined by the DUMAS rapid combustion method using a CN elemental analyser (Vario Max Elementar Analysensysteme, Hanau, Germany). Crude protein (CP) was calculated by multiplying the N concentration with 6.25. The concentrations of neutral detergent fibre (NDF) was analysed according to Van Soest detergent procedure58 and AOAC method 973.18. The methods were adapted to be used with fibre analyser Ankom A220 (Ankom Technology, Macedon, NY, USA).
Polyphenol and tannin analysis with UPLC-DAD-MS/MS
The leaf tannin extracts were prepared for the UPLC-DAD-MS/MS as described by Verma, et al.19. The samples were analyzed with an Acquity UPLC (Waters Corp., Milford, MA, USA) coupled with a XEVO triple-quadrupole mass spectrometer (Waters Corp., Milford, MA, USA) as described by Engström et al.59. The system consisted of a sample manager, a binary solvent manager and a column, coupled with diode-array detector. The UV spectra was recorded. For the elution, acetonitrile (A) and 0.1% aqueous formic acid (B) were used with a constant flow rate of 0.5 mL min–1 with the following elution profile: 0–0.5 min, 0.1% A (isocratic); 0.5–5.0 min, 0.1–30% A (linear gradient); 5.0–8.0 min, 30–45% A (linear gradient); 8.0–11.5 min, column wash and stabilization. The data was recorded from 0 to 6 min for UV–Vis (190–500 nm) and MS data (m/z 100 to 2000). The specifications for the negative electrospray ionization are as follows; capillary voltage: 2.4 kV, desolvation temperature: 650 °C, source temperature: 150 °C, flow rate of desolvation and cone gas (N2): 1000 and 100 L/h, respectively, and collision gas: argon.
Hohenheim gas test
From the 4 pots per cultivar, due to analytical constraints, 3 pots were selected for analysis with the Hohenheim gas test (HGT) in the first harvest. For the same reason, in the second harvest, the biomass from all pots for each cultivar was pooled. The GP and CH4 production from the samples were analyzed using in vitro HGT60. The plant samples (200 ± 1 mg) were added to 100 mL calibrated glass syringes (Haeberle Labortechnik, Lonsee-Ettlenschieß, Germany) and rotated inside the incubator (39 °C) for 24 h. The samples were run in triplicates and each run was performed on two different days leading to 6 replicates per sample. Additionally, to standardize each run, four blanks (no plant material) as well as hay and concentrate standards from the Institute of Animal Science, University of Hohenheim, were used as a reference, and were included in triplicates. The animals were fed a ration consisting of grass hay (3 kg) and concentrate (3 kg) divided into 2 meals (7 a.m. and 4 p.m.). Ruminal fluid was collected prior to morning feeding from two ruminally-cannulated, non-lactating crossbred heifers (Jersey × German Black Pied). It was filtered with cheese cloth into a prewarmed insulated flask, immediately transferred to the lab, and mixed with a freshly prepared buffer solution at 1:2 ratio (v/v). The mixture was continuously stirred and flushed with CO2 while maintained in a water bath (39 °C). The buffered ruminal fluid (30 ml) was added to the syringes containing plant samples and syringes were placed back in the incubator with the rotor set to about one rotation per minute during the period of incubation (24 h). The GP and CH4 production from the samples were measured at the time interval of 8 and 24 h. The GP was recorded from the scale printed onto the syringes and methane concentration in the fermentation gas was measured with an infrared spectrometer (Methan AGM 10, Firma Sensors Europe, Ratingen, Germany). The infrared spectrometer was calibrated with pure nitrogen gas (zero point) and a gas mixture of methane and CO2 (60:40) as standard. The measured GP volume from the samples after 24 h was corrected with an average of the factors obtained from hay and concentrate standards. These constant values were calculated based on the measured and the targeted GP values of 45.9 ml/200 mg DM and 65.13 ml/200 mg DM after 24 h incubation of hay and concentrate standards, respectively.
Additionally, to evaluate the specificity of tannin effect on the gas production parameters, PEG treatment was included to attenuate the effect of tannin on fermentation. The samples were incubated with 200 mg PEG (MW: 6,000 kDa). The PEG treatments were referred as + PEG (samples with PEG) and −PEG (samples without PEG).
Statistical analysis
The statistical analysis was done with using the software R61. A linear mixed model was used with cultivars, harvests, treatment, and their interactions (two- and threefold) as fixed factors. Analytical replicates from the HGT per experimental pots and per date of experiment were used as random factor. Graphical residual analysis was performed and based on that, the data was assumed to be normally distributed and heteroscedastic. An ANOVA and multiple contrast tests were conducted to compare the significance of the influence factors on different levels. All tests of significance were made at P < 0.05.
Second, to assess whether the measured forage quality parameters of PA containing species can explain the fermentation end products, a (forward) model selection was done based on the AIC value (Akaike information criterion) as described by Venables and Ripley62. Therefore, the same model as above was used here as a reference model containing a subset of only PA containing species and the “−PEG” values (without the corresponding PEG treatment effects). In the reference model, the fixed factors were cultivar (only PA containing species), harvest and their interaction effect, and the random effect were the analytical replicates from the HGT per experimental pots and per date of experiment. Subsequently, the previous fixed factors were removed and replaced consecutively with forage trait derived covariates (forage quality and tannin traits). The decision for or against a certain covariate was also based on its significance and its variance inflation factor (VIF; < 5) until the optimal model with the lowest complexity was achieved. The final, covariate-based model was then compared with the reference model.
Data availability
Data sharing not applicable. The data generated during the study is included within this article and supplementary information.
References
Haque, M. N. Dietary manipulation: A sustainable way to mitigate methane emissions from ruminants. J. Anim. Sci. Technol. 60, 1–10. https://doi.org/10.1186/s40781-018-0175-7(2018) (2018).
IPCC, 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Masson-Delmotte, V., P. Zhai, A. Pirani, S.L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M.I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J.B.R. Matthews, T.K. Maycock, T. Waterfield, O. Yelekçi, R. Yu, and B. Zhou (eds.)]. Cambridge University Press (in press).
Lauder, A. R. et al. Offsetting methane emissions—An alternative to emission equivalence metrics. Int. J. Greenh. 12, 419–429. https://doi.org/10.1016/j.ijggc.2012.11.028 (2013).
Hill, J., McSweeney, C., Wright, A. G., Bishop-Hurley, G. & Kalantar-Zadeh, K. Measuring methane production from ruminants. Trends Biotechnol. 34, 26–35. https://doi.org/10.1016/j.tibtech.2015.10.004 (2016).
Van Zanten, H. H. E. et al. Defining a land boundary for sustainable livestock consumption. Glob Change Biol. 24, 4185–4194. https://doi.org/10.1111/gcb.14321 (2018).
Naumann, H. D., Tedeschi, L. O., Zeller, W. E. & Huntley, N. F. The role of condensed tannins in ruminant animal production: Advances, limitations and future directions. Rev. Bras. de Zootec. 46, 929–949. https://doi.org/10.1590/S1806-92902017001200009 (2017).
Mueller-Harvey, I. Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 86, 2010–2037. https://doi.org/10.1002/jsfa.2577 (2006).
Burggraaf, V. T. et al. Morphology and agronomic performance of white clover with increased flowering and condensed tannin concentration. N. Z. J. Agric. Res. 49, 147–155. https://doi.org/10.1080/00288233.2006.9513704 (2006).
Einarsson, R. et al. Crop production and nitrogen use in European cropland and grassland 1961–2019. Sci. Data 8, 288. https://doi.org/10.1038/s41597-021-01061-z (2021).
Salminen, J.-P. & Karonen, M. Chemical ecology of tannins and other phenolics: we need a change in approach. Funct. Ecol. 25, 325–338. https://doi.org/10.1111/j.1365-2435.2010.01826.x (2011).
Zeller, W. E. Activity, purification, and analysis of condensed tannins: current state of affairs and future endeavors. Crop Sci. 59, 886–904. https://doi.org/10.2135/cropsci2018.05.0323 (2019).
Barbehenn, R. V. & Peter Constabel, C. Tannins in plant–herbivore interactions. Phytochemistry 72, 1551–1565. https://doi.org/10.1016/j.phytochem.2011.01.040 (2011).
Chung, Y. H. et al. Enteric methane emission, diet digestibility, and nitrogen excretion from beef heifers fed sainfoin or alfalfa1. J. Anim. Sci. 91, 4861–4874. https://doi.org/10.2527/jas.2013-6498 (2013).
Christensen, R. G. et al. Effects of feeding birdsfoot trefoil hay on neutral detergent fiber digestion, nitrogen utilization efficiency, and lactational performance by dairy cows1. J. Dairy Sci. 98, 7982–7992. https://doi.org/10.3168/jds.2015-9348 (2015).
Jonker, A. & Yu, P. The occurrence, biosynthesis, and molecular structure of proanthocyanidins and their effects on legume forage protein precipitation, digestion and absorption in the ruminant digestive tract. Int. J. Mol. Sci. 18, 1105. https://doi.org/10.3390/ijms18051105 (2017).
Barry, T. N. & McNabb, W. C. The implications of condensed tannins on the nutritive value of temperate forages fed to ruminants. Br. J. Nutr. 81, 263–272. https://doi.org/10.1017/S0007114599000501 (1999).
Verma, S., Taube, F. & Malisch, C. S. Examining the variables leading to apparent incongruity between antimethanogenic potential of tannins and their observed effects in ruminants—A review. Sustainability 13, 2743. https://doi.org/10.3390/su13052743 (2021).
Malisch, C. S. et al. Large variability of proanthocyanidin content and composition in Sainfoin (Onobrychis viciifolia). J. Agric. Food Chem. 63, 10234–10242. https://doi.org/10.1021/acs.jafc.5b04946 (2015).
Verma, S., Salminen, J.-P., Taube, F. & Malisch, C. S. Large inter- and intraspecies variability of polyphenols and proanthocyanidins in eight temperate forage species indicates potential for their exploitation as nutraceuticals. J. Agric. Food Chem. 69, 12445–12455. https://doi.org/10.1021/acs.jafc.1c03898 (2021).
Lorenz, H., Reinsch, T., Kluß, C., Taube, F. & Loges, R. Does the admixture of forage herbs affect the yield performance, yield stability and forage quality of a grass clover ley?. Sustainability 12, 5842. https://doi.org/10.3390/su12145842 (2020).
Hofer, D. et al. Yield of temperate forage grassland species is either largely resistant or resilient to experimental summer drought. J. Appl. Ecol. 53, 1023–1034. https://doi.org/10.1111/1365-2664.12694 (2016).
Mueller-Harvey, I. et al. Benefits of condensed tannins in forage legumes fed to ruminants : Importance of structure, concentration and diet compsition. Crop Sci. 59, 861–885. https://doi.org/10.2135/cropsci2017.06.0369 (2017).
Loza, C. et al. Assessing the potential of diverse forage mixtures to reduce enteric methane emissions in vitro. Animals 11, 1126. https://doi.org/10.3390/ani11041126 (2021).
Min, B. R. et al. Dietary mitigation of enteric methane emissions from ruminants: A review of plant tannin mitigation options. Anim. Nutr. 6, 231–236. https://doi.org/10.1016/j.aninu.2020.05.002 (2020).
van Gastelen, S., Dijkstra, J. & Bannink, A. Are dietary strategies to mitigate enteric methane emission equally effective across dairy cattle, beef cattle, and sheep?. J. Dairy Sci. 102, 6109–6130. https://doi.org/10.3168/jds.2018-15785 (2019).
Hatew, B. et al. Relationship between in vitro and in vivo methane production measured simultaneously with different dietary starch sources and starch levels in dairy cattle. Anim. Feed Sci. Technol. 202, 20–31. https://doi.org/10.1016/j.anifeedsci.2015.01.012 (2015).
Storm, I. M. L. D., Hellwing, A. L. F., Nielsen, N. I. & Madsen, J. Methods for measuring and estimating methane emission from ruminants. Animals 2, 160–183. https://doi.org/10.3390/ani2020160 (2012).
Dewhurst, R. J., Delaby, L., Moloney, A., Boland, T. & Lewis, E. Nutritive value of forage legumes used for grazing and silage. Irish J. Agric. Food Res. 48, 167–187 (2009).
Hakl, J., Fuksa, P., Konečná, J. & Šantrůček, J. Differences in the crude protein fractions of lucerne leaves and stems under different stand structures. Grass Forage Sci. 71, 413–423. https://doi.org/10.1111/gfs.12192 (2016).
Jayanegara, A., Makkar, H. & Becker, K. The use of principal component analysis in identifying and integrating variables related to forage quality and methane production. J. Indones. Trop. Anim. 34, 241–247. https://doi.org/10.14710/jitaa.34.4.241-247 (2009).
Maccarana, L. et al. Methodological factors affecting gas and methane production during in vitro rumen fermentation evaluated by meta-analysis approach. J. Anim. Sci. Biotechnol. 7, 35–35. https://doi.org/10.1186/s40104-016-0094-8 (2016).
Baruah, L., Malik, P. K., Kolte, A. P., Dhali, A. & Bhatta, R. Methane mitigation potential of phyto-sources from Northeast India and their effect on rumen fermentation characteristics and protozoa in vitro. Vet. World 11, 809–818. https://doi.org/10.14202/vetworld.2018.809-818 (2018).
Hassanat, F. & Benchaar, C. Assessment of the effect of condensed (acacia and quebracho) and hydrolysable (chestnut and valonea) tannins on rumen fermentation and methane production in vitro. J. Sci. Food Agric. 93, 332–339. https://doi.org/10.1002/jsfa.5763 (2013).
Naumann, H. et al. Relationships between structures of condensed tannins from texas legumes and methane production during in vitro rumen digestion. Molecules 23, 2123. https://doi.org/10.3390/molecules23092123 (2018).
Jayanegara, A., Makkar, H. P. S. & Becker, K. Addition of purified tannin sources and polyethylene glycol treatment on methane emission and rumen fermentation in vitro. Media Peternakan 38, 57–63. https://doi.org/10.5398/medpet.2015.38.1.57 (2015).
Jayanegara, A., Goel, G., Makkar, H. P. S. & Becker, K. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol. 209, 60–68. https://doi.org/10.1016/j.anifeedsci.2015.08.002 (2015).
Hatew, B. et al. Diversity of condensed tannin structures affects rumen in vitro methane production in sainfoin (Onobrychis viciifolia) accessions. Grass Forage Sci. 70, 474–490. https://doi.org/10.1111/gfs.12125 (2015).
Huyen, N. T. et al. Structural features of condensed tannins affect in vitro ruminal methane production and fermentation characteristics. J. Agric. Sci. 154, 1474–1487. https://doi.org/10.1017/S0021859616000393 (2016).
Salami, S. A. et al. Characterisation of the ruminal fermentation and microbiome in lambs supplemented with hydrolysable and condensed tannins. FEMS Microbiol. Ecol. https://doi.org/10.1093/femsec/fiy061 (2018).
Salminen, J. P., Karonen, M. & Sinkkonen, J. Chemical ecology of tannins: Recent developments in tannin chemistry reveal new structures and structure-activity patterns. Chem.-Eur. J. 17, 2806–2816. https://doi.org/10.1002/chem.201002662 (2011).
Bezabih, M., Pellikaan, W. F., Tolera, A., Khan, N. A. & Hendriks, W. Chemical composition and in vitro total gas and methane production of forage species from the Mid Rift Valley grasslands of Ethiopia. Grass Forage Sci. 69, 635–643. https://doi.org/10.1111/gfs.12091 (2013).
Navarrete, S., Kemp, P. D., Pain, S. J. & Back, P. J. Bioactive compounds, aucubin and acteoside, in plantain (Plantago lanceolata L.) and their effect on in vitro rumen fermentation. Anim. Feed Sci. Technol. 222, 158–167. https://doi.org/10.1016/j.anifeedsci.2016.10.008 (2016).
Basha, N. A., Scogings, P. F. & Nsahlai, I. V. Effects of season, browse species and polyethylene glycol addition on gas production kinetics of forages in the subhumid subtropical savannah, South Africa. J. Sci. Food Agric. 93, 1338–1348. https://doi.org/10.1002/jsfa.5895 (2013).
O’Donovan, L. & Brooker, J. D. Effect of hydrolysable and condensed tannins on growth, morphology and metabolism of Streptococcus gallolyticus (S. caprinus) and Streptococcus bovis. Microbiology 147, 1025–1033. https://doi.org/10.1099/00221287-147-4-1025 (2001).
Bhatta, R. et al. Difference in the nature of tannins on in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations. J. Dairy Sci. 92, 5512–5522. https://doi.org/10.3168/jds.2008-1441 (2009).
Naumann, H. D. et al. Effect of molecular weight and concentration of legume condensed tannins on in vitro larval migration inhibition of Haemonchus contortus. Vet. Parasitol. 199, 93–98. https://doi.org/10.1016/j.vetpar.2013.09.025 (2014).
Jayanegara, A., Goel, G., Makkar, H.P.S., & Becker, K. Reduction in methane emissions from ruminants by plant secondary metabolites: Effects of polyphenols and saponins. Food and Agriculture Organization of the United Nations (FAO) Rome, Italy, 151–157. ISBN 978-92-5-106697-3 (2010).
Hatew, B. et al. Impact of variation in structure of condensed tannins from sainfoin (Onobrychis viciifolia) on in vitro ruminal methane production and fermentation characteristics. J. Anim. Physiol. Anim. Nutr. 100, 348–360. https://doi.org/10.1111/jpn.12336 (2016).
Waghorn, G. C., Douglas, G. B., Niezen, J. H., McNabb, W. C. & Foote, A. G. Forages with condensed tannins-their management and nutritive value for ruminants. Proc. N. Z. Grassl. Assoc., 60, 89−98 (1998).
Woodward, S. L., Waghorn, G. C. & Lassey, K. Early indications that feeding Lotus will reduce methane emissions from ruminants. Proc. N. Z. Soc. Anim. Prod. 61, 23–26 (2001).
Molle, G. et al. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep: Part 1: Effects on feeding behaviour, intake, diet digestibility and performance. Livest. Sci. 123, 138–146. https://doi.org/10.1016/j.livsci.2008.11.018 (2009).
Orlandi, T., Kozloski, G. V., Alves, T. P., Mesquita, F. R. & Ávila, S. C. Digestibility, ruminal fermentation and duodenal flux of amino acids in steers fed grass forage plus concentrate containing increasing levels of Acacia mearnsii tannin extract. Anim. Feed Sci. Technol. 210, 37–45. https://doi.org/10.1016/j.anifeedsci.2015.09.012 (2015).
Patra, A. K. & Yu, Z. Effects of adaptation of in vitro rumen culture to garlic oil, nitrate, and saponin and their combinations on methanogenesis, fermentation, and abundances and diversity of microbial populations. Front. Microbiol. https://doi.org/10.3389/fmicb.2015.01434 (2015).
Niderkorn, V. et al. Effect of increasing the proportion of chicory in forage-based diets on intake and digestion by sheep. Animal 13, 718–726. https://doi.org/10.1017/S1751731118002185 (2019).
Lee, J., Hemmingson, N., Minneé, E. & Clark, C. Management strategies for chicory (Cichorium intybus) and plantain (Plantago lanceolata): Impact on dry matter yield, nutritive characteristics, and plant density. Crop Pasture Sci. 66, 168. https://doi.org/10.1071/CP14181 (2015).
Cong, W.-F., Jing, J., Rasmussen, J., Søegaard, K. & Eriksen, J. Forbs enhance productivity of unfertilised grass-clover leys and support low-carbon bioenergy. Sci. Rep. 7, 1422. https://doi.org/10.1038/s41598-017-01632-4 (2017).
Sanderson, M. A., Labreveux, M., Hall, M. H. & Elwinger, G. F. Nutritive value of chicory and English plantain forage. Crop Sci. 43, 1797. https://doi.org/10.2135/cropsci2003.1797 (2003).
Van Soest, P. J., Robertson, J. B. & Lewis, B. A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583–3597. https://doi.org/10.3168/jds.S0022-0302(91)78551-2 (1991).
Engström, M. T. et al. Rapid qualitative and quantitative analyses of proanthocyanidin oligomers and polymers by UPLC-MS/MS. J. Agric. Food Chem. 62, 3390–3399. https://doi.org/10.1021/jf500745y (2014).
Menke, K. & Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 28, 7–55 (1988).
R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2021).
Venables, B. & Ripley, B. Generalised linear models. In Modern Applied Statistics With S.(4th edition) 183–208 (Springer, 2013).
Acknowledgements
This research was supported by Deutsche Forschungsgemeinschaft (DFG): project number 406534244, grant number MA 8199′/1-1. The authors are grateful to Monika Paschke-Beese, who provided her technical assistance and guidance during in vitro methane measurements. We thank Sören Schneider, Vibha Nihalani, and Rita Kopp for their support during the greenhouse experiment and laboratory analyses. We are grateful to Milla Leppä, Anne Koivuniemi and Ilari Kuukkanen for technical support and guidance during UPLC−MS/MS analysis. We would like to thank Niclas Wowczuk of the Buschwerk Shop GmbH for their support in the planning and implementation of homogeneous growing conditions for the experimental plants during the greenhouse experiment.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Conceptualization, C.S.M.; data curation, C.K. and S.V.; formal analysis, S.V., M.H., and C.S.M.; funding acquisition, C.S.M.; investigation, S.V.; methodology, S.V., M.H. and R.B.; project administration, C.S.M.; supervision, C.S.M. and F.T.; visualization, S.V.; writing—original draft, S.V.; and writing—review and editing, S.V., S.W., J-P. S., M.H., A.S., R.B., C.K., F.T., and C.S.M. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Verma, S., Wolffram, S., Salminen, JP. et al. Linking metabolites in eight bioactive forage species to their in vitro methane reduction potential across several cultivars and harvests. Sci Rep 12, 10454 (2022). https://doi.org/10.1038/s41598-022-14424-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-14424-2
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
