
Abstract
Using animal cells and tissues as precise measuring devices for developing new drugs presents a long-standing challenge for the pharmaceutical industry. Despite the very significant resources that continue to be dedicated to animal testing of new compounds, only qualitative results can be obtained. This often results in both false positives and false negatives. Here, we show how the effect of drugs applied to animal ventricular myocytes can be translated, quantitatively, to estimate a number of different effects of the same drug on human cardiomyocytes. We illustrate and validate our methodology by translating, from animal to human, the effect of dofetilide applied to dog cardiomyocytes, the effect of E-4031 applied to zebrafish cardiomyocytes, and, finally, the effect of sotalol applied to rabbit cardiomyocytes. In all cases, the accuracy of our quantitative estimates are demonstrated. Our computations reveal that, in principle, electrophysiological data from testing using animal ventricular myocytes, can give precise, quantitative estimates of the effect of new compounds on human cardiomyocytes.
Similar content being viewed by others

Introduction
Our main goal was to develop model-based methods evaluating whether measurements of drug effects on animal cardiomyocytes can be translated to corresponding drug effects on human cardiomyocytes. The analysis reveals how effects on membrane currents carried by the same ion channel protein in animal and humans can be translated in order to evaluate and understand human ventricular action potentials. This approach provides a framework revealing unwanted side effects early in the development of novel drug candidates. The usefulness of animal models in understanding human electrophysiology is unquestioned, but limited by the inherent difference between the human and animal action potentials; see e.g.1,2,3. Often, this gives rise to difficulties in direct translation; even if some of the ion channels involved are functionally nearly identical, the overall composition of membrane ion channels can differ substantially between species. Therefore, block of the same ion channel in two different species may yield very different perturbations of the respective action potentials.
This limitation has serious implications for the overall efficiency and effectiveness of drug development. Currently, this process takes 10 to 15 years, with an average cost of ~2.5 billion USD4. Preclinical development, including animal testing for safety predictions, accounts for ~60% of these costs5. However, despite this cost and effort, many promising candidate drugs exhibit toxicity that has not been predicted prior to clinical trials and, ultimately, emerging therapies6; clearly, more quantitatively predictive tools are needed. Assuming that the major currents underlying the ventricular action potential are governed by the same or similar ion channel alpha subunit in an animal and in a human heart cell, and that these proteins have comparable function and are relatively well understood1,7,8,9,10,11, our approach can be used to translate measured pharmacological effects on the action potential in animal models to those in healthy human adult cardiomyocytes.
Previously12,13, we have developed a theoretical approach which allowed translation of measurements obtained on human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) to adult cardiomyocytes. In brief, we have demonstrated that after inverting hiPSC-CM data, and finding the effect of a drug candidate on every ion channel, these findings can be accurately mapped to the case of mature cardiomyocytes. Thus, the effect of the drug on adult human heart cells can be deduced from measurements of hiPSC-CMs. Our aim here is to show that a similar approach can be employed in translation from animal myocytes to human results.
In order to fully translate the effect of a drug from data obtained from either hiPSC-CMs or animals, the effect of the drug must be used to estimate parameters of the associated mathematical model. This is indeed a challenging process; see e.g.14,15,16 with significant uncertainty. Here, in order to focus on the trans-species mapping, we use the techniques developed in13, simplified considerably by assuming that we know which of the ion channels that are affected by a given drug. In this simplified case it is sufficient to only have data on the transmembrane potential.
Results
Here, we demonstrate that it is possible to estimate the effect of a drug candidate on humans solely based on measurements of analogous effects on animal cardiomyocytes. The dog data presented in7 is employed together with our method to estimate the effect that dofetilide will have on humans. Also, in7, the effect of dofetilide on human cardiomyocytes was reported, allowing us to assess the quality of our estimates. Additionally, we used the data provided in17 on zebrafish to estimate the effect of the drug E-4031 on humans. While the effect on human cardiomycytes is not reported in17, this has been reported in18,19, which we again used to assess the accuracy of our estimates. Finally, we used rabbit data presented in20 to estimate the effect of sotalol on human myocytes; data on human cardiomyocytes in response to sotalol has been presented in20,21 and is used here for comparison and assessment.
Quantitative translation from dog to human
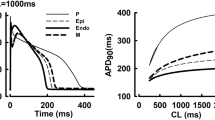
In Fig. 1, our method (see below) is applied to data published in7. In the left panel, experimental data (dotted) is shown for control (blue) and when 50 nM of the IKr blocker dofetilide is applied (red) to canine myocytes. Solid lines show the results of the mathematical model (Supplementary Information). The parameters defining a control model and an IC50 value representing the effect of the drug are estimated and used to model the no drug case (solid blue line) and when the drug has been applied (solid red line).

Dog (left) and human (right) ventricular action potentials in the control case and in the presence of 50 nM of the IKr blocker dofetilide. The dotted lines show measured data from7, and solid lines show simulation results. Note that the drug effect used in the simulation of the human AP was estimated from the dog data.
In the right panel, the control data (dotted blue) and the control mathematical model (solid blue) are illustrated for the human ventricular myocyte. We used the method to estimate the effect of the IKr blocker on the ventricular AP model of the dog, and this effect is translated (Supplementary Information) to the human AP model; this estimated AP (solid red) fits the measured data from7 very well.
In Table 1, we compare selected biomarkers (APD50 and APD90) for the control case of dog cardiomyocytes (measured), dog cardiomyocytes subjected to the drug (measured), the human control (measured), the human control (model), human cardiomyocytes subjected to the drug (measured) and human cardiomyocytes subjected to the drug (estimated as based on the dog data, as described in the Supplementary Information). All experimental data are taken from7. For the dog data, the APD50 is increased by 17.5% when the drug is added while, for human cells, the measured increase in APD50 is 33.3%. When we compare the output from the mathematical model for the control case, and the one where the effect of the drug is added to the model, we observe an increase of 38.0%. Thus, based only on data from the dog, our estimate of the increase of the APD50 for humans differs little from the measured value (38.0 vs 33.3%). The analysis of the APD90 also reveal a similarly small difference (40.2 vs. 44.3%). We conclude that the dog data on APD50/APD90 can be used to estimate the effect of the IKr blocker dofetilide on human cardiomyocytes well. Furthermore, the mathematical model estimates an IC50 value of 17 nM for block of IKr by dofetilide. This value is in good agreement with IC50 values for IKr block by dofetilide found in literature (1–70 nM21,22,23,24).
Quantitative translation from zebrafish to human
In Table 2, the same method was applied to data from zebrafish cardiomyocytes17. Again, we have focused on experimental APD50 and APD90 values for zebrafish in the control case and the increase caused by the application of 1 μM of the IKr blocker E-4031. In addition, we report the increases in APD50/APD90 from measurements of human cardiomyocytes exposed to 1 μM E-4031 from Bussek et al.18 (B) and Jost et al.19 (J).
Finally, we report the increases in APD50 and APD90 estimated for human cardiomyocytes as based on the measurements of zebrafish cardiomyocytes. Again, we observe that the APD changes estimated using the mathematical model for the human case agree well with the measured effects for human cells. For APD50, our method estimates a 58.0% increase, while the increase is 27.7% (B) or 54.2% (J) in the experimental data. Similarly, the method estimates a 62.8% increase of APD90, while the experimental data reports a 31.2% (B) or 65.1% (J) increase. Furthermore, the method estimates an IC50 value of 44 nM for block of IKr by E-4031, in line with corresponding IC50 values found in literature (10–397 nM25,26,27,28,29,30).
Quantitative translation from rabbit to human
Table 3 shows the result when the same method is applied to rabbit ventricular myocyte data from20. In this case, two doses (10 μM and 52 μM) of the IKr and ICaL blocker sotalol were employed. In addition20,21, provide measured data for human cardiomyocytes exposed to 30 μM sotalol. The column at the extreme right in Table 3 reports the increases in human APD50 and APD90 estimated based on the rabbit data. Both the estimated increase in APD50 (24.7%) and APD90 (28.8%) seem to be in good agreement with the measured human values from20,21 (32.3% and 20.6% for APD50 and 37.5% and 28.2% for APD90). The method estimates an IC50 value of 12 μM for block of IKr by sotalol and an IC50 value of 102 μM for block of ICaL. The corresponding IC50 values found in literature are 51–343 μM for IKr21,22,23,24 and 193 μM for ICaL23.
Discussion
The use of animal models to aid in understanding of human anatomy and physiology is centuries old, see e.g.31,32.
In the case of the electrophysiology of the vertebrate heart, this type of comparative study33 has led to deep insight into key physiological mechanisms, but it has been difficult to use these models quantitatively due to inherent species differences. More specifically, while animal cells are routinely used as part of drug development, their quantitative usefulness is hampered by the significant differences in human and animal action potential waveforms. At present, the most useful results are obtained by utilizing observed correlations between human and animal biomarkers (see e.g.34,35,36,37,38). However, no mechanistic relationship has, to date, been derived to examine how drug candidates may affect human and animal cardiomyocytes differentially. This limits the predictive insights into the therapeutic or toxic effects on humans. In this study, we have shown how an antiarrhythmic drug effect on human ventricular myocytes can be revealed solely by measuring an analogous effect of the drug on animal ventricular myocytes. This new investigative capability opens a wide variety of possibilities to improve the testing of novel compounds.
Initially, we have shown that canine data can be used to reliably predict the effect of the drug dofetilide on human myocytes as judged by changes in the common biomarkers APD50 and APD90. Similarly, data from zebrafish action potentials was used to estimate the effect of E-4031 on human ventricular myocytes. Finally, the effect of sotalol on rabbit cardiomyocytes was used to estimate the effect of the same drug on human ventricular myocytes, and the results were revealing.
Our method can only be used to estimate a selected drug effect on ion currents that have significant impact on both the animal and the human action potentials. If one current is very small in an animal but very large in a human, drug effects on that current cannot be estimated using the present technique. One such case is illustrated in Fig. 1 of1, where the action potential of human, canine, rabbit, guinea pig and mouse are illustrated together with associated currents. The substantial differences between the human and mouse action potentials and underlying ion currents suggest that it is difficult to use our method to predict the effect of drugs on human cardiomyocytes by measuring the effect of the drug on mouse cardiomyocytes.
In the present study, our principal aim was to generalize the methods we have developed previously12,13 for extrapolating drug effects obtained using hiPSC-CMs to healthy, mature human ventricular myocytes. Importantly, we found that the an equivalent approach can be used to translate measured pharmacological effects on the action potential in animal models to those in healthy adult human myocytes by employing the same models for the same currents in different cell types. The assumption underlying this approach is that major currents are carried by the same or similar ion channels for which isoforms differences have been measured and are relatively well understood. This is indeed often the case. While sarcolemmal voltage gated ion channels are complex transmembrane proteins of several classes, often composed of more than one interacting or modifying subunit, the alpha subunit of cardiac sodium, potassium, and calcium channels do show very strong primary sequence similarities across species; genome comparisons highlight a number of highly conserved channel families including Nav, Kv, and Cav39. For instance, in epicardial adult ventricular myocytes, channels underpinning late repolarization (delayed rectifier currents rapid and slow, IKr/hERG and IKs/KvLQT-minK, respectively) are conserved among human, canine, guinea pig, and rabbit cells8,40. Again, the transient outward current, Ito, prominent in early depolarization in the rabbit ventricle via Kv1.4/4.2/4.3, is similar in presentation and action potential effects in adult human myocytes, as carried by Kv4.39. The inward rectifying K+ current (IKl), the L-type Ca2+ current (ICaL), and the Na+/K+ pump current (INaK) are also present and effectual in rabbit10 in addition to other species, as in human. Rabbit measurements of calcium handling and excitation-contraction coupling have long been used as a basis to study and model these important phenomenon in human myocytes11, and the molecular constituents of all three major depolarizing sarcolemmal currents, ICaL, INa, and INaCa, are thought to be relatively consistent across species1. To say that conserved sequences implied conservation of overall structure or indeed ion channel function in all cases would be an oversimplification, as differences have been confirmed e.g.41,42. However, the relative conservation of expressed ion channel proteins across mammalian species provides a solid basis for application of our methodology for translation of drug effects between species, given an understanding of the divergence, as well as sufficient data for parameterization and validation.
Methods
Mathematical models of the membrane potential of excitable cells are written on the form
see e.g.43,44,45,46. Here, v is the membrane potential (in mV), t denotes time (in ms) and Ix are the membrane currents (in A/F). Each individual current can be written on the form
where A is the area of the cell membrane (in μm2) and Cm is the specific capacitance of the cell membrane (in pF/μm2); both A and Cm are common parameters for every currents in a given type of cardiomyocyte. Furthermore, Nx is the number of channels of type x, \({g}_{0}^{x}\) is the conductance of a single open channel (in nS), ox is the unitless open probability of the channel. Finally, Ex is the electrochemical equilibrium potential of the channel (in mV). By introducing the constant
we get the simpler expression
In (4) it is important to note that the characteristics of the dynamics of the specific ion channel under consideration is given by the open probability ox; the rest of the expression is either constants or the common term (v − Ex). The dynamics of ox can be modelled using a Markov scheme (see e.g.43,44,47). Here, it is critical to acknowledge that if the ion channel in two different species are identical, then so is the Markov model governing their dynamics. The diagram in Fig. 2 illustrates the assumptions underlying the translation from animal to human.

Upper left; illustration of an animal cardiomyocyte that express three types of ion-channels (proteins) in the myocyte membrane surface (sarcolemma). Two of the proteins are also present in the membrane of the human cardiomyocyte (upper right). Note that the human ventricular myocyte also express an additional type of protein not present in the animal cell. In the lower panels, both the animal and the human cardiomyocytes are subjected to a blocking drug that binds to one of the protein types. An essential assumption for allowing quantitative translation of animal measurements to estimates of the effect of human cells is that, at the level of a single protein, the effect of the drug is independent of whether the protein is expressed in human or an animal cell membrane.
The effect of a specific channel blocker is often expressed in terms of an IC50 value (see e.g.48; here we use the Hill coefficient 1). The IC50 value indicates the concentration that reduces the channel conductance by 50%. We assume that when a channel blocker is applied, the current takes the form
where we have defined the conductance of the single channel to be
In (6), we have introduced the parameter εx = 1/IC50; here D is the drug concentration given in the same unit as the IC50 (often μM). Clearly, \({I}_{x}({{\rm{IC}}}_{50})=\frac{1}{2}{I}_{x}(0),\) where Ix(0) is the current in baseline or control case. Note from (5) that we assume that the blocker only affects the maximum conductance of the single channel. This is a known oversimplification since the effects of drugs may be much more complex; see e.g.47,49,50. However, this assumption greatly simplifies the identification of the drug effect from measurements, and it is sufficient for our purposes.
IC50 for animal current = IC50 for human current
When a drug effect on an ion channel in an animal myocyte is detected, the key observation for translation to humans is that, given the assumption of functional invariance of channels between species (see Fig. 2), the effect of the drug on a single channel protein must be the same whether the protein is expressed in an animal or human. More specifically, if a current in an animal cardiomyocyte is given by
then the associated human current is given by
If a drug is added to the animal channel with a given IC50 value, then, as above, g0 of (7) is replaced by
where ε = 1/IC50. But since the conductance of the single channel is the same for the animal current and the human current, the single channel conductance of the human model (8) is also replaced by the expression given in (9). Hence, when a drug of concentration D is added we have the currents
Therefore, \({I}_{A}({{\rm{IC}}}_{50})=\frac{1}{2}{I}_{A}(0)\), and, similarly, \({I}_{H}({{\rm{IC}}}_{50})=\frac{1}{2}{I}_{H}(0)\), and thus the IC50 value of the animal and human current is the same. In principle, this key observation also is valid when the drug effect is modeled as a part of a complex Markov model, since the effect of the drug, on the single channel, again would be the same for animals and humans as long as the single ion channels are identical.
Details regarding the specific choice of action potential model used in our computations and the method used to identify drug effects on single ion channels from membrane potential measurements are given in the Supplementary information.
Supplementary Information
Translating from animal to human
Let us consider a very simple situation where we assume that both the animal myocyte action potential (a) and the human action potential (h) are generated by only two currents. The mathematical models takes then the form
where y is either a for animal or h for human. Here, clearly \({\rho }_{1}^{y}\), \({\rho }_{2}^{y}\) are different for y = a and y = h due to differences in the ion channel density. However, since the ion channels are the same, the Markov model governing the \({o}_{1}^{y}\), for y = a and y = h are the same.
Suppose now that we have measured an animal dataset sufficient to determine the coefficients ε1 and ε2. We immediately observe that we have a complete mathematical model for both the animal and the human action potentials. Using these principles, we can compute the biomarkers of interest for the human case.
Certainly, mathematical models used to represent action potentials are much more complex than the template model given by (12) in the main text; see e.g.11,51,52. In our computations, we use the Base model developed in13, as it systematically uses the same model for the identical ion channel in two different species. Note that in13 the results of the Base model is compared to the results of the Paci model (for immature cells) and to the Grandi et al. model and the ORd model (adult human cells).
Example
In order to explain and illustrate how it is possible to use animal data to gain quantitative insight concerning effects on human ventricular myocytes, we will present a very simple example. Let us for the sake of argument assume that an animal myocyte action potential is governed by
and the analogous human AP is governed by the very simple model,
Here g1, g2, g3, k1, k2, k3 are conductances and we assume that they are of significant magnitude. Suppose, for instance, that a drug is applied to the animal ventricular myocytes and inversion of the measured data suggests that the drug is a calcium blocker with the IC50 value given by 1/εCaL. Then the animal model in the setting when the drug has been applied is given by
where D denotes the concentration of the applied drug. Since the IC50 value of the single channel is the same for the animal and the human myocytes, the human model takes the form
This model can be used to compute chosen biomarkers of interest for the human action potential. In this particularly simple situation, we can conclude that:
- 1.
One can use the animal datasets to reveal drug effect on human INa and ICaL.
- 2.
One cannot use the animal to get any information about human IKr because the animal does not have IKr.
- 3.
The IKs current in the animal does not create difficulties for the measurements.
The base model of the action potential
In all of our simulations, we use the Base model derived in13. The parameters used to model human, dog, zebrafish, rabbit and guinea pig action potentials are given in Table 4. The remaining parameter values are specified in13, except that the intracellular potassium concentration is increased from 120 mM to 145 mM for the dog case in order to match the resting potential observed in the data from7. Note also that because zebrafish cardiomyocytes have been shown to exhibit a robust T-type calcium current (ICaT)17, the Base model from13, when utilized for the zebrafish simulations, is extended to include a T-type calcium current from53 of the form
where gCaT is the conductance of the channels and ECa is the Nernst equilibrium potential for calcium (see13). Here, d and f are gating variables governed by
Inversion of data
The inversion of data from dog, zebrafish and rabbit action potentials is performed using the inversion procedure described in13. In the inversion, the adjustment factors λKr, λCaL, λNa, λKs, λKl, λto, λNaL, λNaCa, λNaK, λbCl, λbCa, and λpCa for the currents IKr, ICaL, INa, IKs, IKl, Ito, INaL, INaCa, INaK, IbCl, IbCa, and IpCa, are treated as free parameters, in the zebrafish case, the adjustment factor λCaT for the current ICaT is also treated as a free parameter in the inversion, in addition to εKr (and εCaL for sotalol), representing the effect of the drug (see (6)).
Cost function definition
In the inversion, we minimize a cost function of the form
where d numbers the drug doses, Dd, included in the data set (including the control case), j numbers the different cost function terms, Hj, included in the cost function, and wd,j are specified weights for each of the cost function terms and each of the doses. The cost function includes the terms
where APD p is the action potential duration at p percent repolarization (see13). Furthermore, vmin and vmax are the minimum and maximum values of the membrane potential, respectively. The terms marked by an * represent values computed from measured data, and the remaining terms are computed from simulations of the model defined by λ and ε.
Cost function weight
We use the weight 2 for \({H}_{{v}_{{\rm{\max }}}}\), \({H}_{{v}_{{\rm{\min }}}}\) and HAPD50, the weight 5 for HAPD90 and the weight 1 for the remaining terms. In addition, the weights for the control case, w0,j, are multiplied by the total number of doses included in the data set (including the control case).
Minimization procedure
In order to minimize the cost function (14), we apply the continuation-based minimization procedure from13 using 10 iterations with 100 randomly chosen initial guesses. The initial guesses for λ are chosen within 10% above or below the optimal values from the previous iteration, and the initial guesses for εm are chosen within [εm−1/5, 5εm−1], where εm−1 is the optimal ε from the previous iteration. From these initial guesses we run 15 iterations of the Nelder-Mead algorithm54. The starting points for the first iteration of the continuation method are adjusted by hand.
Application of the method to generated data
In the main text, we have shown that we are able to use animal datasets (from dog, zebrafish and rabbit) to estimate the effect of IKr blockers on healthy human ventricular myocytes. However, available published data is limited and we therefore want to indicate that the methodology works more generally than we have been able to prove using available datasets. In Table 5, we have additionally used simulations to generate data. More specifically, we use the zebrafish model to estimate the effect for the human model of blocking the sodium current and the calcium current by 50%. In these inversions, λNa, λCaL and λKr and εNa, εCaL and εKr are treated as free parameters. In the generation of the zebrafish data, INa, ICaL and IKr are all increased by 20%. Furthermore, in order to detect changes in the sodium current, we extend the cost function to include the term
measuring differences in the upstroke velocity. The results in Table 5 illustrate that the method presented here has wide applicability, provided that the ion currents under consideration have significant impact on both animal and human action potentials.
Quantitative translation from guinea pig to zebrafish
The purpose of this paper is to show how a computational approach can be used to translate measured drug responses for animals to corresponding drug responses for humans. We have demonstrated how data from dogs, zebrafish and rabbits can be translated into predictions of human drug responses. In Table 6, we demonstrate how the same approach may be used to translate the effect of a drug from one animal to another. In this case, we consider data from18 of guinea pig cardiomyocytes exposed to four different doses of the IKr blocker E-4031. In addition, measured zebrafish data of 1 μM E-4031 are provided in17. Based on the measured guinea pig data, the mathematical method estimates an IC50 value of 27 nM for block of IKr by E-4031. This is in good agreement with values found in literature (10–397 nM25,26,27,28,29,30) as well as with the IC50 value of 44 nM identified based on zebrafish data in the main text. In Table 6, we observe that the method predicts a 41.7% increase in APD50 and a 35.2% increase in APD90 for zebrafish cardiomyocytes exposed to 1 μM E-4031, in line with the measured increases of 38.7% and 35.4% for APD50 and APD90, respectively.
References
Edwards, A. G. & Louch, W. E. Species-dependent mechanisms of cardiac arrhythmia: a cellular focus. Clinical Medicine Insights: Cardiology, 11, 1179546816686061 (2017).
Yoshida, Y. & Yamanaka, S. Induced pluripotent stem cells 10 years later. Circulation Research 120(12), 1958–1968 (2017).
Ye, L., Ni, X., Zhao, Z.-A., Lei, W. & Hu, S. The application of induced pluripotent stem cells in cardiac disease modeling and drug testing. Journal of Cardiovascular Translational Research 11(5), 366–374 (2018).
DiMasi, J. A., Grabowski, H. G. & Hansen, R. W. Innovation in the pharmaceutical industry: New estimates of R&D costs. Journal of Health Economics 47, 20–33 (2016).
Paul, S. M. et al. How to improve R&D productivity: the pharmaceutical industry's grand challenge. Nature Reviews Drug Discovery 9(3), 203–214 (2010).
Bockorny, M., Chakravarty, S., Schulman, P., Bockorny, B. & Bona, R. Severe heart failure after bortezomib treatment in a patient with multiple myeloma: A case report and review of the literature. Acta Haematologica 128(4), 244–247 (2012).
Jost, N. et al. Ionic mechanisms limiting cardiac repolarization reserve in humans compared to dogs. The Journal of Physiology 591(17), 4189–4206 (2013).
Zicha, S. et al. Molecular basis of species-specific expression of repolarizing K+ currents in the heart. American Journal of Physiology-Heart and Circulatory Physiology 285(4), H1641–H1649 (2003).
Wang, Z. et al. Potential molecular basis of different physiological properties of the transient outward K+ current in rabbit and human atrial myocytes. Circulation Research 84(5), 551–561 (1999).
Gemmell, P., Burrage, K., Rodriguez, B. & Quinn, T. A. Population of computational rabbit-specific ventricular action potential models for investigating sources of variability in cellular repolarisation. Plos One 9(2), e90112 (2014).
Grandi, E., Pasqualini, F. S. & Bers, D. M. A novel computational model of the human ventricular action potential and Ca transient. Journal of Molecular and Cellular Cardiology 48(1), 112–121 (2010).
Tveito, A. et al. Inversion and computational maturation of drug response using human stem cell derived cardiomyocytes in microphysiological systems. Scientific Reports 8(1), 17626 (2018).
Jaeger, K. H. et al. Improved computational identification of drug response using optical measurements of human stem cell derived cardiomyocytes in microphysiological systems. Frontiers in Pharmacology 10, 1648 (2020).
Niederer, S. A., Fink, M., Noble, D. & Smith, N. P. A meta-analysis of cardiacelectrophysiology computational models. Experimental Physiology, 94(5), 486–495, 5 (2009).
Groenendaal, W. et al. Cell-specific cardiac electrophysiology models. Plos computational biology 11(4), e1004242 (2015).
Jaeger, K. H., Wall, S. & Tveito, A. Detecting undetectables: Can conductances of action potential models be changed without appreciable change in the transmembrane potential? Chaos: An Interdisciplinary Journal of Nonlinear Science 29(7), 073102 (2019).
Nemtsas, P., Wettwer, E., Christ, T., Weidinger, G. & Ravens, U. Adult zebrafish heart as a model for human heart? An electrophysiological study. Journal of Molecular and Cellular Cardiology 48(1), 161–171 (2010).
Bussek, A. et al. Tissue slices from adult mammalian hearts as a model for pharmacological drug testing. Cellular Physiology and Biochemistry 24(5-6), 527–536 (2009).
Jost, N. et al. Restricting excessive cardiac action potential and QT prolongation: a vital role for IKs in human ventricular muscle. Circulation 112(10), 1392–1399 (2005).
Baczkó, I., Jost, N., Virág, L., Bösze, Z. & Varró, A. Rabbit models as tools for preclinical cardiac electrophysiological safety testing: importance of repolarization reserve. Progress in Biophysics and Molecular Biology 121(2), 157–168 (2016).
Orvos, P. et al. Evaluation of possible proarrhythmic potency: Comparison of the effect of dofetilide, cisapride, sotalol, terfenadine, and verapamil on hERG and native IKr currents and on cardiac action potential. Toxicological Sciences 168(2), 365–380 (2019).
Crumb, W. J., Vicente, J., Johannesen, L. & Strauss, D. G. An evaluation of 30 clinical drugs against the comprehensive in vitro proarrhythmia assay (CiPA) proposed ion channel panel. Journal of Pharmacological and Toxicological Methods, 81:251–262 Focused Issue on Safety Pharmacology (2016).
Kramer, J. et al. MICE models: superior to the HERG model in predicting Torsade de Pointes. Scientific Reports 3, 2100 (2013).
Qu, Y. & Vargas, H. M. Proarrhythmia risk assessment in human induced pluripotent stem cell-derived cardiomyocytes using the maestro MEA platform. Toxicological Sciences 147(1), 286–295 (2015).
Kim, K.-S. & Kim, E.-J. The phenothiazine drugs inhibit hERG potassium channels. Drug and Chemical Toxicology 28(3), 303–313 (2005).
Gibson, J. K., Yue, Y., Bronson, J., Palmer, C. & Numann, R. Human stem cell-derived cardiomyocytes detect drug-mediated changes in action potentials and ion currents. Journal of Pharmacological and Toxicological Methods 70(3), 255–267 (2014).
Katayama, Y. et al The inter-cell-line reproducibility of hERG assay using the whole-cell patch-clamping. Journal of Pharmacological Sciences, 97 (2005).
McPate, M. J., Duncan, R. S., Witchel, H. J. & Hancox, J. C. Disopyramide is an effective inhibitor of mutant HERG K+ channels involved in variant 1 short QT syndrome. Journal of Molecular and Cellular Cardiology 41(3), 563–566 (2006).
Piper, D. R. et al. Development of the predictor HERG uorescence polarization assay using a membrane protein enrichment approach. Assay and Drug Development Technologies 6(2), 213–223 (2008).
Sanguinetti, M. C. & Jurkiewicz, N. K. Two components of cardiac delayed rectifier K+ current. differential sensitivity to block by class III antiarrhythmic agents. The Journal of General Physiology 96(1), 195–215 (1990).
Shanks, N., Greek, R. & Greek, J. Are animal models predictive for humans? Philosophy, Ethics, and Humanities in Medicine 4(1), 2 (2009).
Hill, A. J. & Iaizzo, P. A. Comparative cardiac anatomy. In Handbook of cardiac anatomy, physiology, and devices, pages 89–114. Springer (2015).
Houser, S. R. et al. Animal models of heart failure: a scientific statement from the American Heart Association. Circulation Research 111(1), 131–150 (2012).
Gong, J. Q. X. & Sobie, E. A. Population-based mechanistic modeling allows for quantitative predictions of drug responses across cell types. NPJ Systems Biology and Applications 4(1), 11 (2018).
Bailey, J., Thew, M. & Balls, M. An analysis of the use of dogs in predicting human toxicology and drug safety. Alternatives to Laboratory Animals 41(5), 335–350 (2013).
Ando, K. et al. QT PRODACT: In vivo QT assay with a conscious monkey for assessment of the potential for drug-induced QT interval prolongation. Journal of pharmacological sciences 99(5), 487–500 (2005).
Bergenholm, L., Collins, T., Evans, N. D., Chappell, M. J. & Parkinson, J. PKPD modelling of PR and QRS intervals in conscious dogs using standard safety pharmacology data. Journal of Pharmacological and Toxicological Methods 79, 34–44 (2016).
Bergenholm, L. et al. Predicting QRS and PR interval prolongations in humans using nonclinical data. British Journal of Pharmacology 174(19), 3268–3283 (2017).
Jegla, T. J., Zmasek, C. M., Batalov, S. & Nayak, S. K. Evolution of the human ion channel set. Combinatorial Chemistry & High Throughput Screening (2009).
Jost, N. et al. A kesoi egyeniranyito kaliumaram gyors (IKr) es lassu komponensenek (IKs) osszehasonlito vizsgalata egeszseges emberi, kutya, nyul es tengerimalac kamrai szivizomsejteken. Cardiologia Hungarica 34, 103–113 (2004).
Blechschmidt, S., Haufe, V., Benndorf, K. & Zimmer, T. Voltage-gated Na+ channel transcript patterns in the mammalian heart are speciesdependent. Progress in Biophysics and Molecular Biology (2008).
Zimmer, T., Haufe, V. & Blechschmidt, S. Voltage-gated sodium channels in the mammalian heart. Global Cardiology Science and Practice (2014).
Rudy, Y. From genes and molecules to organs and organisms: Heart. Comprehensive Biophysics, pages 268–327 (2012).
Rudy, Y. & Silva, J. R. Computational biology in the study of cardiac ion channels and cell electrophysiology. Quarterly Reviews of Biophysics 39(01), 57–116 (2006).
Plonsey, R. & Barr, R. C. Bioelectricity, A Quantitative Approach. Springer (2007).
Sterratt, D., Graham, B., Gillies, A. & Willshaw, D. Principles of Computational Modelling in Neuroscience. Cambridge University Press (2011).
Tveito, A. & Lines, G. T. Computing Characterizations of Drugs for Ion Channels and Receptors Using Markov Models. Springer-Verlag, Lecture Notes, vol. 111 (2016).
Brennan, T., Fink, M. & Rodriguez, B. Multiscale modelling of drug-induced effects on cardiac electrophysiological activity. EuropeanJournal of Pharmaceutical Sciences 36(1), 62–77 (2009).
Clancy, C. E., Zhu, Z. I. & Rudy, Y. Pharmacogenetics and anti-arrhythmic drug therapy: A theoretical investigation. AJP: Heart and Circulatory Physiology 292(1), H66–H75 (2007).
Tveito, A., Maleckar, M. M. & Lines, G. T. Computing optimal properties of drugs using mathematical models of single channel dynamics. Computational and Mathematical. Biophysics 6(1), 41–64 (2018).
Kernik, D. C. et al. A computational model of inducedpluripotent stem-cell derived cardiomyocytes incorporating experimental variability from multiple data sources. The Journal of Physiology (2019).
O’Hara, T., Virág, L., Varró, A. & Rudy, Y. Simulation of the undiseasedhuman cardiac ventricular action potential: Model formulation and experimental validation. PLoS Computational Biology 7(5), e1002061 (2011).
Maltsev, V. A. & Lakatta, E. G. Synergism of coupled subsarcolemmalCa2+ clocks and sarcolemmal voltage clocks confers robust and exible pacemaker function in a novel pacemaker cell model. American Journal of Physiology-Heart and Circulatory Physiology 296(3), H594–H615 (2009).
Nelder, J. A. & Mead, R. A simplex method for function minimization. The Computer Journal 7(4), 308–313 (1965).
Carlsson, L. In vitro and in vivo models for testing arrhythmogenesis in drugs. Journal of Internal Medicine 259(1), 70–80 (2006).
Author information
Authors and Affiliations
Contributions
A.T. and K.H.J. did the mathematical modeling, performed numerical simulations, and wrote the paper; M.M.M., W.R.G. and S.W. reviewed the paper as a whole, and provided editing concerning relevance to existing literature on animal models (W.R.G.), relevance to drug development (S.W.), and applicability of the mathematical model (M.M.M.).
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tveito, A., Jæger, K.H., Maleckar, M.M. et al. Computational translation of drug effects from animal experiments to human ventricular myocytes. Sci Rep 10, 10537 (2020). https://doi.org/10.1038/s41598-020-66910-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-66910-0
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
