
Abstract
Some liquid plant exudates (e.g. resin) can be found preserved in the fossil record. However, due to their high solubility, gums have been assumed to dissolve before fossilisation. The visual appearance of gums (water-soluble polysaccharides) is so similar to other plant exudates, particularly resin, that chemical testing is essential to differentiate them. Remarkably, Welwitschiophyllum leaves from Early Cretaceous, Brazil provide the first chemical confirmation of a preserved gum. This is despite the leaves being exposed to water twice during formation and subsequent weathering of the Crato Formation. The Welwitschiophyllum plant shares the presence of gum ducts inside leaves with its presumed extant relative the gnetalean Welwitschia. This fossil gum presents a chemical signature remarkably similar to the gum in extant Welwitschia and is distinct from those of fossil resins. We show for the first time that a water-soluble plant exudate has been preserved in the fossil record, potentially allowing us to recognise further biomolecules thought to be lost during the fossilisation process.
Similar content being viewed by others

Introduction
A wide variety of vascular plants produce fluid exudates1 e.g. resins and gums, with each group differing in chemical definitions (Table 1). Due to similarity in physical appearance distinguishing exudates based on chemistry is vital, for example gums and resins are visually similar resulting in these terms being used interchangeably1. However, their chemical definitions are very different (Table 1); resins are composed of lipid-soluble terpenoids1,2, while gums are complex, highly branched (non-starch) water-soluble polysaccharides3. A common example of this misunderstanding is the Eucalyptus tree, which is known as a gum tree, but nuclear magnetic resonance analysis of the Eucalyptus exudate shows its composition to be polyphenolic and is therefore actually a kino4 (Table 1). Differences between gum and resin can also be seen in the functional roles within the plant. The main roles of resins are to respond to wounding, as a defence against pathogens and to dissuade herbivory by insects and other organisms2,5,6. Gum is involved in food storage, structural support, and also for wound sealing, but there is no common role across taxa3. Further confusion arises as some plants, e.g. Boswellia and Commiphora species, even produce exudates with a mixture of polysaccharide and resin components (the gum resins myrrh and frankincense respectively)1.
Until now only fossilised plant resin (ambers)7 and latex filaments have been reported preserved in the fossil record8,9. While the fossilisation of fluid exudates might seem unlikely, the fossilisation of resin is relatively common, and extends back some 320 million years to the Carboniferous10, but chemically confirmed gums have never been reported.
The Early Cretaceous (~120 million year old) Crato Formation11 of northeast Brazil (Supplementary Fig. S1) is a well-known laminated limestone deposit that yields exceptionally preserved vertebrates, arthropods, and plants from the Nova Olinda Member12. Investigations of different groups of fossilized animals from the Crato Formation show that they are preserved as various mineral replacements, and their preservation was microbially-mediated13,14,15,16,17.
Amber has also previously been reported from the Crato Formation Lagerstätte, inside fossil plant remains and as isolated clasts18,19,20, and is attributed to conifers18,19. The fossil leaves occur as compressions showing at least some three-dimensionality (Supplementary Fig. S2). An amber-coloured substance is visible in some of the fossil leaves of Welwitschiophyllum brasiliense Dilcher et al. 2005 from the Crato Formation.
Welwitschiophyllum is considered a relative of the extant gymnosperm Welwitschia mirabilis Hooker 1863, the sole member of this gnetalean genus21. The Welwitschiaceae have a sparse macrofossil record, fossils assigned to this family, including Welwitschiophyllum, derive solely from the Crato Formation21. However, the pollen record shows Welwitschiaceae were once a diverse and prevalent group22 that saw a decline with increasing angiosperm pollen23. Today, Welwitschia is restricted to the Namib Desert in Namibia and Southern Angola and has chemically confirmed gum in both the cone and in abaxial ducts within leaves24,25.
We investigated this amber-coloured substance inside fossil Welwitschiophyllum leaves to test whether Welwitschiophyllum produced a resin (now fossilised as amber), or a gum like its presumed extant relative Welwitschia, using Fourier-transform infrared spectroscopy (FTIR) and Attenuated total reflectance (ATR) spectroscopy. We report here the first geochemical evidence for fossilised gum preserved inside Welwitschiophyllum leaves, and suggest areas for future investigation to understand how a ~120 million year old gum may have survived.
Results
Welwitschiophyllum leaves
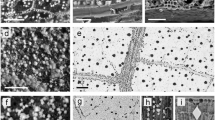
Fossils of Welwitschiophyllum occur as long detached leaves up to 850 mm in length with thin bands of an amber-like substance. (Fig. 1a and Supplementary Fig. S2a). These are particularly visible where the fossil surface has been abraded or removed (Fig. 1b). This substance in the Welwitschiophyllum leaves resembles amber in ducts, lying parallel to the long axis of the leaves (Fig. 1b). These ducts are inferred here as adaxial (upper leaf surface) due to the curvature of the leaf base. However, the absence of preserved cuticle and other anatomical features, means that their precise orientation cannot be confirmed. This constituent arrangement contrasts with the traumatic formation of gum in its presumed relative Welwitschia. Slight compaction of the specimens gives these ducts an ellipsoidal cross section, (Fig. 1c and Supplementary Fig. S3) but they appear to have a repeating pattern showing a principal duct followed by a secondary duct ranging in diameter from 75 µm to 200 µm (Fig. 1d).

Fossil Welwitschiophyllum leaves with gum ducts from the Crato Formation, Brazil. (a) Complete elongate fossil leaf (UERJ 13-P1) showing a curved base and degraded fibrous apex, with a partially abraded surface (lighter part of fossil) exposing the internal leaf tissue and linear gum duct arrangement. (b) Detail from (a) (UERJ 13-P1) where the gum ducts appear as amber-brown structures within Welwitschiophyllum leaf tissue. (c) Transverse thin section through the fossil leaf (UERJ 14-P1) with arrowheads indicating the orange coloured gum ducts within the brown leaf tissue. The black line of tissue may be compressed remains of vascular tissue (below the leaf is the preserving sediment). (d) An oblique thin section of the leaf (UERJ 14-P1) showing the repeating pattern of the amber-coloured gum ducts. Scale bars, (a) 20 mm (b) 3 mm, (c) and (d) 500 μm.
Spectroscopy
Analysis using FTIR and ATR is commonly used on both living and fossil plants showing that complex biomolecules survive and are identifiable in the fossil record26,27,28. FTIR analyses compare living and fossil resin and gum samples (Supplementary Fig. S4). Additionally, ATR analysis confirms that the amber-coloured substance in the fossil leaves, which was extracted and purified for testing, generated a spectrum closely matching those of published gum signatures26 and is remarkably similar to that of Welwitschia gum (Fig. 2a). The ‘noise’ seen in the preserved gum spectrum (spectral line ‘waviness’ over the broader signal detected) was generated possibly due to the very small amount of material available for analysis. It is unlikely but also possibly due to the laser power and low accumulation numbers, but 32 accumulations were made per sample and no other sample showed this feature. Despite the noise, the key features of the spectrum are clearly visible. The diagnostic features of gums are a very large hydroxyl peak at ~3400 cm−1, peak absence at 1516 cm−1 and a very strong peak at 1077 cm−1 26. Using ATR these peaks can be seen both in the fossil Welwitschiophyllum and the extant Welwitschia with the peak at 1077 cm−1 appearing as a shoulder on a strong O-H stretch peak in Welwitschiophyllum (Fig. 2).

ATR spectra of the fossil gum extracted from Welwitschiophyllum (UERJ 13-P1) leaf, and gum from extant Welwitschia. (a) Overview of the ATR spectra with chemical bonds indicated. (b) Detail of the fingerprint regions of gums from both Welwitschiophyllum and Welwitschia with chemical bonds indicated.
Gum solubility
The discovery of in situ preserved plant gum is unusual because of its solubility in water. This is particularly striking in a formation thought to be deposited in a hypersaline lagoon setting. Solubility experiments were undertaken on Welwitschia gum to determine whether the increased salinity of the lagoon may have affected the solubility of the gum in any way (Supplementary Table S1). In the freshwater, brackish, normal marine, and hypersaline water tests the extracted gum dissolved within 49–59 minutes, showing that salinity does not affect solubility, and therefore the preservation (or not) of exposed gum.
Discussion
Our analyses of the amber-coloured substance inside the fossil Welwitschiophyllum leaves shows a distinct chemical spectrum that clearly differs from those of ambers and resins17,18 (compare Supplementary Fig. S4 Brachyphyllum amber), but which closely compares to plant gum spectra, including our comparison sample (see Fig. 2) and those found in the literature26. This means that the recovered substance from the Crato Formation fossil Welwitschiophyllum leaves is a preserved gum and not an amber formed from resin. The chemically detected presence of gum in ducts inside two separate fossil leaves (Fig. 1 and Supplementary Fig. S3) confirms that this is not an isolated occurrence within these Crato Formation fossils.
Due to the soluble nature of gum, its preservation in the fossil record is unexpected. This is particularly notable here as the leaves containing gum were firstly deposited in a hypersaline lagoon, then later this deposit was exposed to continental weathering18. Thus, water featured in both the formation and weathering of the Crato Formation, yet the gum persisted. The gum solubility experiments showed that in each case of differing salinities the extracted Welwitschia gum dissolved, so saline levels appear to have no bearing on gum preservation.
How the gum came to be preserved is currently not understood and further investigation is needed into the taphonomic and diagenetic processes surrounding these gum-preserving fossil leaves. We can speculate that there are at least two factors involved. Firstly, the nature of the microbially-mediated taphonomy and diagenesis in the Crato palaeo-lake setting has been shown to be critical in the preservation of labile structures in animals from the Crato Formation Konservat-Lagerstätte14,15,16,17. Secondly, perhaps only in part, the coriaceous nature of the fossil leaves played a role. Both the surrounding duct tissue and the large amount of resistant embedding leaf tissues would have provided some protection from dissolution in water. In extant Welwitschia the outer walls of the epidermal cells are specialised with three layers, thickening and strengthening the epidermis25, but the preservation of the fossil leaves prohibits epidermal comparison. The regular arrangement of ducts in Welwitschiophyllum suggests that they were formed through duct initiation29, i.e. constituent, as opposed to the stress initiated response known as gummosis30. Their formation was likely to be used for food storage or structural support, signifying that the hydrophilic gum was constituent within the fossil leaves.
Despite the very low preservation potential of a highly water-soluble exudate, the first preserved gum was recovered from the Early Cretaceous. This fossil gum presents a chemical signature remarkably similar to gum in extant Welwitschia and distinct from those of fossil resins. This shows that gum production in plants extends back into the fossil record by at least ~120 million years. This is then the first report of a highly soluble biomolecule recovered from the Crato Formation and future work should focus on how this preserved gum survived. Furthermore, fossilised plants with observed internal ‘resins’ should be chemically confirmed in case further instances of gums or other types of plant exudate can be identified from the fossil record.
Materials and Methods
Image processing
Macro images were taken using Canon EOS 1100D. Images were made into combined figures using Inkscape.
Petrographic thin sections
To examine the fossil plant histology petrographic thin sections were made using standard procedures and examined using a Leica DM750P microscope.
Fossil material extraction and comparison samples
The fossil amber-coloured material for both FTIR and ATR analysis (from Specimen Welwitschiophyllum UERJ 13-P1 and Brachyphyllum UERJ15-P1) was mechanically extracted from the fossil leaf remains using sterile scalpel blades and dental picks under a Leica EZ4W stereomicroscope. The extracted samples were washed in absolute alcohol to minimise contamination. A sample of the limestone matrix from the fossil leaves was also mechanically extracted using sterile scalpel blades and prepared for comparison. All these fossil material samples were then ground into a fine powder using a pre-autoclaved and pre-sterilised glass micro-mortar and pestle (new one for each sample to avoid cross contamination). The resulting powder was then further checked microscopically for any visible impurities, and none were seen.
For comparative purposes analyses were performed on recent exudate from the extant gnetalean Welwitschia mirabilis Hooker, 1863 from the gardens of the South African National Biodiversity Institute, Pretoria (SANBI), and a commercial sample of sandarac was obtained. Reference samples (of sandarac) were washed in isopropanol. These samples were then ground into a fine powder using a pre-autoclaved and pre-sterilised glass micro-mortar and pestle (new one for each sample to avoid cross contamination). These separately prepared powdered modern and fossil exudates were then ready for the spectral analyses.
Spectral analyses
Analysis of gums and resins was performed using FTIR spectroscopy on a PerkinElmer ‘Spectrum 400’ spectrometer, fitted with an ATR sampling accessory (range 4000–550 cm−1, 32 accumulations, 4 cm−1 resolution). KBr pelletisation was not necessary for this machine setup and the pre-powdered samples were applied to the measurement area of the spectrometer with a new and autoclaved micro-spatula (one per measurement). The machine was cleaned by vacuuming off the measured samples thoroughly, then wiping with pure ethanol until all traces were removed and allowed to dry between measurements (test spectra were made to ensure no cross contamination occurred). Sample UERJ 13-P1 (amber-coloured fossil substance from Welwitschiophyllum) was averaged from four scans (with 32 accumulations per sample), the other samples (Welwitschia, Brachyphyllum, sandarac and limestone) had only 1 scan (with 32 accumulations per sample), and the peaks in all cases were normalised.
To confirm the results from the averaged multiple FTIR scans and provide more sensitive data, selected samples were further assessed by hot aqueous extraction and ATR. For ATR analyses (on Welwitschia and Welwitschiophyllum sample (UERJ 13-P1) a small amount of the sample was placed in 0.5 ml of water, which was heated to approximately 90 °C for 10 minutes. A drop of the residual liquid was placed on the ATR crystal, and the water allowed to evaporate, leaving a film of the extracted material on the crystal surface. A spectrum was recorded (with 32 accumulations, to improve the quality of the data, due to the very small amount of material under analysis) and peaks were normalised. Care was taken to thoroughly clean the ATR crystal between sample measurements using warm water and pure ethanol and allowed to dry. Test spectra were made to ensure no cross contamination was occurring between the measurement scans.
A baseline correction for both analyses with reference points at 3715 and 1800 cm−1 was performed. In all cases, the spectrographs were visualised using Spectra Gryph 1.2.10 software.
Solubility experiments
These were completed using a Gallen Kamp magnetic stirrer regulator set at speed 4, Welwitschia gum was placed in freshwater (water-62.5 ml, gum weight-0.08 g), brackish (water- 62.5 ml,1.25 ppt, gum weight-0.08 g), normal marine (water-62.5 ml, 2.18 ppt, gum weight-0.08 g) and hypersaline (water-62.5 ml, 3.12 ppt, gum weight-0.08 g) water.
CITES permit
Analysis of extant Welwitschia mirabilis gum was performed on samples obtained on CITES permit No. 152606.
Data availability
The fossil material from the Crato Formation examined here comprises three isolated leaves of Welwitschiophyllum brasiliense Dilcher et al. 2005 (Specimens UERJ 13-P1, UERJ 14-P1, and UOP-PAL-MC0002) and one specimen of Brachyphyllum obesum Heer, 1875. (UERJ 15-P1). Specimens UERJ 13-P1, UERJ 14-P1 and UERJ 15-P1 are accessioned at Rio Janeiro State University and UOP-PAL-MC0002 is accessioned at the University of Portsmouth. All data generated or analysed during this study are included in this published article (and its Supplementary Information files).
References
Langenheim, J. H. Plant Resins: Chemistry, Evolution, Ecology, and Ethnobotany. Timber Press, Portland, Cambridge (2003).
Langenheim, J. H. Higher plant terpenoids: a phytocentric overview of their ecological roles. Journal of Chemical Ecology 20, 1223–1280 (1994).
BeMiller, J. N. Plant gums. Essential for Life Science John Wiley and Sons Ltd, Chichester 1–6 (2014).
Lambert, J. B., Heckenbach, E. A., Wu, Y. & Santiago-Blay, J. A. Characterization of plant exudates by principal-component and cluster analyses with nuclear magnetic resonance variables. Journal of Natural Products 72, 1028–1035 (2010).
Henwood, A. Recent plant resins and the taphonomy of organisms in amber: a review. Modern Geology 19, 35–59 (1993).
Langenheim, J. H. Biology of amber-producing trees: focus on case studies of Hymenaea and Agathis. In: Anderson, K. B. & Crelling, J. C. (Eds.), Amber, Resinite and Fossil Resin. American Chemical Society, Washington D. C., 1–31 (1995).
Lambert, J. B., Santiago-Blay, J. A. & Anderson, K. B. Chemical signatures of fossilized resins and recent plant exudates. Angewandte Chemie International Edition 47, 9608–9616 (2008).
Simoneit, B. R. T., Otto, A. & Wilde, V. Novel phenolic biomarker triterpenoids of fossil laticifers in Eocene brown coal from Geiseltal, Germany. Organic Geochemistry 34, 121–129 (2003).
Mahlberg, P. G., Field, D. W. & Frye, J. S. Fossil Laticifers from Eocene Brown Coal Deposits of the Geiseltal. American Journal of Botany 71, 1192–1200 (1984).
Bray, P. S. & Anderson, K. B. Identification of Carboniferous (320 million years old) class 1c amber. Science 326(Issue 5949), 132–134 (2009).
Batten, D. J. Spores and pollen from the Crato Formation: biostratigraphic and palaeoenvironmental implications. In: Martill, D. M., Bechly, G. & Loveridge, R. F. (Eds.), The Crato Fossil Beds of Brazil–Window into an Ancient World. Cambridge University Press, Cambridge, 566–573 (2007).
Martill, D. M., Bechly, G. & Loveridge, R. The Crato Fossil Beds of Brazil–Window into an Ancient World. Cambridge University Press, Cambridge (2007).
Barling, N., Martill, D. M., Heads, S. W. & Gallien, F. High fidelity preservation of fossil insects from the Crato Formation (Lower Cretaceous) of Brazil. Cretaceous Research 52, 605–622 (2015).
Varejão, F. G. et al. Exceptional preservation of soft tissues by microbial entombment: insights into the taphonomy of the Crato Konservat-Lagerstätte. Palaios 34, 331–348 (2019).
Osés, G. L. et al. Deciphering the preservation of fossil insects: a case study from the Crato Member, Early Cretaceous of Brazil. PeerJ 4, e2756 (2016).
Osés, G. L. et al. Deciphering pyritization-kerogenization gradient for soft-tissue preservation fish. Scientific Reports 7, 1468 (2017).
Dias, J. J. & Carvalho, I. S. Remarkable fossil crickets preservation from Crato Formation (Aptian, Araripe Basin), Lagerstätten from Brazil. Journal of South American Earth Sciences 98, 102443 (2019).
Martill, D. M., Loveridge, R. F. L., Andrade, J. A. F. G. & Cardoso, A. H. An unusual occurrence of amber in laminated limestones: the Crato Formation lagerstätte (Early Cretaceous) of Brazil. Palaeontology 48, 1399–1408 (2005).
Pereira, R., Carvalho, Id. S., Simoneit, B. R. T. & Azevedo, D. A. Molecular composition and chemosystematic aspects of Cretaceous amber from the Amazonas, Araripe and Recôncavo basins, Brazil. Organic Geochemistry 40, 863–875 (2009).
Batista, M. E. P. et al. New data on the stem and leaf anatomy of two conifers from the Lower Cretaceous of the Araripe Basin, northeastern Brazil, and their taxonomic and paleoecological implications. PLOS One 12, 1–18 (2017).
Dilcher, D. L., Bernardes de Oliveira, M. E., Pons, D. & Lott, T. A. Welwitschiaceae from the Lower Cretaceous of Northeastern Brazil. American Journal of Botany 92, 1294–1310 (2005).
Crane, P. R. The fossil history of the Gnetales. International Journal of Plant Science 157, 50–57 (1996).
Crane, P. R. & Lidgard, S. H. Cretaceous patterns of palynological diversity and the angiosperm radiation. In: Taylor, P. D & Larwood, G. P. (Eds), Major Evolutionary Radiations. Oxford University Press, UK, 377–407 (1990).
Rodin, R. J. Leaf anatomy of Welwitschia. II. A study of mature leaves. American Journal of Botany 45, 96–103 (1958).
Sykes, M. G. V. The anatomy and morphology of the leaves and inflorescences of Welwitschia mirabilis. Philosophical Transactions of the Royal Society B 201, 179–22 (1911).
Tappert, R., Wolfe, A. P., McKellar, R. C., Tappert, M. C. & Muehlenbachs, K. Characterizing modern and fossil gymnosperm exudates using micro-Fourier transform infrared spectroscopy. International Journal of Plant Science 172, 120–138 (2011).
Seyfullah, L. J., Sadowski, E. M. & Schmidt, A. R. Species-level determination of closely related araucarian resins using FTIR spectroscopy and its implications for the provenance for New Zealand amber. PeerJ 3, e1067 (2015).
Vajda, V. et al. Molecular signatures of fossil leaves provide unexpected new evidence for extinct plant relationships. Nature Ecology and Evolution 1, 1093–1099 (2017).
Venkaiah, K. Development, Ultrastructure and Secretion of Gum Ducts in Lannea coromandelica (Houtt.) Merrill (Anacardiaceae). Annals of Botany 69, 449–457 (1992).
Fahn, A. Secretory tissues in plants. Academic Press (1979).
Acknowledgements
We would like to warmly thank the three anonymous reviewers whose comments strengthened the paper. Many thanks to: Dr. Anthony Hitchcock, Karin Behr and Benjamin Festus for assistance obtaining permits and sending the Welwitschia samples (SANBI, Pretoria, SA), Geoff Long (Portsmouth) for thin section production, Dr. Artur Andrade (DNPM Crato) and Dr. Paulo Brito (UERJ Rio de Janeiro) for assistance during fieldwork in Brazil, Professor Mark Jones at The Mary Rose Trust for the use of their micro-FTIR for preliminary analyses. Thanks to Dr Holly Turner for reading through the manuscript. Support for LJS provided by the German Research Foundation (DFG), project number SE2335/3-1.
Author information
Authors and Affiliations
Contributions
D.M.M. and R.F.L. devised the project, D.M.M. and R.F.L. undertook the fieldwork in Brazil, E.A.R., R.F.L. and L.J.S. prepared the material, E.A.R., R.F.L., L.J.S., P.G. ran the analyses. Interpretation was by E.A.R., L.J.S., R.F.L. and P.G. The paper was written by E.A.R., L.J.S., R.F.L., D.M.M. and P.G.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Roberts, E.A., Seyfullah, L.J., Loveridge, R.F. et al. Cretaceous gnetalean yields first preserved plant gum. Sci Rep 10, 3401 (2020). https://doi.org/10.1038/s41598-020-60211-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-60211-2
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
