
Abstract
The alteration of hippocampal function by chronic stress impairs higher order cognitive functions such as prospective memory (PM). However, how chronic stress affects hippocampal subregions related to PM remains largely unknown. In this study, the altered functional network of hippocampal subregions related to PM in chronic stress was explored. College students (N = 21) completed PM tasks and resting-state functional magnetic resonance imaging scans one month prior to (baseline) and during the final examination week (chronic stress). Hippocampal subregions’ seed-based functional connectivity (FC) and PM were compared between baseline and chronic stress. PM performance declined in chronic stress. The FC of the cornu ammonis 2, 3 and dentate gyrus (CA23DG) with the bilateral caudate and precuneus was increased in chronic stress, while the FC of the subicular complex (SUBC) with the left middle frontal gyrus, the left inferior parietal gyrus and the right supramarginal gyrus was decreased. There was a negative correlation between PM performance and the FC of hippocampal subregions. We found chronic stress impairs PM by decreasing the FC of SUBC and increasing the FC of CA23DG. These findings suggest functional changes in hippocampal subregion networks as a mechanism underlying the impairment of PM in chronic stress.
Similar content being viewed by others
Introduction
Chronic stress can be defined as long-term exposure to repeated experiences perceived to be stressful1. Coincident with economic development, chronic stress has increased in modern society, and it has a long-term impact on mental health2. Numerous studies have indicated that chronic stress impairs high-order cognitive function as exemplified by impairment of prospective memory (PM)3,4,5. Previous studies found that PM is sensitive to stress6,7,8. Exposure to chronic stress leads to activation of hypothalamic-pituitary-adrenocortical (HPA) axis and the release of glucocorticoids9. These bind to receptors in specific areas of the brain, such as the hippocampus, a region critical for PM10. A set of recent neuroimaging studies found that acute stress altered cognition-related activity in specific brain regions11,12. However, the effect of chronic stress on PM related to hippocampual function in humans remains unclear.
PM is a form of memory that involves remembering to perform a planned action or recall a planned intention that is supported by the frontoparietal network13,14. There are two kinds of prospective memory: time- and event-based PM (TBPM and EBPM, respectively)15,16. Previous study found the commonalities and differences in the neural substrates of EBPM and TBPM17. Planned information is stored in the hippocampus, as a buffer of memory information18. Previous studies have indicated that the hippocampus is one of the crucial neural substrates of PM10. Exposure to chronic stress leads to reversible impairment of hippocampus morphology, accompanied by dysfunction of PM19,20. Empirical evidence from multiple pharmacological studies in animals has shown that chronic stress has different effects on different hippocampal subregions, such as neuronal atrophy in CA3 and change of GABAergic inhibition in CA121,22. In sum, chronic stress appears to impair the function and/or morphology of the hippocampus related to PM. However, the potential relationship between hippocampal subregions and different kinds of prospective memory in chronic stress remains unclear.
The hippocampus is a crucial subcortical structure for rapid acquisition and persistence of memories10. Chronic stress activates the HPA axis, releasing corticosteroids, for which the hippocampus contains the highest density of receptors23. Besides, chronic stress alters structure and/or neurotransmitter metabolism in hippocampus related to changes of its neurocircuitry, with implications for its stress-associated dysfunctions24,25. Hence, the hippocampus is sensitive to stress through the effect of corticosteroids, which have a significant impact on its functions, such as PM26,27. Importantly, the hippocampus is a heterogeneous brain region that can be divided into three subregions: the cornu ammonis (CA1, CA2, and CA3), the dentate gyrus (DG), and the subicular complex (SUBC)28. On one hand, according to recent research, hippocampal subregions may be involved in different cognitive processes within the memory system29,30,31,32. For instance, Suthana found that the CA2, CA3, and dentate gyrus (CA23DG) function to code new associations with novel information, while the SUBC functions to retrieve learned associations33. Chronic stressors also elicit subregion specific responses34. Previous studies found that chronic stress causes atrophy of CA3 pyramidal neurons but not in the DG35. According to the above findings, chronic stress may cause specific effect on different hippocampal subregion function and/or structure which are involved in the processing of PM.
In the present study, we chose final examinations as a chronic stressor for academic performance from a few days of written exams is the primary factor determining success for freshman in Chinese college36. Performance on final examinations affects scholarship applications and postgraduate recommendations. Hence, college students spend a lot of time preparing for examinations and are inevitably in a state of stress during this period37. In general, there are seven or eight written examination in one week at the end of the term.
We used resting-state functional magnetic resonance imaging (fMRI) to investigate how chronic stress alters the hippocampal subregion network related to PM. We hypothesized that (1) participants would score lower on PM tasks in chronic stress, (2) there would be significant changes in the functional connectivity (FC) of hippocampal subregions under chronic stress compare to baseline, and (3) PM performance would be selectively associated with changes in the FC of different hippocampal subregions in chronic stress.
Results
Sample description
The demographic characteristics are shown in Table 1. Twenty-one college students were recruited in our study (11 male, 10 female; age = 19.89 ± 0.83 years). They came from different majors, including Psychology, Fine Art, Politics and Music. The intervals between the two time points was 31.25 ± 2.67 days (Table 1).
Neuropsychological scores
The average SLSI score was significantly increased in chronic stress compared to baseline (Conflict-SLSI: t = 2.762, p = 0.012; Change-SLSI: t = 3.454, p = 0.003; Total scores-SLSI: t = 2.864, p = 0.010) (Table 1). The IQ (intelligence quotient) of participants was 123.67 ± 10.63 (Table 1).
Performance of PM
Compared to baseline, prospective memory task scores significantly decreased in chronic stress (TBPM: t = 0.00, p = −6.06, EBPM: t = 0.04, p = −2.15) (Table 1). As shown in Table 2, these was significant difference in difference of EBPM and TBPM between baseline and chronic stress (t = 3.342, p = 0.002).
Hippocampal subregion network within-group analysis
The patterns of each hippocampal subregion FC network at baseline and under chronic stress are illustrated in Fig. 1. In both conditions, each functional network of hippocampal subregions involved diffuse subcortical, medial frontal, temporal cortical, parietal and cerebellar sites (p < 0.0001, corrected by FDR). The patterns of FC were similar to the network connections observed in previous studies on the whole and bilateral hippocampus38,39. However, in both chronic stress and baseline conditions, the stronger FC of hippocampal subregional seeds was located in the temporal lobe. In addition, regions that were close to the seed region showed stronger connectivity than other regions in each of the three hippocampal subregions.

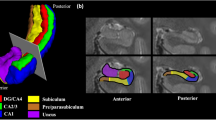
Functional connectivity pattern of hippocampal subregion networks in baseline and chronic stress. Statistical significance was set at p < 0.0001, corrected by FDR. Abbreviations: CA1, Cornu ammonis1; CA23DG, Cornu ammonis 2, 3 and dentate gyrus; SUBC, subicular complex.
Longitudinal changes in hippocampal subregion networks
As shown in Fig. 2, the CA23DG showed greater FC with the bilateral caudate and precuneus in chronic stress compared to baseline. In addition, the SUBC showed decreased FC with the left middle frontal gyrus, left inferior parietal gyrus, and right supramarginal gyrus in chronic stress compared to baseline (Fig. 2). There was no significant change in the CA1 network.

Longitudinal changes in CA23DG and SUBC hippocampal subregional networks. Compared to baseline, (1) the CA23DG subregion showed increased FC with bilateral caudate and precuneus during chronic stress, (2) the SUBC subregion showed decreased FC with the left middle frontal gyrus, left inferior parietal gyrus and right supramarginal gyrus during chronic stress. Significance threshold was set at p < 0.05, AlphaSim corrected. Abbreviations: CA23DG, Cornu ammonis 2, 3 and dentate gyrus; SUBC, subicular complex.
Correlations among hippocampal subregional networks, SLSI scores and PM performance in chronic stress
As shown in Table 3, the FC between the bilateral caudate and the CA23DG was positively correlated with SLSI scores (Conflict-SLSI: r = 0.616, p = 0.011; Stress-SLSI: r = 0.582, p = 0.018; Change-SLSI: r = 0.543, p = 0.030; Emotional reaction-SLSI: r = 0.571, p = 0.021; Total scores- SLSI: r = 0.667, p = 0.005) (Table 3). The FC between the left inferior parietal gyrus and the SUBC was negatively correlated with the SLSI scores (Physiological reaction: r = −0.487, p = 0.047) (Table 3). A significant negative correlation was also found between the FC of hippocampal subregional networks and PM performance in chronic stress. Particularly, the FC between the bilateral caudate and the CA23DG was negatively correlated with EBPM performance (r = −0.440, p = 0.046; Fig. 3). The FC between the left inferior parietal gyrus and the SUBC was negatively correlated with EBPM performance (r = −0.525, p = 0.015; Fig. 4). In addition, the FC of the SUBC with the left inferior parietal gyrus was negatively correlated with TBPM performance (r = −0.496, p = 0.022; Fig. 5). There was no significant result with regard to the moderating effect.

The association between mean functional connectivity of CA23DG-Caudate and performance of event-based prospective memory in chronic stress. Abbreviations: CA23DG: Cornu ammonis 2, 3 and dentate gyrus.

The association between mean functional connectivity of SUBC-LIPG and performance of event-based prospective memory in chronic stress. Abbreviations: SUB, subicular complex; LIPG, left inferior parietal gyrus.

The association between mean functional connectivity of SUBC–LIPG and performance of time-based prospective memory in chronic stress. Abbreviations: SUB, subicular complex; LIPG, left inferior parietal gyrus.
Discussion
In this study, we aimed to investigate chronic-stress-induced modulations in the activity of PM-related hippocampal subregions. The results confirmed our hypothesis regarding the altered FC of hippocampal subregion networks in chronic stress. Our results showed that (a) participants scored lower on the PM task in chronic stress compared to baseline; (b) the CA23DG FC network showed significantly increased FC with the bilateral caudate and precuneus, while the SUBC network showed significantly decreased FC with the left middle frontal gyrus, left inferior parietal gyrus and right supramarginal gyrus in chronic stress; and (c) the altered FC of hippocampal subregions was significantly associated with PM performance in chronic stress. These results may provide insight into the underlying neural mechanism of PM impairment related to hippocampal subregions in chronic stress.
During examination week, participants showed higher stress levels compared to baseline. This finding is consistent with previous studies, which showed that academic examinations are a key factor stress-causing for Chinese students36,37. In addition, we found a correlation between SLSI scores and the FC of hippocampal subregions. This indicated that the altered FC of hippocampal subregions may be related to increased stress levels. Consist with previous study that chronic stress causes hippocampal morphological changes and impairs hippocampal function35. In the present study, participants had lower scores on TBPM and EBPM during chronic stress compared to baseline. A previous study indicated that chronic stress affects cognitive function including PM4. Interestingly, although both EBPM and TBPM were affected by stress in our study, TBPM was affected to a greater extent compared to EBPM. These finding suggest that TBPM is more sensitive to chronic stress than EBPM. This is in agreement with other studies that have found TBPM to be more affected by emotional factors than EBPM8. Moreover, our results support Ellis and Ashbrook’s (1988) model suggesting that emotional factors adjust the allocation of cognitive resources, which are more necessary in TBPM tasks than EBPM tasks15,40,41.
We found an alteration of the intrinsic FC network of CA3DG and SUBC, accompanied by impairment of PM, in chronic stress. Previous studies have indicated that stress activates the HPA, releasing stress-related hormones that bind to receptors in hippocampus, which affects memory-related function42. Our study found opposite FC activation patterns in different hippocampal subregions, with activated FC of the CA23DG network and deactivated FC of the SUBC network. Numerous animal studies have shown that stress has distinct impacts on different hippocampal subregions which may involve in different cognitive processes35,43. Moreover, previous studies reported functional dissociation in hippocampal subregions33. This study concurred with those findings and extended the knowledge by demonstrating chronic stress modulates the FC of hippocampal subregions.
In the current study, the FC of the CA23DG with the caudate was negatively correlated with EBPM scores. Previous studies have shown that TBPM and EBPM performance depends on shared and distinct cognitive abilities that rely on specific brain regions17. The caudate nucleus, which provide an early analysis of the affective properties of the stimuli in a PM task, is activated by aversive stimuli44. Performance on EBPM tasks is largely dependent on the cognitive functions of working memory and executive function, which the caudate nucleus is critically involved in45,46,47,48,49. Therefore, these findings further suggest that altered cooperation of the CA23DG subregion with the caudate nucleus may be a key factor in EBPM under chronic stress.
In chronic stress, the FC of the SUBC with the left inferior parietal gyrus was negatively correlated with both EBPM and TBPM scores. This result suggests that the deactication of the FC of the SUBC and the inferior parietal gyrus under chronic stress related to PM impairment. PM success is linked to the anterior prefrontal cortex, parietal lobe, and hippocampus regions50,51. Previous studies have confirmed that the parietal cortex is a shared substrate of EBPM and TBPM that maintains future intention17. Other studies found that the parietal lobe plays an important role in attention and memory retrieval, which, in turn, play an important role in PM52,53. Furthermore, deactivation of inferior parietal gyrus induced by medicine was found to increase failures of PM54. To summarize, the altered FC of the SUBC and left inferior parietal gyrus in our study may reflect the underlying neurological mechanism whereby chronic stress impairs PM.
This study has two limitations: First, our study had a relatively small sample size. Further studies with larger sample sizes with a more diverse age range are needed. Second, we did not include stress-sensitive psychophysiological measurements, such as cortisol and heart rate. In future studies, the stress level of participants will be measured in different ways, including psychophysiological assessment and self-assessment questionnaire.
To our knowledge, this is the first study to explore the chronic stress-induced specific activation pattern of hippocampal subregions related to impairment of PM. The results suggest that the activation of the FC of SUBC and CA23DG under chronic stress related to impairment of EBPM, while the deactivation of the FC of SUBC is related to both EBPM and TBPM. These findings suggest functional changes in hippocampal subregion networks as a mechanism underlying the impairment of PM in chronic stress.
Methods
Participants
One hundred and five freshmen were recruited from South China Normal University. All participants were right-handed and were selected according to the following criteria: (1) age between 19 and 21 years old, (2) moderate stress as indicated by a the Student-Life Stress Inventory (SLSI) score of >79 and <153, (3) more than 12 years of education, (3) no history of neurological, or psychiatric disorder or head injury, and (4) no history of alcohol/drug dependence. Finally, 25 students (13 male, 12 female) with moderate stress scores were selected as participants. Data were collected at two time points from the participants in May, 2016 (one month prior to examine week; baseline) and June, 2016 (examine week; chronic stress). Informed written consent was obtained from all subjects who participated in the current study. The study was approved by the Research Ethics Board of the South China Normal University. The experiments were performed in accordance with the approved guidelines.
Neuropsychological assessment
The SLSI is a self-administered 51-item Likert-type questionnaire that requires participants to rate their life stress from 1 to 5 (1 = never, 2 = seldom, 3 = occasionally, 4 = often, and 5 = most of the time). The SLSI consists of two parts: stressors and reactions to stressors. The stressors measurement consists of five sections frustrations, conflicts, pressures, changes, and self-imposed stressors while reactions to stressors includes physiological, emotional, behavioral, and cognitive sections. A greater level of stress is indicated by higher scores on stressors and reaction to stress. The SLSI has been shown to have satisfactory reliability and high internal consistency (Cronbach’s α = 0.92)55.
Estimated IQ was assessed with the Wechsler Adult Intelligence Scale-Revised (WAIS-R), and subjects with a total score of less than 80 were excluded56. WAIS-R was performed by experienced clinical psychologists.
Prospective memory assessment
PM performance was measured through a classical dual-task paradigm15 including an ongoing task and a prospective memory task. Ongoing tasks are same in EBPM and TBPM, but the PM task is different. The PM task was divided into two categories: a time-based prospective memory task (TBPM task) and an event-based prospective memory task (EBPM task).
Ongoing-task
For the ongoing task, four Chinese words were displayed in the center of a screen (e.g., 安居乐业). The subject was asked to judge whether the four words formed an idiom. Subjects were instructed to press the “F” key if the four words did not form an idiom (e.g., 直接选举) and the “J” key if they did (e.g., 大材小用).
PM task
In the EBPM task, subjects were instructed to press the space bar if they detected the name of an animal (target event) within the four words (e.g. 如鱼得水). There were five target words (e.g., 虎,马,鼠,鱼,鸡), which appeared at approximately one minute intervals. Non-animal words, such as colors and numbers were used as interference. The participants received one point for each correct response to a target event (total of five target events). Therefore, scores ranged from 0 to 5.
In the TBPM task, a standard clock was placed in the lower left corner of the screen. Subjects were instructed to monitor time throughout the trial and to press the spacebar after one minute. The task lasted for approximately 5 minutes. The participants received one point for each correct response after a full minute. Again, scores ranged from 0–5.
fMRI data acquisition
fMRI data were collected by a clinically approved Siemens Magnetom Avanto 3.0 T (Simens Medical Solutions, Erlangen, Germany) using the Siemens 12-channel receive-only head coil. The imaging sessions included structural T1, a resting condition FC, and two task-related functional acquisitions. Sessions were conducted on the same day and the Siemens Auto Align scout protocol was used to minimize variations in head positioning. For structural analysis, a T1 high-resolution anatomical sequence, 3D MPRAGE (magnetization prepared rapid gradient echo) was performed with the following scan parameters: repetition time (TR) = 2.4 s, echo time (TE) = 3.62 ms, 160 sagittal slices with no gap, field-of-view (FoV) = 234 mm, flip angle (FA) = 8°, in-plane resolution = 1.2 × 1.2 mm² and slice thickness = 1.2 mm. During resting-condition fMRI acquisition, a gradient T2 weighted echo-planar imaging (EPIs) protocol was used and participants were instructed to keep their eyes closed and to think about nothing in particular. The imaging parameters were: 100 volumes, TR = 3 s, TE = 50 ms, FA = 90°, in-plane resolution = 3.4 × 3.4 mm², 30 interleaved slices, slice thickness = 5 mm, imaging matrix of 64 × 64 and FoV = 220 mm. fMRI was acquired using: TR = 2 s, TE = ms, FA = 90°, in-plane resolution and slice thickness = 3.3 mm, 38 ascending interleaved axial slices with no gap and FoV = 212 mm. The functional paradigm protocol was previously described and the paradigm was presented using the fully integrated fMRI system IFIS-SA57.
Image pre-processing
Prior to data processing and analysis, all images were visually inspected to confirm the absence of head motion and brain lesions. Pre-processing was conducted with Statistical Parametric Mapping (SPM12, http://www.fil.ion.ucl.ac.uk/spm), Data Processing Assistant for Resting-Condition fMRI (DPARSF 4.3 Advanced Edition), Resting-Condition fMRI Data Analysis Toolkit (REST 1.8, http://www.restfmri.net) and MATLAB 8.30. The first 10 volumes of the scanning session were discarded to achieve signal stabilization and allow participants to adjust to the scanner noise. Scans with head motion of more than 3 mm maximum displacement in x, y, or z direction or 3° of any angular motion were excluded from analysis. The remaining images were spatially normalized to the standard MMI (Montreal Neurological Institute) echo-planar imaging template, resampled to 3 × 3 × 3 mm³ cubic voxels, and smoothed with a Gaussian kernel 6 × 6 × 6 mm³. To further reduce the effects of confounding factors, the white matter (WM) signal, the cerebrospinal fluid (CSF) signal and six motion parameters were regressed out from the data. Finally, the images were temporally band-pass filtered (0.01–0.08 Hz) and liner trends were removed.
Data from four subjects with poor quality images and head movement more than 3 mm or 3 degree in any direction were excluded from analysis. The final analysis included data from 21 participants.
FC analysis
The Anatomy toolbox in SPM12 (http://www.fz-juelich.de/inm/inm-1/DE/Forschung/docs/SPMAnatomyToolbox/SPMAnatomyToolbox_node.html) was used to identify the three hippocampal subregions (CA1, CA23DG, SUBC) (Fig. 6). Regions of interest were defined according to previous studies58,59. For each subject, a voxel-wise-based FC was computed separately for each hippocampal subregion using the Data Processing Assistant for Resting-State fMRI (dparsf, http://www.rest.restfmri.net). Cross-correlation values between each subregion and the rest of the brain were then calculated. A Fisher transformation was applied to improve the normality of the correlation coefficient60.

Location of hippocampal subregions. Yellow: CA1 (Cornu ammonis1); Red: CA23DG (Cornu ammonis 2, 3 and dentate gyrus); Blue: SUBC (subicular complex).
Statistical analysis
A within group analysis was performed to explore differences in the spatial maps of FC in baseline and chronic stress using one-sample t-tests. Data were corrected for false discovery rate (FDR), and the statistical threshold was set at p < 0.0001. To avoid potential interpretational confounds of negative connectivity caused by correction for the global signal, only positive functional connectivity was examined61.
A between-group analysis was performed to detect changes in PM (TBPM and EBPM) and FC in different hippocampal subregions between chronic stress and baseline (CA1, CA23DG, and SUBC subregion network analysis were performed separately) using paired t tests. Statistical significance was set at p < 0.05 using AlphaSim correction. For measuring extent which chronic stress impact, a between-group analysis was performed to compare the difference of TBPM and EBPM using two-sample t-test.
The relationships between FC of hippocampal subregions and SLSI and PM scores were explored. The mean FC strengths of the clusters showing significant between-group differences were extracted in chronic stress. Spearman’s correlation analysis was performed to investigate the relationship between PM and the FC of hippocampal subregions. A partial correlation analysis was performed to explore the relationship between the FC of hippocampal subregions and SLSI scores, controlling for covariates like gender.
We further explored whether the FC of hippocampal subregions moderates the relationship between the stress level and PM. The SPSS PROCESS macro program, designed by Hayes62, was used to measure the mediating or moderating effect. Within PROCESS, model 1 was selected and the confidence interval was set to 95%. In the moderation models, the SLSI scores were entered as the predictor (X), PM performance as the outcome (Y), and mean FC of hippocampal subregions as the moderator (M). All statistical tests were evaluated at the p < 0.05 significance level and constituted two-tailed tests.
References
Caswell, L. W. et al. Negative associations of chronic stress and cognitive performance in older adult spouse caregivers. Experimental aging research 29, 303–318, https://doi.org/10.1080/03610730303721 (2003).
Bergdahl, J., Larsson, A., Nilsson, L. G., Ahlström, K. R. & Nyberg, L. Treatment of chronic stress in employees: subjective, cognitive and neural correlates. Scand J Psychol 46, 395–402 (2005).
Marin, M.-F. et al. Chronic stress, cognitive functioning and mental health. Neurobiology of Learning and Memory 96, 583–595, https://doi.org/10.1016/j.nlm.2011.02.016 (2011).
Lena, O., Steven, N., Jan, B., Lisbeth, S. B. & Anna, S. N. Cognitive function in outpatients with perceived chronic stress. Scandinavian Journal of Work Environment & Health 33, 223–232 (2007).
Tielbeek, J. J. et al. The impact of chronic stress during adolescence on the development of aggressive behavior: A systematic review on the role of the dopaminergic system in rodents. Neuroscience and biobehavioral reviews 91, 187–197, https://doi.org/10.1016/j.neubiorev.2016.10.009 (2018).
Glienke, K. & Piefke, M. Acute social stress before the planning phase improves memory performance in a complex real life-related prospective memory task. Neurobiology of Learning & Memory 133, 171 (2016).
Ihle, A., Schnitzspahn, K., Rendell, P. G., Luong, C. & Kliegel, M. Age benefits in everyday prospective memory: the influence of personal task importance, use of reminders and everyday stress. Neuropsychology, development, and cognition. Section B, Aging, neuropsychology and cognition 19, 84–101, https://doi.org/10.1080/13825585.2011.629288 (2012).
Nater, U. et al. Psychosocial stress enhances time-based prospective memory in healthy young men. Neurobiol Learn Mem 86, 344–348 (2006).
Peavy, G. M. et al. Effects of chronic stress on memory decline in cognitively normal and mildly impaired older adults. American Journal of Psychiatry 166, 1384 (2009).
Ferbinteanu, J. & Shapiro, M. L. Prospective and Retrospective Memory Coding in the Hippocampus. Neuron 40, 1227–1239 (2003).
Shaozheng, Q. et al. Acute psychological stress reduces working memory-related activity in the dorsolateral prefrontal cortex. Biological Psychiatry 66, 25–32 (2009).
Marle, H. J. F. V., Hermans, E. J., Qin, S. & Fernández, G. Enhanced resting-state connectivity of amygdala in the immediate aftermath of acute psychological stress ✩. Neuroimage 53, 348–354 (2010).
Henry, J. D., MacLeod, M. S., Phillips, L. H. & Crawford, J. R. A Meta-Analytic Review of Prospective Memory and Aging. Psychol & Aging 19, 27 (2004).
Bisiacchi, P. S., Cona, G., Schiff, S. & Basso, D. Modulation of a fronto-parietal network in event-based prospective memory: An rTMS study. Neuropsychologia 49, 2225–2232 (2011).
Einstein, G. O. & McDaniel, M. A. Normal Aging and Prospective Memory. J Exp Psychol Learn Mem Cogn 16, 717–726 (1990).
Einstein, G. O., Mcdaniel, M. A., Richardson, S. L., Guynn, M. J. & Cunfer, A. R. Aging and prospective memory: examining the influences of self-initiated retrieval processes. Journal of Experimental Psychology Learning Memory & Cognition 21, 996 (1995).
Julie, G. et al. How do we process event-based and time-based intentions in the brain? an fMRI study of prospective memory in healthy individuals. Human Brain Mapping 35, 3066–3082 (2014).
Jarrard, L. E. On the role of the hippocampus in learning and memory in the rat. Behav Neural Biol 60, 9–26 (1993).
Kim, J. J., Eun Young, S. & Kosten, T. A. Stress effects in the hippocampus: synaptic plasticity and memory. Stress-the International Journal on the Biology of Stress 9, 1–11 (2006).
Mcewen, B. S. & Magarinos, A. M. Stress effects on morphology and function of the hippocampus. Annals of the New York Academy of Sciences 821, 271–284 (2010).
Lee, V., Mackenzie, G., Hooper, A. & Maguire, J. Reduced tonic inhibition in the dentate gyrus contributes to chronic stress-induced impairments in learning and memory. Hippocampus 26, 1276–1290 (2016).
Mckittrick, C. R. et al. Chronic social stress reduces dendritic arbors in CA3 of hippocampus and decreases binding to serotonin transporter sites. Synapse 36, 85–94 (2015).
Vargha-Khadem, F. et al. Differential effects of early hippocampal pathology on episodic and semantic memory. 277, 376–380 (1997).
Magalhaes, R. et al. The dynamics of stress: a longitudinal MRI study of rat brain structure and connectome. Molecular psychiatry 23, 1998–2006, https://doi.org/10.1038/mp.2017.244 (2018).
Magalhaes, R. et al. A Resting-State Functional MR Imaging and Spectroscopy Study of the Dorsal Hippocampus in the Chronic Unpredictable Stress Rat Model. The Journal of neuroscience: the official journal of the Society for Neuroscience 39, 3640–3650, https://doi.org/10.1523/JNEUROSCI.2192-18.2019 (2019).
McEwen, B. S. & Magarinos, A. M. Stress and hippocampal plasticity: implications for the pathophysiology of affective disorders. Human psychopharmacology 16, S7–S19, https://doi.org/10.1002/hup.266 (2001).
McEwen, B. S. Protective and damaging effects of stress mediators. The New England journal of medicine 338, 171–179, https://doi.org/10.1056/NEJM199801153380307 (1998).
Amunts, K. et al. Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: intersubject variability and probability maps. Anatomy and embryology 210, 343–352, https://doi.org/10.1007/s00429-005-0025-5 (2005).
Awipi, T. & Davachi, L. Content-specific source encoding in the human medial temporal lobe. Journal of Experimental Psychology Learning Memory & Cognition 34, 769 (2008).
Lee, A., Scahill, V. & Graham, K. S. Activating the medial temporal lobe during oddity judgment for faces and scenes. Cerebral Cortex 18, 683 (2008).
Duarte, A., Henson, R. N. & Graham, K. S. Stimulus content and the neural correlates of source memory. Brain research 1373, 110 (2011).
Carr, V. A., Rissman, J. & Wagner, A. D. Imaging the human medial temporal lobe with high-resolution fMRI. Neuron 65, 298–308, https://doi.org/10.1016/j.neuron.2009.12.022 (2010).
Suthana, N., Ekstrom, A., Moshirvaziri, S., Knowlton, B. & Bookheimer, S. Dissociations within human hippocampal subregions during encoding and retrieval of spatial information. Hippocampus 21, 694–701, https://doi.org/10.1002/hipo.20833 (2011).
Hawley, D. F., Morch, K., Christie, B. R. & Leasure, J. L. Differential response of hippocampal subregions to stress and learning. PloS one 7, e53126, https://doi.org/10.1371/journal.pone.0053126 (2012).
Pavlides, C., Nivon, L. G. & McEwen, B. S. Effects of chronic stress on hippocampal long-term potentiation. Hippocampus 12, 245–257, https://doi.org/10.1002/hipo.1116 (2002).
Xiang, Z., Tan, S., Kang, Q., Zhang, B. & Zhu, L. Longitudinal Effects of Examination Stress on Psychological Well-Being and a Possible Mediating Role of Self-Esteem in Chinese High School Students. Journal of Happiness Studies 20, 283–305 (2019).
Duan, H. et al. Chronic stress exposure decreases the cortisol awakening response in healthy young men. Stress 16, 630–637, https://doi.org/10.3109/10253890.2013.840579 (2013).
Bai, F. et al. Aberrant hippocampal subregion networks associated with the classifications of aMCI subjects: a longitudinal resting-state study. PLoS One 6, e29288, https://doi.org/10.1371/journal.pone.0029288 (2011).
Wang, Z. et al. Altered functional connectivity networks of hippocampal subregions in remitted late-onset depression: a longitudinal resting-state study. Neuroscience bulletin 31, 13–21, https://doi.org/10.1007/s12264-014-1489-1 (2015).
Ellis, H. C. & Ashbrook, P. W. Resource allocation model of the effects of depressed mood states on memory. Affect, cognition, and social behavior, 25–43 (1988).
Einstein, G. Retrieval processes in prospective memory: Theoretical approaches and some new empirical findings. Prospective Memory Theory & Applications, 115–141 (1996).
Lupien, S. & Lepage, M. Stress, memory, and the hippocampus: can’t live with it, can’t live without it. Behav. Brain Res. 127, 137–158 (2001).
Magarinos, A. M., Verdugo, J. M. & McEwen, B. S. Chronic stress alters synaptic terminal structure in hippocampus. Proc Natl Acad Sci USA 94, 14002–14008, https://doi.org/10.1073/pnas.94.25.14002 (1997).
Gilbert, S. et al. Effects of Aversive Stimuli on Prospective Memory. An Event-Related fMRI Study. PLoS One 6, e26290, https://doi.org/10.1371/journal.pone.0026290 (2011).
Melrose, R. J., Poulin, R. M. & Stern, C. E. An fMRI investigation of the role of the basal ganglia in reasoning. Brain research 1142, 146–158, https://doi.org/10.1016/j.brainres.2007.01.060 (2007).
Jokinen, P. et al. Impaired cognitive performance in Parkinson’s disease is related to caudate dopaminergic hypofunction and hippocampal atrophy. Parkinsonism & Related Disorders 15, 88–93 (2009).
Mahy, C. E. V. & Moses, L. J. Executive functioning and prospective memory in young children. Cognitive Development 26, 269–281 (2011).
Logie, R., Maylor, E., Sala, S. D. & Smith, G. Working memory in event- and time‐based prospective memory tasks: Effects of secondary demand and age. European Journal of Cognitive Psychology 16, 441–456 (2004).
Rinne, J. O. et al. Cognitive impairment and the brain dopaminergic system in Parkinson disease: [18F]fluorodopa positron emission tomographic study. Archives of neurology 57, 470–475, https://doi.org/10.1001/archneur.57.4.470 (2000).
Burgess, P. W., Gonen-Yaacovi, G. & Volle, E. Functional neuroimaging studies of prospective memory: what have we learnt so far? Neuropsychologia 49, 2246–2257, https://doi.org/10.1016/j.neuropsychologia.2011.02.014 (2011).
Reynolds, J. R., West, R. & Braver, T. J. C. C. Distinct neural circuits support transient and sustained processes in prospective memory and working memory. 19, 1208–1221 (2008).
Reynolds, J. R., West, R. & Braver, T. Distinct neural circuits support transient and sustained processes in prospective memory and working memory. Cereb Cortex 19, 1208–1221, https://doi.org/10.1093/cercor/bhn164 (2009).
Wagner, A. D., Shannon, B. J., Kahn, I. & Buckner, R. L. Parietal lobe contributions to episodic memory retrieval. Trends in Cognitive Sciences 9, 445–453 (2005).
Ramaekers, J. G., Kuypers, K. P., Wingen, M., Heinecke, A. & Formisano, E. Involvement of inferior parietal lobules in prospective memory impairment during acute MDMA (ecstasy) intoxication: an event-related fMRI study. Neuropsychopharmacology Official Publication of the American College of Neuropsychopharmacology 34, 1641–1648 (2009).
Gadzella, B. M. & Others, A. Student-Life Stress Inventory. College Students 12 (1991).
Wechsler, D. WAIS-R manual: Wechsler adult intelligence scale-revised (Psychological Corporation, 1981).
Soares, J. M. et al. Stress Impact on Resting State Brain Networks. Plos One 8, e66500 (2013).
Umeda, S., Nagumo, Y. & Kato, M. Dissociative contributions of medial temporal and frontal regions to prospective remembering. Reviews in the neurosciences 17, 267–278 (2006).
Zeineh, M. M., Engel, S. A., Thompson, P. M. & Bookheimer, S. Y. Dynamics of the hippocampus during encoding and retrieval of face-name pairs. Science 299, 577–580, https://doi.org/10.1126/science.1077775 (2003).
Lowe, M., Mock, B. & Sorenson, J. Functional connectivity in single and multislice echoplanar imaging using resting-state fluctuations. Neuroimage 7, 119–132 (1998).
Etkin, A., Prater, K., Schatzberg, A., Menon, V. & Greicius, M. Disrupted amygdalar subregion functional connectivity and evidence of a compensatory network in generalized anxiety disorder. Arch. Gen. Psychiatry 66, 1361–1372 (2009).
Hayes, A. J. J. O. E. M. Introduction to mediation, moderation, and conditional process analysis. 51, 335–337 (2013).
Acknowledgements
This work was supported by National Natural Science Foundation of China (31871113, 31671135 and 31920103009), the project of Shenzhen Science and Technology Innovation Committee (CYJ20160429185235132, CYJ20160427192001852), and Sanming Project of Medicine in Shenzhen (SZSM201612079, SZSM201512009), and Guangdong Key Project in development of new tools for diagnosis and treatment of autism (2018B030335001).
Author information
Authors and Affiliations
Contributions
J.C., Z.W. and Z.P. designed the study. J.C., L.J., C.X., D.D. and J.L. acquired the data from papers. J.C., Z.W., H.H., G.W. and Z.P. analyzed the data. J.C., Z.W., G.W. and Z.P. wrote the article and all other authors reviewed the article. All authors approved the publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, J., Wei, Z., Han, H. et al. An Effect of Chronic Stress on Prospective Memory via Alteration of Resting-State Hippocampal Subregion Functional Connectivity. Sci Rep 9, 19698 (2019). https://doi.org/10.1038/s41598-019-56111-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-56111-9
This article is cited by
-
The functional and structural changes in the hippocampus of COVID-19 patients
Acta Neurologica Belgica (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
