
Abstract
The Late Triassic saw a flourish of plant–arthropod interactions. By the Late Triassic, insects had developed all distinct strategies of herbivory, notably including some of the earliest occurrences of leaf-mining. Herein we describe exceptionally well-preserved leaf-mine trace fossils on a Cladophlebis Brongniart fern pinnule from the Momonoki Formation, Mine Group, Japan (Middle Carnian), representing the oldest unequivocal leaf-mines from East Asia. The mines all display a distinctive frass trail—a continuous meandering line, which later becomes a broad band containing spheroidal particles—demonstrating larval development. Although the shapes of the frass trails are generally comparable to those of Lepidoptera or Coleoptera, they cannot be unequivocally assigned to a specific extant leaf-mining taxon. Furthermore, elemental analyses by X-ray fluorescence (XRF) reveals that the frass trail comprises phosphate coprolites. The quantitative variations in P, S, and Si between coprolites and leaf veins may reflect physiological processes (e.g., consumption, absorption, and excretion) mediated by plant chemicals. Our findings reinforce the idea that leaf-mining had become a pervasive feeding strategy of herbivorous insects by the Late Triassic.
Similar content being viewed by others


Introduction
Leaf mining is a means of herbivory by which insects consume live foliage while dwelling inside the host-plant tissue1. The biology of leaf-mining has historically attracted much attention and is extensively studied by ecologists2. Extant leaf miners can be identified between species and subfamily level based on mine shape and host-plant taxonomy. Thus plant–insect associations can be reconstructed by analysing mined leaves. Mined leaves also provide information on much of the overall life history of an individual miner: namely, the developmental process, from oviposition, through larval growth and the pupal stage, to adult emergence, is externally traceable, which makes it an ideal system for the study of demography and population dynamics while modelling the rates and causes of mortality (e.g., parasitism, competition) in natural populations3,4. As leaf-mining insects afford a wealth of ecological information, they have frequently been used for studying population, community, and evolutionary ecology of plant–insect interactions5,6,7.
The evolution of leaf-mining is a compelling aspect of the development of dietary niches of insects. Insect groups with mining habits have evolved multiple times in several holometabolous insect orders (e.g., Lepidoptera, Coleoptera, Diptera, Hymenoptera)1,8. Miners are more susceptible to the chemical defences of host plants and are more severely attacked by parasitoids than external feeders9. Thus, they tend to show high host specificity, with many species being monophagous or oligophagous herbivores2,10, and they are embedded in a complex multitrophic network11,12,13. Some leaf-mining insect groups have been used for assessing patterns and processes of macroevolution within a phylogenetic framework14,15,16.
Leaf-mining has also been examined in palaeobiological studies because mine trails can be found from impression/compression fossils of plant leaves. In some cases, the leaf miner responsible for the mine is assignable to a specific insect group by comparing the shape of the fossil mine and the host-plant taxonomy with analogous extant ones17,18,19,20,21; these records can help to time-calibrate insect phylogeny. Furthermore, the specialized damage observed in fossil leaf mines enables us to answer various questions related to dynamic shifts or persistence of plant–insect interactions before and after ecological perturbations22,23,24,25.
Concerning the evolutionary origin of leaf-mining, however, available information is scarce. The earliest credible occurrence of leaf-mining so far dates back to the Middle–Late Triassic, which is much later than other functional feeding groups: namely, sporangivory, boring, external foliage feeding, seed predation, piercing-and-sucking, and galling26,27. Substantial evidence of leaf-mining is, however, very scarce in records preceding the Late Triassic.
Herein we describe a novel type of leaf-mine fossil from the Momonoki Formation, southwestern Japan. Described below are the exquisitely preserved mines in a Cladophlebis fern frond, and the traces preserve their organic remains. The specimen has already been displayed for years in a cabinet at the Mine City Museum of History and Folklore, Yamaguchi, Japan. The fossil locality is dated to be the Middle Carnian (ca. 220 Ma)28, and thus, the mines represent one of the oldest credible leaf-mine fossils worldwide. Additionally, elemental analyses using an X-ray fluorescence (XRF) spectrometer are conducted on the fossil mines, casting new light on this ancient plant–herbivore association from the perspective of ecological stoichiometry.
Materials
Geological setting
The Mine Group, which faces continental China, is located on the northern side of the Median Tectonic Line, which divides the southwest end of Honshu island, Japan (Fig. 1). The stratigraphic setting is subdivided into three strata: the Hirabara, Momonoki, and Aso Formations, in ascending order29,30. Study material was obtained from the Momonoki Formation, which yields the most abundant fossil plants and insects31,32. The Momonoki Formation is dated to the Middle Carnian (ca. 220 Ma) based on U–Pb age data of detrital zircon28, which is consistent with the biostratigraphy of marine and terrestrial invertebrates29,33. The Momonoki Formation exceeds 1000 m in thickness and is conformable with the subjacent Hirabara Formation28,32. The sandstones and muddy sandstones of the Momonoki Formation host impressions of plant and insect macrofossils. The specimen was collected by H.T. at a road cutting on National Route 435 while the road was under construction; the locality also bore many insect fossils32,34,35,36. The Momonoki Formation is a nonmarine deposit characterized by lacustrine, deltaic deposit without marine invertebrate fossils, unlike in the Hirabara and Aso Formations31,37.
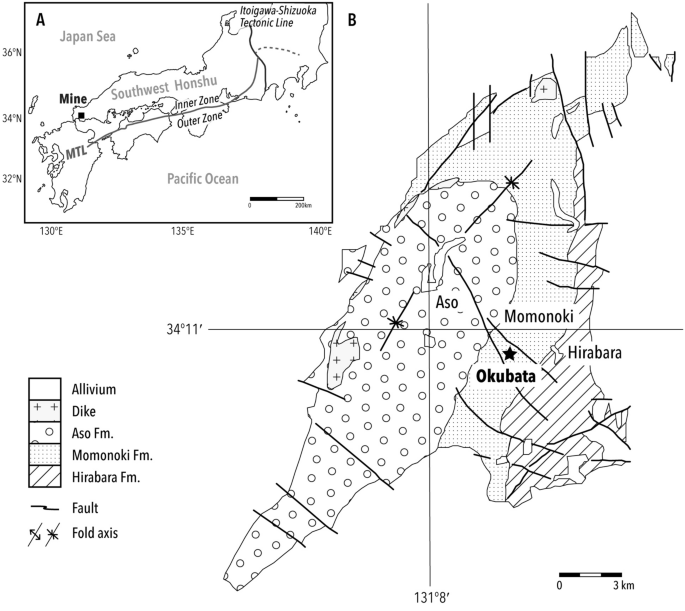
Study site location and geological map. (A) Map showing the fossil site, Mine, Yamaguchi, in southwest Honshu, Japan. Mine is located in the Inner Zone, the region formed by complex faulting along the Median Tectonic Line (MTL) and separated from the Outer Zone (southern side). (B) Geological map of the Mine Group, slightly modified from previous studies38,39. The star denotes the fossil locality, Okubata.
Palaeobiological setting
Plant remains from this locality are primarily represented by impressions of foliage, stem fragments with leaves, and, occasionally, fructifications. The plant assemblage of the Momonoki Formation is composed of ferns, sphenophytes, cycadophytes, ginkgophytes, and conifers40. There are three characteristics of the Momonoki Formation that are distinct from some other coeval fossil plant assemblages41: Sphenopsida (e.g., Neocalamites Halle and Equisetites Sternberg), ferns (e.g., Camptopteris Presl, Clathropteris Brongniart, Dictyophyllum Lindley et Hutton, Cladophlebis), and conifers (e.g., Podozamites Braun, Cycadocarpidium Nathorst) are well represented; cycadophytes and ginkgophytes42 are common; Danaeopsis Schimper and Symopteris Xu (Marattiaceae) ferns are lacking. Liverworts (Pallaviciniites Schuster) are also present43. The floral composition of the Momonoki Formation is part of the Southern Floristic Region of East Asia41,44; the floristic region extends over a broad geographic range, from as far south as Indonesia and north to a small area of northeastern China, and is characterized by the occurrence of Dictyophyllum and Clathropteris41.
A diverse fossil insect fauna has been found at the Momonoki Formation36,45. These fossils are borne from specific stratigraphic layers of dark, fine-grained siltstones that are devoid of coarse detritus31. Most of the insect fossils are isolated wings, but nearly complete bodies are also found36. The recorded insects encompass at least 12 orders: Odonata, Ephemeroptera, Blattodea, Plecoptera, Paraplecoptera, Reculida, Neuroptera, Hymenoptera, Coleoptera, Hemiptera, Mecoptera, Diptera32. In particular, there are rich yields of wings of Coleoptera (Taldycupedidae) and Blattodea (Mancusoblattidae, Mesoblattinidae) amongst the insect fauna of this deposit32,34. Notably, the Momonoki Formation yields true flies36, albeit undescribed, which have only been recorded from a few deposits from the Middle–Late Triassic46,47,48,49,50,51. Although more than 6000 specimens of fossil insects are recorded from this formation30,36, the taxonomic diversity is largely unexplored, and numerous terrestrial and aquatic insect specimens remain undescribed. At present, 18 species in ten families belonging to seven orders are described32,33,34,35,45. The insect fauna of the Mine Group is thought to have an affinity with four remote localities32,33: the Mount Crosby Formation in Australia, the Madygen Formation in Kyrgyzstan, the Djam Djun Formation in Vietnam, and the Yan-Chen Formation in China.
Results
Description of trace fossil
Three leaf mines with similar structures are found on five consecutive pinnules of the basiscopic pinna (white arrowheads in Fig. 2A,B). The plant tissues, including the epidermis and veins, stand out sharply against the profoundly weathered matrix of the slab. The mines broadly cover the lamina, although the margins of mined and remaining areas are not clearly distinguished as breached epidermal tissue, indicating that the miners left some tissue layers unconsumed. The mines originate either from the proximal or distal end of the lobes. Starting points (i.e., oviposition site) are obscure or unseen (orange arrowheads in Fig. 2C,D), lying on a secondary leaf vein. Frass trails appear as narrow sinusoidal threads with a width of 0.5–1.3 mm (1.1 mm on average), accompanying many close-set hairpin curves with occasional loops. The frass trails subsequently expand into broad bands of width 0.5–0.8 mm, with the dense accumulation of faecal material; individual frass pellets are hardly discernible. The frass trails are single, continuous lines but accompanied with a few gaps; altogether, each frass trail covers nearly the entire distal part of a pinnula. Although the trajectories of the frass trails freely extend across the secondary veins, they do not traverse the first veins (i.e., midribs) except those near the leaf apex; such behavioural characteristics thus give the entire mining structure on a pinnule a U-shape. These mines connect to the neighbouring pinnule through the widened base of the pinnule (Fig. 2D–E). The fact that the leaf veins within the mined area are not distorted dictates that the vascular bundles are left unconsumed. The darkened tissue at the end of the mine indicates a possible pupal chamber (pc, Fig. 2D).

Late Triassic leaf-mine fossil on Cladophlebis denticulata of the Momonoki Formation, Yamaguchi, Japan. (A) Specimen (MMHF11-00001a), on which mines on pinnules and chewing marks are marked with white arrowheads and yellow asterisks, respectively, and (B) its counterpart (MMHF11-00001b), at approximately the same scale. (C) Enlargement of pinna with three mines; orange arrowheads signify putative starting point (i.e., oviposition site) of leaf-mines; however, the starting point of the mine on the right is unseen. (D) Enlargement of two pinnules shows the transition of the frass trail, suggesting larval development while mining, and (E) its counterpart at the same scale. Arrowheads in orange and blue denote an oviposition site (os) and possible pupal chamber (pc), respectively.
Locality
Okubata, Omine, Mine, Yamaguchi.
Age
Carnian, Triassic (ca. 220 Ma).
Stratigraphy
Momonoki Formation, Mine Group, Yamaguchi prefecture, Japan.
Material
A single part-and-counterpart specimen (MMHF11-00001a, MMHF11-00001b; Fig. 2A,B, respectively) collected by H.T. from National Route 435 while the road was under construction (Fig. 1).
Host plant
The host plant is considered to be Cladophlebis nebbensis (Brongniart) Nathorst based on the following characteristics. Rachis 3 mm wide, grooved adaxially and rounded abaxially. Pinna subopposite, attached by a broad base to the rachis at an angle of 70–75 degrees, with internodes of 34 mm; apex unknown. Leaves sterile and bipinnate. Pinnules sessile, subopposite, arising at an angle of 71–81 degrees to the pinna rachis, catadromous in order; acroscopic pinnules slightly longer and narrower than basiscopic ones; close-set, occasionally overlapping. Pinnule about 11–14 mm long and 7–8 mm wide (surface area ca. 91.9 mm2), with widened base and obtuse apices; veins catadromous, having distinct primary vein reaching apical margin and 11–13 secondary veins forking once. This species may alternatively be Todites fukutomii Kimura et Ohana52, but this possibility is not considered here because some distinguishing characters (e.g., fertile pinnules and twice-forked secondary veins) are lacking in our specimen.
Remarks
The focal Cladophlebis frond is targeted by external foliage feeders in addition to miners. Excisions bordered by evident reaction rims (the area defined by two yellow asterisks in Fig. 2A) indicate signs of external feeding by a mandibulate folivore.
X-ray fluorescence (XRF) analyses
Elements were quantified at four sample points: (A) frass trail, (B) leaf vein, (C) leaf lamina, (D) rock matrix (Fig. 3). The XRF spectrum detected peaks of ten elements (Al, Si, P, S, K, Ca, Ti, Fe, Sr, Zr); the other peaks were of Rh gas used for the analyses. The values for four elements (Si, P, S, Ca) were particularly notable because these elements could be at least partly biogenic and involved in physiological processes. The leaf lamina was composed of just a thin film of plant tissue, and thus the elemental composition might include that of rock beneath the lamina. The results for the leaf lamina generally showed a similar trend with that of rock; however, P and S, presumably of biogenic origin, were detected, confirming that leaf tissue was preserved on the rock. We report the results below as the total amount of X-ray per sec (cps). Marked variations were found in Si; it was highest in the leaf lamina (376.457 ± 5.394 cps), slightly higher than in rock (366.144 ± 5.323 cps), and lowest in the leaf vein (114.689 ± 3.029 cps). The highest P was recorded in the leaf vein (11.680 ± 1.149 cps), which was followed by P in frass (7.923 ± 0.992 cps), whereas peaks for P were not detected in the lamina or rock. S showed a similar trend as P, but with peaks in frass (29.034 ± 1.614 cps) and the vein (11.680 ± 1.149 cps) being higher than those of P. Ca was consistently detected from all measured sample points and showed only minor differences among them; it was highest in the leaf vein (19.380 ± 1.381 cps), followed by frass (17.519 ± 1.286 cps), rock (16.851 ± 1.251 cps), and leaf lamina (13.895 ± 1.158 cps).

Results of X-ray fluorescence (XRF) analyses conducted with an EA6000VX High Sensitivity XRF Analyzer (Hitachi High-Tech Science Corporation). (Upper panel) Positions of the four sample points are specified on the specimen. (Lower panels) XRF spectra in the range of 0–23 keV illustrated for each sample point (A–D). Vertical axes signify cps (X-ray counts per sec).
Discussion
Significant features of the mines and the possible culprit
The Cladophlebis mining structures from the Momonoki Formation are leaf mines by holometabolous insects and can be distinguished from other feeding methods (e.g., surface-feeding) and also from features of taphonomic origin because they more or less satisfy the following criteria for shapes of insect leaf-mines53: an oviposition site at one end of the mining structure; an enlarged oviposition area as a blotch or curvilinear trace; a sign of the evacuated leaf tissue; width of evacuated plant tissue and frass trail increases from one end to the other; the presence of a frass trail, either particulate or fluidized; response tissue along with the marginal tissue; a distinctive terminus, such as an expanded region (chamber). In this case, features of the frass trails are shared by the three mines on the same pinna (Fig. 2A,B), which are most likely to be produced by the same insect taxon who has a stereotyped strategies for consumption and excretion. Each mine is composed of a continuous, single frass trail, the width of which subsequently expands at the end. The abrupt change in the shapes of faecal tracks can be interpreted as larval development while mining. In the mined pinnules, boundaries between mined and unmined areas are obscure because the frass trails frequently cut across the secondary leaf veins without distorting them. This can be interpreted in two ways: the larvae might mine only epidermal cells and they did not consume mesophylls, as in the case of Phyllocnistis Zeller (Lepidoptera: Gracillariidae)54,55; otherwise, the larvae could mine mesophyll avoiding vascular tissues (e.g., some agromyzid flies56). The latter strategy is possible because the mesophyll of ferns is anatomically well-differentiated into palisade and spongy tissues57.
Mine morphology and host-plant range often provide us with keys to identify leaf-mining insects because leaf-mining insects tend to be associated with a relatively narrow range of plants and show stereotyped, taxon-characteristic behavioural patterns in oviposition and feeding2. However, in general, the convergence among different insect orders/families and variation among closely related species make it difficult to differentiate miners based on their mine shapes53.
The taxonomic affinity of the mines from the Momonoki Formation is herein examined in the light of mine shape, systematic and evolutionary backgrounds of leaf-mining clades, the extant groups of fern-pinnule miners, and the chronological origins of possible culprits. The overall mine shape is not comparable to those of known mining structures by extant fern-miners58. Extant fern-mining insects are found in four orders: Diptera, Coleoptera, Lepidoptera, and Hymenoptera59.
Leaf-mining flies are diverse, encompassing three infraorders, Culicomorpha (e.g., Chironomidae), Bibinomorpha (e.g., Sciaridae), and Muscomorpha (Empidoidea–Muscoidea). Among them, Agromyzidae contain an overwhelming number of leaf-mining taxa. No evidence for the presence of dipteran leaf-mining taxa is available for the Late Triassic, although nematocerans and some of the earliest groups of brachycerans are markedly diverse46,47. Typical leaf mines of Agromyzidae, to which all fossil dipteran leaf mines have been assigned60, make linear-blotch mines61, and mine through mesophylls leaving major veins62; they tend to contain fluidized frass, which is often deposited as two discontinuous rows of pellets19. However, the forms of leaf mines by Agromyzidae greatly vary among taxa63. Flies are relatively diverse as miners of fern pinnules (and stems), represented by Chirosia Rondani (Anthomyiidae)64, Agromyzidae (e.g., Chromatomyia Hardy, Phytoliriomyza Hendel), and Cecidomyiidae59. Among them, Chirosia is a predominant component that use fronds or stems of a range of fern taxa58,65,66,67,68,69,70. In mines of some species of Phytoliriomyza, the frass trail displays a shift from a narrow meandering line to a wider band of the faecal pellets, which, to some extent, resemble those of the mines from the Momonoki Formation.
Leaf-mining taxa of Hymenoptera are mainly composed of sawflies (Tenthredinidae). This order was already diverse by the end of the Triassic71, with the oldest fossils dating back to the Middle Triassic72. Leaf-mining sawflies produce large conspicuous blotch mines53,73 and the larvae deposit cylindrical frass pellets which are scattered irregularly about the mine cavities58,74. The extant fern-feeders of Hymenoptera do not include pinnule-miners, and only Blasticotomidae and Tenthredinidae, as petiole-borers and internal fern-feeders, respectively, are known59.
The coleopteran leaf miners are known from Buprestoidea, Chysomeloidea, Curculionoidea, and a few other clades60; fern-pinnule mining taxa are reported from these superfamilies59. Beetles became widespread worldwide in the Middle to Late Triassic75,76,77. The earliest beetle group, Protocoleoptera, are found from the Momonoki Formation34,45, although they are thought to be saproxylic (i.e., borers of decaying wood), based on some circumstantial evidence78. Linear mines containing granular faecal pellets from the Triassic are often assigned to beetles, e.g., Polyphaga79. Notably, buprestid mines have some distinctive features that are comparable to those of the described mines. The buprestid mines are typically blotchy; they often are full-depth mines, avoiding epidermal and vascular tissues; the frass trails are generally long and stringy when freshly deposited, which later can be fragmented and become granular58,80,81,82,83.
Lepidopteran leaf-miners mainly consist of microlepidopteran groups, including Nepticuloidea, Gracillaroidea, and part of Yponomeutoidea. Based on a recent fossil-calibrated molecular phylogeny84, the appearance of leaf-mining moth clades, represented by the split between Nepticulidae and Opostegidae, dates back to the Late Jurassic; for calibrating this phylogeny, wing-scale fossils of Coelolepida from the Triassic–Jurassic boundary of Germany were used85. The earliest presumed nepticulid leaf mine fossils are known from the Dakota Formation of the Early Cretaceous (102 Ma)18,86,87. The lepidopteran mines exhibit considerable variation in mine shape, tissue consumption, and contents (e.g., faecal pellets). Notably, leaf mines of Ectoedemia (Nepticulidae) typically start as fine, strongly meandering galleries that subsequently become broad blotches87. In addition, typical nepticulids generally leave granular pellets, with abrupt changes in the accumulation pattern in some species. Importantly, these mine features are seen in the Cladophlebis mine from the Momonoki Formation. The Gracillariid moths produce serpentine or blotch mines and the mode of leaf-mining is conserved at the subfamily level88. Many gracillariid taxa (Acrocercopinae, some Gracillariinae, and Ornixolinae, and Lithocollectinae) make a narrow linear mine during the sap-feeding phase of early instars which later become a simple blotch mine. With regard to the fossil and molecular evidence, it is therefore unlikely that a member Nepticulidae caused the Late Triassic mine, although the Nepticuloidea or another early leaf-mining moth group cannot be ruled out.
Altogether, the shape of the Cladophlebis mines does not conform to typical mines of Diptera (Agromyzidae) and Hymenoptera; instead, it shares more features with those of Coleoptera or Lepidoptera in terms of the mine shapes. The assignment of a fossil leaf mine to a particular taxonomic group of insects is subject to uncertainty and limitations19. Available biological accounts of leaf-mining insects are limited, and leaf mines are often not photographed or illustrated. Particularly, in this case, a series of diagnostic features—oviposition habit (e.g., oviposition scar, deposition of eggs), pupal chamber, mining tissue types (e.g., epidermis, parenchyma), relative position of the frass trail in the mine—were not readily recognizable. Furthermore, the mine recorded herein predates the estimated divergence time of major clades of leaf-mining insects in the modern, as examined above. Overall, the potential leaf-miner may be Coleoptera or Lepidoptera, in terms of the general resemblance of the mine shape, the time of appearance, and the presence of records at the order level, although the possibility of Hymenoptera and Diptera is not completely excluded, due to the lack of conclusive diagnostic features. Future taxonomic studies on the insect fossils from the same deposit would provide support for ascertaining the suspect leaf-miner.
The stoichiometric footprint of the studied plant–insect interaction
The elemental analyses indicate quantitative variability in some elements (Si, P, S) that may partly be responsible for physiological processes in nutritional cycles (Fig. 3); plant tissues are deposited and then partly removed by an insect and then the insect metabolizes and excretes the undigested substances. The content of frass is thus the product of absorption, metabolism, and excretion. Compared to the leaf vein, the fossilized frass (coprolites) are shown to be highly phosphatic, which is consistent with the fact that the coprolites are largely composed of calcium phosphate.
Another notable point is the varying intensity of Si among sample points. For the leaf lamina, biogenic and lithologic Si may be conjugated, and thus caution is needed in interpreting our result that the highest intensity of Si was found in the leaf lamina. However, the incremental difference of Si between the frass and leaf vein may be the result of biogenic silica contained in the frass. Biomineralization of silica, especially in the form of phytoliths (SiO2, nH2O), is found in many clades of pteridophytes89,90, and these phytoliths can enhance plant resistance to herbivore feeding91,92. This relatively higher content of Si in the frass coprolites may therefore reflect undigested defensive compounds.
This study illustrates that ecological stoichiometry, a method that traces the flow of energy and elements in ecosystems, can be applied to interactions between plants and endophytic herbivores which occurred 220 million years ago. Similarly, the synchrotron X-ray Fluorescence (SRS-XRF) analyses for exceptionally preserved leaf fossils of Acer pseudoplatanus L. from the Green River Formation (Eocene, ca. 50 Ma)93 have revealed a high concentration of some metal elements (e.g., Cu, S, Zn) in the plants and the faecal materials; however, phosphorus in the leaves was below the detection limits. Future investigations of plant chemical landscapes (nutrition and defence) across various temporal and spatial settings would provide new insights into the macroevolutionary patterns of combat between plants and herbivores.
Origin and early history of leaf-mining
Credible leaf mines are absent or very rare before the Late Triassic27. From the Palaeozoic, at least two types of trace fossils have been previously assigned as possible leaf mines, although this is currently not supported. One type concerns U- or V-shaped structures on pinnules of medullosans from the Upper Carboniferous, or Lower Permian94,95, which were later attributed to fungal or bacterial infection27,96,97. The other types represent a series of small and extensive serpentine structures94,98,99, the ichnotaxon names of which are Asteronomus meandriformis Müller and A. divergens Müller; currently, these are acknowledged as structures of taphonomic origin27. A notable example from the Early Permian is a possible mine on a megaphyll of Glossopteris cf. indica from the Rio Bonito Formation, Morro do Papaléo Mine, Brazil; this structure contained the possible frass of the miner and ended with a terminal expansion, which was possibly a larval/pupal chamber100. Another example is a U-shaped contour on the foliage of Vjaznikopteris rigida Naugolnykh from the Volga River Basin (the P–Tr boundary), European Russia101. However, the identity of these traces as leaf mines is disputable because the above-mentioned criteria for insect mines are not met.
From the Middle–Late Triassic, several distinct shapes of mining structures are known (Fig. 4), and some of them are described as distinct damage types (DT)102. Several sites of the Molteno Formation (Carnian) are a rich source of herbivory and oviposition trace fossils79,103. Two types of leaf mines, one of each from Heidiphyllum elongatum (Morris) foliage (DT41, DT71) and one from Sphenobaiera schenckii (Feistmantel) Florin (DT139), have been recorded from this locality79 (Fig. 4A); additionally, this locality bears an undescribed, well-preserved mine on Cladophlebis fern pinnules (Labandeira, C. C., pers. comm.), although its shape differs considerably from the one from the Momonoki Formation. Another record from the Gondwanan flora is a serpentine mine on Heidiphyllum Retallack foliage, Triassohyponomus dinmorensis Rozefelds et Sobbe, from the Blackstone Formation (Carnian) of the Ipswich Coal Measures Group, Queensland, Australia104,105 (Fig. 4B). Some trace fossils from several other localities of the Middle–Late Triassic have also been assigned to leaf mines; for example, a small, semilinear, frass-laden mining structure (DT40) on foliage of the pteridosperm Scytophyllum bergeri Bornemann, from Monte Agnello, N. Italy (Late Ladinian)106 (Fig. 4C). Some mining structures are also known from the foliage of Nilssonia sturii Krasser from the Lunz Formation (Carnian), in the eastern part of the Northern Calcareous Alps, Austria107 (Fig. 4D); also, two types of mines on some gymnosperm (?Glossophyllum) foliage are reported from Dzhayloucho (Ladinian–Carnian), near Madygen, Kyrghyzstan108,109 (Fig. 4E).

Mining structures known so far from the Middle–Late Triassic. (A) Heidiphyllum foliage mine (DT71) from the Molteno Formation (Carnian)79; two other recorded mines (DT41, DT139) from this formation are unillustrated. (B) Heidiphyllum foliage mine, Triassohyponomus dinmorensis, from the Blackstone Formation (Carnian) of the Ipswich Coal Measures Group, Queensland, Australia105. (C) Pteridosperm foliage mine from Monte Agnello, N. Italy (Late Ladinian)106. (D) Nilssonia foliage mine from the Lunz Formation (Carnian), eastern part of the Northern Calcareous Alps, Austria107. (E) Gymnosperm foliage mine from Dzhayloucho (Ladinian–Carnian), near Madygen, Kyrgyzstan108. (F) Cladophlebis pinnule mines from the Momonoki Formation (our study). (G) Paleogeographic map of the Late Triassic (Carnian) and the approximate locations of fossil mine localities are shown; floral zonation is based on a previous study44. Colours and three-letter acronyms represent floral zones as follows: SIB Siberian Subprovince, MAS Middle Asian Subprovince, NEA Northern East Asian Subprovince, SEA Southern East Asian Subprovince, ARC Arctic Canada Subprovince, NCE North Atlantic/Central European Subprovince, NEW Newark Subprovince, CHD Chinle/Dockum Subprovince, WTE Western Tethydean Subprovince, YZR Yarlung-Zangbo-River Subprovince, ONS Onslow Subprovince, IPS Ipswich Subprovince. Line drawings were made by Y.I. with Adobe Illustrator® 2021.
The Cladophlebis mines (Fig. 4F) described here represent a novel damage type that serves as the oldest credible fossil mine from the Southern Floristic Region of East Asia, the palaeobotanical assemblage of which is geographically and taxonomically distant from any of the above-mentioned floras (Fig. 4G). Our finding, therefore, reinforces the view that leaf-mining had become a pervasive feeding method for plant-feeding insects by the Late Triassic. By this time, they had already colonized a wide range of plant groups: conifers, pteridosperms, cycadophytes, ginkgophytes, and ferns.
Methods
Plant–arthropod interactions were censused for ca. 200 full storage boxes (ca. 536 × 336 cm) of fossil specimens. Thus, the examined surface area of fossils came to no less than 3600 m2; the specimens were collected from several localities of the Momonoki Formation. Although many plant–insect interactions were found and Cladophlebis fern fronds were a dominant component of the fossil floral assemblage, the leaf mines described in this paper were found on only one specimen. Three leaf mines were found on a shale, as one part and counterpart specimen. The fossil type specimen was identified by H.Y. The shale was collected from National Route 435 by H.T. with obtaining permission (Fig. 1). The fossil type specimen is deposited and publicly available in the Mine City Museum of History and Folklore (MMHF), Yamaguchi prefecture, Japan; the catalogue numbers are MMHF11-00001a and MMHF11-00001b. The geological map (Fig. 1) was made by H.Y. based on relevant studies38,39, and later slightly modified by Y.I., using Adobe Illustrator® 2021. The field study on fossil plants comply with relevant institutional, national, and international guidelines and legislation.
Photographs of materials were taken by N.O. with a Nikon D850 using three types of Macro-NIKKOR lens (120 mm F 46.3, 55 mm f 2.8, and 65 mm f 4.5). Photos were later edited by Y.I. with Adobe Photoshop® 2021 to increase the contrast and to erase the background. Areas and lengths were measured by Y.I. with Fiji (Fiji Is Just Image J)110. For preparing Fig. 4A–F, illustrations were made by Y.I. by tracing photos from the relevant literature with water-based drawing pens, scanned with a CanoScan LiDE 400 (Canon) at a resolution of 300 dpi, and edited with Adobe Photoshop® 2021.
To examine the stoichiometric footprint of this ancient plant–insect interaction, elements of the mine trace were analysed by H.Y. via energy-dispersive X-ray fluorescence (XRF) using a EA6000VX High Sensitivity XRF Analyzer (Hitachi High-Tech Science Corporation). An XRF spectrometer measures elements between Na and U in order of increasing atomic number. Elemental compositions were quantitatively measured from three points of the specimen where organic compounds derived from fossil leaves remained (one point was set on a frass trail of a leaf mine where insect's coprolites were visible; each one sample point was set on leaf vein and lamina of the same pinnule), and, as a control point, from the host sedimentary rock. The measurement duration was set at 120 s using a 0.2 × 0.2 mm collimator under an excitation voltage of 50 kV and a tube current of 1000 μA. Output data were visualized by Y.I. with ggplot2111 and formatted with Adobe Illustrator® 2021.
References
Connor, E. F. & Taverner, M. P. The evolution and adaptive significance of the leaf-mining habit. Oikos 79, 6–25. https://doi.org/10.2307/3546085 (1997).
Hespenheide, H. A. Bionomics of leaf-mining insects. Annu. Rev. Entomol. 36, 535–560. https://doi.org/10.1146/annurev.en.36.010191.002535 (1991).
Kato, M. Structure, organization, and response of a species-rich parasitoid community to host leafminer population dynamics. Oecologia 97, 17–25 (1994).
López, R., Carmona, D., Vincini, A. M., Monterubbianesi, G. & Caldiz, D. Population dynamics and damage caused by the leafminer Liriomyza huidobrensis Blanchard (Diptera: Agromyzidae), on seven potato processing varieties grown in temperate environment. Neotrop. Entomol. 39, 108–114. https://doi.org/10.1590/S1519-566X2010000100015 (2010).
Lopez-Vaamonde, C., Godfray, H. C. J. & Cook, J. M. Evolutionary dynamics of host-plant use in a genus of leaf-mining moths. Evolution 57, 1804–1821. https://doi.org/10.1111/j.0014-3820.2003.tb00588.x (2003).
Lopez-Vaamonde, C. et al. Fossil-calibrated molecular phylogenies reveal that leaf-mining moths radiated millions of years after their host plants. J. Evol. Biol. 19, 1314–1326. https://doi.org/10.1111/j.1420-9101.2005.01070.x (2006).
Scheffer, S. J., Lewis, M. L., Hébert, J. B. & Jacobsen, F. Diversity and host plant-use in North American Phytomyza Holly Leafminers (Diptera: Agromyzidae): Colonization, divergence, and specificity in a host-associated radiation. Ann. Entomol. Soc. Am. 114, 59–69. https://doi.org/10.1093/aesa/saaa034 (2021).
Tooker, J. F. & Giron, D. The evolution of endophagy in herbivorous insects. Front. Plant Sci. 11, 581816. https://doi.org/10.3389/fpls.2020.581816 (2020).
Hawkins, B. A. Pattern and Process in Host-Parasitoid Interactions (Cambridge University Press, 1994).
Novotny, V. & Basset, Y. Host specificity of insect herbivores in tropical forests. Proc. R. Soc. B Biol. Sci. 272, 1083–1090. https://doi.org/10.1098/rspb.2004.3023 (2005).
Lewis, O. T. et al. Structure of a diverse tropical forest insect-parasitoid community. J. Anim. Ecol. 71, 855–873. https://doi.org/10.1046/j.1365-2656.2002.00651.x (2002).
Hirao, T. & Murakami, M. Quantitative food webs of lepidopteran leafminers and their parasitoids in a Japanese deciduous forest. Ecol. Res. 23, 159–168. https://doi.org/10.1007/s11284-007-0351-6 (2008).
Pocock, M. J. O., Evans, D. M. & Memmott, J. The robustness and restoration of a network of ecological networks. Science 335, 973–977. https://doi.org/10.1126/science.1214915 (2012).
Leppänen, S. A., Altenhofer, E., Liston, A. D. & Nyman, T. Phylogenetics and evolution of host-plant use in leaf-mining sawflies (Hymenoptera: Tenthredinidae: Heterarthrinae). Mol. Phylogenet. Evol. 64, 331–341. https://doi.org/10.1016/j.ympev.2012.04.005 (2012).
Doorenweerd, C., Van Nieukerken, E. J. & Menken, S. B. J. A global phylogeny of leafmining Ectoedemia moths (Lepidoptera: Nepticulidae): Exploring host plant family shifts and allopatry as drivers of speciation. PLoS ONE 10, 1–20. https://doi.org/10.1371/journal.pone.0119586 (2015).
Nakadai, R. & Kawakita, A. Phylogenetic test of speciation by host shift in leaf cone moths (Caloptilia) feeding on maples (Acer). Ecol. Evol. 6, 4958–4970. https://doi.org/10.1002/ece3.2266 (2016).
Opler, P. A. Fossil lepidopterous leaf mines demonstrate the age of some insect-plant relationships. Science 179, 1321–1323. https://doi.org/10.1126/science.179.4080.1321 (1973).
Labandeira, C. C., Dilcher, D. L., Davis, D. R. & Wagner, D. L. Ninety-seven million years of angiosperm-insect association: Paleobiological insights into the meaning of coevolution. Proc. Natl. Acad. Sci. U. S. A. 91, 12278–12282. https://doi.org/10.1073/pnas.91.25.12278 (1994).
Winkler, I. S., Labandeira, C. C., Wappler, T. & Wilf, P. Distinguishing Agromyzidae (Diptera) leaf mines in the fossil record: New taxa from the Paleogene of North America and Germany and their evolutionary implications. J. Paleontol. 84, 935–954. https://doi.org/10.1666/09-163.1 (2010).
van Nieukerken, E. J., Doorenweerd, C., Hoare, R. J. B. & Davis, D. R. Revised classification and catalogue of global Nepticulidae and Opostegidae (Lepidoptera, Nepticuloidea). Zookeys 2016, 65–246. https://doi.org/10.3897/zookeys.628.9799 (2016).
Maccracken, S. A., Sohn, J.-C., Miller, I. M. & Labandeira, C. C. A new Late Cretaceous leaf mine Leucopteropsa spiralae gen. et sp. nov. (Lepidoptera: Lyonetiidae) represents the first confirmed fossil evidence of the Cemiostominae. J. Syst. Palaeontol. 19, 131–144. https://doi.org/10.1080/14772019.2021.1881177 (2021).
Wilf, P., Labandeira, C. C., Johnson, K. R. & Ellis, B. Decoupled plant and insect diversity after the end-Cretaceous extinction. Science 313, 1112–1115. https://doi.org/10.1126/science.1129569 (2006)
Donovan, M. P., Wilf, P., Labandeira, C. C., Johnson, K. R. & Peppe, D. J. Novel insect leaf-mining after the end-Cretaceous extinction and the demise of Cretaceous leaf miners, Great Plains, USA. PLoS ONE 9, e103542. https://doi.org/10.1371/journal.pone.0103542 (2014).
Donovan, M. P., Iglesias, A., Wilf, P., Labandeira, C. C. & Cúneo, N. R. Rapid recovery of Patagonian plant–insect associations after the end-Cretaceous extinction. Nat. Ecol. Evol. 1, 0012. https://doi.org/10.1038/s41559-016-0012 (2017).
Donovan, M. P., Wilf, P., Iglesias, A., Cúneo, N. R. & Labandeira, C. C. Persistent biotic interactions of a Gondwanan conifer from Cretaceous Patagonia to modern Malesia. Commun. Biol. 3, 708. https://doi.org/10.1038/s42003-020-01428-9 (2020).
Labandeira, C. C. The four phases of plant-arthropod associations in deep time. Geol. Acta 4, 409–438. https://doi.org/10.1344/105.000000344 (2006).
Labandeira, C. C. Silurian to Triassic plant and hexapod clades and their associations: new data, a review, and interpretations. Arthropod Syst. Phylogen. 64, 53–94 (2006).
Wakita, K., Nakagawa, T., Sakata, M., Tanaka, N. & Oyama, N. Phanerozoic accretionary history of Japan and the western Pacific margin. Geol. Mag. https://doi.org/10.1017/s0016756818000742 (2018).
Katayama, M. Stratigraphical study on the Mine Series. J. Geol. Soc. Jpn. 46, 127–141. https://doi.org/10.5575/geosoc.46.127 (1939).
Maeda, H. & Oyama, N. Stratigraphy and fossil assemblages of the Triassic Mine Group and Jurassic Toyora Group in western Yamaguchi Prefecture. J. Geol. Soc. Japan 125, 585–594. https://doi.org/10.5575/geosoc.2019.0020 (2019).
Aizawa, J. Fossil insect-bearing strata of the Triassic Mine Group, Yamaguchi Prefecture. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser. A 10, 91–98 (1991).
Oyama, N. & Maeda, H. Madygella humioi sp. nov. from the Upper Triassic Mine Group, Southwest Japan: The oldest record of a sawfly (Hymenoptera: Symphyta) in East Asia. Paleontol. Res. 24, 64–71 (2020).
Fujiyama, I. Mesozoic insect fauna of East Asia part 1. Introduction and upper Triassic faunas. Bull. Natl. Sci. Mus. 16, 331–386 (1973).
Fujiyama, I. Late Triassic insects from Mine, Yamaguchi, Japan, Part 1. Odonata. Bull. Natl. Sci. Mus. Tokyo Ser. C 17, 49–56 (1991).
Ueda, K. A Triassic fossil of scorpion fly from Mine, Japan. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser. Ser. A 10, 99–103 (1991).
Takahashi, F., Ishida, H., Nohara, M., Doi, E. & Taniguchi, S. Occurrence of insect fossils from the Late Triassic Mine Group. Bull. Mine City Mus. Yamaguchi Prefect. Jpn. 13, 1–27 (1997).
Kametaka, M. Provenance of the Upper Triassic mine group Southwest Japan. J. Geol. Soc. Jpn. 105, 651–667 (1999).
Takahashi, E. & Mikami, T. Triassic. In Geology of Yamaguchi Prefecture (ed. Yamaguchi Museum) 93–108 (Yamaguchi Museum, 1975).
Kiminami, K. Atsu Group and Mine Group. In Monograph on Geology of Japan 6, Chugoku Region (ed. Geological Society of Japan) 85–88 (Asakura Publishing Co., Ltd., 2009).
Naito, G. Plant Fossils from the Mine Group (Mine City Education Comittee, 2000).
Kimura, T. Geographical distribution of Palaeozoic and Mesozoic plants in East and Southeast Asia. Hist. Biogeogr. Plate Tecton. Evol. Jpn. East Asia 1982, 135–200 (1987).
Kimura, T., Naito, G. & Ohana, T. Baiera cf. furcata (Lindley and Hutton) Braun from the Carnic Momonoki Formation, Japan. Bull. Natl. Sci. Mus. 9, 91–114 (1983).
Katagiri, T. Pallaviciniites oishii (comb. Nov.), a thalloid liverwort from the Late Triassic of Japan. Bryologist 118, 245–251. https://doi.org/10.1639/0007-2745-118.3.245 (2015).
Kustatscher, E. et al. Flora of the Late Triassic. In The Late Triassic World, Topics in Geobiology, Vol. 46 (ed. Tanner, L. H.) 545–622 (Springer, 2018). https://doi.org/10.1007/978-3-319-68009-5_13.
Oyama, N., Yukawa, H. & Maeda, H. Mesozoic insect fossils of Japan: Significance of the Upper Triassic insect fauna of the Mine Group, Yamaguchi Pref. Bull. Mine City Mus. Yamaguchi Prefect. Jpn. 33, 1–13 (2020).
Shcherbakov, D. E., Lukashevich, E. D. & Blagoderov, V. Triassic Diptera and initial radiation of the order. Int. J. Dipterol. Res. 6, 75–115 (1995).
Krzemiński, W. & Krzemińska, E. Triassic Diptera: Descriptions, revisions and phylogenetic relations. Acta Zool. Cracov. 46, 153–184 (2003).
Blagoderov, V., Grimaldi, D. A. & Fraser, N. C. How time flies for flies: Diverse Diptera from the Triassic of Virginia and early radiation of the order. Am. Mus. Novit. 3572, 1–39. https://doi.org/10.1206/0003-0082(2007)509[1:HTFFFD]2.0.CO;2 (2007).
Lukashevich, E. D., Przhiboro, A. A., Marchal-Papier, F. & Grauvogel-Stamm, L. The oldest occurrence of immature Diptera (Insecta), Middle Triassic France. Ann. la Société Entomol. Fr. 46, 4–22. https://doi.org/10.1080/00379271.2010.10697636 (2010).
Schmidt, A. R. et al. Arthropods in amber from the Triassic Period. Proc. Natl. Acad. Sci. 109, 14796–14801. https://doi.org/10.1073/pnas.1208464109 (2012).
Lara, M. B. & Lukashevich, E. D. The first Triassic dipteran (Insecta) from South America, with review of Hennigmatidae. Zootaxa 3710, 81–92. https://doi.org/10.11646/zootaxa.3710.1.6 (2013).
Kimura, T. & Ohana, T. Some fossil ferns from the Middle Carnic Momonoki Formation, Yamaguchi prefecture, Japan. Bull. Natl. Sci. Mus. Ser. C Geol. Paleontol. 6, 73–92 (1980).
Hering, E. M. Biology of the Leaf Miners https://doi.org/10.1007/978-94-015-7196-8. (Springer, 1951).
Kirichenko, N. et al. Systematics of Phyllocnistis leaf-mining moths (Lepidoptera, Gracillariidae) feeding on dogwood (Cornus spp.) in Northeast Asia, with the description of three new species. Zookeys 2018, 79–118. https://doi.org/10.3897/zookeys.736.20739 (2018).
Cerdeña, J. et al. Phyllocnistis furcata sp. nov.: A new species of leaf-miner associated with Baccharis (Asteraceae) from Southern Peru (Lepidoptera, Gracillariidae). Zookeys 2020, 121–145. https://doi.org/10.3897/zookeys.996.53958 (2020).
Elb, P. M., Melo-de-Pinna, G. F. & de Menezes, N. L. Morphology and anatomy of leaf miners in two species of Commelinaceae (Commelina diffusa Burm. F. and Floscopa glabrata (Kunth) Hassk). Acta Bot. Brasilica 24, 283–287. https://doi.org/10.1590/S0102-33062010000100030 (2010).
Vasco, A., Moran, R. C. & Ambrose, B. A. The evolution, morphology, and development of fern leaves. Front. Plant Sci. 4, 1–16. https://doi.org/10.3389/fpls.2013.00345 (2013).
Eiseman, C. Leafminers of North America. (Charley Eiseman, 2019).
Yang, J., Wang, X., Duffy, K. & Dai, X. A preliminary world checklist of fern-mining insects. Biodivers. Data J. 9, e62839. https://doi.org/10.3897/BDJ.9.e62839 (2021).
Ding, Q., Labandeira, C. C. & Ren, D. Biology of a leaf miner (Coleoptera) on Liaoningocladus boii (Coniferales) from the Early Cretaceous of northeastern China and the leaf-mining biology of possible insect culprit clades. Arthropod Syst. Phylogen. 72, 281–308 (2014).
Boucher, S. Revision of the Canadian species of Amauromyza Hendel (Diptera: Agromyzidae). Can. Entomol. 144, 733–757. https://doi.org/10.4039/tce.2012.80 (2012).
Scheirs, J., Vandevyvere, I. & De Bruyn, L. Influence of monocotyl leaf anatomy on the feeding pattern of a grass-mining agromyzid (Diptera). Ann. Entomol. Soc. Am. 90, 646–654 (1997).
Boucher, S. Leaf-miner flies (Diptera: Agromyzidae). In Encyclopedia of Entomology (ed. Capinera J. L.) 2163–2169 (Springer, 2008). https://doi.org/10.1007/978-1-4020-6359-6.
Eiseman, C. S. New rearing records for muscoid leafminers (Diptera: Anthomyiidae, Scathophagidae) in the United States. Proc. Entomol. Soc. Wash. 120, 25–50. https://doi.org/10.4289/0013-8797.120.1.25 (2018).
Meikle, A. A. The insects associated with bracken. Agric. Prog. 14, 58–61 (1937).
Lawton, J. H. The structure of the arthropod community on bracken. Bot. J. Linn. Soc. 73, 187–216. https://doi.org/10.1111/j.1095-8339.1976.tb02022.x (1976).
Lawton, J. H., MacGarvin, M. & Heads, P. A. Effects of altitude on the abundance and species richness of insect herbivores on bracken. J. Anim. Ecol. 56, 147–160. https://doi.org/10.2307/4805 (1987).
Cooper-Driver, Gi. A. Insect-fern associations. Entomol. Exp. Appl. 24, 310–316. https://doi.org/10.1111/j.1570-7458.1978.tb02787.x (1978).
Eiseman, C. S. Further Nearctic rearing records for phytophagous muscoid flies (Diptera: Anthomyiidae, Scathophagidae). Proc. Entomol. Soc. Washingt. 122, 595–603. https://doi.org/10.4289/0013-8797.122.3.595 (2020).
Santos, M. G. & Maia, V. C. A synopsis of fern galls in Brazil. Biota Neotrop. 18, e20180513. https://doi.org/10.1590/1676-0611-BN-2018-0513 (2018).
Peters, R. S. et al. Evolutionary history of the Hymenoptera. Curr. Biol. 27, 1013–1018. https://doi.org/10.1016/j.cub.2017.01.027 (2017).
Ronquist, F. et al. A total-evidence approach to dating with fossils, applied to the early radiation of the Hymenoptera. Syst. Biol. 61, 973–999. https://doi.org/10.1093/sysbio/sys058 (2012).
Needham, J. G., Frost, S. W. & Tothill, B. H. Leaf-Mining Insects (Waverly Press, 1928).
Smith, D. R., Eiseman, C. S., Charney, N. D. & Record, S. A new Nearctic Scolioneura (Hymenoptera, Tenthredinidae) mining leaves of Vaccinium (Ericaceae). J. Hymenopt. Res. 43, 1–8. https://doi.org/10.3897/JHR.43.4546 (2015).
Zheng, D. et al. Middle-Late Triassic insect radiation revealed by diverse fossils and isotopic ages from China. Sci. Adv. 4, eaat1380. https://doi.org/10.1126/sciadv.aat1380 (2018).
Zhang, S. Q. et al. Evolutionary history of Coleoptera revealed by extensive sampling of genes and species. Nat. Commun. 9, 1–11. https://doi.org/10.1038/s41467-017-02644-4 (2018).
McKenna, D. D. et al. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. 116, 24729–24737. https://doi.org/10.1073/pnas.1909655116 (2019).
Gimmel, M. L. & Ferro, M. L. General overview of saproxylic Coleoptera. In Saproxylic Insects, Zoological Monographs, Vol. 1 (ed. Ulyshen, M. D.) 51–128 (Springer, 2018). https://doi.org/10.1007/978-3-319-75937-1_2.
Labandeira, C. C., Anderson, J. M. & Anderson, H. M. Expansion of arthropod herbivory in Late Triassic South Africa: The Molteno Biota, Aasvoëlberg 411 site and developmental biology of a gall. In The Late Triassic World, Topics in Geobiology Vol. 46 (ed. Tanner, L. H.) 623–719 (Springer International Publishing AG, 2018).
Fiebrig, K. Eine Schaum bildende Käferlarve Pachyschelus spec. (Bupr. Sap.) Die Ausscheidung von Kautschuk aus der Nahrung und dessen Verwertung zu Schutzzwecken (auch bei Rhynchoten). Z. f. Wiss. Insektenbiol. 4, 333–339 (1908).
Bruch, C. Metamórfosis de Pachyschelus undularius (Burm.). Physis 3, 30–36 (1917).
Hering, E. M. Neotropische Buprestiden-Minen. Arb. Physiol. Angew. Entomol. 9, 241–249 (1942).
Kogan, M. Contribuição ao conhecimento da sistemática e biologia de buprestídeos minadores do gênero Pachyschelus Solier, 1833: (Coleoptera, Buprestidae). Mem. Inst. Oswaldo Cruz 61, 429–457 (1963).
Kawahara, A. Y. et al. Phylogenomics reveals the evolutionary timing and pattern of butterflies and moths. Proc. Natl. Acad. Sci. 116, 22657–22663. https://doi.org/10.1073/pnas.1907847116 (2019).
Van Eldijk, T. J. B. et al. A Triassic-Jurassic window into the evolution of lepidoptera. Sci. Adv. 4, e1701568. https://doi.org/10.1126/sciadv.1701568 (2018).
Sohn, J. C., Labandeira, C. C., Davis, D. & Mitter, C. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world. Zootaxa. https://doi.org/10.11646/zootaxa.3286.1.1 (2012).
Doorenweerd, C., Van Nieukerken, E. J., Sohn, J. C. & Labandeira, C. C. A revised checklist of Nepticulidae fossils (Lepidoptera) indicates an Early Cretaceous origin. Zootaxa 3963, 295–334. https://doi.org/10.11646/zootaxa.3963.3.2 (2015).
Kawahara, A. Y. et al. A molecular phylogeny and revised higher-level classification for the leaf-mining moth family Gracillariidae and its implications for larval host-use evolution. Syst. Entomol. 42, 60–81. https://doi.org/10.1111/syen.12210 (2017).
Mazumdar, J. Phytoliths of pteridophytes. S. Afr. J. Bot. 77, 10–19. https://doi.org/10.1016/j.sajb.2010.07.020 (2011).
Trembath-Reichert, E., Wilson, J. P., McGlynn, S. E. & Fischer, W. W. Four hundred million years of silica biomineralization in land plants. Proc. Natl. Acad. Sci. U. S. A. 112, 5449–5454 https://doi.org/10.1073/pnas.1500289112 (2015).
Hunt, J. W., Dean, A. P., Webster, R. E., Johnson, G. N. & Ennos, A. R. A novel mechanism by which silica defends grasses against herbivory. Ann. Bot. 102, 653–656. https://doi.org/10.1093/aob/mcn130 (2008).
Reynolds, O. L., Keeping, M. G. & Meyer, J. H. Silicon-augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 155, 171–186. https://doi.org/10.1111/j.1744-7348.2009.00348.x (2009).
Edwards, N. P. et al. Leaf metallome preserved over 50 million years. Metallomics 6, 774–782. https://doi.org/10.1039/C3MT00242J (2014).
Müller, A. H. Über Hyponome fossiler und rezenter Insekten, erster Beitrag. Freib. Forschungsh. C 366, 7–27 (1982).
Beck, A. L. & Labandeira, C. C. Early Permian insect folivory on a gigantopterid-dominated riparian flora from north-central Texas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 142, 139–173. https://doi.org/10.1016/S0031-0182(98)00060-1 (1998).
Jarzembowski, E. A. The oldest plant-insect interaction in Croatia: Carboniferous evidence. Geol. Croat. 65(3), 387–392. https://doi.org/10.4154/GC.2012.28 (2002).
Donovan, M. P. & Lucas, S. G. Insect herbivory on the Late Pennsylvanian Kinney Brick Quarry Flora, New Mexico, USA. Kinney Brick Quarry Lagerstätte. N. M. Mus. Nat. Hist. Sci. Bull. 84, 193–207 (2021).
Potonié, R. Ueber das Rothliegende des Thüringer Waldes. Theil II: Die Flora des Rothliegenden von Thüringen. Abh. Preuss. Geol. Landesanst. 9, 1–298 (1893).
Potonié, R. Mitteilungen über mazerierte kohlige Pflanzenfossilien. Z. Bot. 13, 79–88 (1921).
Adami-Rodrigues, K. A., Iannuzzi, R. & Pinto, I. D. Permian plant-insect interactions from a Gondwana flora of southern Brazil. Foss. Strat. 51, 106–126 (2004).
Krassilov, V. A. & Karasev, E. First evidence of plant–arthropod interaction at the Permian–Triassic boundary in the Volga Basin European Russia. Alavesia 2, 247–252 (2008).
Labandeira, C. C., Wilf, P., Johnson, K. & Marsh, F. Guide to insect (and other) damage types on compressed plant fossils. Version 3.0. Smithson. Institution, Washington, DC 25 (2007).
Scott, A. C., Anderson, J. M. & Anderson, H. M. Evidence of plant-insect interactions in the Upper Triassic Molteno formation of South Africa. J. Geol. Soc. London. 161, 401–410. https://doi.org/10.1144/0016-764903-118 (2004).
Tillyard, R. J. Mesozoic Insects of Queensland No. 9. Orthoptera, and Additions to the Protorthoptera, Odonata, Hemiptera, and Planipennia. Proc. Linn. Soc. N. S. W. 47, 447–470 (1922).
Rozefelds, A. C. & Sobbe, I. Problematic insect leaf mines from the Upper Triassic Ipswich Coal Measures of Southeastern Queensland Australia. Alcheringa 11, 51–57 (1987).
Wappler, T., Kustatscher, E. & Dellantonio, E. Plant-insect interactions from Middle Triassic (late Ladinian) of Monte Agnello (Dolomites, N-Italy)-Initial pattern and response to abiotic environmental pertubations. PeerJ 2015, e921. https://doi.org/10.7717/peerj.921 (2015).
Meller, B., Ponomarenko, A. G., Vasilenko, D. V., Fischer, T. C. & Aschauer, B. First beetle elytra, abdomen (Coleoptera) and a mine trace from Lunz (Carnian, Late Triassic, Lunz-am-See, Austria) and their taphonomical and evolutionary aspects. Palaeontology 54, 97–110. https://doi.org/10.1111/j.1475-4983.2010.01009.x (2011).
Vassilenko, D. V. Traces of plant-arthropod interactions from Madygen (Triassic, Kyrgyzstan): Preliminary data. Sovremennaya paleontologia: klassicheskie i noveishie metody 9–16 (2009).
Zherikhin, V. V. Insect Trace Fossils. In History of Insects (ed. Rasnitsyn A. P., Quicke, D. L.) 303–324 (Kluwer Academic Publishers, 2010).
Schindelin, J. et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 9, 676–682. https://doi.org/10.1038/nmeth.2019 (2012).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer-Verlag, 2016).
Acknowledgements
We thank members of the Mine City Museum of History and Folklore and the Mine Fossil Museum; Hiroyuki Makara (ITCFP) and Soichiro Kawabe (FPDM/FPU) for assisting with XRF analyses; Makoto Kato for providing us with valuable information on extant leaf-mining insects; Conrad C. Labandeira for discussion and sharing unpublished data; Robert B. Davis for proofreading the earlier version of the manuscript. This study was supported by a research grant for Environmental Field Research by the Asahi Glass Foundation (Asahi Glass Co., Ltd.), Grant-in-Aid for Scientific Research (KAKENHI) grant numbers JP20K15852 (Y.I.) and JP19J20625 (N.O.) from the Japan Society for the Promotion of Science (JSPS).
Author information
Authors and Affiliations
Contributions
Y.I. conceived the project. Y.I., N.O., and H.Y. contributed to the data collection of trace fossils. Y.I. analysed the data and wrote the paper with input from all co-authors. Y.I. made the illustrations, visualizations, and drawings, and interpreted the data with input from H.Y., N.O.; H.T. and H.Y. identified the host plant, performed XRF analyses, and provided palaeobotanical and geological input. N.O. took photos of the specimens in Fig. 2 and provided palaeoentomological input. H.T. collected the specimen. K.S. contributed to the curation of the museum collection. All co-authors commented on the drafts of the manuscript and approved the final manuscript for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Imada, Y., Oyama, N., Shinoda, K. et al. Oldest leaf mine trace fossil from East Asia provides insight into ancient nutritional flow in a plant–herbivore interaction. Sci Rep 12, 5254 (2022). https://doi.org/10.1038/s41598-022-09262-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-09262-1
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
