
Abstract
Land modified for human use alters matrix shape and composition and is a leading contributor to global biodiversity loss. It can also play a key role in facilitating range expansion and ecosystem invasion by anthrophilic species, as it can alter food abundance and distribution while also influencing predation risk; the relative roles of these processes are key to habitat selection theory. We researched these relative influences by examining human footprint, natural habitat, and predator occurrence on seasonal habitat selection by range-expanding boreal white-tailed deer (Odocoileus virginianus) in the oil sands of western Canada. We hypothesized that polygonal industrial features (e.g. cutblocks, well sites) drive deer distributions as sources of early seral forage, while linear features (e.g. roads, trails, and seismic lines) and habitat associated with predators are avoided by deer. We developed seasonal 2nd -order resource selection models from three years of deer GPS-telemetry data, a camera-trap-based model of predator occurrence, and landscape spatial data to weigh evidence for six competing hypotheses. Deer habitat selection was best explained by the combination of polygonal and linear features, intact deciduous forest, and wolf (Canis lupus) occurrence. Deer strongly selected for linear features such as roads and trails, despite a potential increased risk of wolf encounters. Linear features may attract deer by providing high density forage opportunity in heavily exploited landscapes, facilitating expansion into the boreal north.
Similar content being viewed by others

Introduction
Habitat loss from the conversion of mature forests for human use is the most intensive form of human disturbance to forest species1, but the deleterious effects of fragmentation of otherwise intact forests by road networks and other linear features have been increasingly recognized2,3,4. Imposing anthropogenic landscape features of different shapes and patch composition onto a forest matrix can alter biodiversity indirectly by modifying community structure, shifting species distributions5,6,7 and influencing animal behaviour8. For any given species, landscape changes can manifest as altered abundance and distribution of food resources, as well as altered predation risk – two key components of resource selection for any prey species9,10.
Anthropogenic land-use change often creates suitable conditions for invasive and early seral vegetation that support herbivores that are better adapted to exploit anthropogenic landscapes11,12. Where herbivores capitalize upon novel sources of available forage13, land-use change may lead to expanding distributions. Range-expanding species can be considered invasive in that they negatively impact biodiversity and ecosystem function directly through increased competition and predation14, and indirectly through changes in disturbance regimes, nutrient levels, and micro-climate15,16.
In the western Nearctic boreal forest, extensive forest harvesting and petroleum extraction have altered landscape shape and composition17 outside the range of natural variability18. These disturbance types cumulatively alter species distributions5,19 and favour generalist species. In particular, white-tailed deer (Odocoileus virginianus, hereby referred to as ‘deer’) have thrived in this rapidly changing landscape as evidenced by the expansion of their northern range limit over the past fifty years, with populations increasing in abundance in areas of high human disturbance20,21,22. Historically limited by deep snow, poor quality forage, and cold temperatures, deer are now one of the most pervasive ungulates in western Canada’s boreal forest5,19,23. Deer range expansion is an indirect cause of decline for native subspecies of woodland caribou (Rangifer tarandus caribou)20,24, acting as apparent competitors by inflating the population size of their shared predator, wolves (Canis lupus), thereby increasing predation on caribou25.
The role of human land-use in sustaining boreal deer populations is attributed to early seral vegetation from forestry cutblocks18, petroleum extraction, and transportation networks26,27. Focus has been on polygonal features that create patches of disturbance, such as well pads and forestry cutblocks. There has been less focus on the widespread, high-density linear features (e.g. roads, seismic lines17,28) as sources of forage or predation risk24. While the response of other species, such as wolves (Canis lupus), to linear features is well known26,29 less is known about the role of linear features on ungulate habitat selection in the western boreal forest30,31 though these constitute one of the most intensive and extensive anthropogenic disturbances on western boreal landscapes28.
In eastern North America deer select roadside verges32 and deciduous stands, such that deer abundance increases with road density and forage opportunity33,34. However, the cumulative effects of two different disturbance forms—polygonal early-successional patches and linear features – on deer distribution and reproduction are a key emerging pattern27. For example, in western North America, deer reproductive success increases in areas with intensive resource extraction35. The behavioural component of these responses remains unclear.
More importantly, few studies have examined deer response to cumulative landscape disturbance in the context of predation risk, which is critical for most prey species. The theoretical underpinning of the interaction between anthropogenic landscape change and predation risk lies in optimal foraging theory, which predicts that animals will attempt to maximize energy gain per unit cost; hence deer will seek to maximize foraging opportunities while minimizing their exposure to temperature extremes, predators, and other threats36. Perceived predation risk results in anti-predator avoidance behaviour both temporally and spatially37,38 and can manifest as indirect predation risk when prey avoid landscape features used by predators38. Energetic trade-offs between forage acquisition and predation risk have been observed in deer38 and other ungulates38 including within the context of oil and gas development8. Deer energetic requirements in the boreal shield vary with biological and geographic seasonality, peaking during winter when movement and foraging are limited by deep snow20,39,40, compared to autumn when males compete for mates in the rut, or in spring when females give birth41. Understanding anti-predator response in prey habitat selection studies may better explain the impacts of landscape disturbance on prey distributions and the mechanisms of range expansion; it can focus restoration and management efforts to mitigate changes to spatial predator–prey processes and limit resulting negative impacts to wildlife species, and so remains a key pursuit for landscape ecologists and conservationists.
We ask whether linear and polygonal anthropogenic features, predation risk, or natural habitat best explain seasonal habitat selection of range-expanding Nearctic deer populations. Previous studies on deer near the limits of their northern range have used aerial surveys25, snow tracking23, and camera detections42 to quantify the relative importance of landscape disturbance to deer, but behavioural response requires individual-specific, high-density location data. We use high-frequency GPS collar data to examine seasonal habitat selection by deer at the level of their home range, where landscape disturbance is relevant to range expansion in the population.
We weigh support for six competing hypotheses to explain variability in deer habitat selection: (1) forage acquisition hypothesis, whereby deer select polygonal disturbances due to increased resource subsidies; (2) indirect predation risk hypothesis, whereby deer avoid linear features as heavily-used predator travel corridors; (3) predator-frequency avoidance hypothesis, whereby deer avoid high-use areas measured as predicted monthly occurrence of wolves and black bears; (4) null hypothesis, whereby deer select natural habitats not necessarily associated with subsidy or risk, delineated by different forest and vegetation types; (5) human footprint hypothesis, where deer make trade-offs with avoidance of industrial linear and selection for polygonal features; and (6) cumulative effects hypothesis, whereby deer respond to the combined effects of all natural and anthropogenic variables.
Methods
Study area
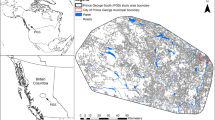
Our sampling frame is the western boreal forest of Canada. Our study area encompasses 3500 km2 of mixed-wood boreal forest in northeast Alberta intersecting the ranges of the Cold Lake and the East-side Athabasca River caribou herds43 (Fig. 1) and represents a high industrial disturbance portion of the frame. Polygonal features and linearization are spread across a gradient of disturbance from oil and gas development and forestry17. This fragmented, grid-like landscape of cleared forest is slow to regenerate28, creating a wide-spread source of early seral forage18 that benefit deer and are easily navigable by predators26,29. Intact natural vegetation is a heterogeneous mosaic of mixed-wood boreal forest dominated by black spruce (Picea mariana) and trembling aspen (Populus tremuloides) in lowland and upland deciduous stands with intersecting bogs, lakes, and rivers.

Seasonal white-tailed deer telemetry data collected in the Christina Lake study area near Conklin, Alberta encompassing 3500 km2 from 2012 to 2014. The area is extensively developed from forest harvesting and petroleum extraction, with features such as 3D seismic lines, legacy seismic lines, and roads appearing in as grey linear features and 0–10-year-old cutblocks as green polygons. Wetlands appear in purple.
Deer telemetry
Thirty-eight female deer were captured and collared in the winters of 2012, 2013, and 2014 in accordance with animal care protocols and permitted by Alberta Environment and Parks (Permits 49365 and 48602). All methods are reported in accordance with ARRIVE guidelines44. Deer were captured using clover traps which minimize deer stress relative to other modes of capture45. Individuals were fitted with LOTEK Iridium Track M 3D telemetry collars programmed to record locations at 2-h fixed intervals. Individuals with < 200 telemetry locations were removed (n = 2) to avoid including individuals with limited home-range coverage, resulting in 36 deer included within the analysis. We defined biological seasons for deer according to their life history stages and geography. Winter occurs from January 1st-April 30th, parturition from May 1st to June 30th, summer from July 1st to September 30th and rut from October 1st to December 31st20.
Estimating predation risk from camera trap data
Predation risk can be characterized as risky times or risky places46. Risky times are indicated by direct proximal predation risk by co-occurring predators; risky places are those where probability of predator encounter is high (but not necessarily occupied by a predator in a given time period46). Here we considered ‘risky places’ as those with high use by boreal deer primary predators: wolves (Canis lupus) and black bears (Ursus americanus)40,47,48. We did not have a direct measure of predation outcomes (e.g. mortalities attributed to predation, concurrently collared predators) so we used the probability of monthly occurrence of a predator at a site (hereafter occurrence frequency) as an indirect measure of risk, which we derived from a concurrent camera-trap survey in the study area5. The camera array of unbaited infra-red Reconyx PC900 Hyperfire remote digital camera was deployed using a stratified random design (see Fisher and Burton, 2018 for more details) at 62 sites from October 2011 to October 2014. Cameras were placed at least 2 km apart and the nearest camera to a road was 50 m with road density used as a model parameter. Predator-distribution models were developed by Fisher and Burton (2018); we used beta coefficients from the best-supported wolf and black bear models of occurrence frequency (Table S1) to extrapolate occurrence frequency for each predator across the study area using ArcMap 10.5 (Fig. S1). We assumed that model-estimated probability of monthly occurrence of each predator species is linearly related to the likelihood of collared deer encountering those predators, which we considered as a measure of predation risk38.
Landscape covariates
To examine deer response to natural habitat characteristics and anthropogenic forms of landscape disturbance relating to forage acquisition and predation risk, we used natural and anthropogenic land cover data quantified in ArcMap 10.5 (Table 1). Forest cover—percent crown closure of dominant overstorey species—were obtained from the Alberta Vegetation Index (AVI49). Anthropogenic landscape features were derived from Alberta Biodiversity Monitoring Institute (ABMI) human footprint maps (ambi.ca), with contemporary additional industrial features supplied by the ABMI Caribou Monitoring Unit (cmu.abmi.ca).
Seasonal resource selection functions
Resource selection functions (RSFs) characterize the probability of a resource unit being selected by an organism in a use-availability framework50. They are estimated by regressing used telemetry locations (1 s) against randomly generated available locations (0 s) within a defined domain of availability51. As our main interest related to habitat selection by a range-expanding ungulate, we were interested in second order selection (Johnson 1980); where individual deer selected their seasonal home ranges from within the surrounding available landscape. We therefore defined the domain of availability for each season as the buffered 100% minimum convex polygon (MCP) surrounding all used deer locations within the associated season. We calculated the mean radius of seasonal home-range areas and used these as buffer distances (winter = 2093 m, parturition = 4249 m, summer = 2172 m, rut = 3903 m) beyond each seasonal MCP; we randomly selected available locations, at a 1:1 used: available ratio, from these areas. Resource variables (Table 1) were extracted for each used and available location using the raster package52 in R 4.1.153. To check that the available sample represented the available landscape, we identified deer with the smallest and largest number of used locations for each season. Next, we created ten bootstrapped available datasets, each consisting of two times the number of used locations randomly sampled from within the respective deer’s seasonal MCPs, extracted covariates, and visually compared the distribution of variables across bootstrapped available samples. Variable distributions sufficiently overlapped across the bootstrapped samples (Figure S2–S4), irrespective of deer sample size (our sampling unit).
We generated six RSF models (Table 2) for each of four biological seasons to examine the drivers of population-scale deer resource selection. We used logistic regression in a Generalized Linear Mixed-Effects Model (GLMM)54. As we had a number of deer that contained data across multiple years, we included “individual-year” as a random effect on the intercept to avoid potential pseudo-replication arising from the dependent nature of telemetry data sampled from individuals and years55. We log-transformed distance variables and standardized percent cover and predator occurrence covariates (mean = 0, s.d. = 1) and tested for multicollinearity54. We excluded collinear variables with r > 0.7 and a variance inflation factor (VIF) > 356.
Models within each season were ranked using an information-theoretic approach based on Akaike Information Criteria corrected for small sample size (AICc) scores where the lowest AICc score reflects the most parsimonious model with the most deviance explained57,58. We used k-fold cross validation, whereby each individual’s data within corresponding seasons were split into k = 10 folds to validate each candidate model51 and each subset was evaluated using models trained with k – 9 alternative subsets.
Ethics and permits
All animals were captured and processed under approved handling protocols by InnoTech Alberta’s Animal Care Committee and permitted by Alberta Environment and Parks (Permit #s 49365 and 48602). All methods are reported in accordance with ARRIVE guidelines.
Results
Telemetry data
A total of 99,148 telemetry locations were obtained over the three-year period with a mean of 2754 locations per individual (range: 254–12,236). These data were subset by season resulting in 46,722 locations in winter, 27,573 in parturition, 9,987 in summer, and 14,866 in rut.
Seasonal drivers of deer habitat selection
Data strongly supported the cumulative effects hypothesis. Deer habitat selection was best predicted by a combination of anthropogenic linear and polygonal features, natural habitat composition, and predation risk, across all seasons (Table 3, Fig. 2). Human footprint was the second-best model across all seasons, and resource subsidies outperformed linear features and predation risk during the summer and rut (Table 3). The strength of deer selection and avoidance of landscape features varied across seasons, although the direction of selection remained constant for the majority of covariates (Fig. 3) and supported the importance of forage acquisition at both polygonal and linear anthropogenic features.

Binned seasonal white-tailed deer population level Resource Selection Function (RSF) relative abundance estimates near Conklin, Alberta Canada with overlaid road network and buffered 100% MCP for (a) Winter (cyan) (b) Parturition (magenta) (c) Summer (red) and (d) Rut (yellow) for combined years from 2012 to 2014. Data are spatially extrapolated to extend < 50 km from MCP boundaries.

Population-level relative probability of selection (mean and 95% confidence intervals) for white-tailed deer in northeastern Alberta as a function of (a) log-transformed distance to cutblocks, (b) log-transformed distance to roads, (c) log-transformed distance to 3D seismic lines and (d) relative probability of wolf occurrence and (e) relative probability of black bear occurrence. Confidence intervals incorporate error from the random effect of individual-year.
Deer strongly selected areas closer to industrial polygonal features (cutblocks and wellsites, Fig. 4a). Selection for cutblocks was weaker in the winter (βcutblock = − 0.53, 95% confidence interval (CI) = − 0.55 to − 0.50) and parturition seasons (βcutblock = − 0.65, CI = − 0.69 to − 0.63), compared to selection in the rut (βcutblock = − 1.01, CI = − 0.1.07 to − 0.95) and summer (βcutblock = − 1.06, CI = − 1.11 to − 1.00; Fig. 3a, 4a). Wellsite selection was relatively constant across all seasons (Fig. 4a). Deer selected roads and trails, while avoiding seismic lines and pipelines most of the year (Fig. 4b) except winter, although selection was extremely weak (βseismic = − 0.034, CI = − 0.049 to − 0.018); confidence intervals overlapped zero for pipelines during parturition (βpipeline = − 0.016, CI = − 0.045 to 0.013; Fig. 4b). Deer avoided 3D seismic lines more strongly than all other linear features, but less so in winter (β3D seismic = 0.28, CI = 0.26 to 0.31; Fig. 4b). Trails were selected in summer (βtrails = − 0.38, CI = − 0.43 to − 0.33) and rut (βtrails = − 0.44, CI = − 0.48 to − 0.39), less so in parturition (βtrails = − 0.27, CI = − 0.30 to − 0.24), and selection was almost neutral in the winter (βtrails = − 0.08, CI = − 0.10 to − 0.06; Fig. 4b). Roads were strongly selected in all seasons (Fig. 4b) though less so in summer (βroads = − 0.17, CI = − 0.22 to − 0.12).

Beta coefficient estimates and 95% confidence intervals from Generalized Linear Mixed-effect Models (GLMMs) for (a) polygonal features (b) linear features, (c) predation risk and (d) natural habitat features, for each season: winter, parturition (red), summer (green) and rut (orange). In panels a, b, and c where variables are a measure of distance, negative beta values represent selection and positive values represent avoidance to a feature. Vegetation and predator variables are interpreted as positive for selection and negative for avoidance.
Deer responded differently to areas occupied by black bears compared to wolves. Deer avoided areas with higher occurrence probability of black bears; as expected this signal was weakest in winter (βbear = − 0.018, CI = − 0.040 to − 0.0038; Fig. 4c) when black bears hibernate, lending confidence to our analysis. In contrast there was little consistent response that deer could avoid risky places with higher wolf occurrence. Contrary to expectations deer selected areas of high wolf occurrence during the rut (βwolf = 0.35, CI = 0.31–0.39), weakly selected areas with higher wolf probability of occurrence during winter (βwolf = 0.087, CI = 0.066–0.11) and parturition (βwolf = 0.13, CI = 0.10–0.16), but avoided wolf areas during the summer (βwolf = − 0.21, CI = − 0.26 to − 0.16.
Lastly, the strongest natural habitat drivers of deer selection were aspen (positive, Fig. 4d) and black spruce (negative, apart from winter; βblack spruce = 0.13 CI = 0.11–0.15). In summary, both polygonal and linear anthropogenic features played a strong role in deer habitat selection, with deer selecting early seral forage polygonal features, and selecting some linear features while avoiding others. Deer did not avoid areas of high predicted wolf predation risk, and in fact selected areas of estimated frequent wolf occurrence.
Discussion
Invading boreal white-tailed deer selected habitats with a combination of high-density polygonal and linear features, interspersed within natural landcover, suggesting forage acquisition drove the most consistent and greatest effect sizes in a model assessing both forage and predation. The boreal forest is changing in landscape composition and ecological community structure as the climate warms and resource extraction increases59,60. The twenty-first century boreal landscape differs from any historical form60, and anthropogenic features have the potential to shape the biotic processes within, including species’ range expansion and invasion. Our results suggest that these features may be contributing to Nearctic range expansion in white-tailed deer in all seasons.
Linear features had the strongest effect on deer habitat selection in the winter and parturition. Industrial linear features are pervasive in the boreal landscape17, and we hypothesized that deer would perceive these features as risky, due to their frequent use by apex predators such as wolves26,29. Deer avoided 3D seismic and pipelines, while selecting roads and trails throughout the year. Selection for these larger linear features may be explained by early seral vegetation forage subsidies at roadsides and verges and through “edge effects” into forest interiors30,61. Deer selected areas closer to roads and trails relative to the available landscape, likely due to an increase in these edge effects. The direction and magnitude of selection may be mediated by perceived risk of predation: features selected by deer are not only expected to have forage subsidies, but also are associated with human activity which may shield deer from predators62. However, deer selected areas farther from 3D seismic lines (generally without much early seral forage) and pipelines, which are less associated with predictable human activity and this behaviour could be inferred as predation avoidance.
Deer also selected areas closer to polygonal block features, particularly during the summer and rut where the resource subsidy hypothesis outcompeted linear features. The addition of both block features and linear features to predation risk and natural heterogeneity better explained deer selection than any set of features alone. The attraction to polygonal and large linear features is problematic as both features continue to expand as demand for oil increases28,63,64. The continuation of deforestation and linearization of the Nearctic boreal forest for seismic exploration, as well as ongoing timber harvesting, will likely sustain and facilitate boreal deer expansion.
Deer selection of habitat features associated with higher predicted predation risk by wolves or bears varied seasonally. During winter and parturition, deer weakly selected areas with greater probability of wolf activity, while strongly selecting it in the rut. This could be interpreted as resulting from two processes. First, wolves are cueing in on areas with abundant prey, especially deer: wolf distribution is strongly and positively associated with moose and deer5,19. Second, deer are either unable to avoid areas with wolves, or do not prioritize risk avoidance, instead prioritizing features that offer abundant resources despite the increased likelihood of encounter with wolves. Deer displayed the weakest association with wolves during the summer and we infer deer change their behaviour to prioritize actively avoiding encounters with wolves during this time when forage is abundant, and fawns are mobile but still vulnerable. Moreover, deer exhibited the weakest predator avoidance behaviour (in this case, strong positive association) during the rut, when mating occurs20. A reduction in vigilance behaviour during the rut was expected, due to deer prioritizing mating and forage acquisition to meet energetic requirements of reproduction and withstanding forage-limiting winter conditions65,66. Deer avoided habitat associated with bears during all seasons when bears are active on the landscape, suggesting spatial anti-predator response. Additionally, black bears have omnivorous diets, and opportunistically depredate fawns of deer and other neonates of boreal Cervidae47. We can therefore not expect bears to display the same increase in activity in deer habitat as we do with wolves. Alternatively, some habitat features avoided by bears are highly selected by deer, such as upland and lowland deciduous forest. Therefore, this relationship may arise due to an incompatibility of resource quality rather than being an example of avoidance behaviour; although bears avoided few features, so we deem this unlikely.
Our measure of predation risk across space (risky places) was the probability of predator occurrence based on monthly predator detections at camera sites over a three-year period5. We were conservative in that our estimates were extrapolated no farther than 50 km from the camera sites, and landscape features were categorized using regression coefficient estimates at the same spatial scale as they were measured in the best-fit models5. Nevertheless, it is possible that the wolf risk map did a poor job of predicting true wolf occurrence, manifesting as a positive relationship between wolves and deer. For example, due to limitations with sample size, we were unable to account for changes in predator distributions across deer biological seasons. Future research could include direct measures of predation risk (e.g. collared predators, deer mortalities) to better understand the extent to which deer make energetic trade-offs in the presence of predators, further exploring the risky times vs. risky places hypotheses67. However, a multi-species telemetry-based movement ecology study is needed for such high-resolution data. In the absence of this expensive research, camera traps can provide a useful index of predator occurrence for spatial inferences, and we hope camera data will inform analyses to greater extent in the future68.
Anthropogenic landscape change is a leading driver of seasonal deer habitat selection behaviour. Landscape change is also associated with deer distribution and reproduction at landscape scales27,35, and given these effects are consistent across the ecological hierarchy (behaviour, distribution, populations), we infer it is a key driver of deer expansion in western boreal landscapes. Range-expanding deer in the Nearctic boreal forest have numerous ecological implications for native fauna and flora. Deer are apparent competitors with declining woodland caribou24—where increasing deer populations have increased wolf populations, driving down caribou26. The recovery of woodland caribou is dependent on the reduction of habitat loss within their ranges and predation pressure by grey wolves69, and hence management of invasive deer. Local increases in deer populations can alter ecosystems by over-browsing, which reduces plant growth, forest understorey diversity, and consequently animal diversity20,70. Estimates for deer abundance are difficult to ascertain across terrain and cover types71,72 and whether carrying capacity for deer has been met in this system is not known. Anthropogenic landscape change facilitating deer invasion may entrain future landscape and biotic change where deer populations are on the rise.
Mitigating the multiple sources of anthropogenic resource subsidies for deer through landscape restoration may eliminate forage subsidies and reduce their ability to persist in harsh winter conditions – and hence curtail their expansion and ancillary biotic changes to the boreal ecosystem. Seismic line restoration, implemented where slow natural regeneration rates and rapid development of new seismic lines call for silvicultural intervention28, is already underway. The current caribou recovery plan for Alberta lists 10,000 km of restored seismic lines as a primary objective in the recovery of declining herds73 and contemporary research suggests there may be some effect on deer74. However, restoration of polygonal features is also needed to return the boreal landscape to one largely lacking sufficient forage suitable for deer range expansion.
As global needs for food, wood, minerals and petroleum increase, landscapes are reshaped and biodiversity declines75. The Canadian boreal forest is a substantial source of wood18 and energy76 while also serving a key carbon sink to mitigate climate change77. As human footprint grows and diversifies, the effects of this reshaped landscape on biota serve as an early model for boreal communities facing environmental change from resource extraction.
Data availability
Seasonal deer data will be available on Dryad.
References
Eisner, R., Seabrook, L. M. & McAlpine, C. A. Are changes in global oil production influencing the rate of deforestation and biodiversity loss?. Biol. Conserv. 196, 147–155. https://doi.org/10.1016/j.biocon.2016.02.017 (2016).
Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 34, 487–515. https://doi.org/10.1146/132419 (2003).
Pfeifer, M. et al. Creation of forest edges has a global impact on forest vertebrates. Nature 551, 187–191. https://doi.org/10.1038/nature24457 (2017).
Tilman, D., May, R., Lehman, C. & Nowak, M. Habitat destruction and the extinction debt. Nature 371, 65–66. https://doi.org/10.1038/371065a0 (1994).
Fisher, J. T. & Burton, C. A. Wildlife winners and losers in an oil sands landscape. Front Ecol. Environ. https://doi.org/10.1002/fee.1807 (2018).
Heim, N., Fisher, J. T., Volpe, J., Clevenger, A. P. & Paczkowski, J. Carnivore community response to anthropogenic landscape change: species-specificity foils generalizations. Landscape Ecol. 34, 2493–2507. https://doi.org/10.1007/s10980-019-00882-z (2019).
Pereira, H. M., Navarro, L. & Martins, I. Global biodiversity change: the bad, the good, and the unknown. Annu. Rev. Environ. Resour. https://doi.org/10.1146/annurev-environ-042911-093511 (2012).
Northrup, J. M., Anderson, C. R. Jr. & Wittemyer, G. Quantifying spatial habitat loss from hydrocarbon development through assessing habitat selection patterns of mule deer. Glob Change Biol. 21, 3961–3970. https://doi.org/10.1111/gcb.13037 (2015).
Holbrook, S. J. & Schmitt, R. J. The combined effects of predation risk and food reward on patch selection. Ecology 69, 125–134. https://doi.org/10.2307/1943167 (1988).
Moody, A. L., Houston, A. I. & McNamara, J. M. Ideal free distributions under predation risk. Behav. Ecol. Sociobiol. 38, 131–143 (1996).
Dietz, H. & Edwards, P. J. Recognition that causal processes change during plant invasion helps explain conflicts in evidence. Ecology 87, 1359–1367 (2006).
Hobbs, R. J. & Huenneke, L. F. Disturbance, diversity, and invasion: implications for conservation. Conserv. Biol. 6, 324–337 (1992).
Van der Graaf, S., Stahl, J., Klimkowska, A. & Drent, J. P. B. Surfing on a green wave—How plant growth drives spring migration in the Barnacle Goose Branta leucopsis. Ardea -Wageningen- 94, 567 (2006).
Parker, I. M. et al. Impact: toward a framework for understanding the ecological effects of invaders. Biol. Invasions 1, 3–19. https://doi.org/10.1023/A:1010034312781 (1999).
Pimentel, D., Zuniga, R. & Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 52, 273–288. https://doi.org/10.1016/j.ecolecon.2004.10.002 (2005).
Shackelford, N. et al. Primed for change: developing ecological restoration for the 21st Century. Restor. Ecol. 21, 297–304. https://doi.org/10.1111/rec.12012 (2013).
Pickell, P. D., Pickell, P. D., Andison, D. W., Coops, N. C. & Gergel, S. E. The spatial patterns of anthropogenic disturbance in the western Canadian boreal forest following oil and gas development. Can. J. For. Res. 45, 732–743. https://doi.org/10.1139/cjfr-2014-0546 (2015).
Fisher, J. T. & Wilkinson, L. The response of mammals to forest fire and timber harvest in the North American boreal forest. Mammal Rev. 35, 51–81 (2005).
Wittische, J., Heckbert, S., James, P. M. A., Burton, A. C. & Fisher, J. T. Community-level modelling of boreal forest mammal distribution in an oil sands landscape. Sci. Total Environ. 755, 142500. https://doi.org/10.1016/j.scitotenv.2020.142500 (2021).
Hewitt, D. G. Biology and management of white-tailed deer (CRC Press, Boca Raton, 2011).
McCabe, R. E. & McCabe, T. R. in White tailed deer: ecology and management Ch. Chapter 2, 19–72 (Stackpole, A Wildlife Management Institute Book, 1984).
Webb, R. The range of white-tailed deer in Alberta (Alberta Fish and Wildlife Division Edmonton, Alberta, 1967).
Dawe, K. L. & Boutin, S. Climate change is the primary driver of white-tailed deer (Odocoileus virginianus) range expansion at the northern extent of its range; land use is secondary. Ecol. Evol. 6, 6435–6451. https://doi.org/10.1002/ece3.2316 (2016).
DeCesare, N. J., Hebblewhite, M., Robinson, H. S. & Musiani, M. Endangered, apparently: the role of apparent competition in endangered species conservation. Anim. Conserv. 13, 353–362. https://doi.org/10.1111/j.1469-1795.2009.00328.x (2010).
Latham, A. D. M., Latham, M. C., McCutchen, N. A. & Boutin, S. Invading white-tailed deer change wolf-caribou dynamics in northeastern Alberta. J. Wildl. Manag. 75, 204–212. https://doi.org/10.1002/jwmg.28 (2011).
Latham, A. D. M., Latham, M. C., Boyce, M. C. & Boutin, S. Movement responses by wolves to industrial linear features and their effect on woodland caribou in northeastern Alberta. Ecol. Appl. 21, 11 (2011).
Fisher, J. T., Burton, A. C., Nolan, L. & Roy, L. Influences of landscape change and winter severity on invasive ungulate persistence in the Nearctic boreal forest. Sci. Rep. 10, 8742. https://doi.org/10.1038/s41598-020-65385-3 (2020).
Dabros, A., Pyper, M. & Castilla, G. Seismic lines in the boreal and arctic ecosystems of North America: environmental impacts, challenges, and opportunities. Environ. Rev. 26, 214–229. https://doi.org/10.1139/er-2017-0080 (2018).
Dickie, M., Serrouya, R., McNay, R. S., Boutin, S. & du Toit, J. Faster and farther: wolf movement on linear features and implications for hunting behaviour. J. Appl. Ecol. 54, 253–263. https://doi.org/10.1111/1365-2664.12732 (2017).
Finnegan, L., MacNearney, D. & Pigeon, K. E. Divergent patterns of understory forage growth after seismic line exploration: implications for caribou habitat restoration. For. Ecol. Manag. 409, 634–652. https://doi.org/10.1016/j.foreco.2017.12.010 (2018).
Prokopenko, C. M., Boyce, M. S., Avgar, T. & Tulloch, A. Characterizing wildlife behavioural responses to roads using integrated step selection analysis. J. Appl. Ecol. 54, 470–479. https://doi.org/10.1111/1365-2664.12768 (2017).
Waring, G. H., Griffis, J. L. & Vaughn, M. E. White-tailed deer roadside behavior, wildlife warning reflectors, and highway mortality. Appl. Anim. Behav. Sci. 29, 215–223. https://doi.org/10.1016/0168-1591(91)90249-W (1991).
Bowman, J., Ray, J. C., Magoun, A. J., Johnson, D. S. & Dawson, F. N. Roads, logging, and the large-mammal community of an eastern Canadian boreal forest. Can. J. Zool. 88, 454–467. https://doi.org/10.1139/z10-019 (2010).
Munro, K. G., Bowman, J. & Fahrig, L. Effect of paved road density on abundance of white-tailed deer. Wildl. Res. 39, 478. https://doi.org/10.1071/wr11152 (2012).
Fisher, J. T. & Burton, A. C. Spatial structure of reproductive success infers mechanisms of ungulate invasion in Nearctic boreal landscapes. Ecol. Evol. 11, 900–911. https://doi.org/10.1002/ece3.7103 (2021).
Kie, J. G. Optimal foraging and risk of predation effects on behavior and social structure in ungulates. J. Mammal. 80, 1114–1129 (1999).
Brown, J. S., Laundré, J. W. & Gurung, M. The ecology of fear: optimal foraging, game theory, and trophic interactions. J. Mammal. 80, 385–399. https://doi.org/10.2307/1383287 (1999).
Kittle, A. M., Fryxell, J. M., Desy, G. E. & Hamr, J. The scale-dependent impact of wolf predation risk on resource selection by three sympatric ungulates. Oecologia 157, 163–175. https://doi.org/10.1007/s00442-008-1051-9 (2008).
Moen, A. N. Energy conservation by white-tailed deer in the winter. Ecology 57, 192–198. https://doi.org/10.2307/1936411 (1976).
Schmidt, K. Winter ecology of nonmigratory Alpine red deer. Oecologia 95, 226–233. https://doi.org/10.1007/BF00323494 (1993).
Kilgo, J. C., Ray, H. S., Vukovich, M., Goode, M. J. & Ruth, C. Predation by coyotes on white-tailed deer neonates in South Carolina. J. Wildl. Manag. https://doi.org/10.1002/jwmg.393 (2012).
Laurent, M., Dickie, M., Becker, M., Serrouya, R. & Boutin, S. Evaluating the mechanisms of landscape change on white-tailed deer populations. J. Wildl. Manag. 85, 340–353. https://doi.org/10.1002/jwmg.21979 (2020).
Schneider, R. R., Hauer, G., Adamowicz, W. L. & Boutin, S. Triage for conserving populations of threatened species: the case of woodland caribou in Alberta. Biol. Conserv. 143, 1603–1611. https://doi.org/10.1016/j.biocon.2010.04.002 (2010).
Kilkenny, C., Browne Wj Fau - Cuthill, I. C., Cuthill Ic Fau - Emerson, M., Emerson M Fau - Altman, D. G. & Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLoS biol. 8(6), e1000412 (2010).
DelGiudice, G. D., Mangipane, B. A., Sampson, B. A. & Kochanny, C. O. Chemical immobilization, body temperature, and post-release mortality of white-tailed deer captured by clover trap and net-gun. Wildl. Soc. Bull. (1973-2006) 29, 1147–1157 (2001).
Droge, E., Creel, S., Becker, M. S. & M’Soka, J. Risky times and risky places interact to affect prey behaviour. Nat. Ecol. Evol. 1, 1123–1128. https://doi.org/10.1038/s41559-017-0220-9 (2017).
Kunkel, K. E. & Mech, L. D. Wolf and bear predation on white-tailed deer fawns in northeastern Minnesota. Can. J. Zool. 72, 1557–1565 (1994).
Latham, A., Latham, M., Knopff, K., Hebblewhite, M. & Boutin, S. Wolves, white-tailed deer, and beaver: Implications of seasonal prey switching for woodland caribou declines. Ecography https://doi.org/10.1111/j.1600-0587.2013.00035.x (2013).
Alberta Environment and Sustainable Resource Development. Alberta Vegetation Index. Accessed October 2016. https://geodiscover.alberta.ca/
Manly, B., McDonald, L., Thomas, D., McDonald, T. & Erickson, W.Resource selection by animals: statistical design and analysis for field studies. Vol. 63, pp. 1-10 (Springer Science & Business Media, 2007).
Boyce, M. S., Vernier, P. R., Nielsen, S. E. & Schmiegelow, F. K. A. Evaluating resource selection functions. Ecol. Model. 157, 281–300. https://doi.org/10.1016/S0304-3800(02)00200-4 (2002).
Hijmans, R. & van Etten, J. Raster: Geographic data analysis and modeling. CRAN R package 2 (2016).
R: A language and environment for statistical computing. (Vienna, Austria, 2013).
Zuur, A., Hilbe, J. & Ieno, E. A Beginner's Guide to GLM and GLMM with R: a frequentist and Bayesian perspective for ecologists. (Highland Statistics, 2013).
Gillies, C. S. et al. Application of random effects to the study of resource selection by animals. J. Anim. Ecol. 75, 887–898. https://doi.org/10.1111/j.1365-2656.2006.01106.x (2006).
Craney, T. A. & Surles, J. G. Model-dependent variance inflation factor cutoff values. Qual. Eng. 14, 391–403. https://doi.org/10.1081/QEN-120001878 (2002).
Akaike, H. Information theory and an extension of the maximum likelihood principle. Selected papers of hirotugu akaike 199–213 (Springer, New York, 1998).
Burnham, K. P. & Anderson, D. R. Multimodel inference: understanding AIC and BIC in model selection. Sociol. Methods Res. 33, 261–304. https://doi.org/10.1177/0049124104268644 (2004).
Boulanger, Y. et al. Climate change impacts on forest landscapes along the Canadian southern boreal forest transition zone. Landscape Ecol. 32, 1415–1431. https://doi.org/10.1007/s10980-016-0421-7 (2017).
Sulla-Menashe, D., Woodcock, C. E. & Friedl, M. A. Canadian boreal forest greening and browning trends: an analysis of biogeographic patterns and the relative roles of disturbance versus climate drivers. Environ. Res. Lett. 13, 014007. https://doi.org/10.1088/1748-9326/aa9b88 (2018).
St-Pierre, F., Drapeau, P. & St-Laurent, M.-H. Drivers of vegetation regrowth on logging roads in the boreal forest: Implications for restoration of woodland caribou habitat. For. Ecol. Manag. 482, 118846. https://doi.org/10.1016/j.foreco.2020.118846 (2021).
Berger, J. Fear, human shields and the redistribution of prey and predators in protected areas. Biol. Let. 3, 620–623. https://doi.org/10.1098/rsbl.2007.0415 (2007).
Heyes, A., Leach, A. & Mason, C. F. The economics of Canadian oil sands. Rev. Environ. Econ. Policy 12, 242–263. https://doi.org/10.1093/reep/rey006 (2018).
Komers, P. E. & Stanojevic, Z. Rates of disturbance vary by data resolution: implications for conservation schedules using the Alberta boreal forest as a case study. Global Change Biol. 19, 2916–2928 (2013).
Hebblewhite, M. & Merrill, E. H. Trade-offs between predation risk and forage differ between migrant strategies in a migratory ungulate. Ecology 90, 3445–3454. https://doi.org/10.1890/08-2090.1 (2009).
Mech, D. L. & Boitani, L. Wolves: behavior, ecology, and conservation Vol. 57 (University of Chicago Press, Chicago, 2004).
Creel, S., Winnie, J. A., Christianson, D. & Liley, S. Time and space in general models of antipredator response: tests with wolves and elk. Anim. Behav. 76, 1139–1146. https://doi.org/10.1016/j.anbehav.2008.07.006 (2008).
Steenweg, R. et al. Scaling-up camera traps: monitoring the planet’s biodiversity with networks of remote sensors. Front. Ecol. Environ. 15, 26–34. https://doi.org/10.1002/fee.1448 (2017).
Hebblewhite, M. Billion dollar boreal woodland caribou and the biodiversity impacts of the global oil and gas industry. Biol. Cons. 206, 102–111. https://doi.org/10.1016/j.biocon.2016.12.014 (2017).
Côté, S. D., Rooney, T. P., Tremblay, J.-P., Dussault, C. & Waller, D. M. Ecological impacts of deer overabundance. Annu. Rev. Ecol. Evol. Syst. 35, 113–147 (2004).
McCullough, D. R. Evaluation of night spotlighting as a deer study technique. J. Wildl. Manag. 46, 963–973. https://doi.org/10.2307/3808229 (1982).
Preston, T., Wildhaber, M., Green, N., Albers, J. & Debenedetto, G. Enumerating white-tailed deer using unmanned aerial vehicles. Wildlife Soc. Bull. https://doi.org/10.1002/wsb.1149 (2021).
Parks, A. E. Provincial woodland caribou range plan. 212 (Edmonton, Alberta, 2017).
Tattersall, E. R., Burgar, J. M., Fisher, J. T. & Burton, A. C. Boreal predator co-occurrences reveal shared use of seismic lines in a working landscape. Ecol. Evol. 10, 1678–1691. https://doi.org/10.1002/ece3.6028 (2020).
Diaz, S. et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science (New York N.Y.) https://doi.org/10.1126/science.aax3100 (2019).
Bayoumi, T. & Muhleisen, M. Energy, the exchange rate, and the economy: macroeconomic benefits of Canada’s oil sands production (International Monetary Fund, Washington, 2006).
Zhu, K., Song, Y. & Qin, C. Forest age improves understanding of the global carbon sink. Proc. Natl. Acad. Sci. 116, 3962. https://doi.org/10.1073/pnas.1900797116 (2019).
Acknowledgements
We thank M. Hiltz, L. Roy, L. Nolan, S. Melenka, S. Heckbert, B. Eaton, K. Richardson, K. Tereschyn, S. Eldridge, J. Dennett, J. Watkins, and T. Zembal at InnoTech Alberta. We further thank D. Pan and J. Cranston for mapping support, and F.E.C. Stewart, G. Chow-Fraser, and S. Frey for valuable comments.
Funding
Field research was primarily supported by InnoTech Alberta (IA), Alberta Environment and Parks (AEP). It was also supported by the Petroleum Technology Alliance of Canada AUPRF, and MEG Energy. SD was supported by Natural Sciences and Engineering Council of Canada (NSERC) CGS scholarship and a MITACS Canada Accelerate Grant. JTF and AL were supported in part by the Oil Sands Monitoring Program, and this research does not necessarily reflect the views of that program.
Author information
Authors and Affiliations
Contributions
S.D. and A.L. developed statistical models for deer. S.D. created predation risk maps and wrote the draft manuscript. A.C.B. and J.T.F. conducted predator abundance modelling. J.T.F. conceived the research, created the design, and led the team. All authors contributed to hypothesis development, manuscript edits, and provided feedback.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Darlington, S., Ladle, A., Burton, A.C. et al. Cumulative effects of human footprint, natural features and predation risk best predict seasonal resource selection by white-tailed deer. Sci Rep 12, 1072 (2022). https://doi.org/10.1038/s41598-022-05018-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-05018-z
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
