
Abstract
Focusing attention on relevant information while ignoring distracting stimuli is essential to the efficacy of working memory. Alpha- and theta-band oscillations have been linked to the inhibition of anticipated and attentionally avoidable distractors. However, the neurophysiological background of the rejection of task-irrelevant stimuli appearing in the focus of attention is not fully understood. We aimed to examine whether theta and alpha-band oscillations serve as an indicator of successful distractor rejection. Twenty-four students were enrolled in the study. 64-channel EEG was recorded during a modified Sternberg working memory task where weak and strong (salient) distractors were presented during the retention period. Event-related spectral perturbation in the alpha frequency band was significantly modulated by the saliency of the distracting stimuli, while theta oscillation was modulated by the need for cognitive control. Moreover, stronger alpha desynchronization to strong relative to weak distracting stimuli significantly increased the probability of mistakenly identifying the presented distractor as a member of the memory sequence. Therefore, our results suggest that alpha activity reflects the vulnerability of attention to distracting salient stimuli.
Similar content being viewed by others

Introduction
Visual working memory enables us to temporarily hold and use the information for further processing but is limited in capacity. A crucial factor in determining working memory capacity is the effective filtering of irrelevant and distracting stimuli1,2, even over the enhancement of task-relevant information3. Former studies suggested that alpha-band oscillations play an important role in distractor filtering4,5,6 and theta-band oscillations have been linked to successful memory manipulation and general cognitive control, which are also essential components of efficient filtering7,8. However, the neural mechanisms and the oscillatory hallmarks of distractor inhibition are not fully understood9.
Oscillatory activity in the alpha frequency band has been linked to top-down modulation of attention10,11 and distractor filtering by several studies4,5,6. Specifically, it has been suggested that alpha oscillation plays an important role in the top-down controlled gating of task-relevant information by the inhibition of task-irrelevant neural populations12,13 by rhythmic synchronization through coherence14. Studies supporting this notion report that the scalp distribution of alpha-band power follows the focus of spatial attention, by decreasing over task-relevant areas and increasing over brain regions that represent distracting information6,15,16,17,18,19, and confirmed a causal relationship between alpha-band oscillations and the attentional suppression of sensory information by using transcranial magnetic stimulation20. However, it has been pointed out recently, that many of the existing evidence that links alpha activity with spatial attention is compatible with an account in which alpha activity supports target selection via signal enhancement21.
Numerous former EEG studies confirmed that increasing cognitive demand enhances frontal theta oscillatory activity22,23,24,25,26. However, recent studies pointed out that theta oscillations over the frontal cortex might reflect the realization of the need for cognitive control in goal-directed high-level cognitive processes such as working memory retention7,27.
Recent studies pointed out that to clarify the role of oscillatory activity in distractor filtering it should be taken into account whether participants can shift their attentional focus before the appearance of the distractors or not, as this influences the filtering process, recruit distinct mechanisms and involve different underlying neural structures28,29,30. Most of the tasks used in previous studies used attentional guidance by presenting targets and distractors bilaterally around a central fixation point along with a cue indicating the to-be memorized hemifield while presenting distractors on the other side of the screen6,30,31,32.
In contrast, there is far less and mixed evidence about the role of theta and alpha-band oscillatory modulation in the rejection of task-irrelevant stimuli when distractors cannot be attentionally avoided29,30,32. Moreover, in most of the previous studies, targets and distractors were displayed simultaneously, and only a few of them applied a sequential presentation33 or used a design, that allows the dissociation of the oscillatory activity related to the processing of targets and distractors34. Furthermore, there are very few studies that took into account that the saliency of the distracting stimuli might potentially influence the oscillatory dynamics32.
We aimed to examine directly whether theta and alpha-band oscillations serve as an indicator of successful distractor rejection. We applied the Sternberg working memory task with independent manipulation of memory set size and distractor saliency, which allows the temporal separation of the encoding and the retention condition including the presentation of distractors. Weak and strong (salient) distractors following memory sequences were presented at the same location as the targets to keep them in the focus of attention.
As previous studies suggest, that cognitive control-related modulations affect the induced oscillations7,35 we analyzed the event-related spectral perturbation (ERSP), which can capture the non-phase locked as well as the evoked oscillations36.
Based on previous research we assumed that the salient distracting stimuli will induce enhanced alpha29,32 and theta oscillations32 compared to the weak (non-salient) distractors, while the increased need for cognitive control related to the rejection of distractors and especially related to the inhibition of salient distractors will enhance theta oscillatory activity, especially in the frontal region7,27,35.
Materials and methods
Participants
The study was carried out in the Department of Psychiatry and Psychotherapy, Semmelweis University, Budapest, Hungary. Altogether 24 students of Semmelweis University (10 females, mean age 24 ± 2.15 years) were enrolled in the study. All participants were right-handed except for one ambidextrous subject. Selection criteria were no history of head injury, stroke, mental retardation, epileptic seizure, acute psychiatric disorder, or substance dependence. All procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Declaration of Helsinki. The National Scientific and Ethical Committee, Budapest, Hungary approved the research. Participants gave their written informed consent before the procedures.
EEG stimuli and procedures
During the EEG recording, participants were seated in a dimly lit, sound-attenuated room. All participants had normal or corrected-to-normal vision. Subjects performed a modified version of the Sternberg task37, an extensively used test of working memory38,39. The original task was adjusted to assess the effect of distractors on working memory retention. The stimuli consisted of white, green, and blue letters, Arial font, point size 60, that were presented on a computer screen at approximately 50 cm with Presentation 13.0 software (Neurobehavioral Systems, Inc.; Albany, CA).
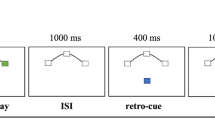
At the onset of each trial, two or six randomly selected white consonants appeared sequentially in the middle of the screen for 1200 ms, separated by a blank screen for 150–250 ms. During the retention period, two green-colored distractors were presented in a randomized order for 1200 ms separated by a blank screen for 1800–2000 ms: a green exclamation mark (weak distractor) and a green consonant that was not part of the previous learning sequence (strong distractor). In the retrieval condition, two blue consonants were presented for 1200 ms separated by a blank screen presented for 1800–2000 ms (Fig. 1). Subjects were instructed to indicate by clicking the mouse buttons (yes-right / no-left) whether the probe letter was part of the presented sequence. The length of the learning sequence, the order of the distractors, and the response assignment were counterbalanced across 144 trials. Efficiency was measured by response accuracy. Working memory capacity was estimated using Cowan's K formula40 for each set-size: K = set size × (hit rate – false alarm rate). Participants completed the task in six equal parts separated by a 3-min rest period. It was carefully monitored that the participants understood the instructions and stayed alert during the session.

The modified Sternberg working memory task. Following the memory sequence, during the retention period of the task, a strong (salient, green consonant) and a weak (non-salient, green exclamation mark) distractor was presented in a randomized order. The Figure was prepared by using Adobe Illustrator CC 2018 software.
EEG recording and processing
EEG was recorded from DC with a low-pass filter at 100 Hz using a high-density 64-channel BioSemi ActiveTwo amplifier41. Electrode caps had an equidistant-layout and covered the whole head. Eye movements were monitored with EOG electrodes placed below the left and above the right external canthi. Data were digitized at a sampling rate of 1024 Hz. Built-in and self-developed functions as well as the freeware EEGLAB toolbox42 in the MATLAB (MathWorks, Natick, MA) development environment were used for subsequent off-line data analyses. EEG was re-referenced to the common average potential and filtered off-line between 0.5 and 45 Hz using zero-phase shift forward, and reverse IIR Butterworth filter.
Epochs from 500 ms pre-stimulus to 1350 ms post-stimulus were extracted from the continuous EEG and corrected for the pre-stimulus baseline adjusted to the presentation of the last items of the learning sequences and of the weak and strong distractors during the retention period.
Removal of muscle, blinking, and eye movement artifacts (detected by EOG) were performed by ADJUST43, and ICA (Independent Component Analysis) based automatic artifact detector. Furthermore, epochs with a voltage exceeding ± 100 μV on any channel were rejected from the analysis.
After artifact rejection, the average number of trials was 132.2 (SD = 11.2) for the learning, 132.3 (SD = 11.0) for the weak distractor, and 132.1 (SD = 13.3) for the strong distractor condition, respectively. For the detailed analysis, we selected 4 scalp regions (frontal, central, right parieto-occipital, left parieto-occipital).
EEG data analysis
Stimulus-related theta (4.1–7 Hz) and alpha (7.1–13 Hz) activity changes were measured by the ERSP, providing a 2-D representation of the mean change in spectral power (in dB) from baseline36. Beyond conventional event-related analytical approaches, ERSP can capture event-related as well as non-phase locked (induced) oscillatory activity. Since distractor filtering is a relatively time-consuming complex cognitive process involving the cooperation of distant cortical regions28 and especially, because it has been shown that cognitive control-related processes modulate the induced oscillatory activity7,35 we applied the ERSP approach in our analysis. To compute the ERSP, baseline spectra are calculated from the EEG immediately preceding each event. The epoch is divided into brief, overlapping data windows, and a moving average of the amplitude spectra is created. Each of these spectral transforms of individual response epochs is then normalized by dividing by their respective mean baseline spectra. Normalized response transforms for many trials are then averaged to produce the average ERSP, plotted as relative spectral log amplitude on a time-by-frequency plane36 (see details in the Supplementary Information).
The analysis was performed on epochs extending from 500 ms before to 1350 ms after stimulus onset in learning, weak distractor, and strong distractor condition, respectively in the 1.5–30 Hz frequency range. The sliding window was 400 ms wide, and it was applied 200 times with an average step size of 6.8 ms. No zero padding was applied. The analyzed time interval lasted from 282 ms before to 1133 ms after stimulus onset. Dynamical changes in oscillatory activity were studied by computing ERSPs for each trial, then averaging them separately for learning, weak distractor, and strong distractor condition44,45.
The ERSP time–frequency matrices were baseline corrected by the average power calculated from the 500 to 200 ms pre-stimulus period for the assessment of the modulatory effect of distractor saliency and memory load to oscillatory activity during the retention period. To compare ERSP across conditions a second analysis was conducted, using the average power calculated from the 500 to 200 ms period before the first item of the learning sequences as a common baseline for the retention and learning conditions. In this analysis, epochs of only the last items of the learning sequences were included to minimize memory load-related differences between the conditions. Note that in this case, baseline values did not originate from the pre-stimulus period and for this reason, the sum of the baseline data points is not necessarily zero. However, thus we could exclude the possibility of finding differences in ERSP caused by differences in baseline activity in the learning and retention period.
Former studies found memory load-related and distractor saliency-related modulation of the alpha activity from 500 to 1300 ms following the stimulus presentation29,30,46. Therefore, based on these prior results and adjusted to the end of the desynchronization in the weak distractor condition we selected the 700–1000 ms time window in the alpha band for further analysis to select a time window past the initial encoding of the stimulus. For the assessment of the theta ERSP, we selected the 300–600 ms time window, as previous studies detected cognitive control-related midfrontal theta power modulation in this time window35,47.
Mean ERSP values were calculated by averaging across electrodes within scalp regions to further attenuate noise. As we aimed to identify and highlight robust distractor rejection-related modulation of electrophysiological activity, every scalp region was involved in the regional analysis. Parieto-occipital region was divided to right and left side to avoid the conflation of the results due to averaging across too many electrodes and since the task-related stimuli were letters, which are generally processed in brain areas located in the left hemisphere48,49,50, which may cause the lateralization of the task-related modulation51,52. To avoid the loss of information due to the aggregation of multiple electrodes, and based on prior studies of cognitive control that selectively examined frontal-midline electrodes27,35,53 we also analyzed the theta activity of the Fz electrode separately.
Statistical analysis
Behavioral results between the 2- and 6-item conditions were compared with paired t-test. The different effects on ERSP in the distractor conditions were tested by three-way analyses of variance (ANOVA) of the distractor type (strong vs. weak) × memory load (2-item vs. 6-item) × region while for the comparison of the learning and distractor conditions two-way ANOVA of the condition (learning vs. strong distractor vs. weak distractor) × region was used. Normal distribution of variables was tested using the Kolmogorov–Smirnov test. Post-hoc pairwise contrasts were conducted to investigate the interactions. Since post-hoc comparisons were evaluated over four regions, Hochberg correction for multiple comparisons was applied to the post-hoc contrasts54,55. To characterize the magnitude of the reported effects we reported the values of effect size56.
The associations between the behavioral results and the task-related modulation of ERSP in the frontal region were assessed by Pearson correlation. Alpha reactivity was characterized by the difference of alpha ERSP in the weak and strong distractor condition (alpha ERSPweak distractor—alpha ERSPstrong distractor) in the 700–1000 ms time window, while theta reactivity was marked by the difference of theta ERSP in the learning and strong distractor condition in the 300–600 ms time window (theta ERSPlearning—theta ERSPstrong distractor).
In the correlational analysis, we focused on the frontal region. Former fMRI studies showed that distraction suppression activates the prefrontal cortex specifically57,58,59,60. Moreover, former EEG studies identified the prefrontal cortex as the main source of top-down attentional control of visual cortical activity during working memory retention mediated by alpha oscillations61 and the origin of theta activity which is modulated by the need for cognitive control7,27, therefore we hypothesized that the task-related modulations of frontal alpha- and theta-band oscillations will reflect the regulation of attentional and cognitive control.
Figures were prepared by using MATLAB and Statistics Toolbox Release 2017a and SAS 9.4 software.
Results
Behavioral results
Response accuracy of the participants was significantly decreased under high memory load compared to low memory load (mean2-items = 96.25% SD = 3.25, mean6-items = 92.02% SD = 5.06, t1,23 = 5.9, p < 0.0001, Cohens’s D = 1.0). Furthermore, subjects made significantly more “distractor commission error” (i.e. participant mistakenly identifies the previously presented distractor as a member of the memory sequence) under high memory load, when they mistakenly identified the strong distractor as part of the learning sequence (mean2-item, distractor commission error = 1.27% SD = 1.14, mean6-item, distractor commission error = 2.50% SD = 2.09, t1,23 = − 2.84, p = 0.009, Cohens’s D = 0.8).
Alpha ERSP in weak and strong distractor conditions
Alpha desynchronization (ERD: decrease in alpha ERSP) was observable (Fig. 2) during the presentation of weak as well as of strong distractors, however, a significantly stronger alpha ERD (F(1,23) = 27.4, p < 0.0001, Cohen’s D = 1.0) was detected in the strong distractor condition compared to the weak distractor condition. Furthermore, alpha ERD was significantly weaker following sequences with high memory load in contrast to low memory load (F(1,23) = 10.8, p = 0.0032, Cohen’s D = 0.3). Moreover, region had a significant effect on alpha ERD (F(3,23) = 11.4, p < 0.0001) with maximum alpha ERD in the right parieto-occipital region during the strong distractor conditions following a low memory load set. Interaction of distractor type and region (F(3,23) = 4.4, p = 0.01) and memory load and region (F(3,23) = 5.1, p = 0.007) had a significant effect as well. Post hoc analyses of these interactions revealed significant memory load-related modulation of alpha ERSP (ERDlow memory set > ERDhigh memory set) in the central (t = 4.1, df = 23, p = 0.0004, Cohen’s D = 0.3) and left parieto-occipital (t = 4.3, df = 23, p = 0.0003, Cohen’s D = 0.4) region, and significant distractor type-related modulation of alpha ERSP (ERDstrong distractor > ERDweak distractor) in every region (t > 3.8, p < 0.05, Cohen’s D > = 0.9), with a maximum difference between distractor types at the central region (t = 7.0, p < 0.0001, Cohen’s D = 1.2) These differences remained significant even after correction for multiple comparisons.

Alpha ERSP in strong and weak distractor conditions. Alpha event-related spectral perturbation (ERSP) in strong and weak distractor conditions (700–1000 ms time window highlighted) and scalp topography of the alpha ERSP groups in the 700–1000 ms time window. The Figures was prepared by using MATLAB and Statistics Toolbox Release 2017a.
Alpha ERSP in learning and distractor conditions
Condition had a significant effect on alpha ERSP (F(2,23) = 19.4, p < 0.0001). Significantly decreased alpha ERSP was found compared to the weak distractor condition in the learning (t = 5.7, df = 23, p < 0.0001, Cohen’s D = 0.4) and in the strong distractor condition (t = 5.3, df = 23, p < 0.0001, Cohen’s D = 0.4), while there was no significant difference in alpha ERSP between the learning and strong distractor conditions (p > 0.05, Cohen’s D = 0.01). Region (F(3,23) = 107.8, p < 0.0001) and interaction of condition and region (F(6,23) = 14.2, p = 0.0001) had a significant effect as well. Post hoc analyses of this interactions revealed significant increase of alpha ERSP during the weak distractor condition relative to the learning and strong distractor condition in every region (t > 3.43, p ≤ 0.0002) with a maximum difference between learning and the weak distractor condition at the right parieto-occipital region (t = 7.23, p < 0.0001, Cohen’s D = 0.6), which remained significant after correction for multiple comparisons, while alpha ERSP in the learning and strong distractor condition did not show difference in any region (p > 0.05) (Supplementary Fig. 2).
Correlational analysis of distractor related alpha oscillatory activity
To explore the relation between attentional control-related modulation of frontal alpha oscillatory activity and task performance, we characterized distractor saliency-related alpha reactivity by the difference of ERSP in the strong and weak distractor condition. Alpha reactivity in the frontal region correlated significantly with mean working memory capacity (kmean, r = 0.44, p = 0.03). Moreover, significant negative correlations were found between the number of “distractor commission errors” and alpha reactivity in the frontal region (r = − 0.47, p = 0.02) (Fig. 4) However, these results did not remain significant after correction for multiple comparisons.
Theta ERSP in weak and strong distractor conditions
Theta synchronization (ERS: increase in theta ERSP) was observable during the presentation of weak as well as of strong distractors, however, theta ERS did not differ significantly in the strong and weak distractor conditions in the 300–600 ms time window (p > 0.05, Cohen’s D = 0.14). Moreover, memory load did not have a significant modulatory effect on theta ERS (p > 0.5, Cohen’s D = 0.10) (Supplementary Fig. 1).
Theta ERSP in learning and distractor conditions
Condition (learning vs weak distractor vs strong distractor) had a significant effect on theta (4–7 Hz) ERSP (F(2,23) = 30.0, p < 0.0001) (Fig. 3). Significantly increased theta ERSP was found during the weak (t = 5.7, df = 23, p < 0.0001, Cohen’s D = 0.3) and the strong distractor condition (t = 6.7, df = 23, p < 0.0001, Cohen’s D = 0.3) compared to the learning condition, while there was no significant difference in theta ERSP between the weak and strong distractor conditions (p > 0.05 Cohen’s D = 0.06). Region (F(3,23) = 43.9, p < 0.0001) and interaction of condition and region (F(6,23) = 7.9, p = 0.0001) had a significant effect as well. Post hoc analyses of this interactions revealed significant increase of theta ERSP during the distractor conditions relative the learning condition, regardless of the distractor type in every region (t ≥ 3.78, p ≤ 0.0001) with a maximum difference between learning and the strong distractor condition at the central region (t = 8.45, p < 0.0001, Cohen’s D = 0.4), which remained significant after correction for multiple comparisons, while theta ERSP in the weak and strong distractor condition did not show any difference in any region (p > 0.05).

Theta ERSP in learning, strong and weak distractor conditions. Theta event-related spectral perturbation (ERSP) in learning, strong and weak distractor conditions (300–600 ms time window highlighted), and scalp topography of the theta ERSP groups in the 300–600 ms time window. The Figure was prepared by using MATLAB and Statistics Toolbox Release 2017a.
We analyzed theta ERSP in the same time window selectively in the Fz electrode as well. Condition had a significant effect on frontal midline theta ERSP (F(2,23) = 27.9, p < 0.0001) (Fig. 3). Significantly higher theta ERSP was found in the strong distractor condition compared to the weak distractor (t = 3.2, df = 23, p < 0.0038, Cohen’s D = 0.2) and to the learning (t = 7.1, df = 23, p < 0.0001, Cohen’s D = 0.5) condition. Furthermore, in the weak distractor condition a significantly increased theta ERSP was detected compared to the learning condition (t = 5.3, df = 23, p < 0.0001, Cohen’s D = 0.3).
Effect of bottom-up differences of visual stimulation on the N1 ERP component and correlational analysis with the ERSP
To rule out that the detected distractor rejection-related modulatory effects were driven by differences in low-level features, we examined the N1 component which is sensitive to physical properties of the stimuli based on former studies62,63,64,65 and its correlation with alpha and theta ERSP. The physical differences between the letters and the exclamation mark did not generate a difference in the N1 component and alpha and theta ERSP did not correlate with the N1 component, suggesting that the differences of alpha and theta ERSP in the later time windows are not the consequences of differences in stimulus-driven activity. Detailed results of these analyses can be found in the Supplementary Information.
Correlational analysis of task-related theta oscillatory activity
To explore the relationship between the cognitive control-related modulation of frontal theta oscillatory activity and task performance, we characterized theta reactivity by the difference of ERSP in the learning and strong distractor condition. In these conditions the presented stimuli were similar except their color, indicating whether they should be learned or ignored. Theta reactivity in the frontal region showed a significant positive correlation with response accuracy (r = 0.42, p = 0.04) (Fig. 4). As one participant had extremely high theta reactivity, we performed the Spearman correlational analysis on the above-mentioned correlation as well, which is robust against the effect of outliers. The correlation between the theta ERSP and the hit rate became marginally significant (Spearman r = 0.39, p = 0.058). However, these results did not remain significant after correction for multiple comparisons.

Correlation of frontal alpha and theta reactivity with behavioral results. We characterized alpha reactivity by the difference of alpha ERSP in the weak and strong distractor condition in the 700–1000 ms time window (alpha ERSPweak distractor—alpha ERSPstrong distractor), while we characterized theta reactivity by the difference of theta ERSP in the learning and strong distractor condition in the 300–600 ms time window (theta ERSPlearning—theta ERSPstrong distractor). The Figure was prepared by using SAS 9.4 software.
Discussion
Alpha frequency band
During the retention period, regardless of the type of distractor, increased alpha ERSP was found following long sequences relative to short sequences. This memory load-related modulation of the alpha band is in line with many previous studies4,38,66. This effect remained significant in the central and left parieto-occipital region even after correction for multiple comparisons. The lateralization of memory load-related modulation is probably due to the phonological nature of the applied stimuli, which is generally processed in brain areas located in the left hemisphere48,49,50, which is also in line with former studies that used letters as memory items and detected more pronounced working memory load-related modulation in the left hemisphere51,52.
However, in contrast to the attributed inhibitory role of alpha enhancement during the increased need for distractor filtering6,13, alpha ERSP decreased with a higher level of distraction. Former studies examining alpha-band modulations during reactive filtering showed similar results29,32. Previous research on contingent capture showed, that salient distractors, that are similar to targets elicit an involuntary shift of spatial attention67,68. As the reduction of alpha power has been suggested as a marker of involuntary capture of attention69, attenuated alpha activity in the presence of a distractor that is highly similar to the targets might indicate the attraction of attention by the salient but distracting stimuli.
This interpretation is also supported by the positive correlation between alpha reactivity (i.e. stronger alpha ERD to strong relative to weak distractors) and the number of “distractor commission errors”, where participants mistakenly identified the previously presented distractor as a member of the memory sequence. This finding can be interpreted as the enhancing distraction of attention by the (stronger) distractor—indicated by a stronger ERD—increased the probability that it would be encoded as a member of the memory sequence, although we have to be aware that this correlation does not necessarily imply a causal relation. The alpha activity was mainly modulated by stimulus quality since stronger alpha ERD was observed in the presence of strong distractors relative to weak distractors (letters vs. exclamation mark), while no significant differences were found between the learning and strong distractor conditions (both stimuli were letters). This corresponds to a former study32, where reactive alpha modulation was present relative to target processing if the distractor had low similarity to the target stimulus, but not in the case of a highly similar distractor.
Therefore, in agreement with former studies29,30,32, our results suggest that in contrast to the inhibition of distractors outside of the attentional focus, alpha oscillatory activity is not directly involved in the rejection of task-irrelevant stimuli when they cannot be attentionally avoided.
Instead, decreasing and increasing alpha oscillatory activity might reflect the balance between externally and internally directed attention, respectively70,71,72. While alpha oscillatory power enhanced with the increasing internal processing demand of higher memory load, it attenuated (i.e. stronger ERD) with the increasing saliency of distractors and therefore the extent of involuntary capture of attention. Therefore, although alpha oscillations might serve as a top-down control mechanism to protect working memory maintenance4, the bottom-up interference of unexpected salient stimuli might attenuate this protection.
Theta frequency band
Theta ERSP was significantly increased during the presentation of weak and strong distractors relative to the learning period. Frontal theta oscillation is an important marker of working memory processing24. Increased frontal theta oscillation has been consequently observed during working memory tasks related to increasing task demands22,23,24, but it has been also considered as a sign of successful memory manipulation, novelty, and conflict detection8,73. Therefore, recently it has been suggested, that frontal theta oscillatory activity is a potential general marker of cognitive control in tasks that involve uncertainty about actions and outcomes7,27. According to this framework, increased theta activity in response to unexpected distractors might signal a need for enhanced cognitive control during reactive filtering7.
Our results showed, that the appearance of weak and strong distractors were both accompanied by increased theta ERSP compared to the learning condition, which suggests that theta modulation is related to the general control processes during the task. Moreover, in line with former results32, salient distractors induced increased frontal midline theta activity compared to the non-salient distractor, which corroborates that theta modulation is related to the need for cognitive control7. However, this modulatory effect was not a robust phenomenon, as at the regional level it did not reach statistical significance.
The connection between theta modulation and cognitive control is also supported by the correlation between response accuracy and frontal theta reactivity related to reactive filtering, as the prefrontal cortex has been suggested as an important source of cognitive control in working memory tasks74.
Limitations
There are some possible limitations of this study. First, our sampling method was non-random, as the participants were all university students, which limits the generalizability of our results. Moreover, the correlational analysis was done by following a ROI-based approach which can be a possible limiting factor as well.
Furthermore, although effect size and sample size are theoretically independent, the small sample size of our study might inflate effect sizes75, which is a further limitation. Therefore, further studies applying similar paradigms on larger and more representative samples are required to verify our preliminary results.
Conclusion
Distractor rejection is accompanied by distinct underlying dynamics of oscillatory activity in the alpha and theta frequency band. Alpha ERSP is mainly modulated by the saliency of the presented stimuli. However, while alpha activity is not directly involved in the filtering mechanism, it reflects the vulnerability of attention to distracting salient stimuli.
In contrast to that, theta ERSP is modulated by the need for cognitive control when faced with unexpected distractors.
Furthermore, our results underline the need for the distinction of proactive and reactive filtering, as these processes are reflected by different oscillatory dynamics, regardless of their common intention30, although further studies applying similar paradigms are required to verify our results.
Data availability
The datasets that are used and analyzed during the current study are available from the corresponding author on reasonable request.
References
Fukuda, K. & Vogel, E. K. Human variation in overriding attentional capture. J. Neurosci. Off. J. Soc. Neurosci. 29, 8726–8733. https://doi.org/10.1523/jneurosci.2145-09.2009 (2009).
Kane, M. J., Conway, A. R. A., Hambrick, D. Z. & Engle, R. W. Variation in Working Memory 21–46 (Oxford University Press, Oxford, 2007).
Gulbinaite, R., Johnson, A., de Jong, R., Morey, C. C. & van Rijn, H. Dissociable mechanisms underlying individual differences in visual working memory capacity. NeuroImage 99, 197–206. https://doi.org/10.1016/j.neuroimage.2014.05.060 (2014).
Bonnefond, M. & Jensen, O. Alpha oscillations serve to protect working memory maintenance against anticipated distracters. Curr. Biol. 22, 1969–1974. https://doi.org/10.1016/j.cub.2012.08.029 (2012).
Freunberger, R., Werkle-Bergner, M., Griesmayr, B., Lindenberger, U. & Klimesch, W. Brain oscillatory correlates of working memory constraints. Brain Res. 1375, 93–102. https://doi.org/10.1016/j.brainres.2010.12.048 (2011).
Sauseng, P. et al. Brain oscillatory substrates of visual short-term memory capacity. CB 19, 1846–1852. https://doi.org/10.1016/j.cub.2009.08.062 (2009).
Cavanagh, J. F. & Frank, M. J. Frontal theta as a mechanism for cognitive control. Trends Cognit. Sci. 18, 414–421. https://doi.org/10.1016/j.tics.2014.04.012 (2014).
Itthipuripat, S., Wessel, J. R. & Aron, A. R. Frontal theta is a signature of successful working memory manipulation. Exp. Brain Res. 224, 255–262. https://doi.org/10.1007/s00221-012-3305-3 (2013).
Gazzaley, A. & Nobre, A. C. Top-down modulation: bridging selective attention and working memory. Trends Cognit. Sci. 16, 129–135. https://doi.org/10.1016/j.tics.2011.11.014 (2012).
Sauseng, P. et al. EEG alpha synchronization and functional coupling during top-down processing in a working memory task. Hum. Brain Mapp. 26, 148–155. https://doi.org/10.1002/hbm.20150 (2005).
Palva, S. & Palva, J. M. New vistas for alpha-frequency band oscillations. Trends Neurosci. 30, 150–158. https://doi.org/10.1016/j.tins.2007.02.001 (2007).
Jensen, O. & Mazaheri, A. Shaping functional architecture by oscillatory alpha activity: gating by inhibition. Front Hum Neurosci 4, 186. https://doi.org/10.3389/fnhum.2010.00186 (2010).
Klimesch, W., Sauseng, P. & Hanslmayr, S. EEG alpha oscillations: the inhibition-timing hypothesis. Brain Res. Rev. 53, 63–88. https://doi.org/10.1016/j.brainresrev.2006.06.003 (2007).
Fries, P. Rhythms for cognition: communication through coherence. Neuron 88, 220–235. https://doi.org/10.1016/j.neuron.2015.09.034 (2015).
Klimesch, W. Alpha-band oscillations, attention, and controlled access to stored information. Trends Cognit. Sci. 16, 606–617. https://doi.org/10.1016/j.tics.2012.10.007 (2012).
Payne, L., Guillory, S. & Sekuler, R. Attention-modulated alpha-band oscillations protect against intrusion of irrelevant information. J. Cognit. Neurosci. 25, 1463–1476. https://doi.org/10.1162/jocn_a_00395 (2013).
Snyder, A. C. & Foxe, J. J. Anticipatory attentional suppression of visual features indexed by oscillatory alpha-band power increases: a high-density electrical mapping study. J. Neurosci. Off. J. Soc. Neurosci. 30, 4024–4032. https://doi.org/10.1523/jneurosci.5684-09.2010 (2010).
Handel, B. F., Haarmeier, T. & Jensen, O. Alpha oscillations correlate with the successful inhibition of unattended stimuli. Journal of cognitive neuroscience 23, 2494–2502. https://doi.org/10.1162/jocn.2010.21557 (2011).
Payne, L. & Sekuler, R. The importance of ignoring: alpha oscillations protect selectivity. Curr. Dir. Psychol. Sci. 23, 171–177. https://doi.org/10.1177/0963721414529145 (2014).
Romei, V., Gross, J. & Thut, G. On the role of prestimulus alpha rhythms over occipito-parietal areas in visual input regulation: correlation or causation?. J. Neurosci. 30, 8692–8697. https://doi.org/10.1523/jneurosci.0160-10.2010 (2010).
Foster, J. J. & Awh, E. The role of alpha oscillations in spatial attention: limited evidence for a suppression account. Curr. Opin. Psychol. 29, 34–40. https://doi.org/10.1016/j.copsyc.2018.11.001 (2019).
Jensen, O. & Tesche, C. D. Frontal theta activity in humans increases with memory load in a working memory task. Eur. J. Neurosci. 15, 1395–1399. https://doi.org/10.1046/j.1460-9568.2002.01975.x (2002).
Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: a review and analysis. Brain Res. Brain Res. Rev. 29, 169–195 (1999).
Hsieh, L. T. & Ranganath, C. Frontal midline theta oscillations during working memory maintenance and episodic encoding and retrieval. NeuroImage 85(Pt 2), 721–729. https://doi.org/10.1016/j.neuroimage.2013.08.003 (2014).
Onton, J., Delorme, A. & Makeig, S. Frontal midline EEG dynamics during working memory. NeuroImage. https://doi.org/10.1016/j.neuroimage.2005.04.014 (2005).
Raghavachari, S. et al. Gating of human theta oscillations by a working memory task. J. Neurosci. Off. J. Soc. Neurosci. https://doi.org/10.1523/JNEUROSCI.21-09-03175.2001 (2001).
Cavanagh, J. F., Zambrano-Vazquez, L. & Allen, J. J. Theta lingua franca: a common mid-frontal substrate for action monitoring processes. Psychophysiology 49, 220–238. https://doi.org/10.1111/j.1469-8986.2011.01293.x (2012).
Geng, J. J. Attentional mechanisms of distractor suppression. Current Direct. Psychol. Sci. 23, 147–153. https://doi.org/10.1177/0963721414525780 (2014).
Schroeder, S. C. Y., Ball, F. & Busch, N. A. The role of alpha oscillations in distractor inhibition during memory retention. Eur. J. Neurosci. 48, 2516–2526. https://doi.org/10.1111/ejn.13852 (2018).
Vissers, M. E., van Driel, J. & Slagter, H. A. Proactive, but not reactive, distractor filtering relies on local modulation of alpha oscillatory activity. J. Cognit. Neurosci. 28, 1964–1979. https://doi.org/10.1162/jocn_a_01017 (2016).
Fukuda, K., Kang, M. S. & Woodman, G. F. Distinct neural mechanisms for spatially lateralized and spatially global visual working memory representations. J. Neurophysiol. 116, 1715–1727. https://doi.org/10.1152/jn.00991.2015 (2016).
van Diepen, R. M., Miller, L. M., Mazaheri, A. & Geng, J. J. The role of alpha activity in spatial and feature-based attention. Neuro https://doi.org/10.1523/eneuro.0204-16.2016 (2016).
Hakim, N., Feldmann-Wüstefeld, T., Awh, E. & Vogel, E. K. Perturbing neural representations of working memory with task-irrelevant interruption. J. Cognit. Neurosci. 32, 558–569. https://doi.org/10.1162/jocn_a_01481 (2020).
Feldmann-Wüstefeld, T. & Vogel, E. K. Neural evidence for the contribution of active suppression during working memory filtering. Cereb. Cortex 29, 529–543. https://doi.org/10.1093/cercor/bhx336 (2019).
Cohen, M. X. & Donner, T. H. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. J. Neurophysiol. 110, 2752–2763. https://doi.org/10.1152/jn.00479.2013 (2013).
Makeig, S., Debener, S., Onton, J. & Delorme, A. Mining event-related brain dynamics. Trends Cognit. Sci. 8, 204–210. https://doi.org/10.1016/j.tics.2004.03.008 (2004).
Sternberg, S. High-speed scanning in human memory. Science 153, 652–654 (1966).
Jensen, O., Gelfand, J., Kounios, J. & Lisman, J. E. Oscillations in the alpha band (9–12 Hz) increase with memory load during retention in a short-term memory task. Cereb. Cortex 12, 877–882. https://doi.org/10.1093/cercor/12.8.877 (2002).
Karrasch, M. et al. Brain oscillatory responses to an auditory-verbal working memory task in mild cognitive impairment and Alzheimer’s disease. Int. J. Psychophysiol. 59, 168–178. https://doi.org/10.1016/j.ijpsycho.2005.04.006 (2006).
40Cowan, N. The magical number 4 in short-term memory: a reconsideration of mental storage capacity. Behav. Brain. Sci. 24, 87–114; discussion 114–185 (2001).
Metting van Rijn, A. C., Peper, A. & Grimbergen, C. A. High-quality recording of bioelectric events. Part 1. Interference reduction, theory and practice. Med. Biol. Eng. Comput. 28, 389–397 (1990).
Delorme, A. & Makeig, S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009 (2004).
Mognon, A., Jovicich, J., Bruzzone, L. & Buiatti, M. ADJUST: an automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology 48, 229–240. https://doi.org/10.1111/j.1469-8986.2010.01061.x (2011).
Herrmann, C. S., Munk, M. H. & Engel, A. K. Cognitive functions of gamma-band activity: memory match and utilization. Trends Cognit. Sci. 8, 347–355. https://doi.org/10.1016/j.tics.2004.06.006 (2004).
Tallon-Baudry, C. & Bertrand, O. Oscillatory gamma activity in humans and its role in object representation. Trends Cognit. Sci. 3, 151–162 (1999).
Heinz, A. J. & Johnson, J. S. Load-dependent increases in delay-period alpha-band power track the gating of task-irrelevant inputs to working memory. Frontiers Hum. Neurosci. 11, 250. https://doi.org/10.3389/fnhum.2017.00250 (2017).
Chinn, L., Pauker, C. & Golob, E. Cognitive control and midline theta adjust across multiple timescales. Neuropsychologia https://doi.org/10.1016/j.neuropsychologia.2018.01.031 (2018).
Awh, E. et al. Dissociation of storage and rehearsal in verbal working memory: evidence from positron emission tomography. Psychol. Sci. 7, 25–31. https://doi.org/10.1111/j.1467-9280.1996.tb00662.x (1996).
Strand, F., Forssberg, H., Klingberg, T. & Norrelgen, F. Phonological working memory with auditory presentation of pseudo-words—an event related fMRI Study. Brain Res. 1212, 48–54. https://doi.org/10.1016/j.brainres.2008.02.097 (2008).
McCandliss, B. D., Cohen, L. & Dehaene, S. The visual word form area: expertise for reading in the fusiform gyrus. Trends Cognit. Sci. 7, 293–299. https://doi.org/10.1016/s1364-6613(03)00134-7 (2003).
Gomarus, H. K., Althaus, M., Wijers, A. A. & Minderaa, R. B. The effects of memory load and stimulus relevance on the EEG during a visual selective memory search task: an ERP and ERD/ERS study. Clin Neurophysiol 117, 871–884. https://doi.org/10.1016/j.clinph.2005.12.008 (2006).
Jensen, O., Gelfand, J., Kounios, J. & Lisman, J. E. Oscillations in the alpha band (9–12 Hz) increase with memory load during retention in a short-term memory task. Cereb. cortex 12, 877–882. https://doi.org/10.1093/cercor/12.8.877 (2002).
Nurislamova, Y. M., Novikov, N. A., Zhozhikashvili, N. A. & Chernyshev, B. V. Enhanced Theta-band coherence between midfrontal and posterior parietal areas reflects post-feedback adjustments in the state of outcome uncertainty. Frontiers Integr. Neurosci. 13, 14–14. https://doi.org/10.3389/fnint.2019.00014 (2019).
Hochberg, Y. A sharper Bonferroni procedure for multiple tests of significance. Biometrika 75, 800–802. https://doi.org/10.1093/biomet/75.4.800 (1988).
Hochberg, Y. & Benjamini, Y. More powerful procedures for multiple significance testing. Stat. Med. 9, 811–818 (1990).
Ferguson, C. J. An effect size primer: a guide for clinicians and researchers. Prof. Psychol. Res. Pract. 40, 532–538. https://doi.org/10.1037/a0015808 (2009).
Dolcos, F., Miller, B., Kragel, P., Jha, A. & McCarthy, G. Regional brain differences in the effect of distraction during the delay interval of a working memory task. Brain Res. https://doi.org/10.1016/j.brainres.2007.03.059 (2007).
Jonides, J., Smith, E., Marshuetz, C., Koeppe, R. & Reuter-Lorenz, P. Inhibition in verbal working memory revealed by brain activation. Proc. Natl. Acad. Sci. U. S. A. https://doi.org/10.1073/pnas.95.14.8410 (1998).
McNab, F. & Klingberg, T. Prefrontal cortex and basal ganglia control access to working memory. Nat. Neurosci. https://doi.org/10.1038/nn2024 (2008).
Clapp, W., Rubens, M. & Gazzaley, A. Mechanisms of working memory disruption by external interference. Cereb. cortex https://doi.org/10.1093/cercor/bhp150 (2010).
Wang, C., Rajagovindan, R., Han, S. & Ding, M. Top-down control of visual alpha oscillations: sources of control signals and their mechanisms of action. Front. Hum. Neurosci. https://doi.org/10.3389/fnhum.2016.00015 (2016).
Ligeza, T. S., Tymorek, A. D. & Wyczesany, M. Top-down and bottom-up competition in visual stimuli processing. Acta Neurobiol. Exp. 77, 305–316 (2017).
Vogel, E. K. & Luck, S. J. The visual N1 component as an index of a discrimination process. Psychophysiology 37, 190–203 (2000).
Hart, S. J., Lucena, N., Cleary, K. M., Belger, A. & Donkers, F. C. Modulation of early and late event-related potentials by emotion. Front. Integr. Neurosci. 6, 102. https://doi.org/10.3389/fnint.2012.00102 (2012).
Luck, S. J. An Introduction to the Event-Related Potential Technique (MIT, Cambridge, 2014).
Palva, S., Kulashekhar, S., Hamalainen, M. & Palva, J. M. Localization of cortical phase and amplitude dynamics during visual working memory encoding and retention. J. Neurosci. Off. J.Soc. Neurosci. 31, 5013–5025. https://doi.org/10.1523/jneurosci.5592-10.2011 (2011).
Folk, C. L. & Remington, R. Top-down modulation of preattentive processing: testing the recovery account of contingent capture. Vis. Cognit. 14, 445–465. https://doi.org/10.1080/13506280500193545 (2006).
Folk, C. L., Remington, R. W. & Johnston, J. C. Involuntary covert orienting is contingent on attentional control settings. J. Exp. Psychol. Hum. Percept Perform. 18, 1030–1044 (1992).
Harris, A. M., Dux, P. E., Jones, C. N. & Mattingley, J. B. Distinct roles of theta and alpha oscillations in the involuntary capture of goal-directed attention. NeuroImage 152, 171–183. https://doi.org/10.1016/j.neuroimage.2017.03.008 (2017).
Benedek, M., Schickel, R. J., Jauk, E., Fink, A. & Neubauer, A. C. Alpha power increases in right parietal cortex reflects focused internal attention. Neuropsychologia 56, 393–400. https://doi.org/10.1016/j.neuropsychologia.2014.02.010 (2014).
Cooper, N. R., Croft, R. J., Dominey, S. J., Burgess, A. P. & Gruzelier, J. H. Paradox lost? Exploring the role of alpha oscillations during externally vs. internally directed attention and the implications for idling and inhibition hypotheses. Int. J. Psychophysiol. 47, 65–74 (2003).
Hanslmayr, S., Gross, J., Klimesch, W. & Shapiro, K. L. The role of alpha oscillations in temporal attention. Brain. Res. Rev. 67, 331–343. https://doi.org/10.1016/j.brainresrev.2011.04.002 (2011).
Sauseng, P., Griesmayr, B., Freunberger, R. & Klimesch, W. Control mechanisms in working memory: a possible function of EEG theta oscillations. Neurosci. Biobehav. Rev. 34, 1015–1022. https://doi.org/10.1016/j.neubiorev.2009.12.006 (2010).
D’Esposito, M. & Postle, B. R. The cognitive neuroscience of working memory. Annu. Rev. Psychol. 66, 115–142. https://doi.org/10.1146/annurev-psych-010814-015031 (2015).
Kühberger, A., Fritz, A. & Scherndl, T. Publication bias in psychology: a diagnosis based on the correlation between effect size and sample size. PLoS ONE 9, e105825. https://doi.org/10.1371/journal.pone.0105825 (2014).
Acknowledgements
The study was supported by the ÚNKP—New National Excellence Program of the Ministry of Human Capacities, by the Bolyai Research Fellowship Program of the Hungarian Academy of Sciences, by the Predoctoral Fellowship Program of Semmelweis University (EFOP-3.6.3-VEKOP-16-2017-00009), and by the Higher Education Institutional Excellence Program of the Ministry of Human Capacities in Hungary, within the framework of the Neurology thematic program of Semmelweis University.
Author information
Authors and Affiliations
Contributions
G.Cs. designed the study, wrote the protocol and contributed to the writing of all sections. Zs.F. managed the literature searches, undertook the statistical analysis, prepared the figures, and wrote the first draft of the manuscript. Cs.M. participated in the execution of measurements and contributed to the writing of the results section. L.T. participated in designing the study and contributed to the writing of the results section. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fodor, Z., Marosi, C., Tombor, L. et al. Salient distractors open the door of perception: alpha desynchronization marks sensory gating in a working memory task. Sci Rep 10, 19179 (2020). https://doi.org/10.1038/s41598-020-76190-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-76190-3
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
