
Abstract
Drought stress severely impairs plant growth and production. Lipoxygenase (LOX), a master regulator for lipid peroxidation, is critical for direct or indirect response to abiotic stresses. Here, we found that drought stress induced the transcription of CmLOX10 in leaves of oriental melon seedlings. Reverse genetic approaches and physiological analyses revealed that silencing CmLOX10 increased drought susceptibility and stomatal aperture in oriental melon seedlings, and that ectopic overexpression of CmLOX10 in Arabidopsis enhanced drought tolerance and decreased the stomatal aperture. Moreover, the transcription of jasmonic acid (JA)-related genes and JA accumulation were significantly induced in CmLOX10-overexpressed Arabidopsis, which were reversely suppressed in CmLOX10-silenced seedlings during the stage of drought stress. Foliar application of JA further verified that JA enhanced drought tolerance and induced stomatal closure in leaves of melon seedlings. In addition, the feedback regulation of CmLOX10 was induced by JA signaling, and the expression level of CmMYC2 was increased by JA and drought treatment. Yeast one-hybrid analysis showed that CmMYC2 directly bound to the promoter of CmLOX10. In summary, we identified the important roles of CmLOX10 in the regulation of drought tolerance in oriental melon seedlings through JA- mediated stomatal closure and JA signaling-mediated feedback through CmMYC2.
Similar content being viewed by others
Introduction
Drought stress is a major environmental stress that severely impairs plant growth and development, even leading to a serious loss in crops yield1. Extensive research has shown that the defense mechanism in drought stress is related to the expression of stress-related genes, the accumulation of metabolites, the synthesis of osmoprotectants and antioxidants, the maintenance of root growth and water uptake, and the modification of transpiration water loss2. In addition, a large number of studies indicated that phytohormones are important in the drought stress response.
Abscisic acid (ABA), a sesquiterpene derived from C40 oxygenated carotenoids through several synthesis steps3, plays critical roles in drought tolerance in plants4. ABA is involved in physiological processes, such as stomatal closure, osmolyte accumulation, and the synthesis of stress-related proteins 5. In recent years, numerous studies suggested JA plays an important role in response to abiotic stresses. Exogenous application of MeJA promoted plant resistance to abiotic stress, namely salt stress 6, chilling 7, drought stress 8 and so on. Drought stress induced the accumulation of JA 9 in plants, then JA enhanced drought stress mainly through regulating stomatal closure and transpirational water loss 10,11,12,13. Treatment with 12-OPDA, a kind of JA precursor, promoted stomatal closure and OPDA levels in leaf of A. thaliana, thereby reducing stomatal aperture and exhibiting higher tolerance on drought stress 14.
Lipoxygenases (LOXs), an essential enzyme for JA synthesis, can catalyze the oxygenization of polyunsaturated fatty acids containing a cis, cis-1, 4- pentadiene 15, and are classified into 9-LOXs and 13-LOXs in plants according to the specific oxidation sites in linoleic acid 16. LOXs play key roles in all kinds of plant physiology process including growth and development 17,18, senescence 19, and especially responses to biotic and abiotic stresses 20. Pepper CaLOX1 and persimmon DkLOX3, belonging to 9-LOX genes, responded to drought and high salinity stress by modulating the expression of ABA- and stress-responsive genes, lipid peroxidation and ROS accumulation in Arabidopsis 21,22. Numerous studies suggested that 13-LOX genes were mainly involved in the response of plants to biotic and abiotic stresses by regulating JA synthesis. Overexpression of TomloxD in transgenic tomatoes resulted in a significant increase of lipoxygenase activity and endogenous JA content, and boosted the tolerant of tomatoes plants to Cladosporium fulvum and high-temperature stress 23. Arabidopsis LOX2 was found to contribute to the majority of JA synthesis upon wounding and osmotic stress. Arabidopsis lox3 mutant displayed salt sensitivity in different developmental stages, and this phenotype could be rescued by MeJA 6. Little is known about the physiological and molecular mechanisms of LOXs response to abiotic stresses in oriental melon.
Previous studies have shown that JA biosynthesis is also regulated by JA signaling mediated-feedback. JA biosynthetic genes, LOX2 and AOS, were induced by MeJA in Arabidopsis 24. Several evidences have showed that AtMYC2 functioned in JA accumulation induced by through directly binding to the promoters of JA biosynthesis and catabolism genes 25. The expression of TomLoxD, OPR3, AOC, and AOS decreased in the MYC2-silenced tomato plants 26. In addition, MYC2 was found to bind to LOX2/3/4, AOS, and OPR327,28 according to the ChIP-qPCR and ChIP-seq analyses.
However, the mechanism of CmLOXs genes in oriental melon, responding to drought stress, except for CmLOX13 enhancing drought tolerance via regulating ABA accumulation and stomatal closure in Arabidopsis, has been unclear. In our previous study, a total of 18 CmLOXs genes were identified from the oriental melon 'YMR' genome 29 In this study, we found many LOXs genes are dramatically upregulated under drought stress. Drought stress induced the transcription of CmLOX10 in leaves of oriental melon seedlings. Reverse genetic approaches and physiological analyses revealed that CmLOX10 positively affected the response of oriental melon on drought stress and also regulates a JA-mediated pathway in oriental melon seedlings.
Results
Expression pattern of CmLOXs in response to drought stress
Our previous study identified 18 CmLOXs, named CmLOX01-18, and five of them were differently induced by abiotic and signal molecules stresses, such as wounding, high temperature, MeJA, ABA and H2O2 29,30. In this study, we were interested in characterizing the role of CmLOXs in response to drought stress. qRT–PCR analysis has shown that the transcription of CmLOX10, CmLOX11, CmLOX12, CmLOX13, and CmLOX18 in oriental melon leaves were strongly induced by 8% PEG6000 treatment, while the expressions of CmLOX02, CmLOX05 and CmLOX06 were almost unchanged, and the other CmLOXs were slightly up-regulated by 8% PEG6000 treatment (Fig. 1). In this study, we determined to further explore the function of the CmLOX10 in response to drought stress.

Expression of eighteen CmLOXs genes at various time points under 8% PEG6000 treatment. The expression of 18S rRNA was used as internal control. The test was repeated three times, and one biological repeat including four plants.
Silenced of CmLOX10 reduced drought tolerance in oriental melon
To investigate the loss-of-function phenotype of CmLOX10 in drought response, we utilized a tobacco rattlev virus (TRV)-induced gene silencing (VIGS) technique to knock down the CmLOX10 gene in oriental melon plants. During four weeks after VIGS, qRT-PCR analysis results verified that CmLOX10 expression level was significantly decreased in the pTRV-CmLOX10 seedlings (Fig. 2A). TRV-10 seedlings displayed no obvious differences in the growth performance and physiology of TRV-0 seedlings under normal growth conditions (Fig. 2B-F). However, after 10 days of drought stress by withholding water, CmLOX10-silenced (TRV-10) seedlings exhibited wilting to a greater extent than that of empty vector control (TRV-0) (Fig. 2B). Previous studies have shown that malondialdehyde (MDA) production can be induced by drought stress, and there is a negative correlation between MDA content and drought resistance 31. Electrolyte leakage (EL), an important indicator of plant cell permeability, reflects the degree of membrane damage. Besides, under stresses, plants inevitably produced reactive oxygen (ROS), whose accumulation in cells caused oxidative damage to membranes (lipid peroxidation), proteins, RNA and DNA molecules 32,33. In this study, drought stress led to drastic accumulation of EL and MDA contents in TRV-10 seedlings, which was higher than that in TRV-0 seedlings (Fig. 2C-D). The results of DAB staining showed few brown spots both on the TRV-0 and TRV-10 leaves under normal condition, while the brown spots on TRV-10 leaves were significantly more than those on the TRV-0 seedlings after drought treatment (Fig. 2E). The H2O2 contents were consistent with the staining results (Fig. 2F). Taken together, the CmLOX10 gene may play a positive role in regulating drought tolerance in oriental melon.

Silencing CmLOX10 in oriental melon decreased drought tolerance. (A) qRT-PCR analysis of CmLOX10 gene expression in CmLOX10-silenced and control oriental melon seedlings. The expression of 18sRNA was used as an internal control. (B) Representative phenotypes of control (TRV-0) and CmLOX10-silenced (TRV-10) plants at the four-leaf stage after 10 d of drought treatment. Electrolyte leakage (C) and MDA contents (D), representative photographs showing DAB staining (E) and H2O2 contents (F) in TRV-0 and TRV-10 plants under control and drought conditions. Error bars show the SD for n = 3. Asterisks indicate significant differences between the TRV-0 and TRV-10 constructs (Student’s t test, *P < 0.05).
Ectopic expression of CmLOX10 enhanced Arabidopsis drought tolerance
To investigate the gain-of-function phenotype of the CmLOX10 in drought response, three CmLOX10-OX transgenic Arabidopsis lines, OX-7, OX-17 and OX-26 were generated (Fig. 3A). There were no differences between CmLOX10-OX plants and wild-type plants under normal growth conditions (Fig. 3B–G). 24-day-old of WT and transgenic plants were withheld watering for 10 d, obvious differences were found in growth and physiology between WT and CmLOX10-OX plants. Comparing to transgenic plants, more WT plants exhibited wilting symptoms (Fig. 3B). After re-watering for 1 day, survival rates of three transgenic lines were above 88%, being 20% higher than that of the wild type (Fig. 3C). Furthermore, physiological results showed that EL, MDA and the H2O2 contents in transgenic plants were significantly lower than that in WT plants without watering for 10 days (Fig. 3D–G), which supported the higher survival rates of transgenic plants under drought stress. These results confirmed that ectopic expression of CmLOX10 enhanced Arabidopsis drought tolerance.

Ectopic expression of CmLOX10 in Arabidopsis enhanced drought tolerance. (A) qRT-PCR analysis of CmLOX10 gene expression in CmLOX10-OX and WT Arabidopsis plants. The expression of Actin7 was used as an internal control. (B) Representative phenotypes of WT and transgenic plants under normal and drought conditions. Then watering for one day to allow the plants to recover. (C) Survival rates were calculated for the seedlings after 1d of re-watering following drought treatment. Electrolyte leakage (D) and MDA level (E), representative photographs showing DAB staining (F) and H2O2 level (G) in WT and transgenic plants under control and drought conditions. WT, wild type; OX-7, OX-17 and OX-26, individual overexpression line. Error bars show the SD for n = 3. Different letters indicate significant differences at P < 0.05 (Duncan’s multiple range test) of levels for the same index in WT, OX-7, OX-17 and OX-26 plants.
CmLOX10 positively regulates stomatal aperture
Root growth and water evaporation are the two major factors that are involved in the drought tolerance of plant. The root length had no differences between CmLOX10-OX and WT plants (Supplementary Fig. S1A), or between TRV-0 and TRV -10 seedlings (Supplementary Fig. S1B). However, Water loss rate, an important indicator of drought stress was higher in TRV-10 leaves than that in TRV-0 leaves (Fig. 4A), and the leaves fresh weight losses of CmLOX10-OX transgenic plants were lower than those of WT (Fig. 4B). Water is lost from plant leaves through stomata, reminding us to measure the stomatal size. Larger stomatal aperture was observed in leaves of TRV-10 seedlings, comparing to TRV-0 seedlings, under normal and drought conditions (Fig. 4C, D). Meanwhile, the stomatal aperture in leaves of CmLOX10-OX seedlings were smaller than that of WT plants under normal and drought conditions (Fig. 4C, E). Therefore, these results implied that CmLOX10 could enhance the resistance of plants on drought stress, which may be related to reducing transpirational water loss through decreasing stomatal aperture.

CmLOX10 contributes to reducing stomatal aperture. (A) Water loss from the leaves of TRV-0 and TRV-10 plants at various times (one-hour interval) after leaf detachment. (B) Water loss from the leaves of WT and CmLOX10-OX plants at various times (one-hour interval) after leaf detachment. (C) Stomatal phenotype of CmLOX10-silenced and CmLOX10-overexpressed plants under different concentrations of PEG (0 and 15%), TRV-0 and WT plants as the control. (D) Stomatal size of TRV-0 and TRV-10 plants under different concentrations of PEG (0 and 15%). (E) Stomatal size of WT and CmLOX10-OX plants under different concentrations of PEG (0 and 15%). Error bars show the SD for n = 3. Asterisks indicate significant differences between the TRV-0 and TRV-10 constructs (Student’s t test, *P < 0.05). Different letters indicate significant differences at P < 0.05 (Duncan’s multiple range test) of levels for the same index in WT, OX-7, OX-17 and OX-26 plants.
CmLOX10 enhanced drought tolerance by regulating endogenous JA synthesis
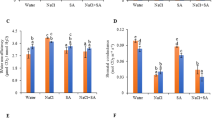
To explore the role of CmLOX10 in response to drought stress, four samples, including CmLOX10-OX and WT Arabidopsis plants under normal and drought conditions with three biological replicates, were used for RNA-seq analysis. Gene ontology (GO) analysis discovered that the DEGs (Fig. 5A) were significantly enriched in biological process including “response to hormone,” “response to abiotic stimulus,” and “jasmonic acid biosynthetic process” between CmLOX10-OX and wild-type plants under drought condition (Fig. 5B). Moreover, the pathways analysis displayed many genes affected by CmLOX10 were involved in diverse pathways. Only hormone metabolism pathway was consistently greater by at least fourfold normalized frequency values in all four comparisons (Table 1). As we all know, ABA and JA (a metabolite of LOXs) are two important phytohormones involved in regulating stomatal closure in response to drought stress in plants 34. Interestingly, the expression of the key genes, NCED3 and NCED5, involved in the synthesis of ABA, had no significant difference in transgenic plants from that in WT leaves, while JA biosynthetic-related genes, AOC3 and OPR3, were upregulated in the CmLOX10-OX lines comparing to WT during drought stress according to the transcriptional data (Fig. 5C, Supplementary Table S2), which prompts us to determine the endogenous ABA and JA levels in transgenic Arabidopsis and oriental melon under normal and drought conditions. As shown in Fig. 5D, there was no significant difference of ABA content between CmLOX10-OX or TRV-0 plants with control plants. JA contents in CmLOX10-OX plants were significantly higher than those of control plants, whereas, JA contents in TRV-10 seedlings were obviously lower than those of control seedlings after drought treatment, respectively (Fig. 5E). These results indicate that CmLOX10 may improve drought tolerance by increasing the synthesis of endogenous JA.

CmLOX10 increased JA synthesis under drought conditions. (A) Venn diagrams of differentially expressed genes by RNA-seq analysis. (B) GO analysis of Up-regulation genes in CmLOX10-OX plants comparing to WT in biological process under drought condition. (C) Expression of key genes in JA and ABA synthesis in WT and CmLOX10-OX lines under normal and drought conditions analyzed by qRT-PCR. (D) ABA contents in Arabidopsis (WT and CmLOX10-OX lines) and oriental melon (TRV-0 and TRV-10) under normal and drought conditions. (E) JA contents in Arabidopsis (WT and CmLOX10-OX lines) and oriental melon (TRV-0 and TRV-10) under normal and drought conditions. AOC, allene oxide cyclase; OPR, 12-OPDA reductase; NCED, 9-cis-epoxycarotenoid dioxygenase. Error bars show the SD for n = 3. Different letters indicate significant differences at P < 0.05 (Duncan’s multiple range test) of levels for the same index in WT, OX-7, OX-17 and OX-26 plants.
In order to explore whether CmLOX10 affects ABA signaling, we first used qRT–PCR analysis to examine the expression pattern of the CmLOX10 gene after ABA treatment. The results showed that the expression of CmLOX10 was not induced by exogenous ABA treatment (Supplementary Fig. S2A). Then, CmLOX10-OX seeds were germinated on 1/2 Murashige and Skoog (MS) medium with different concentrations of ABA. The germination rate, green cotyledon and root length of CmLOX10-OX plants had no significant difference compared with WT plants (Supplementary Fig. S2B, C). In addition, stomatal aperture was measured after ABA treatment, which plays an important role in stomatal closure. However, there was no significant difference in the relative stomatal closure between CmLOX10-OX plants and WT plants(Supplementary Fig. S2D), suggesting that the low transpiration rate of CmLOX10-OX plants was not caused by ABA- mediated stomatal closure.
To investigate the relationship between drought stress and JA level, exogenous JA was applied to oriental melon seedlings before drought treatment. Under normal condition, TRV-0 and TRV-10 seedlings grew very well, and the EL, MDA and H2O2 levels of TRV-10 were similar to those of TRV-0 seedlings. Exposing to drought condition, both TRV-0 and TRV-10 seedlings leaves turned yellow and wilted, and the EL, MDA and H2O2 levels were increased both in leaves of TRV-0 and TRV-10, whereas these physiological indicators were significantly higher in TRV-10 seedlings than in TRV-0 seedlings (Fig. 6A, B). Exogenous JA treatment could alleviate the damage of drought treatment to seedlings and repress the accumulations of EL, MDA and H2O2 in oriental melon seedlings under drought conditions (Fig. 6B). These results indicate that exogenous JA could improve plant drought resistance. To explore whether the larger stomatal aperture in TRV-10 seedlings is caused by JA, oriental melon seedlings were treated with 10 μM JA. The stomatal aperture of both TRV-10 plants and TRV-0 leaves became smaller after JA treatment, but there was a lower relative stomatal closure in TRV-10 leaves comparing to that of TRV-0 leaves (Fig. 6C), suggesting that the part of JA-mediated stomatal closure requires the participation of CmLOX10. Exogenous JA treatment induced the expression of marker genes (SLAC1 and SLAH3) in regulating stomatal closure in TRV-0 and TRV-10 seedlings, thereby reducing stomatal aperture (Fig. 6D).

Exogenous addition of JA can alleviate the damage of drought to oriental melon. The “four leaves stage” oriental melon seedlings were treated with 50 μmol L−1 JA, 15%PEG6000, 50 μmol L−1 JA + 15%PEG6000 for three days, then representative phenotypes (A), and Electrolyte leakage, MDA content and H2O2 level (B) were detected. (C) Stomatal aperture of TRV-0 and TRV-10 plants under different concentrations of JA (0 and 10 μM). (D) Expression of marker genes in regulating stomatal closure in TRV-0 and TRV-10 plants under different concentrations of JA (0 and 50 μM). Error bars indicate SD for three measurements. Asterisks indicate significant differences between the control (TRV-0) and CmLOX10-silenced (TRV-10) constructs (Student’s t test, *P < 0.05).
CmLOX10 expression was feedback regulated by JA signaling
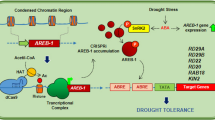
Considering that JA biosynthetic pathway was mainly regulated by the transcriptional regulation, especially the regulation of CmLOX10, we isolated 1346 bp promoter of CmLOX10 from ‘YMR’ and analyzed the putative cis-regulatory elements using the PlantCARE and PLACE databases. Interestingly, within the CmLOX10 promoter region, we found a MeJA-responsive element (CGTCA motif), and qRT-PCR analysis showed that CmLOX10 was induced by JA treatment (Fig. 7A). These results indicate that JA signal transduction pathway might play an important role in CmLOX10 transcriptional regulation. Therefore, we selected CmMYC2 (56.09% homology to AtMYC2), a core transcription factor in the JA signal transduction pathway, as a candidate gene to investigate the regulation of CmLOX10 transcription. In this study, CmMYC2 was induced by JA (Fig. 7B) and PEG6000 (Fig. 7C). In addition, many cis-elements bindings to MYC transcription factor were identified in the 1346 bp promoter region of CmLOX10 (Fig. 7D). Therefore, we focused on analyzing the potential regulation of CmLOX10 transcription by CmMYC2. The specific binding of CmMYC2 to CmLOX10 promoter was verified by Y1H. A 1346 bp promoter sequence upstream of ATG was cloned and constructed into the pAbAi vector. The AbA concentration screening results showed that yeast strain growth was inhibited when SD/-Ura medium supplemented with 200 ng ml−1 AbA (Fig. 7E). Subsequently, P53-pro + AD-P53 and CmLOX10-pro + AD-CmMYC2 were transformed into yeast strain and plated on SD/-Leu medium, finding that the yeast strains were still able to grow; however, yeast transformed CmLOX10-pro + AD-Empty did not grow on the selection medium (Fig. 7E). These findings demonstrated that the transcription of CmLOX10 may be directly regulated by CmMYC2.

CmLOX10 is regulated by a core element of JA signaling CmMYC2 in oriental melon. (A) Expression of the CmLOX10 genes under 50 μM JA treatment. (B) Expression of the CmMYC2 genes under 50 μM JA treatment. (C) Expression of the CmMYC2 genes under 8% PEG6000 treatment. (D) The distribution of major cis-acting elements existed in the CmLOX10 promoter. (E) YIH was used to analyze the CmMYC2 binding CmLOX10 promoter. Error bars indicate SD for three measurements. AbA: Aureobasidin A, AbA200: the AbA concentration is 200 ng mL−1.
Discussion
In plants, LOXs have positive roles in abiotic stress responses, especially osmotic stress, salt stress, mechanical damage and drought stress 22,35 through a variety of mechanisms. However, the function of LOXs remains largely unknown in oriental melon. In this paper, we found that CmLOX10 gene, a 13-LOX gene induced by abiotic and hormonal stresses in oriental melon 30, was quickly and strongly induced by drought stress (Fig. 1). CmLOX10-silenced oriental melon had higher levels of EL, MDA and H2O2 levels were higher than those of control seedlings after 10 days drought treatment (Fig. 2), which was consistent with the drought-sensitive phenotype of CmLOX10-silenced seedlings. In contrast, CmLOX10-OX plants had lower content of EL, MDA and H2O2, supporting the higher survival rates of CmLOX10-OX plants than that of WT plants (Fig. 3). Plants respond to drought stress from the cellular to the whole-plant level. Water loss is crucial for plant tolerance to drought stress 36. The water loss rate was significantly higher or lower in TRV-10 and CmLOX10-OX plants than control, respectively, indicating that the CmLOX10 plays a positive role in response to drought stress.
RNA-seq analysis was used to examine the possible functions of the CmLOX10 at the transcription regulation level. Pathway analysis showed that only the hormone metabolism pathway was consistently greater by at least fourfold normalized frequency values in all four comparisons (Table 1). Previous studies indicated that ABA enhanced plant drought tolerance through regulating stomatal movements 5. In this study, ABA content and transcript of ABA synthetic genes had no obvious differences between CmLOX10-OX and WT plants (Fig. 5C, D). These findings suggest that CmLOX10 enhanced drought stress resistance, not through the ABA synthesis pathway. Similar results were also found in pepper, in which the expressions of NCED3 and NCED5, ABA biosynthesis genes, showed no significant difference between CaLOX1-OX and WT plants under 50 μM ABA treatment 22. GO assay showed that “jasmonic acid biosynthetic process” was enriched in the CmLOX10-OX line (Fig. 5B). In rice, enhanced α-LeA metabolism improved the ability of drought resistance in drought-tolerant landraces/genotypes 37. JA, the metabolite of α-LeA, is a critical signaling molecule in plant defense against biotic and abiotic stresses 38. JA accumulation level indicated that Arabidopsis LOX2, rice OsHI-LOX and OsLOX1, and tobacco NaLOX3 could contribute to JA biosynthesis under wounds and/or herbivores stresses 39,40. The connection between JA and drought tolerance has been widely reported in many higher plants 41. For example, the content of endogenous JA in plants generally increased under water stress 42,43, and exogenous application of JA (or MeJA) at certain concentrations improved drought tolerance 12,44. When plants perceive heat stress, activation of AtOPR3 expression leads to increased JA biosynthesis and accumulation. Subsequently, JA-mediated signaling pathway activated a cascade, resulting in the increase of DREB2A expression and enhancing heat tolerance in plants 45. Normal wound-induced expression of the JA production genes ZmLOX8 and ZmOPR7/8 require LOX10-mediated signaling, whereas normal levels of JA biosynthesis are responses to mechanical damage 46. In this study, we discovered that the key genes of JA biosynthesis (AOC3 and OPR3, Fig. 5C, Supplementary Table S2) were upregulated in CmLOX10-OX lines under drought stress. Meanwhile, increased JA contents in CmLOX10-OX lines were observed compared to WT Arabidopsis, decreased JA content in TRV-10 seedlings were also found, compared to TRV-0 (Fig. 5E). These results suggest that CmLOX10 could play a putative role in drought stress responses by regulating JA synthesis.
JA could be involved in decreasing transpiration loss by regulating stomatal closure 10,47. MeJA-induced stomatal closure is an important physiological response process in plants, which has been reported in various plants such as Arabidopsis 48, barley 49, comfrey50 and pink blue tobacco 51. JA treatment could boost stomatal closure in Atlox2-1 by inducing higher rates of water loss, being consistent with AtLOX2 RNAi plants, compared with the WT, and further investigation indicated that the stomata aperture of Atlox2-1 was wider than that of WT plants under normal growth condition 52. In this study, water loss assays on detached leaves showed that there were higher rates of water loss in TRV-10 seedlings with larger stomata aperture, comparing to TRV-0 seedlings (Fig. 4A, C and D). Comparing to WT plants, CmLOX10-OX plants had lower water loss rates and smaller stomata aperture (Fig. 4B, C and E). CaLOX1-OX plants displayed smaller stomatal aperture than the WT plants under normal and ABA treatment conditions. However, the normalized stomatal aperture of CaLOX1-OX plants was not significantly different from the average stomatal aperture of untreated plants, indicating that the low transpiration rate of CaLOX1-OX plants was not caused by enhanced ABA-induced stomatal closure, which implied that CaLOX1 or CaLOX1-derived oxylipins might play a role in ABA-independent stomatal closure 22. AtLOX1 and AtLOX6 were expressed in guard cells, and AtLOX1 was involved in the regulation of ABA-independent oxylipin-induced stomatal closure at the level of the anion channel SLAC1 to improve defense immunity 53. Our study showed that TRV-10 seedlings accumulated less JA under drought stress with larger stomatal aperture (Fig. 4D, 5E), and displayed smaller stomatal aperture and higher expression of SLAC1 and SLAH3 genes in TRV-0 and TRV-10 plants under adding 10 μM JA treatment (Fig. 6D), while CmLOX10 did not change the sensitivity of Arabidopsis to ABA (Supplementary Fig. S2). All these results demonstrate that CmLOX10 positively regulated drought tolerance through jasmonic acid –dependent pathway in oriental melon.
Interestingly, some studies have shown that JA biosynthesis is also regulated feedback by JA signaling. JA biosynthetic genes LOX2 and AOS were induced by MeJA in Arabidopsis 24. Wounding-induced JA levels were significantly decreased in Arabidopsis coi1-1 mutants 54,55 or COI1-silenced Nicotiana attenuate56. Our research found that CmLOX10 was strongly induced by MeJA (Fig. 7A). Several evidences have pointed out that Arabidopsis thaliana MYC2 functions as the central role in regulating wounding induced JA accumulation by directly binding to the specific motifs (G-box and its variants) in the promoters of genes involved in JA biosynthesis and catabolism 25. The expression of TomLoxD, OPR3, AOC, and AOS decreased in the MYC2-silenced tomato plants26. ChIP-qPCR results indicated that MYC2 directly bond to the promoters of LOX2/3/4, and MeJA treatment enhanced the expression of these genes 27,28,57. In this study, we found that, the expression of CmMYC2 in the oriental melon was strongly induced by MeJA and PEG6000 treatments (Fig. 7B, C). Moreover, many MYC-specific binding sites were found in the promotor of CmLOX10 (Fig. 7D), and Y1H results showed that CmMYC2 directly targeted CmLOX10 promoter (Fig. 7E).
In conclusion, this study provided some evidences that CmLOX10 played an essential role in response to drought stress. CmLOX10 enhanced drought tolerance by promoting JA accumulation and stomatal closure. Although our results suggested that CmMYC2 could directly bind to CmLOX10 promoter, and was involved in regulating drought-induced JA biosynthesis; nevertheless, it was absent of direct evidences for the function of CmMYC2, which needs further to be verified.
Materials and methods
Plant materials and constructs
Arabidopsis thaliana ecotype “Columbia” (WT) were grown in a culture chamber (MLR-350H, SANYO, Japan) at 24 °C, under a 16 h light/8 h dark cycle. The oriental melon (Cucumis melo var. makuwa Makino) inbred “YMR” using for VIGS were raised in culture chamber (MLR-350H, SANYO, Japan) at 25 °C/18 °C, under a 16 h light/8 h dark cycle.
To specifically silence CmLOX10, a 300 bp fragment of the CmLOX10 conserved coding region was inserted into pTRV2 (TRV-0) to generate pTRV2-CmLOX10 (TRV-10) according to Liao’s described 58. CmLOX10 gene contains a 2709 bp open reading frame encoding 902 amino acid 15 and was inserted into the vector 35S:pB7FWG2. CmLOX10-OX plants were generated by using Xing's method 59.
Drought stress treatment
To determine the expression patterns of CmLOXs under drought condition, “Four-leaf stage” oriental melon seedlings treated with 8% PEG6000 were sampled at different times and frozen immediately in liquid nitrogen for RNA preparation.
For drought stress treatment of Arabidopsis plants refers to previous study 59. For drought stress treatment of oriental melon seedlings, “four-leaf stage” TRV-10 and TRV-0 seedlings were treated by withholding water for 10 days, and the physiological data were measured. For exogenous JA treatment, JA (50 μM) was used for three days before drought treatment.
Measurements of electrolyte leakage (EL), malondialdehyde (MDA) and ROS content
EL was measured with minor modification s based on McKersie's method. In brief, fresh rosette leaves (0.1 g) of WT and transgenic Arabidopsis plants and 10 oriental melon fresh leaves disc (d = 1 cm) were incubated in 20 mL ddH2O for one hour at room temperature, determined the conductivities (C1) using a conductivity meter (Model DDS-11A, Rex Instruments Factory, Shanghai, China), subsequently, the leaves were boiled for 10 min, and the conductivities (C2) were recorded after cooling to room temperature. Electrolyte leakage was expressed as C1/C2 × 100%.
In the presence of peroxidase, 3, 3′-diaminobenzidine (DAB) is oxidized by H2O2 to produce a reddish-brown precipitate. The leaves of Arabidopsis and oriental melon treated with drought stress were stained with DAB following the method of Kumar (2014) 60.
MDA and H2O2 contents were determined using MDA Assay Kit (A003-3, Nanjing Jiancheng Bioengineering Institute, China) and H2O2 Assay Kit (A064-1, Nanjing Jiancheng Bioengineering Institute, China), respectively, following the manufacturer’s instructions.
Measurements of seed germination and root length
For germination rate and cotyledons greening after different concentrations of ABA (0, 0.5 and 0.75 μM) treatment were scored on the 7rd day after germination, with three replicated assays according to Xing's description 59.
For Arabidopsis root length experiment, four-day seedlings grown on 1/2 MS medium were transferred to new medium with different concentrations of mannitol (0–500 mM) and ABA (0 and 10 μM) and measured root length after vertical cultured for 7d. For oriental melon root length experiment, “four-leaf stage” plants without watering for 10 days, the roots were observed using root scanner (MICROTEK ScanMaker i800 plus).
Water loss and stomatal aperture assay
For the measurements of water loss, rosette leaves of 3-week-old Arabidopsis (10 plants/ one repeat) and oriental melon (4 plants/one repeat) were detached and weighed immediately (W1), then the plants were left in a plastic tray at 25 °C, 60% relative humidity. The weight (W2) of the leaves was measured at one-hour intervals. The water loss rate was calculated with the following formula: water loss rate (%) = (W1-W2)/W1 × 100%. The experiment repeated three times.
Stomatal aperture measurements were implemented as described previously 61. Epidermal peels of expanded mature leaves were floated in stomatal opening solution exposing to light for 2 h, then transferred the epidermal peels into solutions added different concentrations (0, 15%) of PEG6000, JA (0, 10 μM) and ABA (0, 10 μM) for 2 h before being observed. The average of 90 stomata ratio (width/ length) at least repressed stomatal aperture.
JA and ABA measurement
JA and ABA contents were analyzed using HPLC–MS/MS method according to Xing's description 59.
Transcriptomic analysis
CmLOX10-overexpression line OX-17 and WT plants here were used for transcriptomic analyses. Rosette leaves of WT and OX-17 plants were collected from normal and 8d—drought- stress plants for RNA isolation, and each sample was represented by three replicates. RNA quality and quantified were checked using the NanoDrop and 1% agarose gel electrophoresis. A total of 5 μg of RNA was used to synthesize the subsequent sequencing libraries. After library concentration was tested by Qubit 2.0, the library preparations were then sequenced on an Illumina Hiseq platform. After filtering the raw data, Trimmomatic software was used to remove adapter and low-quality reads to get the clean data. Q20, Q30, and the GC contents of the cleaning data were simultaneously calculated. Then we identified the transcript sequence in the reference Arabidopsis genome to obtain comprehensive transcript information. RPKM (Reads per Kilobase per Million Reads) was used as a standard of gene expression. And DESeqR package (1.18.0) was used for differential expression analysis of drought stress versus control conditions. To control the false discovery rate, Benjamini and Hochberg’s approach were used to adjust the resulting P-values. The functional analysis of differentially expressed genes (absolute value of log FC > 1 and P-Value < 0.05) were used GO and MapMan as the classification source.
Quantitative reverse transcription–PCR (qRT-PCR) analysis
Total RNA was isolated using an ultrapure RNA extraction kit (CWbio. Co. Ltd., Beijing, China) and cDNA were synthesized using PrimeScript RT Master Mix (Perfect Real Time) (Takara, Dalian, China) according to the manufacturer's instructions, respectively. Diluted cDNA of fourfold was using for qRT-PCR analysis according to the SuperReal PreMix Plus (SYBR Green) (Takara, Dalian, China) manufacturer’s protocol. The expression analysis was repeated three times. Four oriental melon plants and ten Arabidopsis plants, respectively, were used at one biological repeat in statistical analysis. The expression of Actin7 and 18sRNA were used as internal reference genes in Arabidopsis and oriental melon, respectively. Primers for the qRT-PCR analysis were listed in Table S1.
Cloned and analysis of CmLOX10 promoter (CmLOX10-pro)
Genomic DNA was extracted from the tender leaves of oriental melon to be used for the cloning of CmLOX10 promoter searched from the melon genome database (https://melonomics.net/). According to the cloned and sequenced fragments, the putative cis-regulatory elements of CmLOX10 promoter were performed using PlantCARE (http:// bioinformatics.psb.ugent.be/webtools/plantcare/html/) and PLACE (http:// www.dna.affrc.go.jp/PLACE/) database.
Yeast one-hybrid (Y1H) assay
Y1H was performed to confirm that the CmMYC2 can directly regulated CmLOX10 according to the instructions of Matchmaker Gold Yeast One-Hybrid Library Screening System (Clontech, USA). The CmLOX10 promoter was subcloned into the pAbAi vector, and the full-length CDSs of CmMYC2 (MELO3C013851) searched from the melon genome database (https://melonomics.net/) was cloned and inserted into the pGADT7 (AD) vectors. The fusion constructs were transformed into Y1HGold strain, then was cultured on SD/-Ura medium supplemented with (0, 100, 150, 200 ng/ml) Aureobasidin A (AbA), to screen for the concentration of AbA inhibition. Finally, the positive control (P53-pro + AD-P53), negative control (CmLOX10-pro + AD-Empty) and the target group (CmLOX10-pro + AD-CmMYC2) were transformed into yeast strain Y1H to and cultured on SD/-Leu medium containing 200 ng/ml AbA.
Statistical analysis
All the experiments were carried out three times in a completely random sampling. We applied one-way ANOVAs with SPSS16.0 statistics program, and presented values as means ± SD. Asterisks above the columns indicate significant differences between the TRV-0 and TRV-10 seedlings (Student’s t test, *P < 0.05). All charts were drawing by Origin 8.0 in this paper.
References
Parmar, R. et al. Transcriptional profiling of contrasting genotypes revealed key candidates and nucleotide variations for drought dissection in Camellia sinensis (L.) O. Kuntze. Sci. Rep. 9, 32. https://doi.org/10.1038/s41598-019-43925-w (2019).
Krasensky, J. & Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 63, 1593–1608. https://doi.org/10.1093/jxb/err460 (2012).
Munoz-Espinoza, V. A., Lopez-Climent, M. F., Casaretto, J. A. & Gomez-Cadenas, A. Water stress responses of tomato mutants impaired in hormone biosynthesis reveal abscisic acid, jasmonic acid and salicylic acid interactions. Front. Plant. Sci. 6, 997. https://doi.org/10.3389/fpls.2015.00997 (2015).
Ashraf, M. Inducing drought tolerance in plants: recent advances. Biotechnol. Adv. 28, 169–183. https://doi.org/10.1016/j.biotechadv.2009.11.005 (2010).
Hoekstra, F. A., Golovina, E. A. & Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 6, 431–438 (2001).
Ding, H. et al. Jasmonate complements the function of Arabidopsis lipoxygenase3 in salinity stress response. Plant Sci. 244, 1–7. https://doi.org/10.1016/j.plantsci.2015.11.009 (2016).
Cao, S., Cai, Y., Yang, Z. & Zheng, Y. MeJA induces chilling tolerance in loquat fruit by regulating proline and γ-aminobutyric acid contents. Food Chem. 133, 1466–1470. https://doi.org/10.1016/j.foodchem.2012.02.035 (2012).
Anjum, S. A. et al. Exogenously applied methyl jasmonate improves the drought tolerance in wheat imposed at early and late developmental stages. Acta Physiol. Plant. https://doi.org/10.1007/s11738-015-2047-9 (2015).
Creelman, R. A. & Mullet, J. E. Jasmonic acid distribution and action in plants: regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. 92, 4114–4119 (1995).
Daszkowska-Golec, A. & Szarejko, I. Open or close the gate - stomata action under the control of phytohormones in drought stress conditions. Front Plant Sci 4, 138. https://doi.org/10.3389/fpls.2013.00138 (2013).
Schaller, A. & Stintzi, A. Enzymes in jasmonate biosynthesis—structure, function, regulation. Phytochemistry 70, 1532–1538. https://doi.org/10.1016/j.phytochem.2009.07.032 (2009).
Mahmood, M., Bidabadi, S. S., Ghobadi, C. & Gray, D. J. Effect of methyl jasmonate treatments on alleviation of polyethylene glycol -mediated water stress in banana (Musa acuminata cv. ‘Berangan’, AAA) shoot tip cultures. Plant Growth Regul. 68, 161–169. https://doi.org/10.1007/s10725-012-9702-6 (2012).
Tanaka, Y. et al. Ethylene inhibits abscisic acid-induced stomatal closure in Arabidopsis. Plant Physiol. 138, 2337–2343. https://doi.org/10.1104/pp.105.063503 (2005).
Savchenko, T. et al. Functional convergence of oxylipin and abscisic acid pathways controls stomatal closure in response to drought. Plant Physiol. 164, 1151–1160. https://doi.org/10.1104/pp.113.234310 (2014).
Cao, S. et al. Heterologous expression and biochemical characterization of two lipoxygenases in oriental melon, Cucumis melo var. makuwa Makino. PLoS ONE 11, e0153801. https://doi.org/10.1371/journal.pone.0153801 (2016).
Feussner, I. & Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 53, 275–297. https://doi.org/10.1146/annurev.arplant.53.100301.135248 (2002).
Yang, X. Y., Jiang, W. J. & Yu, H. J. The expression profiling of the lipoxygenase (LOX) family genes during fruit development, abiotic stress and hormonal treatments in cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 13, 2481–2500. https://doi.org/10.3390/ijms13022481 (2012).
Kolomiets, M. V., Hannapel, D. J., Chen, H., Tymeson, M. & Gladon, R. J. Lipoxygenase is involved in the control of potato tuber development. Plant Cell 13, 613–626 (2001).
Seltmann, M. A. et al. Differential impact of Lipoxygenase 2 and Jasmonates on natural and stress-induced senescence in Arabidopsis. Plant Physiol. 152, 1940–1950. https://doi.org/10.1104/pp.110.153114 (2010).
Nemchenko, A., Kunze, S., Feussner, I. & Kolomiets, M. Duplicate maize 13-lipoxygenase genes are differentially regulated by circadian rhythm, cold stress, wounding, pathogen infection, and hormonal treatments. J. Exp. Bot. 57, 3767–3779. https://doi.org/10.1093/jxb/erl137 (2006).
Hou, Y. et al. The persimmon 9-lipoxygenase Gene DkLOX3 plays positive roles in both promoting senescence and enhancing tolerance to abiotic stress. Front. Plant Sci. 6, 1073. https://doi.org/10.3389/fpls.2015.01073 (2015).
Lim, C. W. et al. The pepper lipoxygenase CaLOX1 plays a role in osmotic, drought and high salinity stress response. Plant Cell Physiol. 56, 930–942. https://doi.org/10.1093/pcp/pcv020 (2015).
Hu, T., Zeng, H., Hu, Z., Qv, X. & Chen, G. Overexpression of the tomato 13-Lipoxygenase gene tomloxd increases generation of endogenous jasmonic acid and resistance to Cladosporium fulvum and high temperature. Plant Mol. Biol. Rep. 31, 1141–1149. https://doi.org/10.1007/s11105-013-0581-4 (2013).
Sasaki, Y. et al. Monitoring of methyl jasmonate-responsive genes in Arabidopsis by cDNA macroarray: self-activation of jasmonic acid biosynthesis and crosstalk with other phytohormone signaling pathways. DNA Res. 8, 153–161 (2001).
Schweizer, F. et al. Arabidopsis basic helix-loop-helix transcription factors MYC2, MYC3, and MYC4 regulate glucosinolate biosynthesis, insect performance, and feeding behavior. Plant Cell 25, 3117–3132. https://doi.org/10.1105/tpc.113.115139 (2013).
Du, M. et al. MYC2 orchestrates a hierarchical transcriptional cascade that regulates jasmonate-mediated plant immunity in tomato. Plant Cell 29, 1883–1906. https://doi.org/10.1105/tpc.16.00953 (2017).
Hou, X., Lee, L. Y., Xia, K., Yan, Y. & Yu, H. DELLAs modulate jasmonate signaling via competitive binding to JAZs. Dev. Cell 19, 884–894. https://doi.org/10.1016/j.devcel.2010.10.024 (2010).
Zhai, Q. et al. Phosphorylation-coupled proteolysis of the transcription factor MYC2 is important for jasmonate-signaled plant immunity. PLoS Genet. 9, e1003422. https://doi.org/10.1371/journal.pgen.1003422 (2013).
Zhang, C. et al. The phylogeny and expression profiles of the lipoxygenase (LOX) family genes in the melon (Cucumis melo L.) genome. Sci. Hortic. 170, 94–102. https://doi.org/10.1016/j.scienta.2014.03.005 (2014).
Liu, J. et al. Effects of abiotic stress and hormones on the expressions of five 13-CmLOXs and enzyme activity in oriental melon (Cucumis melo var. makuwa Makino). J. Integr. Agric. 15, 326–338. https://doi.org/10.1016/s2095-3119(15)61135-2 (2016).
DaCosta, M. & Huang, B. Changes in antioxidant enzyme activities and lipid peroxidation for bentgrass species in response to drought stress. J. Am. Soc. Hortic. Sci. 132, 319–326 (2007).
Apel, K. & Hirt, H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 55, 373–399. https://doi.org/10.1146/annurev.arplant.55.031903.141701 (2004).
Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 7, 405–410 (2002).
Pospíšilová, J. Participation of phytohormones in the stomatal regulation of gas exchange during water stress. Biol. Plant. 46, 491–506 (2003).
Veronico, P. et al. A novel lipoxygenase in pea roots. Its function in wounding and biotic stress. Plant Physiol. 141, 1045–1055. https://doi.org/10.1104/pp.106.081679 (2006).
Yin, M. et al. The Arabidopsis Cys2/His2 zinc finger transcription factor ZAT18 is a positive regulator of plant tolerance to drought stress. J. Exp. Bot. 68, 2991–3005. https://doi.org/10.1093/jxb/erx157 (2017).
Lenka, S. K., Katiyar, A., Chinnusamy, V. & Bansal, K. C. Comparative analysis of drought-responsive transcriptome in Indica rice genotypes with contrasting drought tolerance. Plant Biotechnol. J. 9, 315–327. https://doi.org/10.1111/j.1467-7652.2010.00560.x (2011).
Sasaki-Sekimoto, Y. et al. Coordinated activation of metabolic pathways for antioxidants and defence compounds by jasmonates and their roles in stress tolerance in Arabidopsis. Plant J.: Cell Mol. Biol. 44, 653–668. https://doi.org/10.1111/j.1365-313X.2005.02560.x (2005).
Halitschke, R. & Baldwin, I. T. Antisense LOX expression increases herbivore performance by decreasing defense responses and inhibiting growth-related transcriptional reorganization in Nicotiana attenuata. Plant J.: Cell Mol. Biol. 36, 794–807. https://doi.org/10.1046/j.1365-313x.2003.01921.x (2003).
Zhou, G. et al. Silencing OsHI-LOX makes rice more susceptible to chewing herbivores, but enhances resistance to a phloem feeder. Plant J.: Cell Mol. Biol. 60, 638–648. https://doi.org/10.1111/j.1365-313X.2009.03988.x (2009).
Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 20, 219–229. https://doi.org/10.1016/j.tplants.2015.02.001 (2015).
Zhang, C. & Huang, Z. Effects of endogenous abscisic acid, jasmonic acid, polyamines, and polyamine oxidase activity in tomato seedlings under drought stress. Sci. Hortic. 159, 172–177. https://doi.org/10.1016/j.scienta.2013.05.013 (2013).
Pedranzani, H., Sierra-de-Grado, R., Vigliocco, A., Miersch, O. & Abdala, G. Cold and water stresses produce changes in endogenous jasmonates in two populations of Pinus pinaster Ait. Plant Growth Regul. 52, 111–116. https://doi.org/10.1007/s10725-007-9166-2 (2007).
Alam, M. M., Nahar, K., Hasanuzzaman, M. & Fujita, M. Exogenous jasmonic acid modulates the physiology, antioxidant defense and glyoxalase systems in imparting drought stress tolerance in different Brassica species. Plant Biotechnol. Rep. 8, 279–293. https://doi.org/10.1007/s11816-014-0321-8 (2014).
Tian, X. et al. HEAT SHOCK TRANSCRIPTION FACTOR A1b regulates heat tolerance in wheat and Arabidopsis through OPR3 and jasmonate signaling pathway. Plant Biotechnol. J. https://doi.org/10.1111/pbi.13268 (2019).
Christensen, S. A. et al. The maize lipoxygenase, ZmLOX10, mediates green leaf volatile, jasmonate and herbivore-induced plant volatile production for defense against insect attack. Plant J.: Cell Mol. Biol. 74, 59–73. https://doi.org/10.1111/tpj.12101 (2013).
Munemasa, S. et al. The coronatine-insensitive 1 mutation reveals the hormonal signaling interaction between abscisic acid and methyl jasmonate in Arabidopsis guard cells Specific impairment of ion channel activation and second messenger production. Plant Physiol. 143, 1398–1407. https://doi.org/10.1104/pp.106.091298 (2007).
Suhita, D., Raghavendra, A. S., Kwak, J. M. & Vavasseur, A. Cytoplasmic alkalization precedes reactive oxygen species production during methyl jasmonate-and abscisic acid-induced stomatal closure. Plant Physiol. 134, 1536–1545 (2004).
Tsonev, T., Lazova, G., Stoinova, Z. G. & Popova, L. A possible role for jasmonic acid in adaptation of barley seedlings to salinity stress. J. Plant Growth Regul. 17, 153–159 (1998).
Raghavendra, A. & Reddy, K. B. Action of proline on stomata differs from that of abscisic acid, G-substances, or methyl jasmonate. Plant Physiol. 83, 732–734 (1987).
Suhita, D., Kolla, V. A., Vavasseur, A. & Raghavendra, A. S. Different signaling pathways involved during the suppression of stomatal opening by methyl jasmonate or abscisic acid. Plant Sci. 164, 481–488. https://doi.org/10.1016/s0168-9452(02)00432-6 (2003).
Sun, Y. et al. Lipoxygenase 2 functions in exogenous nitric oxide-induced stomatal closure in Arabidopsis thaliana. Funct. Plant Biol. 42, 1019. https://doi.org/10.1071/fp15151 (2015).
Montillet, J. L. et al. An abscisic acid-independent oxylipin pathway controls stomatal closure and immune defense in Arabidopsis. PLoS Biol. 11, e1001513. https://doi.org/10.1371/journal.pbio.1001513 (2013).
Chung, H. S. et al. Regulation and function of Arabidopsis JASMONATE ZIM-domain genes in response to wounding and herbivory. Plant Physiol 146, 952–964. https://doi.org/10.1104/pp.107.115691 (2008).
Glauser, G. et al. Spatial and temporal dynamics of jasmonate synthesis and accumulation in Arabidopsis in response to wounding. J. Biol. Chem. 283, 16400–16407. https://doi.org/10.1074/jbc.M801760200 (2008).
Paschold, A., Bonaventure, G., Kant, M. R. & Baldwin, I. T. Jasmonate perception regulates jasmonate biosynthesis and JA-Ile metabolism: the case of COI1 in Nicotiana attenuata. Plant Cell Physiol. 49, 1165–1175. https://doi.org/10.1093/pcp/pcn091 (2008).
Niu, Y., Figueroa, P. & Browse, J. Characterization of JAZ-interacting bHLH transcription factors that regulate jasmonate responses in Arabidopsis. J. Exp. Bot. 62, 2143–2154. https://doi.org/10.1093/jxb/erq408 (2011).
Liao, J. J. et al. Overexpression and VIGS system for functional gene validation in oriental melon (Cucumis melo var. makuwa Makino). Plant Cell Tissue Organ Cult. (PCTOC) 137, 275–284. https://doi.org/10.1007/s11240-019-01568-9 (2019).
Xing, Q. et al. The lipoxygenase CmLOX13 from oriental melon enhanced severe drought tolerance via regulating ABA accumulation and stomatal closure in Arabidopsis. Environ. Exp. Bot. 167, 103815. https://doi.org/10.1016/j.envexpbot.2019.103815 (2019).
Kumar, D. et al. Histochemical detection of superoxide and H2O2 accumulation in Brassica juncea seedlings. Bio Prot. 4, e1108 (2014).
Hossain, M. A. et al. Involvement of endogenous abscisic acid in methyl jasmonate-induced stomatal closure in Arabidopsis. Plant Physiol. 156, 430–438. https://doi.org/10.1104/pp.111.172254 (2011).
Acknowledgements
This work was supported by the Agriculture Research System of China (CARS-25).
Author information
Authors and Affiliations
Contributions
LM and LTH: sowing and maintenance of the plants in growth chamber. CSX: acquisition of CmLOX10-OX transgenic plants. LJJ: acquisition of CmLOX10-silenced plants. QHY: direction and coordination of the study. All other experiments and manuscript writing were performed by XQJ.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xing, Q., Liao, J., Cao, S. et al. CmLOX10 positively regulates drought tolerance through jasmonic acid -mediated stomatal closure in oriental melon (Cucumis melo var. makuwa Makino). Sci Rep 10, 17452 (2020). https://doi.org/10.1038/s41598-020-74550-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-74550-7
This article is cited by
-
Genome-wide identification of ZmMYC2 binding sites and target genes in maize
BMC Genomics (2024)
-
CmDOF18 positively regulates salinity tolerance in Chrysanthemum morifolium by activating the oxidoreductase system
BMC Plant Biology (2024)
-
Harnessing Jasmonate, Salicylate, and Microbe Synergy for Abiotic Stress Resilience in Crop Plants
Journal of Plant Growth Regulation (2024)
-
Modulation of the wheat transcriptome by TaZFP13D under well-watered and drought conditions
Plant Molecular Biology (2024)
-
Revisiting the Critical Role of ROS and RNS in Plant Defense
Journal of Plant Growth Regulation (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
