
Abstract
The number of items in an array can be quickly and accurately estimated by dividing the array into subgroups, in a strategy termed “groupitizing.” For example, when memorizing a telephone number, it is better to do so by divide the number into several segments. Different forms of visual grouping can affect the precision of the enumeration of a large set of items. Previous studies have found that when groupitizing, enumeration precision is improved by grouping arrays using visual proximity and color similarity. Based on Gestalt theory, Palmer (Cognit Psychol 24:436, 1992) divided perceptual grouping into intrinsic (e.g., proximity, similarity) and extrinsic (e.g., connectedness, common region) principles. Studies have investigated groupitizing effects on intrinsic grouping. However, to the best of our knowledge, no study has explored groupitizing effects for extrinsic grouping cues. Therefore, this study explored whether extrinsic grouping cues differed from intrinsic grouping cues for groupitizing effects in numerosity perception. The results showed that both extrinsic and intrinsic grouping cues improved enumeration precision. However, extrinsic grouping was more accurate in terms of the sensory precision of the numerosity perception.
Similar content being viewed by others

Introduction
Numerosity perception is a quantitative attribute of entities that is an important dimension of nature. For example, one tree produces more fruits than another, and there are more sheep in one territory than in another. In the process of understanding and adapting to nature, humans and animals have gradually evolved in their ability to perceive numerosity1,2,3.
Three strategies are usually used in numerosity perception: subitizing, counting, and estimation4,5. For a small number of items (usually less than 4), humans can quickly and accurately determine the number of items in the clusters. This is called “subitizing,” which indicates that the items can be understood immediately, without thinking4,6,7. With an increasing number of items (greater than 4), the time required to determine the number of items also increases correspondingly while requiring the coordination of many visual and spatial operations, as the observer determines the number of objects by counting8,9.When the number of items in clusters is large and cannot be counted in a very short time (for example, the presentation time of the stimulus is short), numerosity perception may be inaccurate, pursuant to Weber's Law, the approximate number system (ANS) can be relied upon10. People thus use estimation strategy to determine the approximate number of objects10,11,12.
Beyond subitizing, counting and estimation, recent studies have found that arrays visually divided into sub-groups can be enumerated faster and more accurately than ungrouped arrays; this is called “groupitizing”4,13,14,15,16,17. An increasing number of studies have begun to explore the mechanisms of groupitizing. For example, Starkey and McCandliss (2014) found that groupitizing is positively correlated with the arithmetic ability of children and adults, indicating that the grouping ability may reflect the arithmetic strategies13. Moscoso et al. (2020) suggested that groupitizing is a process based on attention, which depends on the subitizing system, and mathematical ability is correlated with groupitizing15. Ciccione and Dehaene (2020) indicated that in groupitizing, subjects use mental multiplication and mental addition to increase the speed and accuracy of enumeration4. Wege et al. (2021) explained that the numerical information needed for a mental calculation is extracted from grouped arrays, and suggested that the parallel subitizing of dots and groups in grouped arrays may represent the enumeration processes necessary for groupitizing via mental multiplication18.
The groupitizing effect must be based on visual grouping. Research into visual grouping processes and perceptual organization developed against the background of Gestalt theory approximately one hundred years ago19,20,21. Wertheimer (1923) proposed the main principles of perceptual grouping, which specified which regions of images constituted objects or perceptual units, such as similarity, proximity, symmetry, good continuity21. Based on Gestalt theory, Palmer made an important distinction between intrinsic and extrinsic grouping principles. Like most classical Gestalt principles, intrinsic principles are based on the inherent relationships among attributes of grouped elements (e.g., color, shape, size, position). In contrast, the extrinsic principles are based on relationships among elements and other extrinsic elements that induce them to group (e.g., connectedness or common region)22,23,24,25.
Previous studies of groupitizing only involved intrinsic grouping cues (color similarity4,14 and proximity4,13,14,15). To date, no research has explored numerosity perception with extrinsic grouping cues. Thus, this study explored whether extrinsic grouping cues are different from intrinsic grouping cues in numerosity perception. Previous studies have found that extrinsic grouping cues have advantages over intrinsic grouping cues22,26,27,28,29,30,31. Luna et al. suggested that observers respond more quickly to extrinsic cues than to other grouping cues22,32. Quinn and Bhatt reported that young infants (3–4 months old) are sensitive to extrinsic cues, especially common region and connectedness22,28. Therefore, we hypothesized that extrinsic grouping cues would be advantageous in numerosity perception.
In addition, vision research has revealed that shape is crucial for object recognition33,34,35,36. Without other visual information, it is easy for humans to use shapes to identify objects. Human adults and children prefer to classify new objects according to their shape, given conflicting color and texture cues33,36,37. Accordingly, in this study, a shape similarity cue was added to the intrinsic grouping cues to verify whether the shape similarity grouping cues have different effects than other intrinsic grouping cues (i.e., color similarity and proximity).
Methods
Participants
Fifty-three freshman college students (mean age = 19 years, standard deviation = 2.4, range = 18–22) with normal (or corrected-to-normal) vision, and no color blindness were selected. We replicated the experiment in three groups of participants with low, medium, and high levels of math knowledge (for a similar approach, see Dehaene et al., 2020)4. At the highest level, we tested 16 science students majoring in mathematics, all of whom had scored over 120 points on China’s mathematics college entrance examination in 2020. For the medium level, we tested 18 humanities students with math scores between 60 and 90 in the college entrance examination (the mathematics component of the college entrance examination for science is more difficult than for humanities. The maximum score for mathematics is 150). We tested 19 students in the low-level group. They were sports students who had never taken university-level exams in mathematics or related disciplines. The third group was less familiar with mathematics as they had not been taught mathematics for at least one year.
Materials and procedure
Stimuli were presented using E-Prime 2.0. Participants sat in a quiet and dimly light room, 60 cm from a screen monitor (60 Hz). At the beginning of each trial, a fixation point was presented in the center of the screen and remained on the screen throughout the experiment. After 500 ms, a stimulus was displayed for 500 ms, followed by a screen with an input box. Participants estimated the number of stimuli present and entered the estimated result into the input box as quickly and accurately as possible using a numeric keypad (Fig. 1A). Response time was measured from stimulus offset to when the input box was presented. Each condition was tested in separate blocks, and participants were never explicitly informed of the grouping cues.

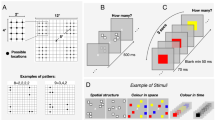
Illustration of the stimuli and procedure. (A) Example of the time course of the experiment. (B) Illustration of how stimulus position was defined in the grouping conditions. (C) Example of stimuli. Stimuli are not depicted to scale. Stimuli figures can be found as supplemental figures on the OSF page.
Stimuli
All stimuli were distributed in a 6° × 6° square grid, which consisted of 144 small squares, where each square was 0.4° × 0.4°, and the array was placed at the intersection of the grids, so that each item had 121 possible positions (Fig. 1B). We tested all numerosities between 5 and 17; there were 13 different numerosities. In the grouping conditions, each numerosity was divided into 2–4 subgroups, and each subgroup contained between 2 and 6 items, configurations were as following: 2, 2, 1; 3, 3; 3, 3, 1; 2, 2, 2, 2; 3, 3, 3; 3, 3, 3, 1; 3, 3, 3, 2; 3, 3, 3, 3; 5, 5, 3; 4, 4, 3, 3; 4, 4, 4, 3; 4, 4, 4, 4; 5, 4, 4, 4. The participants were clearly informed of the range of the numerosity. Each participant completed 390 trials in total, with each numerosity presented in 30 times. “Stimuli Details” files on the OSF page.
Extrinsic cues
Extrinsic cues included connectedness and common region.
Connectedness
In the connectedness conditions, the stimuli were sets of white squares (0.4° × 0.4°) with black borders randomly distributed in the grid. The squares within subgroups were connected by a black line, with the connection at the center of the square. In the no-grouping condition, there was no black line connection, and each item was randomly distributed in the large grid (Fig. 1B).
Common region
In the common region conditions, stimuli were also setting of white squares (0.4° × 0.4°) with black borders. The grid was divided into four quadrants, and the squares of each subgroup were randomly distributed inside the small square boxes (2.5° × 2.5°) in the four quadrants. For example, Fig. 1B is a 3, 3, 3 group with only three subgroups, so there are only three boxes. In the no-grouping condition, there were no small square boxes, and each item was randomly distributed in a large grid (Fig. 1B).
Intrinsic cues
Intrinsic cues included color similarity, shape similarity, and proximity.
Color similarity
The color similarity conditions were the same as those used by Anobile et al. (2020)14. Individual items (0.4° × 0.4°) could be red, blue, yellow, or green, (RGB: 255 0 0; 0 0 255; 255 255 0; 0 255 0, respectively). Colors were arranged from left to right, so that similar colors appeared in a vertical column (see Fig. 1C for a 3, 3, 3 group), where squares were first randomly arranged. The first three squares were colored red (from the left to right), the next three blue, and the remaining three yellow (colors were randomly selected for each group). In the no-grouping condition, positions of the squares were arranged with the same logic, but the colors were randomly assigned.
Shape similarity
The shape similarity condition was similar to the color similarity condition. The only difference was that four shapes replaced the four colors: square, circle, triangle, and diamond; all shapes were 0.4° × 0.4°, and white with black borders (Fig. 1C).
Proximity
The proximity conditions were the same as those used by Anobile et al. (2020)14. Stimuli were arranged into four possible groups of 12 possible positions. Each group (spanning a maximum area of 4° × 2°) was located in the same quadrant and centered at 5° from the central fixation point. Each group was first randomly assigned to one quadrant (between 1 and 4); then, the individual item positions were randomly selected between one of the 12 positions in the selected quadrant. Within each quadrant, the maximum center-to-center distance between each element was 4°, and the minimum was 1°. In the no-grouping condition, each item was randomly distributed in the large grid (Fig. 1C).
Data analysis
To statistically test differences across conditions. We adopted repeated-measures ANOVA, which included the grouping condition (2 levels for grouping and no-grouping), grouping cue (5 levels for connectedness, common region, color similarity, shape similarity, proximity), and numerosity (13 levels, from N5 to N17) as within-subjects factors. Math knowledge (high, medium, and low) was a between-subjects factor.
The median reaction times (RTs) for correct answers were computed for each subject. We excluded trials with RTs more than three standard deviations from the average RTs. Precision was measured by the coefficient of variation (CV), which is a dimensionless precision index that allows cross-numerical comparison of average performance.
Ni is the analyzed numerosity, and i is the standard deviation of the responses to numerosity i.
Data were analyzed by repeated measures ANOVA and t-tests, using JASP statistical package version 0.14.1.0 (https://jasp-stats.org) and IBM SPSS Statistics version 19 (http://www.spss.com/). In addition, we used Bayesian ANOVA inference for additional analysis, because quantifying evidence in favor of both difference and equality was crucial for testing our hypotheses (Wagenmakers et al. 2018)38. We reported the Bayes factors in favor of the alternative (BF10). A BF10 larger than 1 indicated evidence supporting the alternative hypothesis, and a BF10 less than 1 indicated evidence for the null hypothesis. We applied Bonferroni corrections to all post hoc analyses to correct for multiple comparisons.
(The full raw data from this experiment are available on our OSF page. We thank an anonymous reviewer for this suggestion).
Statement
All coauthors agreed with the contents of the manuscript. The study with human subjects was conducted in accordance with the Declaration of Helsinki. This study was approved by the School of Psychology Ethics Committee at Guizhou Normal University. All participants signed informed consent forms prior to the experiment. All methods were carried out in accordance with relevant guidelines and regulations.
Results
As in several previous studies14,15, we also investigated grouping effects on RT and sensory precision (as indexed by the coefficient of variation CV) (Eq. 1). CV is a classical psychophysical parameter; in numerosity perception, this parameter reflects the sensory noise associated with the estimation process: the higher the CV value, the more sensory noise, and thus the less precise the estimates.
Tables 1 and 2 shows the main effect and interaction of ANOVA for RT (Table 1) and CV (Table 2).
Groupitizing and grouping cues
We compared the numerosity perception of extrinsic grouping cues (connectedness, common region) and intrinsic grouping cues (color similarity, shape similarity, proximity), average across numerosities and conditions. A t-test revealed strong statistical evidence for the differences between the extrinsic and intrinsic grouping cues for CV (p < 0.01). As shown in Fig. 2, the CV for extrinsic grouping cues was lower than that for intrinsic grouping cues, indicating that the sensory precision of extrinsic grouping cues was more accurate (less sensory noise) than that of intrinsic grouping cues (Fig. 2). But the extrinsic and intrinsic grouping cues differences for RTs were not significant.

Results of extrinsic and intrinsic grouping cues. CV for extrinsic and intrinsic grouping cues by group condition. ***p < 0.001; **p < 0.01; *p < 0.05.
The ANOVA of RTs revealed a significant main effect of the grouping condition, F (1, 51) = 13.001, p < 0.001***, BF10 > 100, the grouping conditions reacted faster than that in no-grouping conditions. And the main effect of grouping cue was also significant, F (4, 47) = 15.526, p < 0.001***, BF10 > 100. The interaction between grouping cue and grouping condition was significant, F (4, 47) = 18.451, p < 0.001***, BF10 > 100. We performed a simple effects analysis to further test the differences in grouping conditions at different grouping cues. The result can be seen from Fig. 3, for extrinsic grouping cues, there was no significant difference in RT between grouping and no-grouping conditions. But for intrinsic grouping cues, proximity and shape similarity grouping cues had a better grouping effect (the reaction was significantly faster in the grouping condition than in the no-grouping condition) (Fig. 3).

Grouping cue effect. RTs (A) and CV (B) for different grouping cues by group condition (G for grouping condition, NG for no-grouping condition; C for connectedness, CR for common region, P for proximity, S for shape, Color for color). ***p < 0.001; **p < 0.01; *p < 0.05.
The ANOVA of CV revealed a significant main effect of grouping conditions, F (1, 50) = 2.807, p < 0.001***, BF10 > 100, and the grouping conditions had lower CV than no-grouping conditions, which means that grouping condition had less sensory noise and more accuracy. The ANOVA of CV also revealed a significant main effect of grouping cue F (4, 47) = 2.894, p = 0.034*, BF10 > 100; but its interaction with the grouping condition was not significant.
Groupitizing and specific numerosity
ANOVA of RTs revealed a significant main effect of numerosity F (12, 39) = 289.35, p < 0.001***, BF10 > 100. From Fig. 4A, it is evident that RT increased linearly with numerosity, small numerosities reacted significantly faster than large numerosities, and the interaction with the grouping condition was also significant, F (12, 39) = 55.306, p < 0.001***, BF10 > 100. We performed a simple effects analysis to further test the differences in grouping conditions at different numerosities and examined how RTs varied with numerosity in each condition. The results showed that, in the grouping condition, numerosities 6, 9, 12, and 16 had faster RTs than adjacent numbers. In contrast, numbers 7, 11, 13, 17 had slower RTs than their neighbors (Fig. 4A).

Specific numerosities. RTs (A) and CV (B) for different numerosities across group conditions.
ANOVA of CV revealed a significant main effect of numerosity, F (12, 39) = 96.83, p < 0.001***, BF10 > 100. From Fig. 4B, it is evident that CV increased linearly with the numerosity and small numerosities were more accurate than large numerosities. The interaction with the grouping condition was also significant, F (12,39) = 7.395, p < 0.001**, BF10 > 100. Similar to RT, in the grouping condition, numerosities 6, 9, 12, and 16 had lower CV than their neighbors (Fig. 4B). The results were consistent with previous research4,14,15.
Moreover, we found that large numerosities were underestimated for each grouping cue (Fig. 5). In Fig. 5, the dotted line represents the accurate perception of numerosities. Above the dotted line indicates overestimation, while below the dotted line indicates underestimation. From Fig. 5, we can see that under all grouping cues, the subjects began to be underestimated from the value of 13, consistent with the results of previous studies14.

Perceived numerosity. Average perceived numerosity for all grouping cues and tasks, averaged across participants.
Influence of math knowledge
We compared grouping effects for subjects with high, medium, and low levels of math knowledge. ANOVA of RTs revealed a significant main effect of math knowledge, F (2, 52) = 4.798, p = 0.012, BF10 > 100. The subjects in the high math knowledge group had faster RTs, and its interaction with the grouping condition was also significant, F (2, 50) = 1.496, p = 0.004, BF10 > 100. We performed a simple effects analysis to further test the differences in grouping conditions at different levels of math knowledge. We found that the grouping effect was significant only in groups with high math knowledge (see Fig. 6A).

Results for math knowledge. RTs (A) and CV (B) in groups with high, medium, and low levels of math knowledge by group condition. ***p < 0.001; **p < 0.01; *p < 0.05.
ANOVA of CV also revealed a significant main effect of math knowledge, F (2, 52) = 4.798, p = 0.012, BF10 > 100, the subjects in the high math knowledge group had lower CV; but its interaction with the grouping condition was not significant (Fig. 6B).
Discussion
Our results showed that when items were divided into several subgroups, this benefited perception of the numerosity. Furthermore, according to Gestalt theory, perceptual grouping can be divided into extrinsic and intrinsic grouping cues23,24,25. Accordingly, this study explored whether different grouping cues had different influences on groupitizing. The results showed that the sensory precision of extrinsic grouping cues was more accurate than that of intrinsic grouping cues, and the grouping effect was stronger.
The RT for extrinsic grouping cues and intrinsic grouping cues was not significant, inconsistent with Luna et al. (2016), who found that extrinsic grouping cues, especially common regions, were associated with faster RTs than other grouping cues22,32. In addition, Quinn and Bhatt (2015) also found that early infants (4–6 months) were more sensitive to extrinsic grouping cues26,28. Although the RTs for extrinsic grouping cues and intrinsic grouping cues were not significant, the sensory precision of extrinsic grouping cues were more accurate than that of intrinsic grouping cues (Fig. 3). This may indicate that extrinsic grouping cues have a strong advantage of groupitizing. Compared with intrinsic grouping cues, the addition of connecting lines or closing lines led to more visual interference. It also required additional cognitive processing, thus leading to slower responses for extrinsic grouping cues.
Additionally, due to the great similarity between proximity and common region (proximity is distributed in four quadrants, while the common region is divided into four quadrants by the border), we compared the grouping effects of proximity and common region. As shown in Fig. 3A, for RTs, proximity has a grouping effect (the grouping condition reacts significantly faster than the no-grouping condition), while common region shows no grouping effect (the difference between the grouping condition and no-grouping is not significant). For CV, both proximity and common region have a strong grouping effect. The CV under the grouping condition is significantly lower than that under the no-grouping condition, and the grouping effect of the common region is stronger (the difference between the grouping condition and no-grouping condition is more significant, p < 0.001) than proximity. Future research should select preschool children or first-grade primary school children to explore whether children who have not studied mathematics or have no complete magnitude representation system have different groupitizing effects given different grouping cues.
In recent years, significant progress has been made in the visual science of perceptual grouping. Recent studies have focused on the temporal processes and neural basis of intrinsic and extrinsic perceptual grouping26,39. For intrinsic grouping cues, grouping by proximity was found to be related to the positive component at the occipital electrode, whose amplitude peaks 100 to 120 ms after stimulus onset. The collinearity contour integral emerged 130 ms after stimulation40. Grouping by similarity (shape or color) appeared much later, and after 300 ms from stimulus onset, the negative occipito-temporal wave was activated22,39. In contrast, the neural basis of extrinsic grouping principles has received less attention. Montoro et al. (2015) reported the neural effects associated with grouping by common regions. They found that common region grouping cues belong to the category of long-latency grouping principles, which primarily involve activity in extrastriate cortices22,28. Future research should continue to explore the time course and neural mechanisms underlying intrinsic and extrinsic grouping cues of grouping effects.
Interestingly, we found that RTs for grouping by shape similarity were significantly lower than those of the other groupings (Fig. 3). Studies have shown that, in the absence of other visual information, it is easy for human beings to identify objects by shape41,42,43. Adults and children prefer to categorize novel objects according to shapes, given conflicting colors and texture cues. Moreover, shape features play a more important role in inductive reasoning than do color features42,43. Shape similarity is the first strategy used in inductive reasoning in early childhood44. Researchers presented subjects with reference stimuli of color, shape, and texture (such as square, blue, and wooden). They then presented two test stimuli with different shapes, colors, and textures, so that children could judge whether the test stimulus was consistent with the reference stimulus45. The results showed that 2–3 year old children chose shapes as the basis of induction. Future studies should select developing children as participants to explore whether the grouping effect of quantity estimation in shape similarity is different between children and adults.
Regarding math knowledge, for RTs, the interaction between math knowledge and grouping condition was significant (Fig. 6A). The grouping conditions for the high math knowledge group differed significantly in RT, while the middle and low math knowledge groups did not exhibit such a difference, indicating that the groupitizing effect benefited the most with high math knowledge. Many studies have demonstrated that an efficient ANS may be a prerequisite for the typical development of math skills10,11. Therefore, we speculate that the high math knowledge group had a more refined ANS, and that the groupitizing strategy was automatically used in quantity estimation. In the grouping condition, items were visually divided into subgroups, and since individuals with high levels of math knowledge could make better use of groupitizing strategy, RTs in the grouping condition were significantly faster than those in the no-grouping condition. Although sensory precision was higher in the grouping condition for the middle and low math knowledge groups, RTs did not differ among conditions. This may be because, in the grouping condition, when they used groupitizing strategies, they needed to employ more cognitive resources and required more time. In the no-grouping condition, they could not use any strategies; they could only make rough guesses based on their feelings. Thus, RTs were faster but precision was lower. However, for CV, the interaction between math knowledge and grouping condition was not significant (Fig. 6B); this means that high, medium, and low math knowledge groups all have strong grouping effects, which shows that each group benefits from groupitizing in numerosity perception; It further proves that groupitizing is an effective strategy in numerosity perception.
The present study found that, in the grouping condition, numbers 6, 9, 12, and 16 were associated with faster RTs and lower coefficients of variation than adjacent numbers. This may be because those specific numerosities’ configurations (6: 3, 3 = 2 × 3 = 2 groups of 3, 9: 3, 3, 3 = 3 × 3 = 3 groups of 3, 12: 4, 4, 4 = 3 × 4 = 3 groups of 4, 16: 4, 4, 4, 4 = 4 × 4 = 4 groups of 4) were divided into “equal groups,” which let the subjects use mental multiplication in their estimations. This is similar to the result of Dehaene et al. (2020)4, who found that for 5, 7, 11, and other such prime numbers, RTs were slower than for their neighbors, and for non-prime numbers, which could be subdivided into equal numbers, RTs were faster than for their neighbors (Fig. 5).
Conclusion
The present study demonstrates that visually dividing an array into subgroups promotes numerosity perception. Moreover, our research combined the groupitizing effect of numerosity estimation with Gestalt theory for the first time. It also demonstrated a difference between the groupitizing effect of extrinsic versus intrinsic grouping cues, based on Palmer et al.23,24,25. The results thus suggest that it takes longer for to estimate numerosities given extrinsic grouping cues. However, the precision of extrinsic grouping cues is higher than that of intrinsic grouping cues due to a stronger groupitizing effect.
Limitations and future directions
Since the concept of “groupitizing” was proposed by Wender and Roth Kegel (2000)17 and Starkey and McCandless (2014)13, studies have continued to explore the effect of grouping. This study combined the grouping effect and perceptual grouping principles, thus extending the study of groupitizing and the field of perceptual grouping.
Recent studies have begun to explore the shared associative mechanisms between different perceptual features. For example, the theory of magnitude model46,47 proposes that the parietal cortex of human beings processes quantitative information about space, time, and numbers together to optimize action plans and execution. It is necessary to explore the relationship between magnitude representation, space, and time. In this study, we only studied the grouping effect in space. However, subsequent studies should verify the differences in the grouping effect between intrinsic and extrinsic grouping cues in the time dimension.
The participants in this study were adults. Although they were divided into high, middle, and low math knowledge groups, the difference in the math level of adults is not very prominent. Many studies have found that in the process of development and formal arithmetic learning, numerosity perception precision has been greatly improved. In contrast, symbolic mathematics abilities in educated adults may have been stably mapped into their basic non-symbolic representation, making this connection less obvious46,47,48. Future research should explore the differences in groupitizing effects between preschool children with a low number sense and adults, as well as between children with difficulties in math and children without math difficulties.
To date, there has been no electrophysiological or neuroscience study that explores the grouping strategies between intrinsic and extrinsic cues. Future research should be conducted from the perspective of electrophysiology to investigate the neurofunctional links between grouping strategies and intrinsic and extrinsic cues, which would delineate a possible neural hierarchical model for “groupitizing.”
Data availability
Materials and data are available on our OSF page: https://osf.io/m5ay7/.
References
Jevons, W. The power of numerical discrimination. Nature 3, 363–372 (1871).
Deheane, S. The number sense: how the mind creates mathematics. Nature 391 (1998).
Fischer, U., Moeller, K., Bientzle, M., Cress, U. & Nuerk, H. C. Sensori-motor spatial training of number magnitude representation. Psychon. Bull. Rev. 18, 177–183 (2011).
Ciccione, L. & Dehaene, S. Grouping mechanisms in numerosity perception. Open Mind 4, 102–118 (2020).
Burr, D. & Ross, J. A visual sense of number. Curr. Biol. 18, 425–428 (2008).
Revkin, S., Piazza, M., Izard, V., Cohen, L. & Dehaene, S. Does Subitizing Reflect Numerical Estimation? Psychol. Sci. 19, 607–614 (2008).
Pome, A., Anobile, G., Cicchini, G. M. & Burr, D. C. Different Reaction-Times for Subitizing, Estimation, and Texture. J Vis. 19, 14 (2019).
Soltész, F., Szűcs, D. & Szűcs, L. Relationships between magnitude representation, counting and memory in 4-to 7-year-old children: a developmental study. Behav. Brain Funct. 6, 1–14 (2010).
Chapman, C. S. et al. Counting on the motor system: rapid action planning reveals the format- and magnitude-dependent extraction of numerical quantity. J. Vis. 14, 30 (2014).
Odic, D. & Starr, A. An introduction to the approximate number system. Child Dev. Perspect. 12, 223–229 (2018).
Zhou, X., Shen, C., Li, L., Li, D. & Cui, J. Mental numerosity line in the human’s approximate number system. Exp. Psychol. 63, 169–179 (2016).
Dehaene, S. The neural basis of the weber-fechner law: a logarithmic mental number line. Trends Cogn. Sci. 7, 145–147 (2003).
Starkey, G. S. & McCandliss, B. D. The emergence of “groupitizing” in children’s numerical cognition. J. Exp. Child Psychol. 126, 120–137 (2014).
Anobile, G., Castaldi, E., Moscoso, P. A. M., Burr, D. C. & Arrighi, R. “Groupitizing”: a strategy for numerosity estimation. Sci. Rep. UK 10, 1–9 (2020).
Maldonado Moscoso, P. A., Castaldi, E., Burr, D. C., Arrighi, R. & Anobile, G. Grouping strategies in number estimation extend the subitizing range. Sci. Rep. UK 10, 1–9 (2020).
Poom, L., Lindskog, M., Winman, A. & van den Berg, R. Grouping effects in numerosity perception under prolonged viewing conditions. PLoS ONE 14, e207502 (2019).
Wender, K. & Rothkegel, R. Subitizing and its subprocesses. Psychol. Res. 64, 81–92 (2000).
Theresa E. Wege, K. T. M. I. Finding the subitizing in groupitizing evidence for parallel subitizing of dots and groups in grouped arrays. (2021).
Elder, J. H. & Goldberg, R. M. Ecological statistics of gestalt laws for the perceptual organization of contours. J. Vis. (Charlottesv, Va.) 2, 324–353 (2002).
Sasaki, Y. Processing local signals into global patterns. Curr. Opin. Neurobiol. 17, 132–139 (2007).
Graham, L. Gestalt laws of perception. Des. Princ. Pract. 3, 385–394 (2009).
Luna, D., Villalba-Garcia, C., Montoro, P. R. & Hinojosa, J. A. Dominance dynamics of competition between intrinsic and extrinsic grouping cues. Acta Psychol. (Amst.) 170, 146–154 (2016).
Palmer, S. E. Common region: a new principle of perceptual grouping. Cognit. Psychol. 24, 436 (1992).
Palmer, S. E. & Beck, D. M. The repetition discrimination task: an objective method for studying perceptual grouping. Percept. Psychophys. 69, 68 (2007).
Palmer, S. & Rock, I. On the nature and order of organizational processing: a reply to Peterson. Psychon. Bull. Rev. 1, 515–519 (1994).
Montoro, P. R. et al. A Temporo-spatial analysis of the neural correlates of extrinsic perceptual grouping in vision. Neuropsychologia 69, 118–129 (2015).
Han, S. Interactions between proximity and similarity grouping: an event-related brain potential study in humans. Neurosci. Lett. 367, 40–43 (2004).
Montoro, P. R., Villalba-García, C., Luna, D. & Hinojosa, J. A. Common region wins the competition between extrinsic grouping cues: evidence from a task without explicit attention to grouping. Psychon. Bull. Rev. 24, 1856–1861 (2017).
Villalba-Garcia, C., Jimenez, M., Blanco, D., Hinojosa, J. A. & Montoro, P. EXPRESS: competition between perceptual grouping cues in an indirect objective task. Q. J. Exp. Psychol. 92378008 (2021).
Villalba-García, C., Santaniello, G., Luna, D., Montoro, P. R. & Hinojosa, J. A. Temporal brain dynamics of the competition between proximity and shape similarity grouping cues in vision. Neuropsychologia 121, 88–97 (2018).
Devyatko, D. & Pastukhov, A. Extrinsic grouping factors in motion-induced blindness. PLoS ONE 13, e192133 (2018).
Luna, D. & Montoro, P. R. Interactions between intrinsic principles of similarity and proximity and extrinsic principle of common region in visual perception. Perception 40, 1467–1477 (2011).
Elder, J. H. Shape from contour: computation and representation. Ann. Rev. Vis. Sci. 4, 423–450 (2018).
Kennedy, G. J., Orbach, H. S., Gordon, G. E. & Loffler, G. Judging the shape of moving objects: discriminating dynamic angles. J. Vis. 8, 9 (2008).
Chen, N. & Watanabe, K. Color-shape associations affect feature binding. Psychon. Bull. Rev. (2020).
Ayzenberg, V. & Lourenco, S. F. Skeletal descriptions of shape provide unique perceptual information for object recognition. Sci. Rep. UK 9, 9313–9359 (2019).
Burchfield, T. & Greene, E. Evaluating spatiotemporal integration of shape cues. PLoS ONE 15, e224530 (2020).
Wagenmakers, E. et al. Bayesian inference for psychology. Part II: Example applications with JASP. Psychon. Bull. Rev. 25, 58–76 (2018).
Villalba-García, C., Santaniello, G., Luna, D., Montoro, P. & Hinojosa, J. A. Temporal brain dynamics of the competition between proximity and shape similarity grouping cues in vision. Neuropsychologia 121, 88–97 (2018).
Rashal, E., Yeshurun, Y., Kimchi, R., Gauthier, I. & Enns, J. T. The time course of the competition between grouping organizations. J. Exp. Psychol. Hum. Percep. Perform. 43, 608–618 (2017).
Chen, N. & Watanabe, K. Color-shape associations affect feature binding. Psychon. Bull. Rev. 28, 169–177 (2020).
Reeves, A., Fuller, H. & Fine, E. M. The role of attention in binding shape to color. Vis. Res. 45, 3343–3355 (2005).
Graham, S., Kilbreath, C. & Welder, A. Thirteen-month-olds rely on shared labels and shape similarity for inductive inferences. Child Dev. 75, 409–427 (2004).
Welder, A. & Graham, S. The influence of shape similarity and shared labels on infants’ inductive inferences about nonobvious object properties. Child Dev. 72, 1653–1673 (2001).
Walsh, V. A theory of magnitude: common cortical metrics of time, space and quantity. Trends Cognit. Sci. 7, 483–488 (2003).
Feigenson, L., Libertus, M. & Halberda, J. Links between the intuitive sense of number and formal mathematics ability. Child Dev. Perspect. 7, 74–79 (2013).
Halberda, J., Mazzocco, M. & Feigenson, L. Individual differences in non-verbal number acuity correlate with maths achievement. Nature 455, 665–668 (2008).
Braham, E. & Libertus, M. When approximate number acuity predicts math performance: the moderating role of math anxiety. PLoS ONE 13, e195696 (2018).
Acknowledgements
This research was funded from the National Natural Science Foundation of China (Research on the Cognitive Mechanism and Development Characteristics of the Visual Selective Attention Affected on the Number Processing; NO, 31260234. Research on the Cognitive Mechanism and Development Characteristics of the Coding of Number-space Associations; NO, 31860281).
Author information
Authors and Affiliations
Contributions
All authors conceived and planned the experiments. H.Y.Y. and Y.P. developed the study concept and contributed to the study design. Testing and data collection were performed by M.M.L, J.Z. and L.H.C. The data analysis and interpretation was performed by H.Y.Y. under the supervision of Y.P. The manuscript was drafted by H.Y.Y., and Y.P. provided critical revisions. All authors approved the final version of the manuscript for submission.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pan, Y., Yang, H., Li, M. et al. Grouping strategies in numerosity perception between intrinsic and extrinsic grouping cues. Sci Rep 11, 17605 (2021). https://doi.org/10.1038/s41598-021-96944-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-96944-x
This article is cited by
-
Unimpaired groupitizing in children and adolescents with dyscalculia
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
