
Abstract
Site productivity remains a fundamental concern in forestry as a significant driver of resource availability for tree growth. The site index (SI) reflects the overall impact of all environmental factors that determine tree height growth and is the most commonly used indirect proxy for forest site productivity estimated using stand age and height. The SI concept challenges are local variations in climate, soil, and genotype-environmental interactions that lead to variable height growth patterns among ecoregions and cause inappropriate estimation of site productivity. Developing regional models allow us to determine forest growth and SI more appropriately. This study aimed to develop height growth models for the Scots pine in Poland, considering the natural forest region effect. For height growth modelling, we used the growth trajectory data of 855 sample trees, representing the Scots pine entire range of geographic locations and site conditions in Poland. We compared the development of regional height growth models using nonlinear-fixed-effects (NFE) and nonlinear-mixed-effects (NME) modelling approaches. Our results indicate a slightly better fit to the data of the model built using NFE approach. The results showed significant differences between Scots pine growth in natural forest regions I, II, and III located in northern Poland and natural forest regions IV, V, and VI in southern Poland. We compared the development of regional height growth models using NFE and NME modelling approaches. Our results indicate a slightly better fit to the data of the model built using the NFE approach. The developed models show differences in height growth patterns of Scots pines in Poland and revealed that acknowledgement of region as the independent variable could improve the growth prediction and quality of the SI estimation. Differences in climate and soil conditions that distinguish natural forest regions affect Scots pine height growth patterns. Therefore, extending this research to models that directly describe height growth interactions with site variables, such as climate, soil properties, and topography, can provide valuable forest management information.
Similar content being viewed by others

Introduction
Site productivity remains a fundamental concern in forestry as a significant driver of resource availability1. Appropriate estimation of site productivity is critical for forest management decisions2. Additionally, information on site productivity helps monitor and assess climate change impact on forest ecosystems and provide information to support forest management to take effective adaptation measures3. The site index (SI) reflects the overall impact of all environmental factors that determine tree height growth4 and is the most commonly used indirect proxy for forest site productivity estimated using stand age and height2,5,6,7,8,9. An appropriate height growth model is needed to determine the SI accurately.
One of the most critical challenges in the SI concept is a local variation of height growth patterns that result from a variety of factors and could cause the under- or over-estimation of the growth potential and productivity of a site10,11. Species variability caused by sensitivity to environmental factors is the primary reason why a more flexible approach to developing height growth models is recommended to include the impact of local conditions and to accurately reflect height growth variation on a regional scale12.
Climate, soil, and genotype-environmental interactions can favour species adaptation to local ecological conditions, causing variable height growth patterns for the same species among ecoregions10,11. In a study conducted in Sweden, Johansson13 found significant variation in forest height growth curves due to mineral soil type and geographical location. The effect of genetic variability on the height development of trees was reported by Adams et al.14. Genetic variability has also been identified as the primary factor modifying the trajectory of height growth by Buford and Burkhart15, who found that the height growth models developed for different provenances differed in their parameters describing the asymptote. Significant differences in height growth patterns can also occur for the same species, even within a given SI class16. García17 reported that three sources of height growth development variability could be identified: (a) between sites, (b) within sites, and (c) observation error. Because of the inter-regional variability of growth patterns, regional height growth models (RMs) are derived to improve the model's predictive ability11,12,18,19,20. Studies on Pinus pinaster in Spain indicated significant differences in growth patterns observed in different areas because of the effect of genotype-environment interaction11. Research describing regional height growth differences highlights the regional application of the SI concept18,21,22,23,24. The role of regional models may be crucial for sustainable forest management, particularly in solving problems that require accurate information on local growing conditions, where climate and soil type lead to inter-regional variation20.
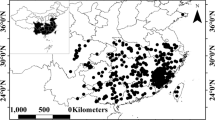
The Scots pine is one of the most important species in Europe and ranges from the boreal region in Northern and Eastern Europe to the mountains of the Mediterranean in Southeastern Europe (Fig. 1). It is also the most important tree species in Poland, covering approximately 58% of the total forested area of the country25. The Scots pine grows under very variable ecological conditions26, which could affect the difference in the shape of the growth curves among ecoregions13. The effective management of such a considerable forest resource requires a thorough assessment of the productivity of stands at a regional level27. Growth modelling is crucial in assessing the consequences of forest management activities in long term planning in forestry28. The use of inappropriate height growth models may result in erroneous estimation of site productivity. This problem could be solved by the development of models, which reflect the difference in growth trajectories resulting from specific growth conditions in a given area. Regional models reflect the impact of regional variability on height growth dynamics resulting from specific climate conditions, soil properties, and differences in local forest management.
In Poland, yield tables with discrete scales are still used in forest management for forest growth and site productivity estimation29,30,31. The forestry practice reports the incorrect determination of site productivity and incremented volume using yield tables, probably because of the inadequacy of Schwappach's tables from differences in growth conditions, which have changed considerably during the 100 years since the yield tables were developed. Pretzsch et al.32 estimated that in Central Europe, site productivity increased by approximately 30–70%. Additionally, the height growth of observed stands differs significantly from that observed 100 years ago33,34 and included in the original tables. Moreover, the tables produce errors by not considering regional growth variability on a countrywide scale.

Natural forest regions in Poland with locations from which stem analysis data were collected (a); the range of Scots pine distribution in Europe (b). The map was generated using ArcGIS software35 (ArcGIS Pro, Version 2.2.0, https://www.esri.com/en-us/arcgis/products/arcgis-pro/overview). The Scots pine range shapefile published by Caudullo et al.36 under the CC BY 4.0 open-access license was the basis for the mapping.
The research adopted the hypothesis that height growth is regionally differentiated. We assumed that height growth patterns are influenced by the variability in site conditions, which can be acknowledged by considering the specific effect of natural forest regions (NFE), distinguished by environmental conditions. Therefore, the research objective was to analyse the variability of height growth patterns between natural forest regions in Poland and develop height growth models for the Scots pine in Poland taking into account the specific effect of the region.
Materials and methods
Collection of the research material
The research material consists of the height growth trajectories of 855 dominant and codominant Scots pines (Table 1). The most substantial part of growth trajectories was collected from the sample plots established within the project "Actual and potential site productivity in Poland for main forest-forming tree species" from 2015 to 2016 and representing the whole variability of growth conditions in Poland for the Scots pine trees. Other tree growth trajectories have been collected in Poland over the last 2 decades as part of research projects conducted by the Department of Forest Resources Management, University of Agriculture, Krakow, the Forest Research Institute in Sękocin Stary, and the Faculty of Forestry, Warsaw University of Life Sciences. The final set of sample plots represent the area of Poland, with a whole range of geographic distributions and site conditions (Fig. 1). The age of the analysed sample trees ranged from 12 to 175 years, with an average of 76 years and heights ranging from 3.50 to 36.75 m (Table 1).
Natural forest regions37 were adopted as the regionalisation unit in Poland. Natural forest regions were distinguished by the natural diversity of the country, in particular: variability of climatic and geological conditions, naturally occurring ranges of principal forest-forming tree species, presence and distribution of natural landscapes classes, and distribution of the primary units of potential natural vegetation37.
Stem analysis
Stem analysis was used to reconstruct the past height growth of the trees. The trees for stem analysis were selected in the direct neighbourhood of the permanent sample plots established in Scots pine stands. For each plot, tree diameter and height close to the top height (TH)—defined as the average height of the 100 thickest trees per 1 ha—were selected and cut for stem analysis (SA). Only trees with typically developed crowns and no visible damage or growth anomalies were selected for SA. After cutting, the total lengths of the trees were measured, and discs were collected for SA. Discs were taken from the base of the felled tree at the height of 0.5 m, and the breast height diameter (DBH) was measured; for trees over 15 m high, subsequent discs were collected at heights of 2.0, 4.0, and successive 2-m intervals to the top of the tree. For trees shorter than 15 m, subsequent discs were collected at heights of 1.5, 2.5, 3.5, and at 1-m intervals further up the tree. The course of growth of the individual trees was reconstructed based on the height of the discs and the number of annual discs. The growth trajectories of all collected trees were visually examined for suppression and release patterns and growth anomalies. As cross-section lengths do not coincide with periodic height growth, the height-age data from the SA were corrected using Carmean's method38. The collected growth series for six natural forest regions were used to develop regional height growth models for Scots pine.
Models and parameters estimation
To develop regional height growth models: nonlinear fixed-effects and nonlinear mixed-effects modelling approaches were used.
A nonlinear mixed-effects modelling approach
The growth series are characterised by a hierarchical structure, which contains information about individual trees and natural forest regions. The mixed-effects modelling approach is a possible solution to handle such data39,40,41,42. To develop the height growth model using a mixed-effects modelling approach, we tested three functions well known in forest growth modelling: Chapman–Richards, Schumacher and Hossfeld:
where H is the top height (m), t is an age, and a1-a3 are estimated parameters.
Given the value of the mean absolute error (MAE), adjusted coefficient of determination (\({\text{R}}_{{{\text{adj}}}}^{2}\)), and the Akaike information criterion (AIC), the possibility of defining different random parameters was achieved, with a maximum of all (three) parameters found in the tested functions on both: trees and natural forest regions as grouping levels. In the case of heteroscedastic data, analysing likelihood ratio test and solutions containing power-type variance function with age as a predictor were applied43. Furthermore, given within-tree autocorrelation, the autoregressive correlation structure of order 1—AR(1)44 and autoregressive–moving average models for the within-group errors was analysed using corAR1 and corARMA(10,0) function from nlme R package respectively41 with RStudio software45.
Development of the models using nonlinear fixed-effect modelling approach with the functions developed using the algebraic difference approach
The algebraic difference approach (ADA) formulated by Bailey and Clutter46 allows for the derivation of models in which the SI depends only on one parameter. Therefore it allows obtaining polymorphic models with single asymptotes or anamorphic models with multiple asymptotes. The generalised algebraic difference approach (GADA) developed by Cieszewski and Bailey47 allows the use of multiple site parameters and models characterised by polymorphism and variable asymptotes for different sites. Height growth models should also show equality of the SI and height at base age and use the same function as the height growth and SI models.
After initial preselection, three ADA models (M2, M4 and M5) and two GADA models (M1, M3) were chosen for further analysis (Table 2). These models meet the criteria mentioned above and have been successfully applied in SI modelling, making them potential candidates for reliable SI models.
Local (site-specific) parameters of individual growth series and global parameters of SI models (M1-M5) were simultaneously fitted using the dummy variable approach (DVA), also known as the varying parameter method53. In the DVA, global and site-specific parameters are simultaneously estimated, resulting in the same parameter estimates regardless of the selected base age; therefore, it is a base-age invariant method and produces unbiased estimates for base-age invariant equation54. The DVA uses a starting value of height as the site-specific parameter for each growth series, assuming all particular growth series measurements have the same starting value. The dynamic model takes the following form Eq. (4).
where H is tree height, αSIi is the site-specific parameter for tree i, whose starting value for fitting is the observed SI value at the specified base age, di is a dummy variable (being 1 for tree i or 0 for other trees), b is a vector for global parameters, Tj is any age, T0 is an initial condition, and εij is the error term. As a result of parameter estimation, an expression containing simulated variables and a site-specific parameter allows the calculation of a unique site parameter for each tree. Due to the longitudinal nature of growth series data, subsequent height observations from the same growth trajectory are significantly correlated, and the assumption of independent errors may be violated. Therefore, the error structured modelling approach with a linear k-order autoregressive error was used in parameter estimation (Eq. 5).
where εij is the error observed at the jth measurement on ith tree, φk—are the k parameters to be estimated for the autoregressive process of order k, rk is a dummy variable equal to 1 when j > k and equal to zero, when j < k, and uij is random noise.
The quality of the fit was evaluated using MAE, \({\text{R}}_{{{\text{adj}}}}^{2}\), and AIC. The criteria mentioned above, and the evaluation of the residuals homoscedasticity, which was carried out based on the distribution of residual values against the predicted values, were used to select the function best fitted to empirical data.
The potential improvement of height growth prediction was analysed by expanding the height growth model parameters as a function of regions. For the calculation of the full model, taking into account the specific effect of region, the vector of global ith model parameters (bi) was expanded by the expansion term (δiRj), which is specific for the given jth region (Rj) and ith global parameter (i). Six dummy variables (Rj) representing regions were used to simultaneous fitting the expansion terms specific for each ith parameter (bi) and jth region.
The significance of the expansion terms was tested using t-statistics and was assumed as a statistical test for the difference between a given global parameter bi and the regional parameter values. Regional parameter values are the sum of the global parameters bi and expansion term (δiRj). Moreover, to compare the fits of the full (regional) and reduced (global) model, we used the anova() function with the regression objects as two separate arguments41. The anova() function takes model objects as arguments and returns an ANOVA to check that a more complex model is much better at capturing data than a simpler model.
The full model trajectories were directly compared by plotting the differences between observed and model trajectories. Parameter estimation was carried out in R using the procedures and suitably defined model form45.
Results
Nonlinear mixed-effects models
The range (0.359–0.710, Table 3) of the mean absolute error indicates that the application for the mixed-effects modelling approach when elaborating the height growth models allows obtaining high accuracy. The obtained results indicate that a better fit characterises the functions with defined two random parameters than one random parameter. The Chapman–Richards function with parameters a and b, which refer respectively to the asymptote and the growth rate, defined as random effects, was characterised by the best fit (Table 3). Note the shape of the function is expressed as a parameter c is estimated as a fixed-effect.
Moreover, analysis of the residuals for the best fitted Chapman-Richards model (estimated parameters shown in Table 4) indicated heteroscedasticity (Fig. 4). The assumed variance function did not model heteroscedasticity well. Likewise, the likelihood test indicated that the variance function has a nonsignificant influence on model fit. Alike autoregressive correlation structure of order 1—AR(1) autoregressive–moving average models for the within-group errors ARMA(10,0) has significant influence on model fit (p < 0.0001 for likelihood test). Whereas ARMA(10,0) solution allows to reach the best goodness-of-fit measures (Table 4).
Nonlinear fixed-effects models
All preselected functions showed a good fit to the data and explained (in most cases) close to 99% (adjusted R2) of the height growth variation (Table 5). The error structured modelling with the 10-order autoregressive error was used in parameter estimation. All developed models prediction errors were relatively low, with root mean absolute error (MAE) varying from 0.59 to 0.69 m.
Considering the largest part of the explained variance, the smallest average absolute error and the AIC value, the function M1 was selected as best fitted to the data. Next function M1 was used to develop the model taking into account the regional variability. For fitting of the full model, taking into account the specific effect of region, the parameters of the model M1 were expanded as a function of regions (Eq. 6).
where \(R = Z_{0} + \left( {Z_{0}^{2} + \frac{{2\left( {b_{2} + \delta_{{2R_{j} }} \times R_{j} } \right)H_{0} }}{{T_{0}^{{\left( {b_{1} + \delta_{{1R_{j} }} \times Rj} \right)}} }}} \right)^{0.5}\); \(Z_{0} = H_{0} + \left( {b_{3} + \delta_{{3R_{j} }} \times R_{j} } \right)\).
The extended full (regional) model explains 0.47% more residual variance, while all expansion terms are statistically significant (t < 0.001, Table 6). Besides, the ANOVA result indicates that more complex, the regional model is significantly better (p < 0.01) than the reduced (global) model and thus favour the regional model.
The residuals of the developed regional growth model are homoscedastic in the entire range of values predicted for individual trees and in individual regions (Fig. 2).

Relationship between residual and predicted height using the site index model for Scots pine in Poland developed using nonlinear fixed-effect modelling approach with the parameters expanded as a function of regions (I–VI).
The developed regional model does not cause systematic errors in SI classes, age classes and tree height classes (Fig. 3). The mean residuals slightly deviate from 0 only in the least common SI below 15 m, and the equally rare age class above 120 years and the height class above 35 m (Fig. 3).

Mean values of the residuals of the regional nonlinear fixed-effect model in site index, age and height classes.
Comparison of regional models developed with the use of nonlinear mixed-effects and nonlinear fixed-effects modelling approach
Based on the model fit statistics and graphical error analysis, it can be concluded that the regional model developed using the fixed-effects approach is characterised by better fit statistics (Table 7) and the distribution of errors (Fig. 4).

Plot of residuals vs predicted value of height for regional models developed using mixed-effects (left), and fixed-effects (right) modelling approaches.
Analysis of regional variability of height growth patterns based on a model developed using the fixed-effects modelling approach
The height growth patterns in individual regions show that it changes directionally from the north and northwest to south and southeast Poland (Fig. 5). In the NFR located in the northwestern (I, III) and northern (II) lowlands of Poland, the height increment rate is also maintained in older stands. In the case of NFR located in the highlands of southern and southeastern Poland, the height increment in old age is inhibited, and therefore the growth curves are more asymptotic (Fig. 5).

Regional nonlinear fixed-effects model height trajectories against the observed growth series from stem analysis collected in natural forest regions I-VI.
Growth trajectories of models developed for individual regions were directly compared using the height at base age 40 as the reference level (Fig. 6). A direct comparison of the height growth trajectories for different SI values determined for the base age of 40 years shows a significant diversification of the height growth patterns between regions, especially in more fertile sites. Up to the base age of 40, the height growth in individual regions is very similar. However, above the base age, a considerable diversification of the growth pattern is observed, especially for most fertile sites with SI equal to 23 or 17 m. Intensive height increment persists for the longest in older stands in the lowlands of northwest and northern Poland, especially in NFR I and III, which are under the influence of the Atlantic climate. Scots pine NFR II located in northeastern Poland grows only slightly more slowly. In the stands growing in regions VI and IV located in the highlands of southeastern Poland, where the climate is continental, the growth is slowed down in older stands. Scots pines from the NRF V located in southern Poland lowlands show weaker growth in old age in the most fertile sites, but in the poorer, their growth dynamics persist until old age (Fig. 6).

Top height trajectories estimated from the regional fixed-effects model for Scots pines in six natural forest regions (base age 40 years).
Discussion
We compared the development of regional height growth models using nonlinear fixed-effects and nonlinear mixed-effects modelling approaches. Our results indicate a better fit to the data of the model developed using the nonlinear fixed-effects modelling approach. Comparison of fixed-effects and mixed-effects models was conducted by Nigh40 who also found a better fit of models built with GADA. Similar results were also obtained by Cieszewski55, who demonstrated that self-referencing GADA models based on the nonlinear fixed-effects approach possess all the desirable properties associated with logical behaviour of the model and estimation statistics, while the nonlinear mixed-effects models lack the well-behaved model properties. The reason for the better fit of the models may be that the nonlinear fixed-effect models ar by definition more flexible than the nonlinear mixed-effect models55. Moreover, models can significantly overestimate variances based on untenable assumptions and have more restrictions imposed regarding the site effects' distributional properties. Cieszewski55 points out that this is because the NME model fitting process imposes an additional constraint in that the random effect must conform to the arbitrary assumption of randomness, meaning that it must always have a greater or equal sum of square errors when the same models are fit into the same dataset as site models. Our results also confirm well-behaved properties of the nonlinear fixed-effects self-referencing models and their better suitability for modelling TH growth and SI compared to mixed-effects models.
The developed models allowed us to compare the differences in Scots pine height growth dynamics between natural forest regions in Poland. We found that height growth is regionally differentiated. The most relevant differences in growth patterns are observed from the northwest to the southeast gradient. The shape of the growth trajectory for stands located in natural forest regions from the north and northwestern Poland (I–III) differs for younger and older trees. In these natural forest regions, the stands also reach higher absolute heights than those of southern regions. Our results show that the use of the regional model Poland reduces errors in the height growth prediction.
Regional variation in growth patterns was previously reported; these variations have usually been interpreted as the result of variability in climate, geology, soil type, geographical location, and genotype13,14,15.
We found significant differences between the height growth patterns from NRF located in lowlands of northern- northwestern and northeastern Poland and highlands of southern- southeastern Poland. This may be because natural forest regions in northern Poland are under the influence of Atlantic climate and have deep podsolic, brown podsolic, and brown glacial soils, whereas natural forest regions VI and IV are under the influence of continental climate and grow on upland soils with less depth. The depth and physical properties of soil and the sand and clay contents lead to varying soil fertility and productivity56,57. Additionally, the availability of water is strongly correlated with the growth of stands and impacts the productivity of sites58. A linear increase in net primary production with increased water availability was observed at dry sites59. The amount of annual precipitation in Poland changes from south to north and, with soil fertility and depth, can explain the difference in height growth variability between the northern and southern regions.
We found that growth trajectories for individual regions vary with age. The growth of stands in southern and southeastern Poland is slowing rapidly, whereas northern and northeastern stands have more continuous rapid height growth. The difference in growth pattern in regions IV, V, and VI observed in older age can be explained as the impact of soil depth. When soil depth is not a limiting factor in the youngest age classes, the impacts of climate and other factors are more important for growth dynamics, but with the transition to older age classes and increased interspecific competition, soil depth likely begins to limit the growth dynamic. An evident slowdown of growth dynamics in oldest age classes in natural forest regions IV and VI may be the result of a combination of shallower and less fertile podsolic and brown podsolic soils and low annual rainfall. In regions, I, II, and III, geological and climatic conditions are more favourable for Scots pines and likely result in a larger height increment in the older stands.
The significant height growth differences in different regions indicate that the use of one generalised height growth model for the entire country may result in the inappropriate estimation of site productivity and have direct forest management consequences. The importance of regional models may increase because the projections of climate change in Poland predict accentuated differences in site conditions between the northwest and southeast regions60. The obtained results concerning the spatial variability of Scots pines' height growth dynamics should be considered when making forest management decisions concerning the rotation age or intensity of silvicultural treatments.
The amount of carbon stored in forests is more significant than in any other terrestrial ecosystem61, and tree growth data play a significant role in carbon sequestration and biomass production calculations62. Several conversion methods have been developed for different types of forest to obtain reliable estimates63,64,65. However, our results show that not considering specific regional growth conditions and the resulting differentiated growth patterns by using only global models for these calculations may result in an over- or under-estimation of regional height growth, biomass production, and carbon sequestration. Many studies on the estimation of biomass production and carbon sequestration emphasise that the regional approach is promising66,67; therefore, the use of models developed on a regional scale shows particular importance for effective forest management.
With the development of geographical information systems and technologies, tools such as the mapping SI are increasingly common68. Many problems regarding the stability of forest stands and disturbance of forests in Europe concern relatively small areas. Numerous studies found that SI mapping and analyses, (particularly on a small scale) provide information for a more dynamic forest growth assessment and should be an essential component of regional planning and management69,70. Therefore the use of advanced forestry solutions focused on smaller areas, regional growth models, and the analysis of local trends have great potential for the fast detection of problems and the precise formulation of answers to stand instability and forest disturbances.
We used the height growth trajectories of dominant Scots pines collected using the unified methodology to develop a regional model; however, collection methods and data processing in different regions should be carefully executed. Differences in methodology or data sources can lead to significant inconsistencies between growth models that are not based on ecological or biological causes18. A given species growth patterns in different ecological or geographical conditions are significant and should be determined before developing local models for small areas. The differences in local model predictions may not be large enough in small geographical regions to justify the development costs18. In this situation, smaller regions with low climatic and geomorphological variability should be grouped to develop a single model for larger areas.
Conclusions
We compared the development of regional height growth models using nonlinear-fixed-effects and nonlinear-mixed-effects modelling approaches. Our results indicate a slightly better fit to the data of the model using the nonlinear fixed-effects approach. The developed models show differences in height growth patterns of Scots pines in Poland and revealed that acknowledgement of region as the independent variable could improve the growth prediction and quality of the SI estimation. The presented study showed regional differences in height growth patterns of Scots pines in Poland. Differences in climate and soil conditions that distinguish natural forest regions affect Scots pine height growth patterns. Therefore, extending this research to models that directly describe height growth interactions with site variables, such as climate, soil properties, and topography, can provide valuable forest management information.
Data availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Bontemps, J. D. & Bouriaud, O. Predictive approaches to forest site productivity: Recent trends, challenges and future perspectives. Forestry 87, 109–128 (2014).
Skovsgaard, J. P. & Vanclay, J. K. Forest site productivity: a review of the evolution of dendrometric concepts for even-aged stands. Forestry 81, 13–31 (2008).
Albert, M. & Schmidt, M. Climate-sensitive modelling of site-productivity relationships for Norway spruce (Picea abies (L.) Karst) and common beech (Fagus sylvatica L.). For. Ecol. Manag. 259, 739–749 (2010).
Véga, C. & St-Onge, B. Mapping site index and age by linking a time series of canopy height models with growth curves. For. Ecol. Manag. 257, 951–959 (2009).
Hägglund, B. & Lundmark, J. E. Site index estimation by means of site properties of Scots pine and Norway spruce in Sweden. Stud. For. Suec. 138, 5–38 (1977).
Johansson, T. Site index curves for common alder and grey alder growing on different types of forest soil in Sweden. Scand. J. For. Res. 14, 441–453 (1999).
Raulier, F., Lambert, M.-C., Pothier, D. & Ung, C.-H. Impact of dominant tree dynamics on site index curves. For. Ecol. Manag. 184, 65–78 (2003).
Corral Rivas, J. J., Álvarez González, J. G., Ruíz González, A. D. & von Gadow, K. Compatible height and site index models for five pine species in El Salto, Durango (Mexico). For. Ecol. Manag. 201, 145–160 (2004).
Tewari, V. P., Rivas, J. J. C., VilČko, F. & Von Gadow, K. Height-Growth and site index equations for social forestry plantations of acacia nilotica and eucalyptus hybrid in gujarat state of India. For. Trees Livelihoods 17, 125–140 (2007).
Monserud, R. A. & Rehfeldt, G. E. Genetic and environmental components of variation of site index in inland douglas-fir. For. Sci. 36, 1–9 (1990).
Alvarez-González, J. G., Ruiz-González, A. D., Rodríguez-Soalleiro, R. & Barrio-Anta, M. Ecorregional site index models for Pinus pinaster in Galicia (northwestern Spain). Ann. For. Sci. 62, 115–127 (2005).
Bravo-Oviedo, A., Tomé, M., Bravo, F., Montero, G. & del Río, M. Dominant height growth equations including site attributes in the generalised algebraic difference approach. Can. J. For. Res. 38, 2348–2358 (2008).
Johansson, T. Site index curves for Norway spruce plantations on farmland with different soil types. Stud. For. Suec. 198, 1–19 (1995).
Adams, J. P., Matney, T. G., Land, S. B. Jr., Belli, K. L. & Duzan, H. W. Jr. Incorporating genetic parameters into a loblolly pine growth-and-yield model. Can. J. For. Res. 36, 1959–1967 (2006).
Buford, M. A. & Burkhart, H. E. Genetic improvement effects on growth and yield of loblolly pine plantations. For. Sci. 33, 707–724 (1987).
Monserud, R. A. Height growth and site index curves for inland Douglas-fir based on stem analysis data and forest habitat type. For. Sci. 30, 943–965 (1984).
García, O. Dynamical implications of the variability representation in site-index modelling. Eur. J. For. Res. 130, 671–675 (2010).
Calama, R., Cañadas, N. & Montero, G. Inter-regional variability in site index models for even-aged stands of stone pine (Pinus pinea L.) in Spain. Ann. For. Sci. 60, 259–269 (2003).
Adame, P., Hynynen, J., Cañellas, I. & del Río, M. Individual-tree diameter growth model for rebollo oak (Quercus pyrenaica Willd.) coppices. For. Ecol. Manag. 255, 1011–1022 (2008).
Bravo-Oviedo, A., del Río, M. & Montero, G. Geographic variation and parameter assessment in generalised algebraic difference site index modelling. For. Ecol. Manag. 247, 107–119 (2007).
Bontemps, J. & Bouriaud, O. Predictive approaches to forest site productivity: recent trends, challenges and future perspectives. Forestry 87, 1–20 (2013).
Claessens, H., Pauwels, D., Thibaut, A. & Rondeux, J. Site index curves and autecology of ash, sycamore and cherry in Wallonia (Southern Belgium). Forestry 72, 171–182 (1999).
Karlsson, K. Height growth patterns of Scots pine and Norway spruce in the coastal areas of western Finland. For. Ecol. Manag. 135, 205–216 (2000).
Martín-Benito, D., Gea-Izquierdo, G., del Río, M. & Cañellas, I. Long-term trends in dominant-height growth of black pine using dynamic models. For. Ecol. Manag. 256, 1230–1238 (2008).
Dyrekcja Generalna Lasów Państwowych, Raport o stanie lasów w Polsce (2018).
Socha, J. Long-term effect of wetland drainage on the productivity of Scots pine stands in Poland. For. Ecol. Manag. 274, 172–180 (2012).
Bravo, F. & Montero, G. Site index estimation in Scots pine (Pinus sylvestris L.) stands in the High Ebro Basin (northern Spain) using soil attributes. Forestry 74, 395–406 (2001).
Gadow, K.; Hui, G. Modelling Forest Development, Vol. 57, Forestry Sciences (Springer Netherlands, Dordrecht, 1999). ISBN 978–1–4020–0276–2.
Schwappach, A. Ertragstafeln der Wichtigeren Holzarten (Druckerei Merkur, Prag, 1943).
Szymkiewicz, B. Niektóre zagadnienia dotyczące tablic zasobności drzewostanów sosnowych. Pr. IBL Seria A 67 (1948)
Szymkiewicz, B. Tablice Zasobności i Przyrostu Drzewostanów (Państwowe Wydawnictwo Rolnicze i Leśne, Warszawa, 2001). ISBN 83–09–01745–6.
Pretzsch, H., Biber, P., Schütze, G., Uhl, E. & Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 5, 4967 (2014).
Socha, J. & Orzeł, S. Dynamic site index curves for Scots pine (Pinus sylvestris L.) in southern Poland. Sylwan 157, 26–38 (2013).
Socha, J., Tymińska-Czabańska, L., Grabska, E. & Orzeł, S. Site index models for main forest-forming tree species in Poland. Forests 11, 8–10 (2020).
Esri Inc. ArcGIS Pro (Version 2.2.0). Esri Inc. https://www.esri.com/en-us/arcgis/products/arcgis-pro/overview (2020).
Caudullo, G., Welk, E. & San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Br. 12, 662–666 (2017).
Zielony, R., Kliczkowska, A. Regionalizacja Przyrodniczo-Leśna Polski 2010 (2012). ISBN 9788361633624.
Carmean, W. H. Site index curves for upland oaks in the central states. For. Sci. 18, 109–120 (1972).
Mehtätalo, L. & Lappi, J. Biometry for Forestry and Environmental Data: with Examples in R 1st edn. (Chapman and Hall/CRC, Boca Raton, 2020).
Nigh, G. Engelmann spruce site index models: a comparison of model functions and parameterisations. PLoS ONE 10, e0124079 (2015).
Pinheiro J., Bates D., & DebRoy S. S. D. nlme: Linear and Nonlinear Mixed Effects Models (2020).
Wang, M., Borders, B. E. & Zhao, D. An empirical comparison of two subject-specific approaches to dominant heights modeling: the dummy variable method and the mixed model method. For. Ecol. Manag. 255, 2659–2669 (2008).
Bronisz, K. & Mehtätalo, L. Mixed-effects generalised height–diameter model for young silver birch stands on post-agricultural lands. For. Ecol. Manage. 460, 117901 (2020).
Wang, M., Bhatti, J., Wang, Y. & Varem-Sanders, T. Examining the gain in model prediction accuracy using serial autocorrelation for dominant height prediction. For. Sci. 57, 241–251 (2011).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/ (2020).
Bailey, R. L. & Clutter, J. L. Base-age invariant polymorphic site curves. For. Sci. 20, 155–159 (1974).
Cieszewski, J. & Bailey, L. Generalized algebraic difference approach: theory based derivation of dynamic site equations with polymorphism and variable asymptotes. For. Sci. 46, 116–126 (2000).
Cieszewski, C. J. Three methods of deriving advanced dynamic site equations demonstrated on inland Douglas-fir site curves. Can. J. For. Res. For. 31, 165–173 (2001).
Krumland, B. & Eng, H. Site index systems for major young-growth forest and Woodland species in North California. Calif. For. 4, 1–220 (2005).
Cieszewski, C. J. Developing a well-behaved dynamic site equation using a modified hossfeld IV function Y 3 = (axm)/(c + x m–1), a simplified mixed-model and scant subalpine fir data. For. Sci. 49, 539–554 (2003).
Sharma, R. P., Brunner, A., Eid, T. & Øyen, B.-H. Modelling dominant height growth from national forest inventory individual tree data with short time series and large age errors. For. Ecol. Manag. 262, 2162–2175 (2011).
Anta, M. B. et al. Development of a basal area growth system for maritime pine in northwestern Spain using the generalised algebraic difference approach. Can. J. For. Res. 36, 1461–1474 (2006).
Cieszewski, C. J., Harrison, M. & Martin, S. W. Examples of practical methods for unbiased parameter estimation in self-referencing functions. In Proceedings of the First International Conference on Measurements and Quantitative Methods and Management and The 1999 Southern Mensur (ed. Cieszewski, C. J.) (2000).
Nunes, L., Patrício, M., Tomé, J. & Tomé, M. Modeling dominant height growth of maritime pine in Portugal using GADA methodology with parameters depending on soil and climate variables. Ann. For. Sci. 68, 311–323 (2011).
Cieszewski, C. J. Comparing properties of self-referencing models based on nonlinear-fixed-effects versus nonlinear-mixed-effects modeling approaches. Math. Comput. For. Nat. Sci. 10, 46–57 (2018).
Seynave, I. et al. Picea abies site index prediction by environmental factors and understorey vegetation: a two-scale approach based on survey databases. Can. J. For. Res. 35, 10 (2005).
Brandl, S. et al. Static site indices from different national forest inventories: harmonisation and prediction from site conditions. Ann. For. Sci. 75, 1–17 (2018).
Breda, N., Huc, R. & André Granier, E. D. Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 63, 625–644 (2006).
Loik, M. E., Breshears, D. D., Lauenroth, W. K. & Belnap, J. A multi-scale perspective of water pulses in dryland ecosystems: climatology and ecohydrology of the western USA. Oecologia 141, 269–281 (2004).
Kundzewicz, Z. W., Hov, Ø., Okruszko, T. Zmiany klimatu i ich wpływ na wybrane sektory w Polsce. 257 (2017).
Barrio Anta, M. & Dieguez-Aranda, U. Site quality of pedunculate oak (Quercus robur L.) stands in Galicia (northwest Spain). Eur. J. For. Res. 124, 19–28 (2005).
Bowman, D. M. J. S., Brienen, R. J. W., Gloor, E., Phillips, O. L. & Prior, L. D. Detecting trends in tree growth: not so simple. Trends Plant Sci. 18, 11–17 (2013).
Isaev, A., Korovin, G., Zamolodkchikov, D., Utkin, A. & Pryaznikov, A. Carbon stock and depostion in phytomass of the Russian forests 247–256 (Kluwer, Amsterdam, 1995).
Fang, J. Y. & Wang, Z. M. Forest biomass estimation at regional and global levels, with special reference to China’s forest biomass. Ecol. Res. 16, 587–592 (2001).
Schroeder, P., Brown, S., Mo, J., Birdsey, R. & Cieszewski, C. Biomass estimation for temperate broadleaf forests of the United States using inventory data. For. Sci. 43, 424–434 (1997).
Bravo-Oviedo, A., Gallardo-Andrés, C., del Río, M. & Montero, G. Regional changes of Pinus pinaster site index in Spain using a climate-based dominant height model. Can. J. For. Res. 40, 2036–2048 (2010).
Woodbury, P. B., Smith, J. E., Weinstein, D. & Laurence, J. Assessing potential climate change effects on loblolly pine growth: a probabilistic regional modeling approach. For. Ecol. Manag. 107, 99–116 (1998).
Latta, G., Temesgen, H. & Barrett, T. M. B. M. Mapping and imputing potential productivity of Pacific Northwest forests using climate variables. Can. J. For. Res. 39, 1197–1207 (2009).
Coops, N. C., Hember, R. & Waring, R. H. Assessing the impact of current and projected climates on Douglas-Fir productivity in British Columbia, Canada, using a process-based model (3-PG). Can. J. For. Res. 40, 511–524 (2010).
Coops, N. C., Coggins, S. B. & Kurz, W. a Mapping the environmental limitations to growth of coastal Douglas-fir stands on Vancouver Island, British Columbia. Tree Physiol. 27, 805–815 (2007).
Acknowledgements
We would like to thank the Forest Management and Geodesy Bureau from Sekocin Stary for cooperation in implementing the project "Actual and potential site productivity in Poland for the main forest forming tree species", particular for help in collecting the height growth data. We thank Professor Arkadiusz Bruchwald for sharing stem analysis data collected within his research projects.
Funding
This work was supported by the project I-MAESTRO. Project Innovative forest MAnagEment STrategies for a Resilient biOeconomy under climate change and disturbances (I-MAESTRO) is supported under the umbrella of ForestValue ERA-NET Cofund by the National Science Centre, Poland and French Ministry of Agriculture, Agrifood, and Forestry; French Ministry of Higher Education, Research and Innovation, German Federal Ministry of Food and Agriculture (BMEL) via Agency for Renewable Resources (FNR), Slovenian Ministry of Education, Science and Sport (MIZS). ForestValue has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement N° 773324. Data collection was supported by the project “Actual and potential site productivity in Poland for the main forest forming tree species”, financed by the General Directorate of State Forests [agreement no. ER-2717–11/14, 15th April 2014].
Author information
Authors and Affiliations
Contributions
Conceived and designed the study: J.S. Collected data: J.S. Analysed the data: J.S., K.B., L.T.C. Wrote the paper: J.S., L.T.C., K.B., S.Z., P.H. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Socha, J., Tymińska-Czabańska, L., Bronisz, K. et al. Regional height growth models for Scots pine in Poland. Sci Rep 11, 10330 (2021). https://doi.org/10.1038/s41598-021-89826-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-89826-9
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
