
Abstract
Gyrocotylideans are evolutionary ancient parasitic flatworms, and like their hosts—a relict group of holocephalan fishes (Chimaeriformes)—they are considered to be “living fossils” of a vanished past. However, the species diversity, host associations and biogeography of these most basal tapeworms are poorly known. Herein, we provide evidence of a conspicuous contrast between the genetic and morphological data based on an examination of newly collected and properly processed Gyrocotyle specimens (hologenophores) isolated from holocephalans off Taiwan and Argentina. Our molecular data, inferred from three genes (COI, 28S rRNA, 18S rRNA), showed unexpected genetic interrelationships among isolates of the genus Gyrocotyle, because each of the four genotypes from Taiwan clustered with isolates of distinct gyrocotylideans from the North Atlantic. Three genotypes of Gyrocotyle from Taiwan were morphologically almost indistinguishable from each other but represented distinct genetic lineages; a single specimen of Gyrocotyle sp. genotype 4 exhibited a clear genetic and morphological distinctness, though its formal description as a new species would be premature. Additionally, specimens of Gyrocotyle rugosa Diesing, 1850, from the type host Callorhinchus callorynchus from Argentina, provided the first genetic data on the type species of the genus and enabled us to characterise it, which is necessary for future taxonomic studies. The finding of some specimens of Gyrocotyle sp. genotype 3 in Chimaera phantasma, and another one in C. cf. argiloba, together with the putative conspecificity of an unidentified gyrocotylidean from Callorhinchus milii off Australia and G. rugosa from C. callorynchus off Argentina, represent evidence that one gyrocotylidean species may parasitise more than one holocephalan host species. Existing taxonomic problems and conflicts between morphological and molecular data on species of Gyrocotyle can only be resolved if hologenophores from type hosts and localities of nominal taxa are properly characterised genetically and morphologically.
Similar content being viewed by others
Introduction
The order Gyrocotylidea Poche, 1926, represents an enigmatic and the most basal extant group of tapeworms, with a unique monozoic body and a posteriorly located funnel-shaped adhesive organ1,2. This relatively small taxon currently has 10 valid species, which are considered strictly host-specific parasites of the spiral intestine of holocephalans (Chimaeriformes), mostly deep-sea dwelling cartilaginous fishes3. Gyrocotylideans are distributed worldwide and have been reported from 15 out of the 56 (25%) recognised extant species of holocephalans, with a high prevalence that may reach up to 100%3,4. To the best of our knowledge, each holocephalan that has been examined for endohelminths thus far hosts at least one gyrocotylidean species5. This suggests that major parts of gyrocotylidean diversity may be undiscovered. Recently, two new species of Gyrocotyle, from Harriotta raleighana Goode & Bean and Hydrolagus mirabilis (Collett), respectively, were described based only on genetic data, because the authors considered morphology to be totally unsuitable for species characterisation6.
To date, most of the species have been described from the North Atlantic5,6,7,8,9,10, whereas a few taxa have been reported from the southern seas5,11,12,13, including the type species Gyrocotyle rugosa Diesing, 1850. The only gyrocotylidean recorded in east Asian waters was found in Chimaera phantasma Jordan & Snyder from the coast of Japan and was identified as Gyrocotyle fimbriata Watson, 1911, in a brief report of Ichihara14 and in two unidentified species of Hydrolagus and Harriotta off Indonesia3. However, the former finding needs verification, because G. fimbriata was originally described as a parasite of Hydrolagus colliei (Lay & Bennett) from the west coast of the United States of America15.
In fact, reliable morphological identification is currently barely possible because of insufficient original descriptions dated mostly to the first half of the last century and overlapping diagnostic traits among the individual species3,16. Genetic identification is also limited, as sequence data are available, with one exception, for a few species from the North Atlantic, and some of them are most probably misidentified6,17,18,19,20. The reliable identification of many species is thus impossible and was often based only on their host and geographic origin.
However, some holocephalans may host more than a single gyrocotylidean species. Chimaera monstrosa L. may apparently be infected with Gyrocotyle urna (Grube & Wagener, 1852), Gyrocotyle confusa van der Land & Dienske, 1968, and Gyrocotyle nybelini (Fuhrmann, 1931). Another holocephalan, Hydrolagus colliei, may harbour G. fimbriata and G. parvispinosa van der Land & Dienske, 1968, and Hydrolagus affinis (de Brito Capello) was found to be the host of Gyrocotyle major van der Land & Templeman, 1968 and Gyrocotyle abyssicola van der Land & Templeman, 19689,10,21,22,23. Additionally, Manter24 and Bandoni & Brooks16 questioned the strict host specificity of gyrocotylideans and reported G. urna and G. fimbriata as parasites of more than one holocephalan species.
Herein, we suggest an approach based on examination of voucher specimens of sequenced samples, i.e., hologenophores (see25). We document the unexpected genetic divergence of morphologically almost indistinguishable gyrocotylideans from the East China Sea and also provide the first detailed molecular and morphological characterisation of Gyrocotyle rugosa from the type host as a baseline for future studies.
Results
Molecular analysis of Gyrocotyle spp
In total, 17 specimens of Gyrocotyle spp. from Taiwan were genotyped and four distinct genotypes were identified based on partial sequences of the COI, 28S rRNA and 18S rRNA genes. The pairwise divergence (uncorrected p-value) within the particular genotypes (i.e., intragenotypic divergence) ranged from 0.2 to 0.8% for the COI dataset, whereas the sequences of 28S rDNA were identical (the value was not calculated for 18S rRNA, because a single sequence was obtained for each genotype). The intergenotypic pairwise divergences were 6.1‒9.1% for COI, 1.2‒3.9% for 28S rRNA, and 0.9‒1.2% for 18S rRNA. Considering isolates of Gyrocotyle rugosa from Callorhinchus callorynchus L. off Argentina, identical sequences of both COI and 28S rRNA genes were obtained from two specimens sequenced. The pairwise divergence between them and possibly a conspecific isolate of Gyrocotyle sp. from Callorhinchus milii Bory de Saint-Vincent off Australia was 0.5% and 0.2% for the 28S rRNA and 18S rRNA genes, respectively.
For phylogenetic analysis, amphilinidean, caryophyllidean and spathebothriidean taxa were tested as potential outgroups. However, the numerous gaps in alignments caused a considerable loss of signal due to the removal of ambiguously aligned sequence positions. As rooted trees with isolates of none of the three tapeworm orders provided robust support for phylogenetic relationships among the gyrocotylidean taxa, their genetic relationships are herein depicted as an un-rooted network of two concatenated datasets, i.e. 28S rRNA + 18S rRNA genes and COI + 28S rRNA + 18S rRNA genes.
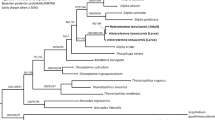
Out of 17 specimens genotyped, only the specimens genetically unique in at least one genetic marker were chosen for the further analysis (for the complete list of the material examined, see Supplementary Table S1). Among the unique isolates, the four genotypes of Gyrocotyle off Taiwan did not cluster together but formed separate groups spread among other gyrocotylideans in both analyses (Fig. 1a,b). The individuals of Gyrocotyle sp. genotype 1 from Chimaera phantasma exhibited the highest genetic similarity with the specimens of Gyrocotyle urna from Chimaera monstrosa off Norway and an isolate identified as “Gyrocotyle rugosa” from Hydrolagus colliei in the Gulf of Alaska. Two specimens of Gyrocotyle sp. genotype 2 from Chimaera cf. argiloba diverged from the lineage with the isolates of Gyrocotyle discoveryi Bray, Waeschenbach, Littlewood, Harvolsen & Olson 2020 from Hydrolagus mirabilis off Ireland, and the specimens of Gyrocotyle sp. genotype 3 from C. phantasma and C. cf. argiloba grouped with the isolate identified as “Gyrocotyle urna” from Chimaera monstrosa off Ireland. The specimen of Gyrocotyle sp. genotype 4 appeared in the branch with the isolate of Gyrocotyle confusa from Chimaera monstrosa off Norway, both being closely related to Gyrocotyle haffii Bray, Waeschenbach, Littlewood, Harvolsen & Olson 2020 from Harriotta raileghana off the North Atlantic and Gyrocotyle nybelini from Chimaera monstrosa off Norway, which had by far the longest branch in both analyses (Fig. 1a,b).

Estimation of genetic interrelationships among the isolates of the genus Gyrocotyle. Maximum likelihood analysis performed on two concatenated datasets of two (28S rRNA + 18S rRNA) (a) and three (COI + 28S rRNA + 18S rRNA), (b) partial genes depicted as unrooted networks. Note the genetic diversity of the four genotypes of Gyrocotyle sp. from Taiwan, each of which clustered with G. urna off Norway + ‘G. rugosa’ off Alaska, G. discovery off Ireland, ‘G. urna’ off Ireland, and G. confusa off Norway, respectively, rather than together. Also note the cluster of Gyrocotyle sp. genotype 3 specimens isolated from Chimaera phantasma and C. cf. argiloba, respectively, and the close relatedness of Gyrocotyle rugosa from Callorhinchus callorynchus off Argentina with the isolate of Gyrocotyle sp. from Callorhinchus milii off Australia. Our original isolates are in black, with a micrograph of the morphological voucher; asterisks mark the hologenophores. The ultrafast bootstrap supports over 89% and 94% are shown as open circles and black dots, respectively. The full length of the branches and gene concatenations are in the rectangular box, the identification of isolates in Table 1.
Additionally, our specimen of Gyrocotyle rugosa off Argentina grouped with the only available isolate from the Southern hemisphere, Gyrocotyle sp. from Callorhinchus milii off Australia. These two specimens formed a relatively long branch, which was closely related to the group of Gyrocotyle sp. genotype 2 off Taiwan + Gyrocotyle discoveryi in the 28S rRNA + 18S rRNA gene network (Fig. 1a), though they exhibited a different topology when the COI gene was included (Fig. 1b). Moreover, the addition of the COI data increased the bootstrap support for some nodes when comparing with the dataset of ribosomal genes only.
Morphological characterisation of specimens off Taiwan and Argentina
The specimens of Gyrocotyle sp. genotypes 1, 2 and 3 off Taiwan are similar in their general morphology, i.e., they possess heavily plicated lateral margins, a wide funnel and a large complex rosette. The copulatory papilla and external and internal seminal vesicles are present, and the male pore opens slightly posterior to the level of the vaginal pore. The ovary is U-shaped, the uterus coiled, and its distal part forms a well-developed uterine sac containing unembryonated eggs. Two excretory pores open posterolateral to the uterine pore (Figs. 2a‒f, 3a‒d).

Whole mounts of Gyrocotyle spp. off Taiwan and Argentina. Morphological vouchers of Gyrocotyle sp. genotype 1 ex Chimaera phantasma (a), specimen TW212-1*; (b), specimen TW13-1, Gyrocotyle sp. genotype 2 ex Chimaera cf. argiloba (c), specimen TW138*; (d), specimen TW139*, Gyrocotyle sp. genotype 3 ex C. cf. argiloba (e), specimen TW215* and C. phantasma (f), specimen TW10-1, Gyrocotyle sp. genotype 4 ex C. cf. argiloba (g), specimen TW141*, all from Taiwan; and Gyrocotyle rugosa ex Callorhinchus callorynchus Argentina (h), specimen AR89*. Field numbers with asterisks above indicate the hologenophores. ac acetabulum, do dorsal opening, ep excretory pore, gn genital notch, mg male gonopore, ov ovary, rs receptaculum seminis, te testes, us uterine sac, ut uterus, va vagina.

Scanning electron micrographs of Gyrocotyle sp. genotype 1 ex Chimaera phantasma (Taiwan). Ventral view of a juvenile specimen TW12-3; note the distribution of body spines restricted near the rosette, lateral plication, and weakly developed genital pores (a). The anterior body part of an adult TW12-3 specimen (b), intrauterine egg with smooth surface and shallow operculum (c), detail of an acetabular spine (d). ao acetabular opening, as acetabular spines, bs body spines, gn genital notch, gp genital papilla, op operculum, up uterine pore.
However, these three genotypes can be distinguished from each other by the unique combination of morphological traits (Supplementary Table S2). Specimens of Gyrocotyle sp. genotype 1 possess a large acetabulum and a rosette of the same width or wider than the maximum body width. The acetabular spines are less numerous, and the dorsal body spines are small and distributed mostly on the posterior half of the body, whereas ventral spines are few and restricted to near the rosette. The testicular field reaches one-fourth of the uterus length, the ejaculatory duct is straight, and the uterine sac is middle-sized (Fig. 2a,b).
Gyrocotyle sp. genotype 2 is characterised by a large acetabulum and the rosette is not wider than the maximum body width. The acetabular spines are numerous and the body spines are small, with dorsal spines distributed on the posterior half of the body and near the lateral margins towards the acetabulum, whereas the ventral body spines are restricted to near the rosette; anterolateral spines (posterior to the acetabular spines) are also present. The testicular field reaches one-half of the uterus, the ejaculatory duct is coiled, and the uterine sac is middle-sized (Fig. 2c,d).
Gyrocotyle sp. genotype 3 possesses a small acetabulum and the rosette is not wider than the maximum body width. The acetabular spines are less numerous, body spines are large and distributed over the whole body surface. They are more numerous on the dorsal than on the ventral side, and their density is highest on the funnel, decreasing towards the acetabulum. The testicular field reaches one fourth of the uterus, the ejaculatory duct is straight, and the uterine sac is large (Fig. 2e,f).
The single specimen of Gyrocotyle sp. genotype 4 markedly differs from the specimens of all the genotypes described above by its lateral margins with few deep folds, numerous acetabular spines, a relatively narrow funnel and rosette, and the ovary being far from both the dorsal opening and the posterior body margin (Fig. 2g).
Gyrocotyle rugosa from Callorhinchus callorynchus off Argentina is typified by possessing an elongated body with crenulate lateral margins, a small rosette, a weakly developed uterine sac, a branched uterus, a V-shaped ovary and embryonated intrauterine eggs (Fig. 2h).
Molecular identification of Chimaera spp. off Taiwan
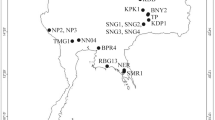
The analysis of the hosts’ COI gene distinguished two host genotypes (Fig. 4). The first genotype clustered within the Chimaera phantasma clade, whereas the second one nested with the isolates of Chimaera argiloba Last, White & Pogonoski off Australia, Indonesia and New Caledonia. Although the identification of the second Taiwanese genotype as Chimaera cf. argiloba is provisional due to its distinct geographical origin, it is evident that we examined two different host species from the East China Sea.

Hosts of gyrocotylideans off Taiwan and their phylogenetic positions. Maximum likelihood analysis on a partial COI gene dataset distinguished two host genotypes, i.e., Chimaera phantasma and C. cf. argiloba. Newly generated sequences are in black. The ultrafast bootstrap supports over 89% and 94% are depicted as open circles and black dots, respectively. The scale bar represents the number of substitutions. Numbers in parentheses are accession numbers in the GenBank database.
Discussion
Almost 50 years ago, Simmons26 called gyrocotylideans a “century-old enigma” and this status still persists despite the advent of more advanced identification methods3. The poor understanding of the group (e.g., the complete life cycle of none of the species is known) is linked with the scarcity of available data and the biological peculiarities of these tapeworms and their holocephalan hosts. In particular, most of the host species are rarely available deep-sea dwellers, which often could not be examined fresh or were frozen with their parasites prior to examination. If isolated alive, gyrocotylideans exhibit an unusual morphological variability due to the contraction of their large bodies and as a result of different fixative procedures which were tested to ensure their relaxation (e.g.27). Despite these issues, several comprehensive studies have been conducted, e.g.15,16,21,28, which provided deep insight into the biology, ecology and taxonomy of these enigmatic tapeworms. Nevertheless, the poor quality of the specimens studied and the use of different, not always appropriate, methods of parasite fixation, unintentionally affected the quality of morphological descriptions of most gyrocotylidean species, which prevented the establishing of clear morphological borders to delimit individual species. As a result, the informative value of morphological traits used for species delimitation should be re-assessed, based on the simultaneous use of molecular data, i.e., the use of hologenophores to match morphology and molecular data. Existing problems with species delimitation and morphological variability even led to complete omission of morphological characterisation of two new species described just recently6.
Herein, the genotyping of the Gyrocotyle spp. specimens acquired in Taiwan revealed four distinct genotypes, each one more related to the North Atlantic isolates identified as “Gyrocotyle urna” off Ireland (the isolate is genetically diverse from G. urna off Norway), “G. rugosa” off Alaska (probably misidentified, see below), G. discoveryi off Ireland and G. confusa off Norway, respectively, than to each other.
In addition to casting doubts on the restriction of gyrocotylideans to individual oceans, our data also question the proclaimed strict host specificity3,7, because specimens of Gyrocotyle sp. genotype 3 were found in two hosts species, which are not the closest relatives to one another—C. phantasma and C. cf. argiloba (Fig. 4). Broader host specificity was also reported for G. fimbriata, which was found in Hydrolagus colliei and Chimaera phantasma, and for G. rugosa, recorded in Callorhinchus callorynchus and C. milii14,15,24,29. Gyrocotyle urna was also found in several holocephalans, including Chimaera monstrosa, Callorhinchus callorynchus, Hydrolagus ogilbyi Waite and H. colliei24,29,30. In contrast, Bandoni & Brooks16 revised the host spectrum of this parasite, considering C. monstrosa as the only host of G. urna.
The suitability of the molecular markers employed for this group also requires attention, because a considerable amount of phylogenetic information was also lost in the un-rooted dataset due to treatment of the numerous gaps in the 28S rRNA alignment. The involvement of partial COI gene sequences seemed to be informative for estimating gyrocotylidean phylogeny, because we obtained a no-gap COI alignment and improved support for some nodes in the three-gene network. The suitability of this marker requires assessment employing further taxa, because except for our isolates off Taiwan and Argentina, only a single sequence of the COI gene (i.e., that of G. urna off Norway; GenBank acc. no. JQ268546) is currently available.
A single specimen of Gyrocotyle sp. genotype 4 was conspicuously different morphologically from the remaining ones by having few folds on the lateral margins, many acetabular spines, a narrow funnel and a small rosette. However, its formal description as a new species would be premature, because only a single specimen was found. Morphological differences among the specimens of the other genotypes were not so obvious, even though a careful examination of the hologenophores allowed us to find several morphological traits that were characteristic for particular genotypes (see “Results” section). Among them, the number of acetabular spines and the distribution of the body spines and their size may be potentially useful for species differentiation, especially because the body contraction can hardly affect them. Since body contraction cannot be absolutely excluded even when live specimens are properly fixed, its effect could be overcome to some degree by an evaluation of ratios related to the main body dimensions (e.g., length of uterine sac/total body length) rather than comparison of total measurements of internal structures.
The specimens off Taiwan most probably represent several new species, but we decided not to describe them formally as new taxa, mainly because of the shortage of comparative data. In addition to these specimens, two hologenophores of Gyrocotyle rugosa off Argentina were examined, which made it possible to characterise the type species of the genus. The host of G. rugosa described by Diesing10 was questionable until Callorhynchus antarcticus (= C. callorynchus—see31) off New Zealand was finally established as its currently accepted type host3,32. Gyrocotyle rugosa was found in coastal waters of South America, South Africa and New Zealand as a parasite of C. callorynchus and C. milii, suggesting its broader host specificity16,24. Our specimens from C. callorynchus off Argentina were identified as G. rugosa based on crenulated (i.e., without any folds) lateral margins, a tiny uterine sac, a branched uterus and embryonated eggs in the uterine sac; the latter two traits are unique to this species21. Genetically, it clustered with an unspecified isolate of Gyrocotyle from C. milii off Australia, and these specimens seem to be conspecific.
In contrast, an isolate from Hydrolagus colliei off Alaska identified as G. rugosa (GenBank acc. nos. AF286925 and AF124455) was apparently misidentified, because (i) it was found in an unrelated definitive host (H. colliei belongs to the family Chimaeridae, whereas the type host to the family Callorhinchidae), (ii) its distant geographic origin (the type locality of G. rugosa is unclear, but it is definitely in the Southern hemisphere), and (iii) its genetic divergence from our isolate of G. rugosa from the type host off Argentina. The isolate from H. colliei may represent Gyrocotyle fimbriata or G. parvispinosa, which have been reported from this host off the Pacific coast of North America, but its identification was not possible because morphological vouchers were not available to the present authors.
Gyrocotylideans were generally considered to be oioxenous, i.e. strictly specific parasites sensu Euzet and Combes33, with each gyrocotylidean species parasitising a single holocephalan species. Although several species were reported from two or more hosts species16,24, these findings are usually considered as misidentifications due to the unclear taxonomy of the order. Moreover, some holocephalans, such as Ch. monstrosa, H. colliei, H. affinis, and Ca. callorynchus, were often found to harbour two or more gyrocotylidean species, one common and the other rare9,10,21,22,23. Our findings of Gyrocotyle sp. genotypes 1 and 3 in Ch. phantasma and Gyrocotyle sp. genotypes 2, 3 and 4 in Ch. cf. argiloba suggested stenoxenous host specificity (i.e., the occurrence in a few closely related hosts) of gyrocotylideans, because the specimens of genotype 3 were found in both species of Chimaera. The obvious genetic similarity of our G. rugosa specimen from Ca. callorynchus and the isolate of Gyrocotyle sp. from Ca. milii also questions the strict specificity of this group, but morphological vouchers of the latter, which are necessary for the confirmation of their conspecificity, are not available.
Our genetic analyses provided insight into the interrelationships among the gyrocotylideans, even though the absence of a suitable outgroup did not enable us to broadly assess the possible evolutionary scenario of this earliest evolving group of tapeworms. Moreover, genetic data on only half of the nominal species of Gyrocotyle are available, not considering the possibility of misidentifications of previously sequenced specimens, for which hologenophores are not available. However, some clues of host-parasite coevolution can be inferred from the network. The mutual genetic distance of species/genotypes from the same host species suggests multiple colonisation events rather than co-speciation with their hosts within the order. It seems that G. phantasma might have been colonised by Gyrocotyle sp. genotype 1 or genotype 3, because these two genotypes are not the closest relatives in our analyses. The same pattern is obvious for C. cf. argiloba parasitised by Gyrocotyle sp. genotype 2, 3 and 4, and also for C. monstrosa, which harbours G. urna, G. confusa and G. nybelini. Indeed, Colin et al.27 considered these species from C. monstrosa to be conspecific, but our genetic data support the validity of three separate and genetically distant species. Moreover, G. nybelini formed by far the most distant lineage among all isolates, which may suggest the validity of the genus Gyrocotyloides Furhmann, 1931.
Genetic divergence of congeneric tapeworms from the same host species was also observed in several elasmobranch/teleost-cestode assemblages, e.g., Acanthobothrium spp. (Onchoproteocephalidea) and the mumburarr whipray Urogymnus acanthobothrium Last, White & Kyne; Echeneibothrium spp. (Rhinebothriidea) and the yellownose skate Dipturus chilensis (Guichenot); and Pseudoendorchis spp. (Onchoproteocephalidea) and the catfish Pimelodus maculatus Lacepède34,35,36.
The aim of this paper was to provide new insight into the phylogenetic relationships within the enigmatic order Gyrocotylidea, but, in particular, to demonstrate the lack of geographical patterns in the distribution of most its species and the limited suitability of current morphological characteristics for species circumscription. Herein, we have outlined a methodology (fixation of live specimens with hot fixative and the exclusive use of hologenophores) that should be used in future taxonomic, ecological and biogeographical studies of gyrocotylideans in order to reliably circumscribe their actual species diversity and to unravel associations with their hosts, a relict group of marine vertebrates. Gyrocotylideans represent one of the key groups of parasitic flatworms (Neodermata) in terms of a better understanding of their evolutionary history and the switch of free-living flatworms to parasitism.
Material and methods
Fifteen individuals of Chimaera spp. (Chimaeridae) from the fish market in Yilan City, northern Taiwan, and several individuals of Callorhinchus callorynchus (Callorhinchidae) off Mar del Plata, Argentina, were examined from 2004 to 2019. Their morphological identification followed Ebert et al.37; isolation of their genomic DNA and amplification of partial mitochondrial cytochrome c oxidase subunit 1 (COI) genes were performed according to Holmes et al.38. Purification of the amplicons, sequencing method and processing of raw sequence data were same as for the parasites (see below).
Live tapeworms isolated from the spiral valve of the above-mentioned holocephalans (prevalence 100%) were processed using standard helminthological techniques (see39); specimens were fixed with almost boiling water and then placed in 70% non-denaturated ethanol. This approach allowed the isolation of DNA from a small piece of the parasite tissue, whereas the rest of the body was used for morphological observations as a hologenophore (see25). Additionally, paragenophores (an individual putatively conspecific with a hologenophore) were included in our analyses, because no mixed infections, i.e., two different parasite species (genotypes in our case) in a single host individual were recorded. The vouchers were deposited in the Helminthological collection of the Institute of Parasitology, Biology Centre of the Czech Academy of Sciences, České Budějovice, Czech Republic (IPCAS C-869–872).
Isolation of genomic DNA and amplification of two nuclear genes (18S rRNA and 28S rRNA) followed Olson et al.40. The partial COI gene was amplified by newly designed GcotCO1F (5′-ACTTTAGATCATAAGCGTATTGG-3′) and GcotCO1R1 (5′-AGCATAGTTATACCAGCAGC-3′) primers (courtesy of H. Hansen, Norwegian Veterinary Institute, Oslo, Norway); PCR was initiated by denaturation at 95 °C for 5 min, followed by 35 cycles of 95 °C for 1 min, 50 °C for 1 min, and 72 °C for 90 s, and terminated at 72 °C for 5 min. The amplicons were enzymatically purified41 and Sanger sequenced using PCR primers; nuclear genes were also sequenced using internal primers40,42. Contiguous sequences were aligned and inspected for ambiguous bases using Geneious43. In total, 26 original sequences of gyrocotylideans and Chimaera spp. were submitted to the GenBank database (accession numbers in Table 1).
Phylogenetic analyses were computed based on two concatenated datasets of our original sequences and those available from GenBank. The sequences of three genes were separately aligned by E-INS-I (18S rRNA and 28S rRNA) and L-INS-I (COI) algorithms in MAFFT ver.7.38844; the alignments were inspected by eye in Geneious and doubtful positions were removed. Substitution models for 18S rRNA (GTR + F + I), 28S rRNA (TVM + F + I + G4) and COI (TIM3 + F + I, HKY + F, and K3Pu + F + G4 for the 1st, 2nd, and 3rd codon positions, respectively) gene alignments were estimated under AICc criterion with an implemented test of model violation in ModelFinder using IQ-Tree ver. 2.0.545,46,47. IQ-Tree was also employed in the construction of phylogenetic trees under the Maximum Likelihood method with ultrafast bootstrapping (1000 replicates)48. These methods were also applied for host molecular identification employing TIM3e + G4, F81 + F, and TIM2 + F + G4 substitution models for the 1st, 2nd, and 3rd COI codon positions, respectively.
Data availability
The datasets generated during the current study are available from the corresponding author upon request.
References
Waeschenbach, A. & Littlewood, D. T. J. A molecular framework for the Cestoda. In Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth Vol. 25 (eds Caira, J. N. & Jensen, K.) 431–451 (University of Kansas, 2017).
Gibson, D. I. Order Gyrocotylidea Poche, 1926. In Keys to the Cestode Parasites of Vertebrates (eds Khalil, L. F. et al.) 21–43 (CAB International, 1994).
Kuchta, R., Scholz, T. & Hansen, H. Gyrocotylidea Poche, 1926. In Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth Vol. 25 (eds Caira, J. N. & Jensen, K.) 191–199 (University of Kansas, 2017).
Froese, R. & Pauly, D. Fishbase. World Wide Web Electronic Publication (2020) (accessed December 2020); www.fishbase.org.
Klimpel, S., Busch, M. W., Kellermanns, E., Kleinertz, S. & Palm, H. W. Metazoan Deep-Sea Fish Parasites 383 (Verlag Natur & Wissenschaft, 2009).
Bray, R. A., Waeschenbach, A., Littlewood, D. T. J., Halvorsen, O. & Olson, P. D. Molecular circumscription of new species of Gyrocotyle Diesing, 1850 (Cestoda) from deep-sea chimaeriform holocephalans in the North Atlantic. Syst. Parasitol. 97, 285–296. https://doi.org/10.1007/s11230-020-09912-w (2020).
Wagener, G. R. Ueber einen neuen in der Chimaera monstrosa gefunden Eingeweide-Wurm, Amphiptyches urna Grube and Wagener. Arch. Anat. Physiol. Wiss. Med. 543–554 (1852).
Fuhrmann, O. Dritte Klasse der Cladus Plathelminthes. Cestoidea. In Handbuch der Zoologie Vol. 2 (eds Kükenthal, W. & Krumbach, T.) 141–416 (Walter de Gruyter and Co, 1931).
van der Land, J. & Dienske, H. Two new species of Gyrocotyle (Monogenea) from chimaerids (Holocephali). Zool. Med. 43, 97–105 (1968).
van der Land, J. & Templeman, T. Two new species of Gyrocotyle (Monogenea) from Hydrolagus affinis (Brito Capello) (Holocephali). J. Fish. Res. Board Can. 25, 2365–2385 (1968).
Diesing, K. M. Systema Helminthum (W. Braumüller, Vindobonae, 1850).
MacDonagh, E. J. Parásitos de peces comestibles. III—Dos cestodarios: Gyrocotyle rugosa del “Pez gallo” y Gyrocotyle máxima n. sp. del “Gatuso”. La Semana Méd. Buenos Aires 34, 1232–1235 (1927).
Haswell, W. A. On a Gyrocotyle from Chimaera ogilbyi and on Gyrocotyle in general. Proc. Linn. Soc. 27, 48–55 (1902).
Ichihara, A. Gyrocotyle fimbriata Watson, 1911 from Chimaera fantasma in Suruga Bay. Jpn. J. Parasitol. 25, 13 (1976).
Watson, E. E. The genus Gyrocotyle and its significance for problems of cestode structure and phylogeny. Univ. Calif. Publ. Zool. 6, 353–468 (1911).
Bandoni, S. M. & Brooks, D. R. Revision and phylogenetic analysis of the Gyrocotylidea Poche, 1926 (Platyhelminthes: Cercomeria: Cercomeromorpha). Can. J. Zool. 65, 2369–2389 (1987).
Olson, P. D., Littlewood, D. T. J., Bray, R. A. & Mariaux, J. Interrelationships and evolution of the tapeworms (Platyhelminthes: Cestoda). Mol. Phylogenet. Evol. 19, 443–467. https://doi.org/10.1006/mpev.2001.0930 (2001).
Olson, P. D. & Caira, J. N. Evolution of the major lineages of tapeworms (Platyhelminthes: Cestoidea) inferred from 18S ribosomal DNA and elongation factor-1 alpha. J. Parasitol. 85, 1134–1159. https://doi.org/10.2307/3285679 (1999).
Waeschenbach, A., Webster, B. L. & Littlewood, D. T. J. Adding resolution to ordinal level relationships of tapeworms (Platyhelminthes: Cestoda) with large fragments of mtDNA. Mol. Phylogenet. Evol. 63, 834–847. https://doi.org/10.1016/j.ympev.2012.02.020 (2012).
Lockyer, A. E., Olson, P. D. & Littlewood, D. T. J. Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): Implications and a review of the cercomer theory. Biol. J. Linn. Soc. 78, 155–171. https://doi.org/10.1046/j.1095-8312.2003.00141.x (2003).
Williams, H. H., Colin, J. A. & Halvorsen, O. Biology of gyrocotylideans with emphasis on reproduction, population ecology and phylogeny. Parasitology 95, 173–207. https://doi.org/10.1017/s0031182000057656 (1987).
Bristow, G. A. On the distribution, ecology and evolution of Gyrocotyle urna, G. confusa and G. nybelini (Cercomeromorpha: Gyrocotylidea) and their host Chimaera monstrosa (Holocephalida: Chimaeridae) in Norwegian waters, with a review of the species question. Sarsia 77, 119–124 (1992).
Karlsbakk, E., Aspholm, P. E., Berg, V., Hareide, N. R. & Berland, B. Some parasites of the small-eyed rabbitfish, Hydrolagus affinis (Capello, 1867) (Holocephali), caught in deep waters off SW Greenland. Sarsia 87, 179–184. https://doi.org/10.1080/003648202320205256 (2002).
Manter, H. W. Studies on Gyrocotyle rugosa Diesing 1850, a cestodarian parasite of the elephant fish, Callorhynchus milii. Zool. Publ. Victoria Univ. Wellington 1, 1–11 (1951).
Pleijel, F. et al. Phylogenies without roots? A plea for the use of vouchers in molecular phylogenetic studies. Mol. Phylogenet. Evol. 48, 369–371. https://doi.org/10.1016/j.ympev.2008.03.024 (2008).
Simmons, J. E. Gyrocotyle: A century-old enigma. In Symbiosis in the Sea (ed. Vernberg, W. B.) 195–218 (University of South Carolina Press, 1974).
Colin, J. A., Williams, H. H. & Halvorsen, O. One or three gyrocotylideans (Platyhelminthes) in Chimaera monstrosa (Holocephali)?. J. Parasitol. 72, 10–21 (1986).
Lynch, J. E. Redescription of the species of Gyrocotyle from the ratfish, Hydrolagus colliei (Lay and Bennet), with notes on the morphology and taxonomy of the genus. J. Parasitol. 31, 418–446 (1945).
Allison, F. R. & Coakley, A. The two species of Gyrocotyle in the elephant fish, Callorhynchus milii (Bory). J. R. Soc. N. Z. 3, 381–392 (1973).
Wardle, R. A. The Cestoda of Canadian fishes II. The Hudson Bay drainage system. Contrib. Can. Biol. Fish. (New Ser.) 7, 377–403 (1932).
Didier, D. A. Phylogenetic systematics of extant chimaeroid fishes (Holocephali, Chimaeroidei). Am. Mus. Novit. 3119, 1–86 (1995).
Monticelli, F. Notes on some Entozoa in the collection of the British Museum. Proc. Zool. Soc. of Lond. 321–325 (1889).
Euzet, L. & Combes, C. Les problèmes de l’espèce chez les animaux parasites. Mém. Soc. Zool. France 40, 239–285 (1980).
Fyler, C. A. & Caira, J. N. Phylogenetic status of four new species of Acanthobothrium (Cestoda: Tetraphyllidea) parasitic on the wedgefish Rhynchobatus laevis (Elasmobranchii: Rhynchobatidae): Implications for interpreting host associations. Invertebr. Syst. 24, 419–433. https://doi.org/10.1071/IS10034 (2010).
Bueno, V. M. & Caira, J. N. Redescription and molecular assessment of relationships among three species of Echeneibothrium (Rhinebothriidea: Echeneibothriidae) parasitizing the yellownose skate, Dipturus chilensis, Chile. J. Parasitol. 103, 268–284. https://doi.org/10.1645/16-177 (2017).
Alves, P. V., de Chambrier, A., Luque, J. L. & Scholz, T. Integrative taxonomy reveals hidden cestode diversity in Pimelodus catfishes in the Neotropics. Zool. Scr. 50, 210–224. https://doi.org/10.1111/zsc.12465 (2021).
Ebert, D. A. et al. An annotated checklist of the chondrichthyans of Taiwan. Zootaxa 3752, 279–386. https://doi.org/10.11646/zootaxa.3752.1.17 (2013).
Holmes, B. H., Steinke, D. & Ward, R. D. Identification of shark and ray fins using DNA barcoding. Fish Res. 95, 280–288. https://doi.org/10.1016/j.fishres.2008.09.036 (2009).
Oros, M., Scholz, T., Hanzelová, V. & Mackiewicz, J. S. Scolex morphology of monozoic cestodes (Caryophyllidea) from the Palaearctic region: A useful tool for species identification. Folia Parasitol. 57, 37–46 (2010).
Olson, P. D., Cribb, T. H., Tkach, V. V., Bray, R. A. & Littlewood, D. T. J. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 33, 733–755. https://doi.org/10.1016/S0020-7519(03)00049-3 (2003).
Werle, E., Schneider, C., Renner, M., Völker, M. & Fiehn, W. Convenient single-step, one tube purification of PCR products for direct sequencing. Nucl. Acids Res. 22, 4354–4355 (1994).
Littlewood, D. T. J., Curini-Galletti, M. & Herniou, E. A. The interrelationships of Proseriata (Platyhelminthes: Seriata) flatworms tested with molecules and morphology. Mol. Phylogenet. Evol. 16, 449–466 (2000).
Kearse, M. et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649 (2012).
Katoh, K. & Standley, D. M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 30, 772–780 (2013).
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., von Haeseler, A. & Jermiin, L. S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 14, 587–589. https://doi.org/10.1038/nmeth.4285 (2017).
Minh, B. Q. et al. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37, 1530–1534. https://doi.org/10.1093/molbev/msaa015 (2020).
Naser-Khdour, S., Minh, B. Q., Zhang, W., Stone, E. A. & Lanfear, R. The prevalence and impact of model violations in phylogenetic analysis. Genome Biol. Evol. 11, 3341–3352. https://doi.org/10.1093/gbe/evz193 (2019).
Hoang, D. T., Chernomor, O., von Haeseler, A., Minh, B. Q. & Vinh, L. S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 35, 518–522. https://doi.org/10.1093/molbev/msx281 (2018).
Littlewood, D. T. J., Rohde, K. & Clough, K. A. The phylogenetic position of Udonella (Platyhelminthes). Int. J. Parasitol. 28, 1241–1250. https://doi.org/10.1016/s0020-7519(98)00108-8 (1998).
Olson, P. D., Poddubnaya, L. G., Littlewood, D. T. J. & Scholz, T. On the position of Archigetes and its bearing on the early evolution of the tapeworms. J. Parasitol. 94, 898–904. https://doi.org/10.1645/Ge-1456.1 (2008).
Acknowledgements
We are obliged to Shin-Pin Huang (Biodiversity Research Museum, Academia Sinica, Taiwan) for his help with fish identification, to Jesús S. Hernández-Orts for providing specimens from Argentina, to Haakon Hansen (Norwegian Veterinary Institute, Oslo, Norway) for providing his unpublished COI primers, and two anonymous reviewers for their helpful suggestions.
Funding
This research was financially supported by a bilateral research project between the Slovak Academy of Sciences and the Ministry of Sciences and Technology, Taiwan (Nos. SAS-MOST JRP 2016/7 & MOST 106-2923-B-038-001-MY3), the Grant Agency of the Ministry of Education of the Slovak Republic (VEGA No. 2/0126/20), the Czech Science Foundation (Project No. 19-28399X), and the Institute of Parasitology, Czech Academy of Sciences (RVO: 60077344).
Author information
Authors and Affiliations
Contributions
Mi.O., C.K.F. and D.B. conceived the study; Mi.O., D.B., P.S., C.K.F., R.K., Ma.O. and H.W.C. examined fish and collected parasites. D.B. performed molecular analyses, and D.B. and Mi.O. performed morphological examination. D.B., Mi.O., R.K. and T.S. prepared the draft of manuscript. All authors discussed the results and commented on the manuscript. All authors also revised the manuscript after its peer review.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Barčák, D., Fan, CK., Sonko, P. et al. Hidden diversity of the most basal tapeworms (Cestoda, Gyrocotylidea), the enigmatic parasites of holocephalans (Chimaeriformes). Sci Rep 11, 5492 (2021). https://doi.org/10.1038/s41598-021-84613-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-84613-y
This article is cited by
-
Land and deep-sea mining: the challenges of comparing biodiversity impacts
Biodiversity and Conservation (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
