
Abstract
Iris germanica L. is a perennial herbaceous plant that has been widely cultivated worldwide and is popular for its elegant and vibrantly colorful flowers. Selection of appropriate reference genes is the prerequisite for accurate normalization of target gene expression by quantitative real-time PCR. However, to date, the most suitable reference genes for flowering stages have not been elucidated in I. germanica. In this study, eight candidate reference genes were examined for the normalization of RT-qPCR in three I. germanica cultivars, and their stability were evaluated by four different algorithms (GeNorm, NormFinder, BestKeeper, and Ref-finder). The results revealed that IgUBC and IgGAPDH were the most stable reference genes in ‘00246’ and ‘Elizabeth’, and IgTUB and IgUBC showed stable expression in ‘2010200’. IgUBC and IgGAPDH were the most stable in all samples, while IgUBQ showed the least stability. Finally, to validate the reliability of the selected reference genes, the expression patterns of IgFT (Flowering Locus T gene) was analyzed and emphasized the importance of appropriate reference gene selection. This work presented the first systematic study of reference genes selection during flower bud development and provided guidance to research of the molecular mechanisms of flowering stages in I. germanica.
Similar content being viewed by others


Introduction
Quantitative real-time PCR (RT-qPCR) is a reliable and widely used technique to quantify target gene expression patterns in various fields of biological research, due to its high sensitivity, accuracy and reproducibility1,2,3. However, the accuracy of RT-qPCR is influenced by various factors, including the quantity of mRNA templates, enzymatic efficiency in cDNA synthesis and PCR amplification4. Thus, to avoid bias, it is necessary to select reliable reference genes that are steadily expressed under different experimental conditions before determining the expression pattern of a target gene by RT-qPCR.
Traditional reference genes, such as elongation factor 1 alpha (EF1α), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), actin (ACT) and ubiquitin (UBQ), are mostly involved in intermediary metabolism or other basic cellular processes5,6,7, and are therefore commonly accepted for normalization without the need for any validation for their stability. However, numerous studies indicate that the expression levels of these genes vary considerably at different developmental stages or throughout the entire lifecycle of plants8,9. A systematic study for each tested species is recommended for identifying the best potential reference genes. Furthermore, several statistical algorithms, such as GeNorm10, NormFinder11 and BestKeeper12, have been developed for the evaluation of potential reference gene(s) in different experimental systems.
Flowers of higher plants are reproductive organs that are widely studied and are important ornamental characteristics of ornamental plants. A number of reference genes have been reported for different flowering stages of various ornamental plants species, such as chrysanthemum13, petunia14, azalea15 and tree peony16, but little information is available concerning reference genes for Iris. Iris germanica L., which is often referred to as Pogon iris, is one of the most important ornamental species in the Iris genus17, the posture of the flower is peculiar and the flowers are rich and have high ornamental and economic value. However, the spring-time flowering habit and short duration of flowering of I. germanica hinder its year-round supply and economic benefits. To enable additional detailed and in-depth studies of the expression level of key genes involved in flowering, it is necessary to identify the stability of candidate reference genes at various flowering stages in I. germanica. Recently, studies have been shown that ACT11 performed well in different tissues but poorly in different flower development stages in I. germanica cultivar ‘Huangjinjia’. Furthermore, TUA performed best in different flower development stages but was the worst in different tissues18. Nevertheless, limited information is available concerning reference genes for flowering stages in I. germanica.
In the present study, the expression stability of eight candidate reference genes, IgEF1α, IgGAPDH, IgACT6, IgUBQ, IgUBC (ubiquitin-protein ligase), IgEF1β (elongation factor 1β), IgPGK (phosphoglycerate kinase), IgTUB (beta-tubulin), was validated by RT-qPCR during flower development in different cultivars of I. germanica. Four statistical algorithms, GeNorm10, BestKeeper12, NormFinder11, and Ref-finder (http://www.leonxie.com/referencegene.php) were used to evaluate the most suitable reference genes needed for normalization. To verify the usefulness of the selected reference genes, the relative expression levels of IgFT, a putative homolog of Flowering Locus T (FT) gene in Arabidopsis, was analyzed during flower bud development and in different genotypes of I. germanica. This is the first report about the selection of reference genes during flower bud development and will benefit future studies on gene expression of flowering stages in I. germanica and other related species.
Results
Assessment of primer specificity and amplification efficiency of reference genes
The gene names, primer sequences and amplicon length characteristics of the 8 reference genes are summarized in Table 1. The primer specificities were verified by agarose gel electrophoresis, only a specific product of the expected size was observed, and no primer dimers were detected (Supplementary Fig. S1). In addition, only a single peak was found in the melting curves of the amplified products of all genes, indicating that no primer dimers were generated (Supplementary Fig. S2). The amplification efficiency (E) of each reference gene varied from 98.47% for IgGAPDH to 101.74% for IgUBQ, and the correlation coefficients (R2) ranged from 0.9984 to 0.9998 (Table 1). The amplicon size ranged from 107 for IgACT6 to 295 bp for IgUBQ.
Expression levels and profiles of reference genes
The quantification cycle (Cq) values of eight reference genes were assayed by RT-qPCR analysis, with lower Cq values representing relatively higher mRNA transcript levels. Cq values for each of the 8 candidate reference genes in three different genotypes of I. germanica are listed in supplementary Table S1. The average Cq values of these reference genes ranged from 19.83 to 27.61 for the highest and lowest expression levels, respectively, across all samples (Fig. 1). In addition, the standard deviations (SD) of Cq values were used to determine reference gene stability levels. Genes with relatively high SD of Cq values showed more variable expression than did those with lower SD. IgGAPDH showed the least variation in gene expression (23.69 ± 0.36) and presented the lowest SD, while IgEF1β showed the most variable levels of expression (23.16 ± 1.38).

Cq values of the 8 candidate reference genes in all samples of I. germanica. Line across the box-plot of Cq value depicts median values. The outside box is determined by the 25th and 75th percentiles. The whiskers represent the maximum and minimum values.
Expression stability of reference genes
To further select the most appropriate reference gene for RT-qPCR-based analysis in the investigation of flower development across three different I. germanica cultivars, four software programs, GeNorm, NormFinder, BestKeeper and RefFinder were used to analyze the expression stability of each reference gene.
GeNorm analysis
GeNorm program was used to evaluate the expression stability of the 8 candidate reference genes by calculating average expression stability (M) values based on the average pairwise variation among all the tested genes. According to GeNorm algorithm, stably expressed genes had M values below 1.5, and a relatively low M value indicates a relatively stable expression10. In this study, all of the tested genes showed high expression stability, with M-values of < 1.5, indicating that they all conformed to basic requirements for function as reference genes. IgUBC and IgGAPDH were the most stable reference genes in both ‘00246’ and ‘Elizabeth’, while IgTUB and IgGAPDH were identified as the most stable in ‘2010200’. In terms of the total samples tested, IgUBC and IgGAPDH were recommended as the most stable reference genes. In contrast, IgUBQ with the highest M value was identified as the least stable reference gene in all of the samples (Fig. 2).

Average expression stability values (M) of 8 candidate reference genes predicted by GeNorm software. The most stable genes are on the right, while the least stable genes are on the left. (A) ‘00246’, (B) ‘2010200’, (C) ‘Elizabeth’, and (D) all samples.
The optimal number of reference genes was also measured by determining the pairwise variation between sequentially ranked genes (Vn/Vn + 1) based on the GeNorm algorithm (Fig. 3). Generally speaking, a cutoff of 0.15 (Vn value) has been recommended as the threshold to determine the optimal number of reference genes10. Our results reveal that the V2/3 values of the ‘00246’, ‘2010200’ and ‘Elizabeth’ samples were lower than 0.15 (Fig. 3), suggesting that two reference genes were sufficient for accurate normalization. However, the value of 0.15 should not be a fixed threshold, and higher cutoff values of Vn/n + 1 have been shown in several reports19,20. Our data showed small variation between V2/3 and V3/4 across all the samples, suggesting that two reference genes were sufficient for normalization (Fig. 3), which was similar to results in bermudagrass21 and Kentucky bluegrass22.

Pairwise variation (V) of 8 candidate reference genes were calculated by GeNorm. Vn/Vn + 1 value were used to determine the optimal number of reference genes.
NormFinder analysis
NormFinder is used to determine the stability value of reference genes, based on inter- and intragroup variance in different sample groups11. The stability value is then calculated, with a relatively low stability value meaning that the gene is relatively stable. Reference gene stability values were calculated by NormFinder, as shown in Table 2. IgUBC and IgGAPDH were the two most stable genes among the total group, while IgUBQ was the least stable. The top two most stably expressed genes were IgUBC and IgGAPDH in both ‘00246’ and ‘2010200’, and IgUBC and IgTUB in ‘Elizabeth’. The ranking order generated by this method was slightly different from that of GeNorm.
BestKeeper analysis
BestKeeper evaluates the stability of reference genes based on the standard deviation (SD) and coefficients of variation (CV) of Cq values, with relatively low SD and CV representing relatively high stability12. The results of BestKeeper analysis are listed in Table 3. IgGAPDH and IgUBC were recommended as the most stable genes in ‘00246’, ‘Elizabeth’ and across all the samples, which was similar to the results from the GeNorm and NormFinder analysis. In ‘2010200’ samples, IgTUB and IgACT6 were detected as the most stable genes via BestKeeper analysis, whereas IgACT6 was ranked fourth by GeNorm and sixth by NormFinder.
RefFinder analysis
RefFinder (http://www.leonxie.com/referencegene.php) was used to obtain the comprehensive rankings of reference genes by integrating three common analysis programs: GeNorm, NormFinder and BestKeeper23. The final comprehensive rankings of the three algorithms were integrated by RefFinder and the results are shown in Table 4. Across all the samples, the ranking order was (from the most stable to the least stable) as follows: IgUBC > IgGAPDH > IgTUB > IgPGK > IgEF1ɑ > IgACT6 > IgEF1β > IgUBQ. This order is similar to the results of the GeNorm and NormFinder analysis. IgGAPDH and IgUBC were ranked as the most stable genes in ‘00246’ and ‘Elizabeth’, and IgTUB and IgUBC were the most stable genes in ‘2010200’. On the other hand, IgUBQ was the most unstable gene in all the samples, ‘00246’ and ‘2010200’, and IgEF1β was the least stable gene in ‘Elizabeth’. In the all samples, IgUBC and IgGAPDH were purported to be the most stable reference genes, while IgUBQ showed the least stability.
Validation of the selected reference genes
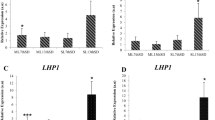
To validate the reliability of the reference genes, the relative expression patterns of IgFT were examined using different combinations of reference genes in the three cultivars. The two most stable reference genes (IgGAPDH and IgUBC for ‘00246’ and ‘Elizabeth’, IgTUB and IgUBC for ‘2010200’) and the least stable reference genes (IgUBQ for ‘00246’ and ‘2010200’, IgEF1β for ‘Elizabeth’) selected from the analyses described above were used either alone or in combination for RT-qPCR analyses. As shown in Fig. 4, although the overall relative expression patterns of IgFT showed similar trends, differences were found when the data were normalized to those of the different reference genes. When the least stable gene IgUBQ was used as the reference gene, the normalized expression levels of IgFT in ‘00246’ and ‘2010200’ significantly decreased compared with those normalized using IgGAPDH or IgUBC alone, the combination of IgGAPDH + IgUBC (for ‘00246’), IgTUB or IgUBC alone or the combination of IgTUB + IgUBC (for ‘2010200’) (Fig. 4A, B). However, when the least stable gene IgEF1β was used for normalization, the expression level of IgFT dramatically increased compared to that of IgGAPDH, IgUBC, or the combination of IgGAPDH + IgUBC in ‘Elizabeth’ (Fig. 4C). The combination of IgGAPDH/IgUBC was recommended as the optimum pair of reference genes for ‘00246’ and ‘Elizabeth’, and IgTUB/IgUBC was the best suited pair of reference genes for accurate normalization in ‘2010200’. IgUBC was the most suitable reference gene for three different I. germanica cultivars.

Relative quantification of IgFT gene expression in different cultivars of I. germanica using validated reference genes for normalization. (A) The expression level of IgFT in ‘00246’. (B) The expression level of IgFT in ‘2010200’. (C) The expression level of IgFT in ‘Elizabeth’. The bars represent standard errors. The relative expression level is reported as the mean of three biological replicates (n = 3), with bars indicating the standard deviation. Different letters indicate a statistically significant difference in each condition (P < 0.05, student’s t-test).
Discussion
Gene expression analyses are extremely important for revealing the molecular mechanisms that regulate important plant traits24,25. RT-qPCR has become the most powerful technique for quantification studies at the mRNA transcript level13. Selecting the appropriate reference genes is a necessary prerequisite for reliable RT-qPCR-based analysis. Ideally, an accurate reference gene should display stable expression in different tissues, in different organs, at different developmental stages and across different treatments26. In this study, we performed a systematic evaluation of 8 reference genes at different flowering stages and different genotypes of I. germanica cultivars. This study is the first attempt to identify the reference genes suitable for RT-qPCR normalization in flowering stages of I. germanica. The expression stability of various reference genes differed among the cultivars. Similar results were reported in tree peony, Panax ginseng and strawberry16,27,28. Different genetic backgrounds and biological processes between cultivars may have important effects on the expression stability of reference genes. Indeed, the selection and validation of reliable reference genes for quantitative analysis of gene expression analysis were necessary for the different cultivars.
Three programs GeNorm, NormFinder and BestKeeper, which are based on different algorithms and analytical procedures, are widely used to select the most reliable reference genes by researchers29,30. In this study, we found discrepancies in the reference gene stability rankings and validation data generated by the three different algorithms above. The rankings created by GeNorm and NormFinder were similar, but they showed quite distinct differences from the ranking obtained by BestKeeper. For instance, IgACT6 was ranked among the top two stable genes by BestKeeper in ‘2010200’ but was ranked in the middle or bottom portion by GeNorm and NormFinder. Moreover, across all the samples, IgACT6 was ranked among the top four stable genes by BestKeeper, whereas it was ranked seventh by GeNorm and NormFinder. These results are similar to those of many previous studies, possibly due to the different principles among the algorithms13,31. RefFinder, a comprehensive statistical program that integrates data from GeNorm, NormFinder, and BestKeeper, is used to evaluate the overall stability of reference gene expression32 Based on the comprehensive analysis by Ref-finder, IgGAPDH and IgUBC for ‘00246’ and ‘Elizabeth’ and IgTUB and IgUBC for ‘2010200’ were identified as the most stable reference genes for RT-qPCR of target gene expression studies. These results suggest that all 8 reference genes exhibited differential stability among the three cultivars.
In this study, we evaluated 8 genes that have been widely used as candidate reference genes in many species. The results indicate that it is better to select different reference genes according to different biological samples. Based on the results of our study involving different flower developmental stages, IgUBC, IgGAPDH and IgTUB were good candidates for normalization in all of the samples. Similar studies have also been conducted in other species, such as Rhododendrons33, Chinese cabbage34, Chrysanthemum lavandulifolium35 and Silybum marianum36. Moreover, UBQ was determined to be one of the most stable reference genes under NaCl and Pb stress in Iris. lactea var. chinensis37, but this gene was ranked as the least stable reference genes in both ‘00246’ and ‘2010200’ in our study, which was similar to the findings in flower buds of Iris bulleyana38 and Rhizophora apiculate39. EF1α and EF1β were determined to be the best suitable reference genes for all samples of various tissues in soybean40. In our study, these two genes ranked very low in all the samples, and IgEF1β exhibited the most unstable expression values in ‘Elizabeth’, which was similar to results in Moso bamboo41. ACT6 and PGK were determined to be the most stable reference genes for proper normalization in flower buds of Iris bulleyana38 and Chrysanthemum across ploidy levels42 and meiosis and somatic tissues of wheat43, while these two genes were found to be not well suited in our study, similar to reports in I. lactea var. chinensis37. TUB, a member of the Tubulin gene family, has also been widely used as a reliable reference gene in Primula forbesii44 and peach45. Similarly, in our study, IgTUB was determined as the most stable reference gene in the flowers of ‘2010200’. However, this gene ranked very low under all the tested conditions in Iris bulleyana38. GAPDH has been reported as the most stable reference gene under PEG and cold stress in I. lactea var. chinensis37, but it showed unstable expression under various environmental conditions in garlic plants46 and under PEG and NaCl treatments in Glehnia littoralis47. In our study, IgGAPDH was the most stable reference gene across all flower developmental stages in ‘00246’. In addition, IgUBC was ranked first in ‘Elizabeth’ and was also the most stable reference gene in all the samples, which was similar to the findings in all samples of I. lactea var. chinensis48. These results highlight the fact that the choice of reference genes for normalization should be specific. Even though the samples belong to the same type and are from the same species (but belong to different lines), they may have different sets of reference genes. Therefore, it is necessary to select and verify reliable reference genes for quantitative gene expression analysis, whether for different species or for different cultivars.
To illustrate the reliability of the reference genes, the expression levels of the IgFT gene were normalized by using the most stable or least stable reference genes. The results showed that the relative expression level of IgFT exhibited a clear pattern in all three cultivars when the stable reference genes IgGAPDH, IgUBC, and IgTUB or a combination of them were used (Fig. 4). The relative transcript abundance presented conflicting results when the least stable genes, IgUBQ or IgEF1β were used. Therefore, the selection of suitable internal control genes is critically important for normalization of target gene expression data by RT-qPCR.
In summary, the current study provides the first comprehensive analysis of the selection of stable reference genes as internal controls for RT-qPCR-based analysis of target gene expression in different flowering stages and different genotypes of I. germanica cultivars. IgGAPDH combined with IgUBC was recommended as the optimal reference gene in ‘00246’ and ‘Elizabeth’, while IgUBC/IgTUB was identified as the best combination for ‘2010200’. This research is the first report on the validation of candidate reference genes across flower developmental stages of three different I. germanica cultivars, which will provide basic data for research on the molecular mechanism underlying flower development in this species, and lays a foundation for similar studies in other related species.
Materials and methods
Plant material
Iris germanica L. materials were introduced from Xi'an, Beach, Xi'an, Shanxi Province (CHN, latitude 34° 15′ N, longitude 108° 56′ E) under the permission of competent authority and cultivated in the Iris Resource Collection Garden of Institute of Botany, Nanjing Sun Yat-Sen Memorial Botanical Garden (CHN, latitude 32° 01′ N, longitude 118° 48′ E). All experiments are carried out with relevant institutional, national, and international guidelines and legislation. Three different genotypes of I. germanica cultivars including ‘00246’ (an early-flowering cultivar), ‘2010200’ (an intermediate-flowering cultivar), and ‘Elizabeth’ (a late-flowering cultivar), were used as plant materials; these cultivars can be used to study the flowering gene expression of I. germanica in different flowering stages. For each cultivar, flower buds samples were collected from plants into four flowering stages: in flowering stage 1 (FS1), the size of flower bud is less than 1.00 mm in length, in flowering stage 2 (FS2), the size of flower bud is between 1.00 to 2.00 mm, in flowering stage 3 (FS3), the size of flower bud is between 2.00 to 3.00 mm, in flowering stage 4 (FS4), the size of flower bud is between 4.00 to 5.00 mm. The traits of three different genotypes of I. germanica cultivars were shown in Table S2. The samples were immediately frozen in liquid nitrogen and stored at − 80 ℃ until RNA extraction. The experiment included three biological replicates.
RNA isolation and cDNA synthesis
Total RNA was extracted using RNA simple Total Kit (TaKaRa Dalian, China) according to the manufacturer’s instructions. Total RNA was pretreated with RNase-free DNase I (TaKaRa, Dalian, China) at 37 ℃ for 30 min to eliminate potential DNA contamination. The integrity of the total RNA was assessed by 1.5% (w/v) agarose gel electrophoresis and the concentration of the samples was determined by a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, USA). Only RNA samples showing an A260/A280 ratio of 1.9–2.1 and A260/A230 ratio > 2.0 were used for subsequent analysis. For qPCR based analysis, first strand cDNA was synthesized using 1 μg total RNA in a volume of 20 μL with the PrimeScript RT Reagent Kit (TaKaRa, Dalian, China) according to the manufacturer’s instructions.
Selection of reference genes and PCR primer design
Eight candidate reference genes, IgEF1α, IgGAPDH, IgACT6, IgUBQ, IgUBC, IgEF1β, IgPGK and IgTUB were selected from homologs of traditional housekeeping genes previously used for flower development13,27. The putative homologs of 8 reference genes were identified from the transcriptomic data sequences of Iris lactea var. chinensis49. All the candidate reference genes were cloned in Stage 4 samples and confirmed through sequencing. Primers were designed using Primer 5.0 software (Premier Biosoft International) with melting temperatures (TM) of 55–65 ℃, primer lengths of 18–25 bp and amplicon lengths of 107–295 bp (Table 1). The performance of the primers was tested by qPCR and the specificity of the primer amplicons was further verified by 2% (w/v) gel electrophoresis.
Quantitative real-time PCR (RT-qPCR)
qPCR was performed using a Mastercycler ep realplex 2S device (Eppendorf, Germany) in conjunction with SYBR Premix Ex Taq II (TaKaRa, Dalian, China). Reactions were performed in a total volume of 20 μL containing 5 μL of diluted cDNA, 0.6 μL of each of forward and reverse primer (10 μM), 10 μL of 2 × SYBR Premix and 3.8 μL of ddH2O. The amplification program comprised an initial denaturation step (95℃ for 2 min), followed by 40 cycles of 95℃ for 5 s, 60℃ for 30 s, and 72 ℃ for 30 s, and a melting curve protocol (60–95 ℃ with a temperature increment of 0.5 ℃ s−1). Each RT-qPCR was performed for three biological and three technical replicates, and negative controls were included for each primer pair. Amplification efficiency (E) and correlation coefficient (R2) values were obtained from standard curves generated using a tenfold diluted cDNA series, the starting quantity of cDNA was 500 ng/μL50.
Data analysis
The stability of the eight candidate reference genes was assessed using GeNorm10, NormFinder software11, BestKeeper12, and Ref-finder (http://www.leonxie.com/referencegene.php). For GeNorm and NormFinder analysis, quantification cycle (Cq) values were transformed into relative expression levels using the formula: 2−∆Cq, in which ∆Cq = each corresponding Cq value-the minimum Cq value16. The expression stability measurement (M) was determined by the GeNorm program based on the average pairwise variations (V) among all the other reference genes. NormFinder program estimates intra- and intergroup variations, and the lowest stability is ranked the highest. Calculations of the BestKeeper program are calculated based on the coefficients of variation (CV) and the standard deviations (SD) of the Cq values, and the lowest CV and SD were used as detection indexes for the most-stable reference genes. Ref-finder is an online tool that integrates the results of the currently available major computational programs, including GeNorm (M values), NormFinder (stability values), BestKeeper (CV and SD), and ∆Cq values.
Validation of selected reference genes
To validate the influence of the choice of different reference genes on the final normalized outcome, the relative expression levels of IgFT which plays an important role in promoting flowering51 in three cultivars were analyzed using individual stably expressed or unstably expressed genes or a combination of stable reference genes, as determined by GeNorm48. The primers used for IgFT are presented in Table 1. The fold change of gene expression was calculated using the 2−∆∆Ct method24.
Conclusions
This research provided the first systematic analysis for the selection of stable reference genes as the internal control in RT-qPCR analysis in different flowering stages and different genotypes of I. germanica cultivars. Analysis using GeNorm, NormFinder, BestKeeper, and Ref-finder revealed that IgUBC, IgGAPDH, and IgTUB could be considered as appropriate reference genes for gene expression analysis in future molecular researches that aim to understand the mechanisms of flowering stages in I. germanica.
References
VanGuilder, H. D., Vrana, K. E. & Freeman, W. M. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques 44, 619–626. https://doi.org/10.2144/000112776 (2008).
Bustin, S. A. et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622. https://doi.org/10.1373/clinchem.2008.112797 (2009).
Artico, S., Nardeli, S. M., Brilhante, O., Grossi-de-Sa, M. F. & Alves-Ferreira, M. Identification and evaluation of new reference genes in Gossypium hirsutum for accurate normalization of real-time quantitative RT-PCR data. BMC Plant Biol. 10, 49. https://doi.org/10.1186/1471-2229-10-49 (2010).
Derveaux, S., Vandesompele, J. & Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 50, 227–230. https://doi.org/10.1016/j.ymeth.2009.11.001 (2010).
Kozera, B. & Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 54, 391–406. https://doi.org/10.1007/s13353-013-0173-x (2013).
Xiao, X. L. et al. Validation of suitable reference genes for gene expression analysis in the halophyte Salicornia europaea by real-time quantitative PCR. Front. Plant Sci. 5, 788. https://doi.org/10.3389/fpls.2014.00788 (2014).
Delporte, M., Legrand, G., Hilbert, J. L. & Gagneul, D. Selection and validation of reference genes for quantitative real-time PCR analysis of gene expression in Cichorium intybus. Front. Plant Sci. 6, 651. https://doi.org/10.3389/fpls.2015.00651 (2015).
Niu, X. P. et al. Selection of reliable reference genes for quantitative real-time PCR gene expression analysis in Jute (Corchorus capsularis) under stress treatments. Front. Plant Sci. 6, 848. https://doi.org/10.3389/fpls.2015.00848 (2015).
Zhang, Y. X. et al. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Salix matsudana under different abiotic stresses. Sci. Rep. 7, 40290. https://doi.org/10.1038/srep40290 (2017).
Vandesompele, J. et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. https://doi.org/10.1186/gb-2002-3-7-research0034 (2002).
Andersen, C. L., Jensen, J. L. & Ørntoft, T. F. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64, 5245–5250. https://doi.org/10.1158/0008-5472.CAN-04-0496 (2004).
Pfaffl, M. W., Tichopad, A., Prgomet, C. & Neuvians, T. P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: bestkeeper-Excel-based tool using pair-wise correlations. Biotechnol. Lett. 26, 509–515. https://doi.org/10.1023/B:BILE.0000019559.84305.47 (2004).
Qi, S. et al. Reference gene selection for RT-qPCR analysis of flower development in Chrysanthemum morifolium and Chrysanthemum lavandulifolium. Front. Plant Sci. 7, 287. https://doi.org/10.3389/fpls.2016.00287 (2016).
Mallona, I., Lischewski, S., Weiss, J., Hause, B. & Egea-Cortines, M. Validation of reference genes for quantitative real-time PCR during leaf and flower development in Petunia hybrida. BMC Plant Biol. 10, 4–14. https://doi.org/10.1186/1471-2229-10-4 (2010).
Yi, S. J. et al. Selection of reliable reference genes for gene expression studies in Rhododendron micranthum Turcz. Sci. Hortic-Amsterdam 138, 128–133. https://doi.org/10.1016/j.scienta.2012.02.013 (2012).
Li, J., Han, J. G., Hu, Y. H. & Yang, J. Selection of reference genes for quantitative real-time PCR during flower development in tree peony (Paeonia suffruticosa Andr). Front. Plant Sci. 7, 516. https://doi.org/10.3389/fpls.2016.00516 (2016).
Linnegar, S. & Hewitt, J. Irises (Octopus Publishing Group, 2008).
Wang, Y. J. et al. Reference gene selection for qRT-PCR normalization in Iris germanica L. Phyton Int. J. Exp. Bot. 90, 277–290. https://doi.org/10.32604/phyton.2020.011545 (2020).
Silveira, É. D., Alves-Ferreira, M., Guimarães, L. A., da Silva, F. R. & Carneiro, V. T. Selection of reference genes for quantitative real-time PCR expression studies in the apomictic and sexual grass Brachiaria brizantha. BMC Plant Biol. 9, 84. https://doi.org/10.1186/1471-2229-9-84 (2009).
Marum, L., Miguel, A., Ricardo, C. P. & Miguel, C. Reference gene selection for quantitative real-time PCR normalization in Quercus suber. PLoS ONE 7, e35113. https://doi.org/10.1371/journal.pone.0035113 (2012).
Niu, K. J., Shi, Y. & Ma, H. L. Selection of candidate reference genes for gene expression analysis in Kentucky Bluegrass (Poa pratensis L.) under abiotic stress. Front. Plant Sci. 8, 193. https://doi.org/10.3389/fpls.2017.00193 (2017).
Chen, Y. et al. Selection and validation of reference genes for target gene analysis with quantitative RT-PCR in leaves and roots of bermudagrass under four different abiotic stresses. Physiol. Plant. 155, 138–148. https://doi.org/10.1111/ppl.12302 (2014).
Xie, F. L., Xiao, P., Chen, D. L., Xu, L. & Zhang, B. H. miRDeepFinder: a miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 80, 75–84. https://doi.org/10.1007/s11103-012-9885-2 (2012).
Huggett, J. F., Dheda, K., Bustin, S. A. & Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 6(279–284), 2012. https://doi.org/10.1038/sj.gene.6364190 (2005).
Chen, Y. et al. Cold acclimation induces freezing tolerance via antioxidative enzymes, proline metabolism and gene expression changes in two chrysanthemum species. Mol. Biol. Rep. 41, 815–822. https://doi.org/10.1007/s11033-013-2921-8 (2014).
Chapman, J. R. & Waldenström, J. With reference to reference genes: a systematic review of endogenous controls in gene expression studies. PLoS ONE 10, e0141853. https://doi.org/10.1371/journal.pone.0141853 (2015).
Wang, M. & Lu, S. Validation of suitable reference genes for quantitative gene expression analysis in Panax ginseng. Front. Plant Sci. 6, 1259. https://doi.org/10.3389/fpls.2015.01259 (2016).
Galli, G. G. et al. YAP drives growth by controlling transcriptional pause release from dynamic enhancers. Mol. Cell 60, 328–337. https://doi.org/10.1016/j.molcel.2015.09.001 (2015).
He, Y., Yan, H., Hua, W., Huang, Y. & Wang, Z. Selection and validation of reference genes for quantitative real-time PCR in Gentiana macrophylla. Front. Plant Sci. 7, 945. https://doi.org/10.3389/fpls.2016.00945 (2016).
Tang, F. et al. Selection and validation of reference genes for quantitative expression analysis of miRNAs and mRNAs in Poplar. Plant Methods 15, 35. https://doi.org/10.1186/s13007-019-0420-1 (2019).
Shivhare, R. & Lata, C. Selection of suitable reference genes for assessing gene expression in pearl millet under different abiotic stresses and their combinations. Sci. Rep. 6, 23036. https://doi.org/10.1038/srep23036 (2016).
Kim, H. et al. RNA-Seq analysis of spatiotemporal gene expression patterns during fruit development revealed reference genes for transcript normalization in plums. Plant Mol. Biol. Rep. 33, 1634–1649. https://doi.org/10.1007/s11105-015-0860-3 (2015).
Xiao, Z. et al. Selection of reliable reference genes for gene expression studies on Rhododendron molle G. Don. Front. Plant Sci. 7, 1547. https://doi.org/10.3389/fpls.2016.01547 (2016).
Wang, C. et al. Identification and validation of reference genes for RT-qPCR analysis in non-heading Chinese cabbage flowers. Front. Plant Sci. 7, 811. https://doi.org/10.3389/fpls.2016.00811 (2016).
Fu, J., Wang, Y., Huang, H., Zhang, C. & Dai, S. L. Reference gene selection for RT-qPCR analysis of Chrysanthemum lavandulifolium during its flowering stages. Mol. Breed. 31, 205–215. https://doi.org/10.1007/s11032-012-9784-x (2013).
Fulvio, F., Martinelli, T. & Paris, R. Selection and validation of reference genes for RT-qPCR normalization indifferent tissues of milk thistle (Silybum marianum, Gaert). Gene Gene https://doi.org/10.1016/j.gene.2020.145272 (2020).
Gu, C. S. et al. Validation of reference genes for RT-qPCR normalization in Iris. lacteavar. chinensis leaves under different experimental conditions. Sci. Hortic-Amsterdam 175, 144–149. https://doi.org/10.1016/j.scienta.2014.06.011 (2014).
Ma, L. L. et al. Selection and validation of reference genes for quantitative real-time PCR analysis in Iris bulleyana during flower color variation. J. Nucl. Agric. Sci. 33, 1707–1716 (2019).
Saddhe, A., Malvankar, M. & Kumar, K. Selection of reference genes for quantitative real-time PCR analysis in halophytic plant Rhizophora apiculate. PeerJ 6, e5226. https://doi.org/10.7717/peerj.5226 (2018).
Ma, R., Xu, S., Zhao, Y., Xia, B. & Wang, R. Selection and validation of appropriate reference genes for quantitative real-time PCR analysis of gene expression in Lycoris aurea. Front. Plant Sci. 7, 536. https://doi.org/10.3389/fpls.2016.00536 (2016).
Fan, C. et al. Selection of reference genes for quantitative real-time PCR in bamboo (Phyllostachys edulis). PLoS ONE 8, e56573. https://doi.org/10.1371/journal.pone.0056573 (2013).
Wang, H. B., Chen, S. M., Jiang, J. F., Zhang, F. & Chen, F. D. Reference gene selection for cross-species and cross-ploidy level comparisons in Chrysanthemum spp. Sci. Rep. 5, 8094. https://doi.org/10.1038/srep08094 (2015).
Garrido, J., Aguilar, M. & Prieto, P. Identification and validation of reference genes for RT-qPCR normalization in wheat meiosis. Sci. Rep. 10, 2726. https://doi.org/10.1038/s41598-020-59580-5 (2020).
Jia, Y. et al. Reference gene selection and validation by qRT-PCR during flower development and in different organs of Primula forbesii. J. Hortic. Sci. Biotech. 95, 383–394. https://doi.org/10.1080/14620316.2019.1681909 (2019).
Tong, Z., Gao, Z., Wang, F., Zhou, J. & Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 10, 71. https://doi.org/10.1186/1471-2199-10-71 (2009).
Wang, G. L. et al. Selection of reliable reference genes for quantitative RT-PCR in garlic under salt stress. PeerJ 7, e7319. https://doi.org/10.7717/peerj.7319 (2019).
Li, L., Li, N., Fang, H., Qi, X. & Zhou, Y. Selection and validation of reference genes for normalisation of gene expression in Glehnia littoralis. Sci. Rep. 10, 7374. https://doi.org/10.1038/s41598-020-63917-5 (2020).
Gu, C. S. et al. Reference gene selection for quantitative real-time RT-PCR normalization in Iris. lactea var. chinensis roots under cadmium, lead, and salt stress conditions. Sci. World J. 2014, 532713. https://doi.org/10.1155/2014/532713 (2014).
Gu, G. S. et al. De novo characterization of the Iris lactea var. chinensis transcriptome and an analysis of genes under cadmium or lead exposure. Ecotox Environ. Safe 144, 507–513. https://doi.org/10.1016/j.ecoenv.2017.06.071 (2017).
Taylor, S., Wakem, M., Dijkman, G., Alsarraj, M. & Nguyen, M. A practical approach to RT-qPCR-Publishing data that conform to the MIQE guidelines. Methods 50, S1–S5. https://doi.org/10.1016/j.ymeth.2010.01.005 (2010).
Zhang, H. L. et al. Precocious flowering in trees: the Flowering Locus T gene as a research and breeding tool in Populus. J Exp Bot 61, 2549–2560. https://doi.org/10.1093/jxb/erq092 (2010).
Acknowledgements
The work was supported by the National Natural Science Foundation of China (31801901), the Natural Science Foundation of Jiangsu (BK20180314, BK20180317), the Foundation of Key Laboratory of Landscaping (KF201901), Ministry of Agriculture and Rural Affairs, P. R. China, and the Jiangsu Key Laboratory for the Research and Utilization of Plant Resources (JSPKLB201814).
Author information
Authors and Affiliations
Contributions
Y.W., H.Y. conceived and designed the experiments. Y.W., Y.Z., Q.L., T.Z. and L.L. performed the experiments and collected the data. Y.W., H.T. and S.H. prepared reagents/materials/analysis tools. Y.W. and C.G. analyzed the data; Y.W. wrote the paper. Y.W. and H.Y. revised the paper. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, Y., Zhang, Y., Liu, Q. et al. Selection and validation of appropriate reference genes for RT-qPCR analysis of flowering stages and different genotypes of Iris germanica L. Sci Rep 11, 9901 (2021). https://doi.org/10.1038/s41598-021-89100-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-89100-y
This article is cited by
-
Reference genes for qPCR expression in black tiger shrimp, Penaeus monodon
Molecular Biology Reports (2024)
-
Selection and validation of appropriate reference genes for RT–qPCR analysis of Nitraria sibirica under various abiotic stresses
BMC Plant Biology (2022)
-
Identification of appropriate housekeeping genes for gene expression studies in human renal cell carcinoma under hypoxic conditions
Molecular Biology Reports (2022)
-
Validation of reference genes for quantitative real-time PCR in chemical exposed and at different age’s brackish water flea Diaphanosoma celebensis
Scientific Reports (2021)
-
Stability analysis of reference genes for RT-qPCR assays involving compatible and incompatible Ralstonia solanacearum-tomato ‘Hawaii 7996’ interactions
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
