
Abstract
The Nudiviridae are a family of large double-stranded DNA viruses that infects the cells of the gut in invertebrates, including insects and crustaceans. The phylogenetic range of the family has recently been enhanced via the description of viruses infecting penaeid shrimp, crangonid shrimp, homarid lobsters and portunid crabs. Here we extend this by presenting the genome of another nudivirus infecting the amphipod Dikerogammarus haemobaphes. The virus, which infects cells of the host hepatopancreas, has a circular genome of 119,754 bp in length, and encodes a predicted 106 open reading frames. This novel virus encodes all the conserved nudiviral genes (sharing 57 gene homologues with other crustacean-infecting nudiviruses) but appears to lack the p6.9 gene. Phylogenetic analysis revealed that this virus branches before the other crustacean-infecting nudiviruses and shares low levels of gene/protein similarity to the Gammanudivirus genus. Comparison of gene synteny from known crustacean-infecting nudiviruses reveals conservation between Homarus gammarus nudivirus and Penaeus monodon nudivirus; however, three genomic rearrangements in this novel amphipod virus appear to break the gene synteny between this and the ones infecting lobsters and penaeid shrimp. We explore the evolutionary history and systematics of this novel virus, suggesting that it be included in the novel Epsilonnudivirus genus (Nudiviridae).
Similar content being viewed by others

Introduction
The family Nudiviridae comprises a group of non-occluded, double-stranded DNA (dsDNA) viruses infecting arthropods. The family includes two recognised genera, Alphanudivirus and Betanudivirus, with two other genera, Gammanudivirus and Deltanudivirus, recently proposed to contain aquatic-host-infecting viruses 1,2. Alphanudivirus contains two species; Gryllus bimaculatus nudivirus and Oryctes rhinoceros nudivirus, while Betanudivirus contains a single species, Heliothis zea nudivirus 3. In addition, several other insect-infecting nudiviruses have been morphologically and/or genomically characterized but remain to be formally recognised 4. In recent years, closely related viruses have been identified infecting marine crustaceans, using genomics and ultrastructural data. The most complete descriptions include Penaeus monodon nudivirus (PmNV) infecting the farmed penaeid shrimp Penaeus monodon1 and recently, Homarus gammarus nudivirus (HgNV) infecting juveniles of the European lobster, Homarus gammarus2.
Other crustaceans are also infected with putative nudiviruses, often referred to in the published literature as “bacilliform viruses” in lieu of available genomic data5,6,7,8,9,10,11. In addition to descriptions of putative nudiviruses in these decapod crustaceans, observations of nudivirus-like infections of Amphipoda have also been reported; e.g. for Dikerogammarus villosus 12, Dikerogammarus haemobaphes 11, Gammarus roeselii 8, Pontogammarus robustoides 7 and Gammarus varsoviensis 7. In all cases, putative nudivirus infection is observed within cells of the hepatopancreas, causing nuclear hypertrophy but no observable host immune response to infection, in histological Section8,11. Histopathologically, nudiviral infection results in nuclear hypertrophy of the hepatopancreatocytes, caused primarily by a growing viroplasm; this is explored in full for the virus in D. haemobaphes in Bojko et al. 11. Also in D. haemobaphes, infection prevalence of up to 77.7% has been correlated with altered behaviour in infected animals 11. The behavioural change was associated with increased activity, which positively correlated with viral burden, potentially indicating some benefit to viral transmission through increased movement 11. Many of the amphipod hosts in which putative nudiviruses have been reported are non-native or invasive species present outside of their native ranges. Infection with this viral family may therefore have potential for transboundary transmission when their hosts are present in their invasive range 7,10,11.
Genomic data is currently lacking for all the putative nudiviruses infecting amphipods. In this study, we provide full genome characterisation of a nudivirus infecting the amphipod D. haemobaphes collected from outside of its native range. We use these data to provisionally name the virus as Dikerogammarus haemobaphes nudivirus (DhNV) and place the virus within a newly suggested genus Epsilonnudivirus of the family Nudiviridae.
Results
Genome structure of DhNV
The circular genome of DhNV is 119,754 bp and contains 106 hypothetical ORFs, with 56 on the positive strand and 50 on the negative strand (Fig. 1). Fifty-nine of the ORFs met our comparative e-value threshold of < 0.001 and were directly comparable to other members of the Nudiviridae. Up to 17% and 37% of the ORFs aligned most closely with genes from HgNV and PmNV, respectively. Two ORFs, DhNV_008 and DhNV_002, scored above 50% similarity to protein sequences on BLASTp. DhNV_008 was 52.33% similar to pif-2 from HgNV (QBB28614) with per os infectivity as the only identified protein domain. DhNV-002 was 50% similar to a hypothetical protein from PmNV (YP_009051845) where the cytoplasmic, non-cytoplasmic, tmhelix, and transmembrane domains were identified. Among the 47 ORFs that provided no similarity to other protein sequences within the threshold, InterProScan assessment identified 20 ORFs with functional domains. A protein signature match to the inhibitor of apoptosis repeat superfamily in DhNV_059 may indicate the presence of a homolog of the Iap nudivirus gene; however, BLASTp annotations did not yield any similarity results to the Iap gene found in other nudiviruses. The remaining 19 ORFs contained proteins with Zinc finger domains (DhNV_045), Tmhelix (DhNV_044), signal peptide (DhNV_084), p-loop containing nucleoside triphosphate hydrolase (DhNV_047), non-cytoplasmic domain (DhNV_025, 028, and 057), disorder predictions (DhNV_011, 020, 024, 072, 077, and 078), predicted to be cytoplasmic (DhNV_019 and 065), or had ‘coil’ feature(s) (DhNV_022, 026, 042, and 086).

The ‘Dikerogammarus haemobaphes nudivirus’ circular genome with open reading frames (ORF) graphically plotted across the 119,754 bp sequence. Locus tags for each similar virus are denoted in the place of DhNV genes, where possible. Gold regions and text indicate predicted genes on the positive strand and black regions and text indicate those on the negative strand. Below the genome, the green plot displays genome coverage across the genome using the MiSeq and HiSeq trimmed sequence reads. ORFs with significant similarity (e-value < 0.001) to other nudivirus genes are listed as the reference gene number on the corresponding viral genome. The circular plot was developed in Circa (www.omgenomics.com/circa/) and the coverage map from CLC genomics workbench v.12 (Qiagen).
Seven genes involved with DNA processing were identified: DNA polymerase, helicase, two copies of helicase 2, integrase, DNA ligase, and fen-1. Five genes involved with RNA transcription were also identified: p47, lef-4, lef-5, lef-8, and lef-9. Eight genes known to be involved in per os infectivity were found: vp91 (pif-8), pif-1, pif-2, pif-3, odv-e28 (pif-4), odv-e56 (pif-5), ac68 (pif-6), and p74 (pif-0), along with the 11 k gene. Nine genes can be grouped into packaging, assembly, and release: two copies of ac92 (p33), two copies of odv-e66, two copies of vlf-1, 38 k, ac81, and 31 K (vp39). The recently identified p6.9 gene in crustacean-infecting nudivirus genomes, a baculovirus core gene associated with encapsulation of the viral genome, could not be found in DhNV despite similarity searches and checks for hypothetical SRSR repeat regions common to the p6.9 protein. A putative DUTPase, cg30-1, p-loop NTPase, guanosine monophosphate kinase, esterase, and p51 were also identified. Definitively, 20 out of the 21 core baculovirus genes conserved among nudivirus were identified in the DhNV genome (Table 1).
Multiple genes were specific to DhNV; however, some were similar to other protein groups from various taxa. One ORF, DhNV_076, revealed 31.98% to 35.97% similarity to proteins from seven different organisms. The LOC108666550-like protein from HgNV is 32.35% similar (85% coverage) to DhNV_076 (Table 1). Uncharacterized proteins from Hyalella azteca (QBB28676, sim. 35.97% cov. 82%), an amphipod native to North America; Penaeus vannamei (XP_027235023, sim. 34.07% cov. 83%), the Whiteleg shrimp; Crassostrea gigas (XP_011433877, sim. 31.98% cov. 85%), the Pacific Oyster; Armadillidium vulgare (RXG54766, sim. 34.15% cov. 83%), the common pill-bug; and Tigriopus californicus (TRY76277, similarity 33.26% coverage 88%), a North American coastal copepod, all met the e-value threshold, as did the actin-binding IPP-like protein from Brachionus plicatilis (RMZ96256 similarity 33.65% coverage 87%). Only the undescribed PANTHER protein family, PTHR38566, was identified as a domain in DhNV_076.
Gene synteny among the Epsilonnudivirus, Gammanudivirus and Deltanudivirus genera
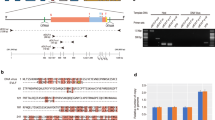
A comparison of gene synteny between DhNV and three other nudiviruses determined that DhNV had a different gene synteny to members of the Gammanudivirus and Deltanudivirus genera (Fig. 2). Comparison between ToNV and DhNV revealed a high level of genomic rearrangement, where the 32 genes that showed genetic similarity (e < 0.001) with those ORFs on the DhNV genome were located across the respective genomes, showing little conserved synteny (Fig. 2a). Comparison between DhNV and PmNV/HgNV revealed higher levels of gene synteny (Fig. 2b,c). A comparison using all three viruses identified 12 major regions of genetic novelty in the DhNV genome (Fig. 2d). This included 47 hypothetical ORFs that were unique to DhNV and showed little genetic/protein relatedness to other nudiviruses within the e-value threshold of < 0.001, one of which (DhNV_070) showed highest similarity to a gene from Pyricularia oryzae, a fungal plant pathogen, and another (DhNV_029) with highest similarity to Sucra jujuba nucleopolyhedrovirus (Table 1).

(a) Gene synteny among the Epsilonnudivirus, Gammandivirus and Deltanuvirus reveals a gene order among decapod-infecting nudiviruses, which is missing from peracarid-infecting nudiviruses. The genomes are annotated with positive strand (gold) and negative strand (silver) coding regions. In Fig. 2a, Dikerogammarus haemobaphes nudivirus gene synteny (white) is compared to Tipula oleracea nudivirus gene synteny (pink). Ribbons connecting the two genomes link up the homologous gene and its location on the viral genome. Scale ticks = 2 kb. The comparative plots were developed in Circa (www.omgenomics.com/circa/). (b) Dikerogammarus haemobaphes nudivirus gene synteny (white) is compared to Penaeus monodon nudivirus gene synteny (blue). Ribbons connecting the two genomes link up the homologous gene and its location on the viral genome. Scale ticks = 2 kb. The comparative plots were developed in Circa (www.omgenomics.com/circa/). (c) Dikerogammarus haemobaphes nudivirus gene synteny (white) is compared to Hommarus gammarus nudivirus gene synteny (green). Ribbons connecting the two genomes link up the homologous gene and its location on the viral genome. Scale ticks = 2 kb. The comparative plots were developed in Circa (www.omgenomics.com/circa/). (d) All four virus are compared together, indicating regions of novelty in the Dikerogammarus haemoabphes nudivirus genome. The white triangles on the DhNV genome highlight the areas of novel sequence information that do not correspond to genes on the other nudivirus genomes. Scale ticks = 10 kb. The comparative plots were developed in Circa (www.omgenomics.com/circa/). (e) The crustacean nuidviruses contain 24 nudivirus core genes (VLF-1, ODV-E66 and Helicase 2 are duplicated) (p6.9 is missing from Dikerogammarus haemobaphes nudivirus) (orange) and 11 other gene homologs conserved across the crustacean-infecting nudiviruses (yellow). These conserved homologues are based on similarity, synteny and functional identity. Comparison with DhNV (peracarid-infecting nudivirus) highlights three main areas of gene reorganization. ‘X’ corresponds to a rearrangement of the DhNV_032, DhNV_34 and pif-1 genes, ‘Y’ corresponds to a rearrangement of the vlf-1 and p74 genes and finally, ‘Z’ corresponds to a rearrangement of six genes (vlf-1, ac68, DhNV_080, ac81 and both helicase 2 homologues). The Gammanudivirus members that infect decapods share the same gene synteny across these conserved motifs.
Using the protein similarity data, we determined that there were 11 crustacean-infecting nudivirus genes (DhNV_006, 024, 032, 034, 050, 062, 067, 080, 103, 104 and 106) that show conservation across the crustacean-infecting nudiviruses (Table 1) (i.e. present in PmNV, HgNV and DhNV) but appear absent from other nudiviruses that do not infect crustaceans. Using these genes in addition to the conserved baculovirus core genes across DhNV, HgNV and PmNV 1,2, 35 genes were comparable in a “gene-block” fashion relative to their genomic loci (Fig. 2e). This revealed three major rearrangement events. Reordering of the DhNV_032, DhNV_034, and pif-1 gene block, which is reversed in PmNV and HgNV (Fig. 2, ‘X’). Reordering of the vlf-1 and p74 gene block, which is reversed in PmNV and HgNV (Fig. 2, ‘Y’) and a larger rearrangement of 6 genes (vlf-1, ac68, DhNV_080, ac81, and both copies of helicase 2), which is reversed in PmNV and HgNV and overlaps the ‘Y’ rearrangement event (Fig. 2, ‘Z’).
Morphological and phylogenetic comparison to other Nudiviridae
A concatenated maximum likelihood phylogenetic analysis of eight nudiviruses and one baculovirus (outgroup) using 18 core nudivirus genes (see Sect. 4) supported the positioning of DhNV outside of the two crustacean-infecting nudiviruses with bootstrap values of 100% (Fig. 3). Within this grouping, DhNV is an early branching member of the Gammanudivirus genus and may constitute a different genus altogether. The Betanudivirus genus branches outside of the Gammanudivirus cluster and the Deltanudivirus member ToNV is the earliest branching member of these three genera. The Alphanudivirus group represents the most phylogenetically distinct nudivirus genus represented on our diagram (Fig. 3).

A concatenated phylogeny using 18 core nudivirus genes among 8 nudiviruses with LoobMNV as an outgroup. Node labels indicate bootstrap support in percent. Nudivirus genera are displayed in coloured groups. Illustrations of virion morphology are presented to the right of the tree and are based off electron micrographs of electron dense cores surrounded by a membrane. Capsid length approximations and averages are displayed from relevant publications cited in the methods. Genome accession numbers include: DiNV (NC_040699), OrNV (MN623374), GbNV (NC_009240), HzNV2 (NC_004156), HgNV (MK439999), PmNV (NC_024692), DhNV (MT488302), ToNV (NC_026242) and LoobMNPV (NC_043520). The tree was annotated in FigTree v.1.4.3. For additional information on DhNV virion morphology and pathology, please consult Bojko et al.11.
Illustrations of virus morphology provide another dimension of comparison among the Nudiviridae species (Fig. 3). DhNV virions consist of a double membrane surrounding an electron-dense core measuring (n = 30, mean ± SD) 302 ± 13 nm in length and 55 ± 4 nm at its diameter 11. The rod-shaped structure is maintained across all the nudiviruses. DhNV represents one of the larger nudiviruses discovered to date, second to HzNV2, which has a length of 382 ± 30 nm.
A second concatenated maximum likelihood phylogenetic analysis of putative iap and pif-2 genes supported DhNV as an earlier branch of the crustacean-infecting nudiviruses. The addition of Macrobrachium rosenbergii nudivirus CN-SL2011 (MrNV) (NCBI:txid1217568), which only has the aforementioned genes available, branched in the Gammanudivirus genus. ToNV (Deltanudivirus) is the earliest branch of these genera, followed by HzNV2 (Betanudivirus), and the four crustacean-infecting nudiviruses. The Alphanudiviruses represent the most phylogenetically distinct lineage among nudiviruses in this tree (Fig. 4), following the same general phylogenetic theme as the details in Fig. 3.

A concatenated phylogeny using iap and pif-2 genes from 8 nudiviruses with LoobMNPV as an outgroup. Nodes are assigned bootstrap support values from 1,000 bootstrap replicates. Accession numbers include: OrNV (MN623374), GbNV (NC_009240), ToNV (NC_026242), HzNV2 (NC_004156), DhNV (MT488302), MrNV (JQ804994; JQ804993), PmNV (NC_024692), HgNV (MK439999) and LoobMNPV (NC_043520). The tree was annotated in FigTree v.1.4.3.
Discussion
We provide a full genome characterisation of DhNV, a novel member of the Nudiviridae infecting the freshwater amphipod host, Dikerogammarus haemobaphes. The genome size, ORFs and morphology of this virus correspond with related viruses from crustaceans and insects. The identification of this virus is discussed relative to its genetic and protein content, its gene synteny and the gene synteny of related viruses, and finally, its phylogenetic relatedness to other Nudiviridae. These combined data suggest a novel genus may be appropriate: Epsilonnudivirus.
A novel member of the Nudiviridae (Epsilonnudivirus) from an amphipod
Using a combination of the core genes conserved across the Nudiviridae, we show that DhNV is most related to the Gammanudivirus genus; however, with a low level of protein similarity at most loci (< 50%) it seems pertinent to explore the erection of a new genus. Our concatenated phylogenetic analysis of eight nudiviruses representing four genera is concordant with previously published trees 2. DhNV appears to branch early from the three marine nudiviruses (Figs. 3, 4), suggesting an ancestral position to the HgNV, MrNV and PmNV.
The DhNV genome encoded all core nudivirus genes, apart from p6.9, a nucleotide-binding protein. These proteins function for DNA processing, RNA transcription, and per os infectivity 13. The p6.9 gene, which is responsible for the encapsulation of the viral genome, is characterized by a serine-arginine repeat region that could not be identified from the DhNV genome and was not present at the predicted locus where p6.9 lies in other Gammanudivirus members: between lef-5 (DhNV_051) and vlf-1 (DhNV_055). In addition to the core genes (n = 24, including three repeat homologues), most genes show similarity to other Nudiviridae under a conservative e-value threshold (< 0.001), providing strong evidence that this virus belongs within the Nudiviridae (Table 1).
The primary source of protein similarity information for DhNV ORF’s came from PmNV and HgNV, two genomically characterised viruses from the Gammanudivirus genus. Gammanudivirus members contain unique apoptosis inhibitor genes that lack a predicted RING domain 1 and appear twice in the HgNV genome. DhNV_059 represents a homolog of the Iap nudivirus gene in DhNV, where an inhibitor of apoptosis repeat domain was detected but is relatively different from existing Baculovirus homologues 1. In addition to family-level gene conservation, we identified 11 “crustacean-infecting nudivirus” genes that are conserved among those that infect crustaceans. Using a gene-block approach, we identified that PmNV and HgNV share gene synteny, where the DhNV genome exhibits three reorganization events, termed ‘X’, ‘Y’ and ‘Z’ (Fig. 3). These rearrangements are visible only in this virus, alongside a low average protein similarity of ~ 50%, and may indicate that a fifth nudivirus genus could be erected to hold peracarid-infecting nudiviruses. We suggest Epsilonnudivirus. In further work, greater genomic availability of viruses from peracarid hosts could help to better define these demarcation criteria.
Further genomic characterisation of peracarid-infecting nudiviruses may also help to identify the evolutionary history of DhNV, especially with regards to genes that show relatedness outside the Nudiviridae. Examples include DhNV_029, which shares 35.19% similarity to the cg30-1 gene (YP_009186763) from Sucra jujuba nucleopolyhedrovirus (Table 1), a butterfly-infecting baculovirus. This is the first of two ORFs with zinc finger, RING-type domains in the DhNV genome; with DhNV_045 being the second (Table 1). These do show some relation to HgNV and PmNV, where both HgNV and PmNV contain three proteins with Zinc finger, RING-type domains: HgNV_019, 064, and 067 and KN57gp_003, 033, and 049 respectively 1,2. DhNV_070 also shows high similarity to a non-nudivirus organism. A hypothetical protein from Pyricularia oryzae (Table 1), the fungal pathogen that causes rice blast disease, shows 41.18% similarity to DhNV_070. Protein domain analysis using InterProScan revealed mainly cytoplasmic and disorder protein domains from the P. oryzae sequence (XP_003712544) while DhNV_070 yielded a detailed signature match to proline rich extensin, commonly found in plant cell walls. This extensin domain does not appear in any Gammanudivirus protein. Finally, DhNV_076 shows some similarity to a homologue of HgNV (LOC108666550-like protein); however, both also show high levels of similarity to ORFs of invertebrate taxa, which lacks an identified domain or function. Such a conserved gene encoded by these viruses may reflect an ancient horizontal gene acquisition from a host during their evolutionary history.
New perspectives surrounding the Nudiviridae
Nudivirus infections often delay development of their arthropod hosts, eventually causing death 4. However, high prevalence of nudiviruses in hosts apparently displaying few clinical signs of infection may also suggest some host benefit of retaining such sub-clinical infections 2,5,10,11. While the exact relationship between DhNV and host survival still requires testing, a significant association with increased activity may subsequently increase invasive capabilities of the host 11. Examining the genome of DhNV revealed several conserved and convergent traits of crustacean nudiviruses, highlighting potential genes for diagnostic development and further research into functional roles during host infection and survival within the environment. Further sequencing and characterisation of many hypothetical proteins will provide more insight into the evolutionary history and host relationship of DhNV relative to other Nudiviridae. Through genomic analysis, phylogeny, and virion morphology it is evident that the Nudiviridae in Crustacea are highly derived from their insect relatives and a great diversity of currently undescribed taxa likely reside in other arthropod hosts on land and in water.
Materials and methods
Collection of infected Dikerogammarus haemobaphes and next generation sequencing
Dikerogammarus haemobaphes were collected, dissected and underwent DNA extraction as explained by Bojko et al. 11, who also explore virion morphology and pathology associated with the discovery of a novel nudivirus. Stored DNA from a single individual displaying the characteristic pathology of bacilliform virus infection was selected for next generation sequencing using Illumina MiSeq and Illumina HiSeq. The DNA extract underwent library preparation for Illumina MiSeq using the NEXTERA XT library preparation kit, according to manufacturer’s protocol (Illumina, UK). The library underwent quality screening using a bioanalyzer (Agilent), was quantified using a QuantiFluor fluorimeter (Promega), was denatured using sodium hydroxide and diluted to 10 pM in Illumina HT1 hybridisation buffer for sequencing via an Illumina V3-600 cartridge. The same DNA extract was used to produce a library for Illumina HiSeq using the Illumina TruSeq DNA PCR-Free library preparation kit, according to manufacturer’s protocols. The library underwent quality screening using a bioanalyzer (Agilent), was quantified using a QuantiFluor fluorimeter (Promega), was denatured using sodium hydroxide and diluted to 10 pM in Illumina HT1 hybridisation buffer for sequencing on an Illumina HiSeq 3,000 with a 2 × 150 cartridge.
MiSeq and HiSeq outputs were trimmed in silico using Illuminaclip v0.32 (Trimmomatic: LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 MINLEN:36) 14 and pooled into paired and unpaired sequence files. The paired sequence data from each technique were paired-end-combined using PEAR v0.9.8 (settings: overlap similarity minimum, 20 bp) 15 to increase the read length of paired reads by combining the reads into single sequence reads. These reads were assembled using SPAdes v3.13.0 16 with default parameters and k-mer lengths 21, 33, 55, 77, 99 and 127, to produce 228,433 scaffolds with a maximum read length of 119,824 bp and minimum read length of 128 bp.
Identification and annotation of the viral genome
Scaffolds above 100,000 bp were extracted from the dataset and annotated using PROKKA v1.11 17. The subsequent output was assessed for similarity to existing sequence data using NCBI, Blastp nr database. This identified a raw contiguous sequence of 119,824 bp as the genome of DhNV, which was subsequently circularized and checked for average coverage using CLC Genomics workbench v11 to result in a genome of 119,754 bp (coverage: 157.93X). PROKKA v1.11 17 and GeneMarkS 18 was used to annotate the viral genome (parameters: virus). A combination of these two tools resulted in 95 identical open reading frames (ORFs), 8 frames with high similarity but different gene size and three ORFs identified just by PROKKA. Combined, this provided 106 ORFs for annotation. The protein product of the 106 ORFs were compared to existing information using BLASTp via the NCBI repository (GenBank) with a cut-off e-value of < 0.001. The protein sequences were also assessed using the InterProScan tool (ebi.ac.uk/interpro/) to identify domains and predicted function. Twenty-one conserved core baculovirus/nudivirus genes were identified; however, P6.9 was not found within the genome of DhNV after analysis using BLASTp, ExPASy 19, GeneMarkS and InterProScan.
The gene synteny data for DhNV was compared to two crustacean-infecting viruses, Homarus gammarus nudivirus (HgNV) and Penaeus monodon nudivirus (PmNV) (Gammanudiviridae), and an insect-infecting virus, Tipula oleracea nudivirus (ToNV) (Deltanudiviridae), whose data were obtained from NCBI accessions: KJ184318, MK439999, NC_026242, respectively. The data were plotted using Circa (omgenomics.com/circa).
The annotated viral genome is available through NCBI accession: MT488302.
Phylogenetic analysis of DhNV among the Nudiviridae
A concatenated phylogenetic tree was developed using 18 of the 21 identified nudivirus core proteins from DhNV and seven other nudiviruses: Drosophila innubila nudivirus (DiNV), Oryctes rhinocerous nudivirus (OrNV), Gryllus bimaculatus nudivirus (GbNV), Helicoverpa (syn. Heliothis) zea nudivirus-2 (HzNV2), HgNV, PmNV, and ToNV. A baculovirus outgroup, Lonomia obliqua multiple nucleopolyhedrovirus (LoobMNPV) was used to root the tree. The p47 (missing from GbNV), Helicase 2, vlf-1, and p6.9 were not included as they are not present in the genomes of all the tested nudiviruses or are duplicated in DhNV. The remaining conserved proteins, 38 k, ac81, DNA polymerase, helicase, lef-4, lef-5, lef-8, lef-9, ac92 (p33), p74 (pif-0), pif-1, pif-2, pif-3, odv-e28 (pif-4), odv-e56 (pif-5), pif-6, vp91 (pif-8) and 31 K (vp39), were aligned using Geneious v10 using MAFFT with default parameters. In HzNV2, Lef-9 was trimmed using Geneious due to its fusion with p47 1. IQtree was used to produce the maximum likelihood phylogenetic tree, which included 13,795 positions using the VT + F + I + G4 model (according to BIC) with 1,000 bootstrap replicates. Subsequently, the tree was imported into Figtree v1.4.3 for annotation. Transmission electron micrographs from each nudivirus were used to create illustrations of the virions with approximations of nucleocapsid size 2,11,20,21,22,23,24,25. A second concatenated tree was produced using putative iap and pif-2 genes from DhNV and seven other nudiviruses: OrNV, GbNV, HzNV2, HgNV, PmNV, ToNV, in addition to recently obtained Macrobrachium rosenbergii nudivirus CN-SL2011 (MrNV) (NCBI:txid1217568) sequences, which includes just two protein coding genes (iap and pif-2). DiNV was excluded from this tree as it lacks an identifiable iap ORF. The baculovirus, Lonomia obliqua multiple nucleopolyhedrovirus (LoobMNPV) was used to root the tree. Genes were trimmed using Geneious v10 and aligned using MAFFT with default parameters. IQtree produced a phylogenetic tree using the Blosum62 + G4 model (according to BIC) with 1,000 bootstrap replicates. The tree was imported into Figtree v1.4.4 for final annotation.
Data availability
Sequence data from this study are available through NCBI as stated herein. Biological materials from the host are available from the Cefas Aquatic Registry and Repository.
References
Yang, Y. T. et al. The genome and occlusion bodies of marine Penaeus monodon nudivirus (PmNV, also known as MBV and PemoNPV) suggest that it should be assigned to a new nudivirus genus that is distinct from the terrestrial nudiviruses. BMC Genomics 15, 628 (2014).
Holt, C. C. et al. The first clawed lobster virus Homarus gammarus nudivirus (HgNV n. sp.) expands the diversity of the Nudiviridae. Sci. Rep. 9, 10086 (2019).
Adams, M. J., Lefkowitz, E. J., King, A. M. Q. & Carstens, E. B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2014). Arch. Virol. 159(10), 2831–2841 (2014).
Bézier, A. et al. The genome of the nucleopolyhedrosis causing virus from Tipula oleracea (ToNV) sheds new light into the Nudiviridae family. J. Virol. 89, 3008 (2014).
Stentiford, G. D., Bateman, K. & Feist, S. W. Pathology and ultrastructure of an intranuclear bacilliform virus (IBV) infecting brown shrimp Crangon crangon (Decapoda: Crangonidae). Dis. Aquat. Organ. 58, 89–97 (2004).
Bateman, K. S. & Stentiford, G. D. Cancer pagurus bacilliform virus (CpBV) infecting juvenile European edible crabs C pagurus from UK waters. Dis. Aquat. Organ. 79, 147–151 (2008).
Bojko, J. & Ovcharenko, M. Pathogens and other symbionts of the Amphipoda: taxonomic diversity and pathological significance. Dis. Aquat. Organs. 136, 3–36 (2019).
Bojko, J. et al. Parasites, pathogens and commensals in the “low-impact” non-native amphipod host Gammarus roeselii. Parasit. Vect. 10, 193 (2017).
Bateman, K. S. & Stentiford, G. D. A taxonomic review of viruses infecting crustaceans with an emphasis on wild hosts. J. Invertebr. Pathol. 147, 86–110 (2017).
Bojko, J. et al. Green crab Carcinus maenas symbiont profiles along a North Atlantic invasion route. Dis. Aquat. Organ. 128(2), 147–168 (2018).
Bojko, J. et al. Pathogens of Dikerogammarus haemobaphes regulate host activity and survival, but also threaten native amphipod populations in the UK. Dis. Aquat. Organ. 136, 63–78 (2019).
Bojko, J. et al. Baseline histopathological survey of a recently invading island population of ‘killer shrimp’ Dikerogammarus villosus. Dis. Aquat. Organ. 106, 241–253 (2019).
Beperet, I., Simón, O., Williams, T., López-Ferber, M. & Caballero, P. The ‘11K’ gene family members sf68, sf95 and sf138 modulate transmissibility and insecticidal properties of Spodoptera frugiperda multiple nucleopolyhedrovirus. J. Invertebr. Pathol. 127, 101–109 (2015).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30(15), 2114–2120 (2014).
Zhang, J., Kobert, K., Flouri, T. & Stamatakis, A. PEAR: a fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 30, 614–620 (2013).
Nurk, S., Meleshko, D., Korobeynikov, A. & Pevzner, P. A. metaSPAdes: a new versatile metagenomic assembler. Genome Res. 27, 824–834 (2017).
Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069 (2014).
Besemer, J., Lomsadze, A. & Borodovsky, M. GeneMark S (2001) a self-training method for prediction of gene starts in microbial genomes Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 29, 2607–2618 (2001).
Gasteiger, E. et al. ExPASy: the proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 31, 3784–3788 (2003).
Lightner, D. V., Redman, R. M. & Bell, T. A. Observations on the geographic-distribution, pathogenesis and morphology of the baculovirus from Penaeus monodon Fabricius. Aquaculture 32(3–4), 209–233 (1983).
Huger, A. M. A new virus disease of crickets (Orthoptera: Gryllidae) causing macronucleosis of fatbody. J. Invertebr. Pathol. 45, 108–111 (1985).
Raina, A. K. & Adams, J. R. Gonad-specific virus of corn earworm. Nature 374, 770–770 (1995).
Wolff, J. L. C. et al. Identification and characterization of a baculovirus from Lonomia obliqua (Lepidoptera: Saturniidae). J. Invertebr. Pathol. 79, 137–145 (2002).
Huger, A. M. The Oryctes virus: Its detection, identification, and implementation in biological control of the coconut palm rhinoceros beetle, Oryctes rhinoceros (Coleoptera: Scarabaeidae). J. Invertebr. Pathol. 89, 78–84 (2005).
Unckless, R. L. A DNA Virus of Drosophila. PLoS ONE 6, e26564 (2011).
Acknowledgements
The authors acknowledge NERC funding to J.B. (Award #: 1368300); GDS and DB were funded by Cefas contract #FB002, which supported the collection of NGS data along with personal funds from DCB (University of Florida).
Author information
Authors and Affiliations
Contributions
J.B., G.D.S. and D.B. designed the study. J.B. collected the animals. J.B. and T.A. conducted the bioinformatic analyses, including phylogenetics and annotation. T.A., G.D.S., D.B., D.C.B. and J.B. contributed to the text of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Allain, T.W., Stentiford, G.D., Bass, D. et al. A novel nudivirus infecting the invasive demon shrimp Dikerogammarus haemobaphes (Amphipoda). Sci Rep 10, 14816 (2020). https://doi.org/10.1038/s41598-020-71776-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-71776-3
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
