
Abstract
Nutrients from dehydrated sewage sludge play an essential role in the development of many plants such as Terminalia argentea, in the recovery of degraded areas. The aims were to assess the abundance, diversity and species richness of phytophagous, pollinators and predators arthropods, as well as the percentage of defoliation of T. argentea trees, fertilized (or not) with dehydrated sewage sludge in a degraded area. The abundance, diversity and species richness of phytophagous Coleoptera and total predators (predator insects + protocooperating ants + spiders); abundance and species richness of Diptera, pollinator insects, spiders, and predators (predator insects + spiders) were higher on trees fertilized with dehydrated sewage sludge. The abundance of phytophagous Coleoptera declined with the presence of phytophagous Hemiptera and protocooperating ants; population of phytophagous Orthoptera declined in response to phytophagous Coleoptera and total predators; the numbers of the leafminer Lyriomyza sp. directly increased with the numbers of spiders. The ecological indices of phytophagous, pollinators, and predator arthopods increased on Terminalia argentea trees fertilized with dehydrated sewage sludge; such a better ecological indices in fertilized than in unfertilized trees, show it more suitable for the recovery of degraded areas. We discuss the competition between phytophagous insects groups as well as herbivory reduction by predators.
Similar content being viewed by others


Introduction
Sewage sludge, a residual and semi-solid material, produced as a by-product during domestic and industrial waste water treatment, is rich in organic matter, shows potential for fertilization and production of seedling substrates1,2. Sewage sludge can be used safely in agriculture and forests plantations as fertilizer and in the recovery of degraded areas, with a low-cost alternative to reduce the environmental impacts and to avoid contamination of the human food chain3,4,5. Furthermore, dehydrated sewage sludge (DSS) does not affect the heavy metal contents in grains of maize, Zea mays L. (Poales: Poaceae) and cowpea, Vigna unguiculata (L.) Walp. (Fabales: Fabaceae)6.
Terminalia argentea Mart. & Zucc (Combretaceae), a secondary native tree from the Southeastern and Central-western Brazil, is used for landscaping, wood and coal production, civil construction and the recovery of degraded areas7. Continuous release of exudates by T. argentea in the trunk is typical due to pathogens attack, affecting the constant visitation by Trigona branneri (Crockere) and Mesembrinella bicolor (Fabricius) (Hymenoptera: Apidae)8.
Insect diversity may be used to assess the recovery of degraded area, as these organisms easily respond to environmental changes9. Different orders of insects, with a large number of families and species, including Coleoptera, are widely used as a bioindicator5,10. Nutritional indices and chemical plant defenses are associated with factors such as fertilization and plant development (i.e. age), affecting phytophagous insects and therefore, the natural enemies’ diversity5,11,12,13. Sewage sludge increases the humus content in the soil and it is rich in macro (e.g. N, P and K) and micronutrients (e.g. Cu and Zn)14, favoring plants and, consequently, insect development.
The aims here were to assess for 24 months the ecological indices (abundance, diversity and species richness) and ecological processes (herbivory and predation) of phytophagous, pollinators and predators arthropods on T. argentea trees, fertilized (or not) with DSS in a degraded area. We hypothesize that (i) T. argentea trees resemble living islands, and that the fertilization with DSS may increase the canopy size (canopy islands), and thus accommodate larger numbers of phytophagous, pollinators and predators arthropods (> the equilibrium theory of island biogeography—ETIB)5,15,16,17; (ii) there is competition between groups of phytophagous insects, such as hemipterans, coleopterans and orthopterans18,19; and (iii) arthropod predators, such as insects and spiders, reduce the number of phytophagous insects and thus herbivory on T. argentea trees19,20,21.
Results
Terminalia argentea trees and arthropods
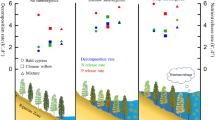
The phytophagous Coleoptera and the abundance, diversity and species richness of total predators and Diptera, pollinators, spiders, predator abundance and species richness were higher (P < 0.05) on T. argentea trees fertilized with DSS (Table 1). Percentage of defoliation and phytophagous Coleoptera Psiloptera sp. (Buprestidae), Cerambycidae, Cerotoma sp., Lamprosoma sp., Parasyphraea sp. (Chrysomelidae) and Cratosomus sp. (Curculionidae); Euxesta sp. (Diptera: Otitidae), Lepidoptera caterpillars and Tropidacris collaris Stoll (Orthoptera: Romaleidae); pollinators Trigona spinipes Fabricius (Hymenoptera: Apidae); and predators Araneidae and Salticidae (Araneae), Podisus sp. (Hemiptera: Pentatomidae), Polybia sp. (Hymenoptera: Vespidae), protocooperating ants (Hymenoptera: Formicidae) and Mantis religiosa L. (Mantodea: Mantidae) were higher (P < 0.05) on T. argentea trees fertilized with DSS (Tables 2 and 3). The abundance of Coleoptera, Diptera, Hemiptera and Orthoptera, spiders and protocooperating ants; the diversity of Coleoptera, protocooperating ants and total predator; the species richness of Coleoptera, Diptera, pollinators, protocooperating ants, spiders, predators and total predator; the percentage of defoliation; the numbers of phytophagous insects Cratosomus sp., Euxesta sp., Lamprosoma sp., Lepidoptera, Parasyphraea sp. e T. collaris, pollinators T. spinipes, and predators Pentatomidae and Polybia sp. increased with the total numbers of T. argentea leaves (Fig. 1).

Estimated network structures based on the Spearman correlation (P < 0.05) generated for total leaves per tree, defoliation (%), and the abundances of Aphirape uncifera, Araneidae, spiders, phytophagous Coleoptera, Clytini, Cratosomus sp., Diptera, Disonycha brasiliensis, Euxesta sp., protocooperating ants, phytophagous Hemiptera, Lamprosoma sp., Lepidoptera, Mantis religiosa, Lyriomyza sp. mines, phytophagous Orthoptera, Oxyopidae, Parasyphraea sp., Podisus sp., Polybia sp., Tettigoniidae, Trigona spinipes, and Tropidacris collaris; the diversity (D.) of protocooperating ants, phytophagous Coleoptera, total predators (predators + spiders + protocooperating ants), and phytophagous Orthoptera; and species richness (SR) of spiders, phytophagous Coleoptera, Diptera, protocooperating ants, Orthoptera, pollinators, predators, and total predadors on Terminalia argentea trees. n = 48.
Competition between phytophagous insects
The abundance of phytophagous Hemiptera and pollinators reduced (P < 0.05) the abundance of phytophagous Coleoptera, phytophagous Orthoptera and T. spinipes; the abundance of phytophagous Hemiptera and phytophagous Orthoptera reduced Clytini (Coleoptera: Chrysomelidae); the species richness of pollinators reduced (P < 0.05) phytophagous Coleoptera; and the species richness of phytophagous Hemiptera reduced pollinators (Table 4, Fig. 1).
Predators and phytophagous insects
The abundance of Araneidae and spiders reduced (P < 0.05) the number of the leafminer Lyriomyza sp.; the abundance of protocooperating ants reduced phytophagous Coleoptera, Euxesta sp., pollinators, and T. spinipes; the abundance of total predators reduced phytophagous Orthoptera, pollinators and T. spinipes; and the abundance of Polybia sp. reduced Clytini. The diversity and species richness of total predators reduced (P < 0.05) the numbers of phytophagous Coleoptera and Orthoptera, respectively. On the other hand, the abundance of pollinators and T. spinipes increased (P < 0.05) spiders; the abundance of T. spinipes increased Araneidae; the abundance of phytophagous Coleoptera increased total predators and the leafminer Lyriomyza sp.; the abundance of Lepidoptera caterpillars and T. spinipes increased M. religiosa; the abundance of the leafminer of Lyriomyza sp. and T. spinipes increased Podisus sp.; the abundance of Polybia sp., spiders and protocooperating ants increased phytophagous Hemiptera; the abundance of Oxyopidae (Araneae) increased Euxesta sp.; the abundance of Cratosomus sp., phytophagous Hemiptera and Orthoptera increased Araneidae; the abundance of Disonycha brasiliensis Lima (Coleoptera: Chrysomelidae) and Tettigoniidae (Orthoptera) increased Aphirape uncifera Tullgren (Araneae: Salticidae). The diversity of Orthoptera increased (P < 0.05) the abundance of A. uncifera; the diversity of phytophagous Hemiptera increased protocooperating ants; the species richness of phytophagous Coleoptera and pollinators increased the abundance of M. religiosa and Podisus sp.; and the species richness of phytophagous Orthoptera and pollinators increased the abundance of Araneidae (Table 4, Fig. 1).
Discussion
The highest ecological indices (abundance, diversity and species richness) of phytophagous, pollinator and predators arthropods on T. argentea, fertilized with dehydrated sewage sludge (DSS), are related to a higher nitrogen levels6 and consequently a better development of these plants (e.g. > leaves/tree = > ETIB)5,15. The apparent competition between Coleoptera and Hemiptera for space and food, and the negative effect between protocooperating ants and phytophagous Coleoptera, are in accordance to findings on Caryocar brasiliense Camb. (Malpighiales: Caryocaraceae) trees19,21.
The highest ecological indices of phytophagous Cerambycidae, Cerotoma sp., Cratosomus sp., Euxesta sp., Lamprosoma sp., Lepidoptera caterpillars, Parasyphraea sp., Psiloptera sp., T. collaris and Hemiptera, ; pollinator T. spinipes; predators Araneidae, Salticidae, Pentatomidae and Polybia sp.; protocooperating ants and ecological processes (herbivory) on T. argentea trees fertilized with DSS, may be due to the highest numbers of leaves of this plant (> ETIB). Leaves are food resource with a better quality for these phytophagous insects, which in turn may attract a higher number of predators. Such an observation confirms the first hypothesis (i.e. ETIB), that the diversity and abundance of phytophagous insects, pollinators and their predators are usually higher on larger trees with higher leaf mass5,15,16,17. Thus, trees such as T. argentea, may seem as islands (as proposed by ETIB), and those with lower leaf mass present a higher chance to get extinct the endangered species5,17,22,23. In addition, the number of free amino acids and proteins in leaves, pollen and/or nectar production and quality (more protein and amino acids) in flowers, are superior in plants with higher nitrogen fertilization levels, e.g. T. argentea trees fertilized with DSS, increasing the attractiveness to phytophagous and pollinator insects6,24,25,26. Dehydrated sewage sludge used as a biofertilizer improved the macrofauna recovery, including scarab beetles’ larvae and adults in degraded soils of the Cerrado (Brazilian Savanna) biome27.
The abundance of phytophagous Hemiptera and pollinators (e.g. T. spinipes) reduced the number of phytophagous Coleoptera; whilst this insect order reduced the numbers of Orthoptera and T. spinipes, as well as those of phytophagous Hemiptera and phytophagous Orthoptera reduced Clytini. These correlations confirm the second hypothesis that there was competition between those insect groups for space and feeding. Moreover, protocooperating ants, associated with phytophagous Hemiptera, for instance, may have attacked beetles. However, further studies are needed to elucidate this hypothesis. Competition between defoliators (e.g. Coleoptera), sucking and galling insect species for space and feeding was observed on C. brasiliense trees17,19.
Trigona spinipes, by flying in flocks with aggressive behavior, chases other pollinators, such as Apis mellifera L. and Tetragonisca angustula Latreille (Hymenoptera: Apidae)28, and also likely other insects (e.g. beetles); beyond damages shoot and plant growth tissues to remove fibers for nests construction5,29,30. Food web studies are intricate due to interactions among host plants, phytophagous, predators and parasitoids insects, soil and climatic conditions31. Only a few studies have examined food webs in complex ecosystems, such as in the Cerrado18,31,32.
Spiders, the dominant predators group (excluding the protocooperating ants), correlated negatively with some phytophagous insects (e.g. Lyriomyza sp. and Orthoptera), confirming the third hypothesis on the negative correlation between phytophagous insects and predators. On the other hand, T. spinipes is perhaps the major prey to spiders on T. argentea trees. Spiders are important in the biological control of phytophagous (r = − 0.73; P = 0.00) and leafminer insects (r = − 0.62; P = 0.01) on C. brasiliense trees19,21. Spiders are important in pest control in agroforestry systems, especially in tropical regions21,33,34,35 since a wide range of pest insects can get caught in their webs, resulting in deaths36. The importance of these arthropods for biological control was confirmed by population reduction of Epiphyas postvittana (Walker) (Lepidoptera: Tortricidae) on Malus domestica Bork (Rosaceae) and Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae) on Citrus sinensis (L.) Osbeck (Rutaceae)37,38. In addition to spiders, the protocooperating ants were very abundant on T. argentea trees fertilized with DSS, probably due to the highest numbers of phytophagous insects—protocooperation39,40,41. The increased abundance of protocooperating ants reduced the numbers of phytophagous Coleoptera and T. spinipes on T. argentea trees, as observed in C. brasiliense, where the highest number of these ants reduced defoliation by beetles19,21. In addition, ants are bioindicators in the recovery of degraded area because they respond quickly to environmental complexity and by interacting mutually with other insects42,43,44,45. The abundance of the predatory wasp Polybia sp. was higher on fertilized plants probably due to a higher numbers of caterpillars (Lepidoptera) and the leafminer Lyriomyza sp.. Predatory wasps (Vespidae) are important natural enemies in agricultural systems such as Brassica campestris L. and kale B. oleracea L. var. acephala DC (Brassicales: Brassicaceae); Arabian coffee Coffea arabica L. (Gentianales: Rubiaceae) and tomato Solanum lycopersicon L. (Solanales: Solanaceae), preying mainly on caterpillars and leafminers (Lepidoptera)46,47,48,49.
In general, arthropod predators on T. argentea trees reduced herbivory by insects. However, in a few cases the presence of arthropod predators increased the numbers of phytophagous such as the leafminer Lyriomyza sp., likely by reducing competition with other more dominant groups (e.g. phytophagous Coleoptera). It shows how complex are interactions in food webs in natural and agroforestry systems18,19,21,31,32,36. Predators are often generalist in their feeding habits, and the greatest complexity of canopy architecture increases niches options for phytophagous insects and consequently for the natural enemy diversity50. For example, sewage sludge increases the richness of the ground beetle Carabidae (Coleoptera) in the area of Oxford, USA51.
The largest T. argentea tree canopy size (> ETIB) fertilized with DSS may explain the largest abundance of phytophagous insects (> defoliation), pollinators and predators, showing that this plant is adequate to recovery degraded areas. There was competition between groups of phytophagous insects and predator arthropods in high populations and consequent herbivory reduction.
Material and methods
Study
The study was conducted in a degraded area at the “Instituto de Ciências Agrárias (ICA)” of the “Universidade Federal de Minas Gerais (UFMG)”, Montes Claros, Minas Gerais, Brazil (S 16º51′38″ W 44º55′00″ 943 m) from March 2015 to February 2017 (24 months; arthropod collection period). The area presents soil loss and changes in soil chemistry and hydrology due to degradation52,53. Köppen’s climate classification54 defines this area as tropical dry climate; annual rainfall, 1,000–1,300 mm, with dry winter; annual mean temperature, ≥ 18 °C. The type of soil is litolic neosoil55 and chemical and physical details were described5.
Study design
Seeds were collected from five-years old Terminalia argentea trees at ICA/UFMG campus before sowing. Terminalia argentea seedlings were produced in March 2014 by sowing one seed per plastic polybag (8 × 12 cm), and these kept in a nursery covered with black shed net. The mixed substrate contained 30% organic materials (i.e. two parts of debris gardening pruning < 5 cm in length, and one of brown bovine manure), 30% clay soil, 30% sand, and 10% of mineral fertilizer (i.e. 160 g reactive natural phosphate per seedling)5. The soil pH in the pits (40 × 40 × 40 cm) was corrected with dolomitic limestone with anhydrous carbonate mineral composed of calcium magnesium carbonate (90% relative total neutralization power) (187 g per pit), increasing base saturation to 50%56. Natural phosphate (80 g per pit), fritted trace elements (FTE) (10 g/pit), and marble roch dust (1 kg per pit) were added when needed. Thirty-centimeters tall T. argentea seedlings were planted in pits in a two-meters spacing , in six parallel lines on flat terrain with two-meters spacing lines, with four trees per treatment (fertilized or not with dehydrated sewage sludge—DSS) per line. The seedlings in experimental area were supplied with water until the beginning of the rainy season. The seedlings with five-cm long branches were pruned with a sterilized razor, eliminating the additional shoots (i.e. others different from the leader shoot) and branches up to 1/3 of crown height. The experimental design was in random blocks with two levels of fertilization (i.e. a single dose of 20 L of DSS per pit or none fertilization) and 24 replications with one plant each5.
DSS (with 5% mean moisture content) was obtained from a sewage treatment plant (STP) in Juramento, Minas Gerais, Brazil. The STP is operated by the Minas Gerais Sanitation Company – “Companhia de Saneamento de Minas Gerais S.A. (COPASA)”. The STP is highly efficient, removing more than 90% of the organic material from the domestic waste water. The sewage sludge is dumped off into coarse sand tanks, staying there for three months to reduce the amount of thermotolerant coliforms (and other pathogenic microrganisms) and reach the ideal levels for agricultural use that is < 103 of the most likely number per g of total solids (as recommended by the National Council for the Environment—“Conselho Nacional do Meio Ambiente—CONAMA”). The chemical and biological characteristics of the DSS were described5,6.
Arthropods
Insects and spiders were visually counted, every two weeks, on the adaxial and abaxial surfaces of the leaves between 7:00 and 11:00 AM at the apical, middle and basal canopy in the northerly, southerly, easterly and westerly directions, in 12 leaves per plant (i.e. 27,648 leaves from 48 T. argentea trees) during 24 months. Only insects and spiders collected for identification were removed from trees during the assessment. At least three specimens per insect or spider species were collected using aspirator, stored in glass flasks with 70% ethanol or mounted, separated into morphospecies, and sent for identification. Insect defoliation was assessed visually as the leaf area loss on a 0–100% scales with 5% increments for removed leaf area57,58.
Ecological indices
To avoid pseudoreplication, mean numbers of data per tree were ever used. Ecological indices (abundance, diversity, and species richness) were calculated for each species per tree in the treatments (fertilization or not with DSS) using the software BioDiversity Professional, Version 259. The arthropod diversity was calculated using the Hill’s formula60,61 and the species richness with the Simpson indices62,63. The predator (i.e. insects and spiders) and prey ratio on T. argentea was calculated per tree. Predators were classified as spiders (most important group), predators (predators + spiders) and total predators (spiders + predators + protocooperating ants).
Statistical analyses
Data on defoliation percentage, abundance, diversity, and species richness of phytophagous insects, pollinators and predators were submitted to non-parametric statistical hypothesis, the Wilcoxon signed-rank test (P < 0.05)64, using the statistical program “Sistema para Análises Estatísticas e Genéticas” (SAEG), version 9.165. Simple regression analyzes and parameters (P < 0.05) were performed with SAEG to test the interactions between groups of phytophagous, pollinators and predators, and foliar mass (see41).
The Spearman correlation matrix, among the most significant characteristics, was calculated. The matrices were submitted to correlation networks66. Edge thickness was controlled by application of a cut value of 0.28 (from which the Spearman correlation becomes significant, meaning that only edges with |rij|≥ 0.28 are highlighted). These analyses were performed in R version 3.4.167. The correlation network procedure was performed using the package qgraph66.
Ethics
No specific permits are required to Terminalia argentea tree in Brazil. The laboratory and field studies did not involve endangered or protected species.
Data availability
All data generated or analyzed during this study are included in this manuscript.
References
Frišták, V., Pipíška, M. & Soja, G. Pyrolysis treatment of sewage sludge: a promising way to produce phosphorus fertilizer. J. Clean. Prod. 172, 1772–1778. https://doi.org/10.1016/j.jclepro.2017.12.015 (2018).
Antonkiewicz, J. et al. A mixture of cellulose production waste with municipal sewage as new material for an ecological management of wastes. Ecotoxicol. Environ. Saf. 169, 607–614. https://doi.org/10.1016/j.ecoenv.2018.11.070 (2019).
Kimberley, M. O., Wang, H., Wilks, P. J., Fisher, C. R. & Magesan, G. N. Economic analysis of growth response from a pine plantation forest applied with biosolids. Forest Ecol. Manag. 189, 345–351. https://doi.org/10.1016/j.foreco.2003.09.003 (2004).
Caldeira, M. V. W. et al. Lodo de esgoto como componente de substrato para produção de mudas de Acacia mangium Wild. Comun. Sci. 5, 34–43 (2014).
Silva, J. L. et al. Diversity of arthropods on Acacia mangium (Fabaceae) and production of this plant with dehydrated sewage sludge in degraded area. R. Soc. Open Sci. 7, 2. https://doi.org/10.1098/rsos.191196 (2020).
Nogueira, T. A. R. et al. Metais pesados e patógenos em milho e feijão caupi consorciados, adubados com lodo de esgoto. Rev. Bras. Eng. Agríc. Ambient. 11, 331–338. https://doi.org/10.1590/S1415-43662007000300014 (2007).
Lorenzi, H. Árvores Brasileiras: Manual de Identificação e Cultivo de Plantas Arbóreas Nativas do Brasil (Instituto Plantarum, Nova, 2002).
Boff, S., Graciolli, G., Boaretto, A. G. & Marques, M. R. Visiting insects of exudated gums by Terminalia argentea Mart. & Zucc (Combretaceae). Rev. Bras. Entomol. 52, 477–479. https://doi.org/10.1590/S0085-56262008000300025 (2008).
Santos, M. S. et al. Riqueza de formigas (Hymenoptera, Formicidade) da serapilheira em fragmentos de floresta semidecídua da Mata Atlântica na região do Alto do Rio Grande, MG, Brasil. Iheringi. Sér. Zool. 96, 95–101. https://doi.org/10.1590/S0073-47212006000100017 (2006).
Davis, A. J. et al. Dung beetles as indicators of change in the forests of northern Borneo. J. Appl. Ecol. 38, 593–616. https://doi.org/10.1046/j.1365-2664.2001.00619.x (2001).
Bowers, M. D. & Stamp, N. E. Effects of plant-age, genotype, and herbivory on Plantago performance and chemistry. Ecology 74, 1778–1791. https://doi.org/10.2307/1939936 (1993).
Coley, P. D. & Barone, J. A. Herbivory and plant defenses in tropical forests. Annu. Rev. Ecol. Syst. 27, 305–335, https://doi.org/10.1146/annurev.ecolsys.27.1.305 (1996.)
Leite, G. L. D., Picanço, M., Zanuncio, J. C., Moreira, M. D. & Jham, G. N. Hosting capacity of horticultural plants for insect pests in Brazil. Chil. J. Agric. Res. 71, 383–389. https://doi.org/10.4067/S0718-58392011000300006 (2011).
Mass, K. D. B. Biossólido Como Substrato na Produção de Mudas de Timburi. Graduate theses and dissertations (2010).
MacArthur, R. H. & Wilson, E. O. The Theory of Island Biogeography (Princeton University Press, Princeton, 1967).
Espírito Santo, M. M., Neves, F. S., Andrade Neto, F. R. & Fernandes, G. W. Plant architecture and meristem dynamics as the mechanisms determining the diversity of gall-inducing insects. Oecologia 153, 353–364. https://doi.org/10.1007/s00442-007-0737-8 (2007).
Leite, G. L. D. et al. Architectural diversity and galling insects on Caryocar brasiliense trees. Sci. Rep. 7, 16677. https://doi.org/10.1038/s41598-017-16954-6 (2017).
Morris, R. J., Lewis, O. T. & Godfray, H. C. J. Experimental evidence for apparent competition in a tropical forest food web. Nature 428, 310–313. https://doi.org/10.1038/nature02394 (2004).
Leite, G. L. D. et al. Habitat complexity and Caryocar brasiliense herbivores (Insecta; Arachnida; Araneae). Fla. Entomol. 95, 819–830. https://doi.org/10.1653/024.095.0402 (2012).
Auslander, M., Nevo, E. & Inbar, M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. J. Arid Environ. 55, 405–416. https://doi.org/10.1016/S0140-1963(02)00281-1 (2003).
Leite, G. L. D. et al. Seasonal damage caused by herbivorous insects on Caryocar brasiliense (Caryocaraceae) trees in the Brazilian savanna. Rev. Colombiana Entomol. 38, 108–113 (2012).
Kitahara, M. & Fujii, K. An island biogeographical approach to the analysis of butterfy community patterns in newly designed parks. Res. Popul. Ecol. 91, 23–35. https://doi.org/10.1007/BF02765247 (1997).
Burns, K. C. Native–exotic richness relationships: a biogeographic approach using turnover in island plant populations. Ecology 97, 2932–2938, https://doi.org/10.1002/ecy.1579 (2016)
Taiz, L., Zeiger, E., Moller, I. M. & Murphy, A. Fisiologia e desenvolvimento vegetal. (Artmed, 2017).
Tiedge, K. & Lohaus, G. Nectar sugars and amino acids in day- and night-flowering Nicotiana species are more strongly shaped by pollinators’ preferences than organic acids and inorganic ions. PLoS ONE 12, 1–25. https://doi.org/10.1371/journal.pone.0176865 (2017).
Stabler, D., Power, E. F., Borland, A. M., Barnes, J. D. & Wright, G. A. A method for analysing small samples of floral pollen for free and protein-bound amino acids. Methods Ecol. Evol. 9, 430–438. https://doi.org/10.1111/2041-210X.12867 (2018).
Kitamura, A. E., Alves, M. C., Sanches, S., Akihiro, L. G. & Antonio, P. G. Recuperação de um solo degradado com a aplicação de adubos verdes e lodo de esgoto. Rev. Bras. Cienc. Solo 32, 405–416. https://doi.org/10.1590/S0100-06832008000100038 (2008).
Serra, B. D. V. & Campos, L. A. Polinização entomófila de abobrinha, Cucurbita moschata (Cucurbitaceae). Neotrop. Entomol. 39, 153–159. https://doi.org/10.1590/S1519-566X2010000200002 (2010).
Silva, F. W. S. et al. Spatial distribution of arthropods on Acacia mangium (Fabales: Fabaceae) trees as windbreaks in the Cerrado. Fla. Entomol. 97, 631–638. https://doi.org/10.1653/024.097.0240 (2014).
Damascena, J. G. et al. Spatial distribution of phytophagous insects, natural enemies, and pollinators on Leucaena leucocephala (Fabaceae) trees in the Cerrado. Fla. Entomol. 100, 558–565. https://doi.org/10.1653/024.100.0311 (2017).
Gratton, C. & Denno, R. F. Seasonal shift from bottom-up to top-down impact in phytophagous insect populations. Oecology 134, 487–495. https://doi.org/10.1007/s00442-002-1137-8 (2003).
Marquis, R. J., Diniz, I. R. & Morais, H. C. Patterns and correlates of the interspecific variation in foliar insect herbivory and pathogen attack in Brazilian Cerrado. J. Trop. Ecol. 17, 127–148. https://doi.org/10.1017/S0266467401001080 (2001).
Landis, D., Wratten, S. D. & Gurr, G. M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Ann. Rev. Entomol. 45, 175–201. https://doi.org/10.1146/annurev.ento.45.1.175 (2000).
Langellotto, G. A. Aggregation of Invertebrate Predators in Complex-Structured Habitats: Role of Altered Cannibalism, Intraguild Predation, Prey Availability, and Microclimate. Graduate theses and dissertations (2003).
Halaj, J., Halpern, C. B. & Yi, H. Responses of litter-dwelling spiders and carabid beetles to varying levels and patterns of green-tree retention. Forest Ecol. Manag. 255, 887–900. https://doi.org/10.1016/j.foreco.2007.09.083 (2008).
Sunderland, K. & Samu, F. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: a review. Entomol. Exp. Appl. 95, 1–13. https://doi.org/10.1046/j.1570-7458.2000.00635.x (2000).
Amalin, D. M., Reiskind, J., Peña, J. E. & Mcsorley, R. Predatory behavior of three species of sac spiders attacking citrus leafminer. J. Arachnol. 29, 72–81. https://doi.org/10.1636/0161-8202(2001)029[0072:PBOTSO]2.0.CO;2 (2001).
Hogg, B. N., Mills, N. J. & Daane, K. M. Temporal patterns in the abundance and species composition of spiders on host plants of the invasive moth Epiphyas postvittana (Lepidoptera: Tortricidae). Environ. Entomol. 146, 502–510. https://doi.org/10.1093/ee/nvx065 (2017).
Del-Claro, K. & Oliveira, P. S. Conditional outcomes in a neotropical treehopper-ant association: temporal and species-specific variation in ant protection and homopteran fecundity. Oecologia 124, 156–165. https://doi.org/10.1007/s004420050002 (2000).
Schmitz, O. J. Indirect Effects in Communities and Ecosystems: The Role of Trophic and Nontrophic Interactions (Princeton University Press, Princeton, 2006).
Costa, S. S. D. Insects and growth of Terminalia argentea Mart & Zucc (Combretaceae) fertilized with dehydrated sewage sludge. Graduate theses and dissertations (2019).
Economo, E. P. et al. Global phylogenetic structure of the hyperdiverse ant genus Pheidole reveals the repeated evolution of macroecological patterns. Proc. R. Soc. B. 282, 1–10. https://doi.org/10.1098/rspb.2014.1416 (2014).
Pérez-Lachaud, G. & Lachaud, J. P. Arboreal ant colonies as ‘hot-points’ of cryptic diversity for myrmecophiles: the weaver ant Camponotus sp. aff. textor and its interaction network with its associates. PLoS ONE 9, 1–8. https://doi.org/10.1371/journal.pone.0100155 (2014).
Chomicki, G., Ward, P. S. & Renner, S. S. Macroevolutionary assembly of ant/plant symbioses: Pseudomyrmex ants and their ant-housing plants in the Neotropics. Proc. R. Soc. B. 282, 1–9. https://doi.org/10.1098/rspb.2015.2200 (2015).
Sanches, A. Fidelity and promiscuity in an ant-plant mutualism: a case study of triplaris and Pseudomyrmex. PLoS ONE 10, 1–19. https://doi.org/10.1371/journal.pone.0143535 (2015).
Miranda, M. M. M., Picanço, M., Leite, G. L. D., Zanuncio, J. C. & Clercq, P. Sampling and non-action levels for predators and parasitoids of virus vectors and leaf miners of tomato in Brazil. Med. Facul. Landb. Univ. Gent 63, 519–523 (1998).
Leite, G. L. D., Oliveira, I. R., Guedes, R. N. C. & Picanço, M. Comportamento de predação de Protonectarina sylveirae (Saussure) (Hymenoptera: Vespidae) em mostarda. Agro-Ciencia (Chillán) 17, 93–96 (2001).
Picanço, M., Ribeiro, L. J., Leite, G. L. D. & Gusmão, M. R. Seletividade de inseticidas a Polybia ignobilis (Haliday) (Hymenoptera: Vespidae) predador de Ascia monuste orseis (Godart) (Lepidoptera: Pieridae). An. Soc. Entomol. Bras. 27, 85–90. https://doi.org/10.1590/S0301-80591998000100011 (1998).
Picanço, M. C. et al. Ecology of Vespidae (Hymenoptera) predators in Coffea arabica plantations. Sociobiology 59, 1269–1280 (2012).
Condon, M. A. et al. Lethal interactions between parasites and prey increase niche diversity in a tropical community. Science 343, 1240–1244. https://doi.org/10.1126/science.1245007 (2014).
Larsen, K. J., Purrington, F. F., Brewer, S. R. & Taylor, D. H. Influence of sewage sludge and fertilizer on the ground geetle (Coleoptera: Carabidae) fauna of an old-field community. Environ. Entomol. 25, 452–459. https://doi.org/10.1093/ee/25.2.452 (1996).
Milton, S. J., Dean, W. R. J., du Plessis, M. A. & Siegfried, W. R. A conceptual model of arid rangeland degradation. Bioscience 44, 70–76. https://doi.org/10.2307/1312204 (1994).
Whisenant, S. G. Repairing damaged wildlands: a process-oriented, landscape-scale approach. Restor. Ecol. 9, 249–249. https://doi.org/10.1046/j.1526-100x.2001.009002249.x (1999).
Alvares, C. A., Stape, J. L., Sentelhas, P. C., Gonçalves, J. L. M. & Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 22, 711–728. https://doi.org/10.1127/0941-2948/2013/0507 (2013).
Santana, P. H. L., Frazão, L. A., Santos, L. D. F., Fernandes, L. A. & Sampaio, R. A. Soil attributes and production of Eucalyptus in monoculture and silvopastoral systems in the north of Minas Gerais, Brazil. J. Agric. Sci. Technol. 6, 361–370. https://doi.org/10.17265/2161-6264/2016.06.001 (2016).
Kopittke, P. M. & Menzies, N. W. A review of the use of the basic cation saturation ratio and the “ideal” soil. Soil Sci. Soc. Am. J. 71, 259–265. https://doi.org/10.2136/sssaj2006.0186 (2007).
Sastawa, B. M., Lawan, M. & Maina, Y. T. Management of insect pests of soybean: effects of sowing date and intercropping on damage and grain yield in the Nigerian Sudan savanna. Crop Prot. 23, 155–161. https://doi.org/10.1016/j.cropro.2003.07.007 (2004).
Mizumachi, E., Mori, A., Osawa, N., Akiyama, R. & Tokuchi, N. Shoot development and extension of Quercus serrata saplings in response to insect damage and nutrient conditions. Ann. Bot. 98, 219–226. https://doi.org/10.1093/aob/mcl091 (2006).
Krebs, C. J. Bray-Curtis cluster analysis. 1. Biodiversity Pro Versão 2. https://biodiversity-pro.software.informer.com (1989).
Hill, M. O. Diversity and evenness: a unifying notation and its consequences. Ecology 54, 427–432. https://doi.org/10.2307/1934352 (1973).
Jost, L. Entropy and diversity. Oikos 113, 363–375. https://doi.org/10.1111/j.2006.0030-1299.14714.x (2006).
Begon, M., Townsend, C. R., Harper, J. L. Ecologia: de indivíduos a ecossistemas. (Artmed, 2007).
Lazo, J. A., Valdés, N. V., Sampaio, R. A. & Leite, G. L. D. Diversidad zoológica asociada a um silvopastoreo leucaena-guineacon diferentes edades de establecimiento. Pesq. Agropec. Bras. 42, 1667–1674. https://doi.org/10.1590/S0100-204X2007001200001 (2007).
Wilcoxon, F. Individual comparisons by ranking methods. Biometrics Bull. 1, 80–83. https://doi.org/10.2307/3001968 (1945).
SAEG—Sistema para Análises Estatísticas. https://arquivo.ufv.br/saeg/. (accessed on 30 june 2018) (2007).
Epskamp, S., Cramer, A. O. J., Waldorp, L. J., Schmittmann, V. D. & Borsboom, D. qgraph: Network visualizations of relationships in psychometric data. J. Stat. Softw. 48, 1–18. https://doi.org/10.18637/jss.v048.i04 (2012).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. https://www.R-project.org/ (2014).
Acknowledgements
We would like to thank to Dr. Antônio Domingos Brescovit (Instituto Butantan, São Paulo, Brasil—Aracnidae) and Dr. Ayr de Moura Bello (Fundação Oswaldo Cruz, Rio de Janeiro, Brasil—Coleoptera) for arthropod species identifications. The voucher number for spiders is IBSP 36921–36924 (Instituto Butantan, São Paulo, Brasil) and that of insects is 1595/02 and 1597/02 (Centro de Estudos Faunísticos e Ambientais, Universidade Federal do Paraná, Curitiba, Paraná, Brasil). The study was financially supported by the following Brazilian agencies “Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)”, “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES-Finance Code 001)”, “Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG)”, and “Programa Cooperativo sobre Proteção Florestal (PROTEF)” of the “Instituto de Pesquisas e Estudos Florestais (IPEF)”.
Author information
Authors and Affiliations
Contributions
The study was conceived and designed by G.L.D.L. and M.A.S., data were collected by F.W.S.S. and J.C.N.C. with support from G.L.D.L., data analysis was performed by G.L.D.L. and A.M.A., figures, tables and manuscript preparation was done by G.L.D.L., F.W.S.S., G.L.T., A.M.A., M.A.S., J.C.Z. and J.C.L. All authors contributed to revisions and approve the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Carvalho, J.C.N., Silva, F.W.S., Leite, G.L.D. et al. Does fertilization with dehydrated sewage sludge affect Terminalia argentea (Combretaceae) and associated arthropods community in a degraded area?. Sci Rep 10, 11811 (2020). https://doi.org/10.1038/s41598-020-68747-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-68747-z
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
