
Abstract
Honeydew honey is increasingly valued due to its pronounced antibacterial potential; however, the underlying mechanism and compounds responsible for the strong antibacterial activity of honeydew honey are still unknown. The aim of this study was to investigate the inhibition of bacterial growth of 23 honeydew honey samples. Activity of bee-derived glucose oxidase (GOX) enzyme, the content of defensin-1 (Def-1) and hydrogen peroxide (H2O2), and total polyphenol content were determined in the 23 honey samples. Our results demonstrated that antibacterial activity of honeydew honey was equivalent to medical-grade manuka and kanuka honey and was abolished by catalase. Although H2O2 is an important factor in the inhibition of bacterial growth, polyphenolic compounds and their interaction with H2O2 are the key factors responsible for high antibacterial activity of honeydew honey. In addition, our results indicated that the antibacterial activity of honeydew honey is not dependent on GOX-mediated production of H2O2 or the presence of Def-1.
Similar content being viewed by others


Introduction
Honeydew honey is produced by bees from sugar-rich secretions of trees and plants or exudates of plant-sucking insects (Hemiptera). In addition to its attractive sensorial characteristics, which differ from blossom honey, honeydew honey is valued due to its pronounced antibacterial potential1. The botanical origin of honeydew honey is from conifers or deciduous trees. A fir honeydew honey is one of the most known type of conifer honeydew honey in Europe2.
Honeydew honey possesses strong biological properties including antibacterial1,3,4, antibiofilm5, anti-inflammatory6, antioxidant7, as well as wound healing activity8. The antibacterial and antibiofilm activity of honeydew honey is comparable to manuka honey, which is currently used as medical-grade honey in clinical applications. Furthermore, a Slovak fir honeydew honey has been successfully clinically tested for the treatment of infected gluteo-femoral fistulas9, lower leg ulcers10, contact lens-induced corneal ulcers11, and as a prophylactic agent for endophthalmitis12.
The mechanism underlying the antibacterial and antibiofilm activity of honeydew honey has not been fully elucidated. It has been proposed that hydrogen peroxide (H2O2), produced by enzymatic conversion of glucose in diluted honey, and the bee-derived antibacterial peptide defensin-1 (Def-1) are considered the major components responsible for the antibacterial activity of natural honey, with the exception of manuka honey13. Although the levels of H2O2 in honey may differ between different types of honey, regardless of its botanical and geographical origin14, it has been suggested that honeydew honey produces higher amounts of H2O2 compared with nectar honey15. Honeydew honey is known for a high content of phenolic acids and flavonoids possessing antioxidant and pro-oxidant properties16. Thus, polyphenols, at concentration found in honeydew honey, could be involved in the generation of substantial amounts of H2O217,18.
In the present study, we aimed to (i) determine the antibacterial activity of Slovak honeydew honey samples against the two most frequently isolated wound pathogens, (ii) investigate the mechanism of how honeydew honey inhibits bacterial growth, and (iii) elucidate the contribution of Def-1 and glucose oxidase (GOX)-mediated H2O2 antibacterial activity to the overall antibacterial activity of honeydew honey.
Results
Antibacterial activity of honeydew honey
Minimum inhibitory concentration (MIC) values of 23 honeydew honey samples against the tested bacterial strains are shown in Fig. 1. Honey samples were more effective against of the Gram-positive S. aureus compared with the Gram-negative P. aeruginosa. For S. aureus, the MIC values of the honey samples ranged from 2.0 to 12.5%. The MICs of honey against P. aeruginosa ranged from 7.5 to 15%. Honey samples # 4–8 possessed the strongest antibacterial activity against the tested bacteria.

Antibacterial activity of honeydew honey samples (n = 23) and medical-grade manuka and kanuka honey against Staphylococcus aureus and Pseudomonas aeruginosa isolates. Activity was determined with a minimum inhibitory concentration (MIC) assay. The MIC was defined as the lowest concentration of honey solution (%) inhibiting bacterial growth. K, kanuka honey; M, manuka honey.
Besides MIC determination, a minimum bactericidal concentration (MBC) of honey samples was evaluated. The values of MIC and MBC were comparable and the overall inhibitory and bactericidal profiles of honey samples were identical (Fig. S1).
Kanuka and manuka honey, which are both used as medical-grade honeys, exhibited comparable or less effective antibacterial activity compared with the honeydew honey samples. Similar to honeydew honey, both types of New Zealand honey were more effective against S. aureus than P. aeruginosa.
Comparison of GOX activity and H2O2 production in honey samples
The GOX activity determined in 20% honey solutions ranged from 21 to 50 mU/ml in all honeydew honey samples, with an average value of 35.7 mU/ml (Fig. 2A). The lowest enzymatic activity was monitored in both types of medical honey (<20 mU/ml), suggesting that methylglyoxal, present in both samples, may have structurally modified the GOX enzyme. The concentration of H2O2 in honeydew honey samples was determined in 40% honey solutions and ranged from 0.3 to 3.4 mM after 24 h incubation at 37 °C (Fig. 2B). The average value of H2O2 concentration was 1.8 mM. Two honeydew honey samples (#16 and #21) showed a weak production of H2O2, with values below 1 mM. As expected, kanuka and manuka honey accumulated the lowest levels of H2O2. No significant correlations were found between the values of GOX activity and H2O2 concentration among the honeydew honey samples (r = 0.26, P = 0.22). Therefore, we assumed that other compound(s) of botanical origin present in honeydew honey participate in the production of H2O2. Potential candidates responsible for the elevated levels of H2O2 include phenolic compounds.

Glucose oxidase (GOX) activity and hydrogen peroxide (H2O2) production in honeydew honey samples (n = 23) and medical-grade manuka and kanuka honey. (A) GOX activity was determined in 20% (w/v) honey solutions with a GOX assay kit. (B) H2O2 production was measured in 40% (w/v) honey samples with a modified GOX assay kit. The data are expressed as the mean values with standard error of the mean (SEM).
Total polyphenol and Def-1 content in honey samples
Def-1 is a key, but quantitatively variable, antibacterial factor in honey. Def-1 content was examined in the 2.5, 5 and 10% honey samples in PBS, using the newly developed competitive ELISA and the results were expressed as a percentage of total protein (Fig. 3A). The amount of Def-1 in each honey sample, including kanuka and manuka honey, is shown in Fig. 3A. The two honeydew honey samples #9 and #10 contained the highest amount of Def-1. Similar to the H2O2 production capacity, kanuka and Manuka honey had the lowest content of Def-1 among all the tested samples. Overall content of Def-1 in the honeydew honey samples did not correlate with their antibacterial activity against S. aureus (r = 0.17, P = 0.45). Def-1, at the concentration observed in the honeydew honey samples, did not exert its antibacterial activity at the MICs of the honeydew honey samples. The obtained data also suggest that the antibacterial action of kanuka and manuka honey dependeds on methylglyoxal or other unknown botanical compounds rather than bee-derived antibacterial factors.

Content of defensin-1 (Def-1) and total polyphenol in honeydew honey samples (n = 23) and medical-grade manuka and kanuka honey. (A) Def-1 content was quantified in honey samples using indirect competitive enzyme-linked immunosorbent assay and the results were expressed as a percentage of total protein. (B) Total polyphenol (TP) content was determined with a Folin Ciocalteu Phenolic Content Quantification Assay Kit in 20% (w/v) honeydew honey solutions. Gallic acid (GAE) was used as the reference standard compound and results were expressed as GAE equivalents (mg/ml). Data are expressed as the mean values with standard error of the mean (SEM).
A direct antibacterial activity of phenolic compounds has not been shown since their concentrations in honey are too low to determine overall activity. However, phenolic compounds may act in synergy via pro-oxidative action by generating elevated levels of H2O2 in honey, which mediates the inhibition of bacterial growth as a result of oxidative stress. Total phenolic content determined in the 20% (w/v) honeydew honey solutions is shown in Fig. 3B.
Contribution of bee-derived antibacterial compounds to the antibacterial activity of honeydew honey
To investigate the contribution of bee-derived antibacterial compounds to the overall antibacterial activity of honeydew honey, samples were treated with a proteolytic enzyme, proteinase K. Upon proteinase K treatment, all proteinous content in the 50% (w/v) honey solutions was completely digested (data not shown). Untreated and proteinase K-treated honeydew samples were subjected to immunoblot analysis of GOX and Def-1. As expected, GOX and Def-1 were present in all untreated samples (Fig. 4). On the other hand, no immunoreactive bands for GOX and Def-1 were observed in treated samples, confirming the proteolytic effectiveness of proteinase K.

Detection of glucose oxidase (GOX) and defensin-1 (Def-1) in honeydew honey samples (n = 23) and medical-grade manuka and kanuka honey following proteinase K treatment by immunoblotting. Aliquots (15 μl) of 50% (w/v) honey solution with or without proteinase K were resolved by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and 16.5% Tricine-SDS-PAGE. After semi-dry blotting procedure, the blocked membrane was incubated overnight with a rabbit polyclonal antibody against honeybee GOX or Def-1 (1:2000). Shown are cropped blots. (The blots with indicated cropping lines are shown in Supplementary Fig. S2). Immunoreactive bands were detected in solution containing dissolved SigmaFast 3,3-diaminobenzidine tablets (GOX) or detected using an enhanced chemiluminescence Immobilon Western kit (Def-1).
MIC values were determined in untreated and proteinase K-treated honey samples. No significant changes were identified in the MIC values between untreated and proteinase-treated honey against both tested bacteria (Fig. 5), suggesting that bee-derived components, including GOX-mediated H2O2, are not responsible for the pronounced antibacterial activity of the honeydews honey. Although H2O2 is a major antibacterial compound of honeydew honey, its source might be solely of botanical origin.

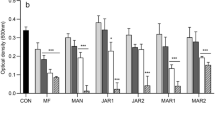
Antibacterial activity of honeydew honey samples (n = 23) and medical-grade manuka and kanuka honey following catalase and proteinase K treatment against (A) Staphylococcus aureus and (B) Pseudomonas aeruginosa isolates. The 50% (w/v) honey solutions were treated with catalase (2000–5000 U/mg protein) at a final concentration ranging from 1000 to 2500 U/ml at room temperature for 2 h or proteinase K (30 U/mg) at a final concentration of 50 μg/ml at 37 °C for 30 min. The antibacterial activity was determined with a minimum inhibitory concentration (MIC) assay. The MIC was defined as the lowest concentration of honey solution (%) inhibiting bacterial growth. K, kanuka honey; M, manuka honey.
Role of H2O2 in honeydew honey antibacterial activity
To determine the role of H2O2 in honeydew honey antibacterial activity, 50% (w/v) honey solutions were treated with catalase for 2 h and MICs were determined. The MIC values of honey samples with or without catalase treatment against both tested bacteria are shown in Fig. 5. Catalase-treated samples had lower antibacterial activity with average MIC values of 30%. Furthermore, in seven samples, the MIC values were greater than 30% following catalase treatment. The effect of catalase on the MIC values in all honeydew honey samples was highly significant (NumDF = 2, DenDF = 268.43, F = 2136.38, P < 0.001) and predicted values were three times larger (Fig. 6) compared with untreated and protease K-treated samples. Interestingly, statistically significant changes (NumDF = 1, DenDF = 268.27, F = 8.52, P = 0.004)) were observed following catalase treatment between S. aureus and P. aeruginosa (Fig. 6). S. aureus was more resistant to the honey solution after the removal of H2O2 compared with P. aeruginosa.

Graphical representation of the results of the general linear mixed model with response variable treatment type, bacterial species, and their two-way interaction. (Pseudomonas aeruginosa – red and Staphylococcus aureus – blue).
Interestingly, the MIC of kanuka honey following catalase treatment was also changed, suggesting that the antibacterial activity of kanuka honey is, at least in part, due to the generation of H2O2. As expected, no remarkable changes in antibacterial activity were observed in manuka honey. Surprisingly, the antibacterial activity of manuka honey against P. aeruginosa following catalase treatment was even higher than without enzyme treatment.
Despite the fact that the antibacterial activities of honeydew honey samples were comparable, H2O2 production was not uniform. Statistical analysis did not show any correlation between antibacterial activity and the level of H2O2 in the different honey samples (r = −0.38, P = 0.08 and r = −0.03, P = 0.89). The honey samples with the highest level of H2O2 were not the most active.
Discussion
Honey is increasingly valued for its antibacterial and wound healing activity and effectiveness as a treatment of chronic wounds and burns. However, every natural honey exhibits a certain antibacterial activity and only those with a strong and relatively stable antibacterial activity have been selected for medical and clinical use. In this study, we focused on honeydew honey as a source of potential medical-grade honey to elucidate its antibacterial activity and to investigate the role of bee-derived antibacterial components on its overall antibacterial activity.
A plethora of studies has demonstrated the important role of Def-1 and H2O2 in the antibacterial activity of honey.
Def-1 is an antibacterial peptide belonging to the insect defensin group that is composed of 51 amino acids with a molecular weight of 5.52 kDa. Bee Def-1 is effective against Gram-positive bacteria19,20,21; however, some studies using recombinant Def-1 have also reported its activity against Gram-negative bacteria including P. aeruginosa and Salmonella choleraesuis22,23. According to our recent study24, Def-1 is a common but quantitatively variable component in honey and its levels in different blossom honey samples are correlated with antibacterial activity against S. aureus. Recently, Def-1 has also been shown to be an immunomodualtor in wound healing where it positively contributes to cutaneous wound closure25. In the present study, the amount of Def-1 in honeydew honey did not correlate with the overall antibacterial activity of samples against S. aureus, suggesting that its contribution at low dilutions (5–10%) is negligible.
The most important antibacterial factor in honey is H2O2, which is produced by GOX-mediated conversion of glucose to gluconic acid under aerobic conditions in diluted honey26. Therefore, GOX plays an important role in the generation of H2O215. Although H2O2 at concentrations found in honey does not kill bacteria, it is able to interact with bacterial cell proliferative signals, and thus affects bacterial growth even at high dilutions of honey27. Some researchers have suggested that the levels of H2O2 in honey may differ between types of honey regardless of botanical and geographical origin14. A few studies have attempted to determine the concentration of H2O2 in different honey samples and to evaluate its effect on the overall antibacterial activity15,27,28,29,30. Many studies have shown a greater correlation between antibacterial activity against S. aureus and the amount of H2O2 in honey samples. In the present study, there was no obvious relationship between the level of H2O2 and the antibacterial activity of the honeydew honey samples; the honey samples with the highest level of H2O2 were not the most active. Antibacterial activity was comparable among all honeydew honey samples despite variations in the concentration of H2O2. In addition, the removal of H2O2 by catalase did alter the antibacterial activity, as the activity following catalase treatment was comparable among all the honeydew honey samples. Similarly, Roshan and co-workers (2017) found no relationship between the concentration of H2O2 and overall antibacterial activity of eight different Australian honey samples. Researchers have suggested that low levels of H2O2 in different types of honey may be a result of the presence of phytochemicals other than methylglyoxal that might contribute to overall activity30.
We assume that plant-derived polyphenolic compounds, often reported in honey samples, may contribute to or modulate antibacterial activity. In the presence of transition metal ions (Cu and Fe) and peroxides, polyphenols, well-known dietary antioxidants, can act as pro-oxidants by accelerating hydroxyl radical formation and oxidative strand breakage in DNA31. In fact, polyphenols work in two ways to promote antibacterial activity: by directly producing H2O2, and by reducing Fe (III) to Fe (II), which triggers the Fenton reaction to create more potent reactive oxygen species such as hydroxyl radicals. A key factor in determining whether polyphenolic compounds exhibit antioxidative or antibacterial properties is pH value32. In weakly alkaline conditions (pH 7.0–8.0), polyphenols can exhibit pro-oxidative activity and inhibit bacterial growth. In the present study, the determination of honey MIC values and H2O2 concentration was carried out at pH 7.3 and 7.0, respectively. In these experimental conditions, it is obvious that the polyphenols in honeydew honey may act in synergy and inhibit bacterial growth. As mentioned, we measured the accumulation of H2O2 in diluted honey samples, but found no correlation between the concentration of H2O2 and antibacterial activity. It is likely that besides accumulation of H2O2, polyphenol-mediated production of hydroxyl radicals in honeydew honey significantly affects the overall antibacterial activity. Moreover, it has been shown that the chemical interaction of honey polyphenols with H2O2 results in the generation of products responsible for the degradation of bacterial DNA33. Interestingly, H2O2 alone is not able to induce DNA breaks but is somehow involved in this process. Although H2O2 is necessary for accelerating the auto-oxidation of polyphenols via hydrolysis and serves as a source of hydroxyl radicals, polyphenol concentration is a critical factor for antibacterial activity.
Total polyphenol content was reported to be high in other studies of honeydew honey samples7,34,35. In the present study, the total polyphenol content did not differ substantially between the tested honeydew honey samples (with the exception of sample #15), with an average value of 50 mg GAE/100 g of honey. It will be important to determine the critical concentration limit of total polyphenols for their behavior as pro-oxidant agents.
Several studies have attempted to identify biologically-active polyphenols and flavonoids from honeydew honey of different botanical and geographical origin6,36,37. Ferulic acid, quercetin, kaempherol and particular p-coumaric acid were the most abundant polyphenols in honeydew honeys. In a very recent studies38, combination of p-coumaric acid with bacteriocin showed synergistic effects against planktonic cells of food-borne bacteria. Similarly, combination of plant polyphenols and H2O2 was shown to be more effective against bacteria than H2O2 alone and can induce oxidative stress-related responses in bacteria32. It has been hypothesized that antibacterial effect of plant polyphenols is linked to polyphenols-induced H2O2 and an increase of endogenous reactive oxygen species generation39.
Until now, no study had been conducted to determine the role of bee-derived antibacterial compounds in honeydew honey. Here, we showed that the antibacterial activity of honeydew honey is not dependent on GOX-mediated production of H2O2 or the presence of Def-1. This is an interesting observation in the light of current knowledge regarding the antibacterial properties of honey. The antibacterial potential of honeydew honey is therefore not associated with the production capacity of GOX and Def-1 in particular bee colonies, which has a great impact on the biological properties of honey.
In conclusion, the tested honeydew honey samples showed equivalent or, in some cases, higher antibacterial activity compared with medical-grade kanuka and manuka honey. This strong antibacterial activity was abolished by treatment with catalase. Although H2O2 is an important factor in the inhibition of bacterial growth, phytochemicals such as polyphenolic compounds and their interaction with H2O2 are the key factors responsible for the antibacterial activity of honeydew honey. In addition, our results indicate that the antibacterial activity of honeydew honey is not dependent on GOX-mediated production of H2O2 or the presence of Def-1.
Materials and Methods
Honey samples
Honey samples (n = 28) collected in 2016 were received from beekeepers from several regions of Slovakia. The samples were selected according to their electrical conductivity value (Codex Alimentarius, Bogdanov 2002). Five samples were excluded from the study, as they did not meet the criteria for honeydew honey. Overall, 23 honeydew honey samples were used for further analysis. The honey samples were also compared to medical grade honey, Manuka Honey MGO 550 purchased from Manuka Health (New Zealand), and raw kanuka honey (HLKN1, No. 5054), kindly provided by Dr. Shaun Holt (honeylab, New Zealand).
Microorganisms
The antibacterial activity of honey samples was assessed against the isolates Pseudomonas aeruginosa CCM1960 and Staphylococcus aureus CCM4223, obtained from the Department of Medical Microbiology, Slovak Medical University (Bratislava, Slovakia). These bacterial isolates are the two most frequently isolated pathogens from wounds.
Determination of honey antibacterial activity
The antibacterial efficacy of the honey samples was evaluated with a minimum inhibitory concentration (MIC) assay as described by Bucekova et al. (2014). Briefly, overnight bacterial culture was suspended in phosphate-buffered saline (PBS), pH 7.2, and the turbidity of the suspension was adjusted to 108 colony-forming unit (CFU)/ml and diluted with Mueller-Hinton broth (MHB) medium (pH 7.3 ± 0.1) to a final concentration of 106 CFU/ml. Then, 10- μl aliquots of suspension were inoculated into each well of sterile 96-well polystyrene plates (Sarstedt, Germany). The final volume in each well was 100 μl, consisting of 90 μl of sterile medium or diluted honey and 10 μl of bacterial suspension. After 18 h of incubation at 37 °C, bacterial growth inhibition was determined by monitoring the optical density at 490 nm. The MIC was defined as the lowest concentration of honey inhibiting bacterial growth. All tests were performed in triplicate and repeated three times.
Serial dilutions of each honey sample were prepared from a 50% (w/v) honey solution, resulting in final concentrations of 45, 35, 30, 25, 20, 18, 16, 14, 12, 10, 8, 6, and 4%.
Besides MIC determination, MBC was evaluated in honey samples40. The viability of bacteria in wells with no turbidity was determined by spreading 100 µl onto an MHB agar plate and incubating at 37 °C for 24 h. The lowest honey that resulted in no survival of viable bacteria was recorded as MBC.
Enzymatic treatment of honey samples with catalase and proteinase K
The 50% (w/v) honey solutions were treated with catalase (2000–5000 U/mg protein; Sigma-Aldrich, UK) at a final concentration ranging from 1000–2500 U/ml at room temperature for 2 h or proteinase K (30 U/mg; Promega, WI, USA) at a final concentration of 50 μg/ml at 37 °C for 30 min. Catalase- and proteinase K-treated honey samples were then used in the antibacterial assay to determine MIC values against S. aureus and P. aeruginosa.
To determine the effectiveness of both enzymes, the accumulation of H2O2 was measured in catalase-treated samples after 24 h incubation at 37 °C and the presence of GOX and Def-1 in proteinase K-treated samples were determined by immunoblot analysis (see 2.9.).
Determination of GOX activity
The bee-derived GOX activity was determined by with a Megazyme GOX assay kit (Megazyme International Ireland Ltd., Bray, Ireland), which is based on the oxidative catalysis of β-D-glucose to D-glucono-δ-lactone, with the concurrent release of H2O2. The resultant H2O2 reacts with p-hydroxybenzoic acid and 4-aminoantipyrine in the presence of peroxidase to form a quinoneimine dye complex, which has a strong absorbance at 510 nm. The enzyme activity was determined in 20% (w/v) honey solutions in a 96-well microplate according to the manufacturer’s instructions.
Determination of H2O2 concentration
The H2O2 concentration in the honey samples was determined with a Megazyme GOX assay kit (Megazyme International Ireland Ltd), which is based on H2O2 release. For a standard, H2O2 diluted to 9.8–312.5 μM was used. Briefly, 40% (w/v) honey solutions were incubated at 37 °C for 24 h. Each honey sample and standard was tested in duplicate in a 96-well microplate and the absorbance was measured at 510 nm using Synergy HT microplate reader (BioTek Instruments, VT, USA).
Quantification of Def-1
Bee-derived peptide Def-1 was quantified as described by Valachova et al. (2016). Briefly, serial dilutions of each honey sample were prepared from a 50% (w/v) solution of honey, resulting in final concentrations of 2.5, 5, and 10%. An enzyme-linked immunosorbent assay (ELISA) was performed using a rabbit polyclonal anti-honeybee Def-1 antibody raised against a synthetic peptide corresponding to the C-terminus of bee Def-1 (CRKTSFKDLWDKRFG)24. The amount of Def-1 measured was expressed as a percentage of total proteins, which were measured using the Quick Start Bradford Protein Assay (Bio-Rad, CA, USA) according to the manufacturer’s instructions. All measurements were performed in triplicate and repeated three times.
Total phenolic content
Total phenolic content was determined with a Folin Ciocalteu Phenolic Content Quantification Assay Kit (BioQuoChem, Spain) in a 20% (w/v) honey solution in a 96-well microplate according to the manufacturer’s instructions. Gallic (GAE) acid was used as the reference standard and results were expressed as GAE equivalents (mg/ml). Absorbance was measured at 700 nm at 37 °C.
Detection of GOX and Def-1 by immunoblotting
Aliquots (15 μl) of 50% (w/v) honey solution with or without proteinase K were resolved by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and 16.5% Tricine-SDS-PAGE using a Mini-Protean II electrophoresis cell (Bio-Rad). The proteins were transferred onto a 0.22-μm nitrocellulose Advantec membrane (Sigma-Aldrich) in 48 mM Tris, 39 mM glycine and 20% methanol using the semi-dry blotting procedure. The membrane was blocked for 1 h in a Tris-buffered saline-Tween (TBST) buffer (50 mM Tris–HCl, pH 7.5, 200 mM NaCl, and 0.05% Tween 20) containing 5% non-fat dried milk and incubated overnight with a rabbit polyclonal antibody against honeybee GOX or Def-1 (1:2000 in TBST). After washing with TBST, the membranes were incubated for 2 h in blocking buffer containing goat anti-rabbit horseradish peroxidase-linked antibodies (1:2500 in TBST; Promega). Immunoreactive bands were detected in solution containing dissolved SigmaFast 3,3-diaminobenzidine tablets (Sigma-Aldrich) or detected using enhanced chemiluminescence Immobilon Western kit (Millipore, MA, USA).
Statistical analysis
A general linear mixed model was used to analyse the differences in MIC values following treatment with catalase and proteinase K as well as untreated samples. As every sample was tested for each treatment and both P. aeruginosa and S. aureus, the identity of samples was controlled by including a random part of the model with the sample ID. This approach allowed for the exclusion of inter-sample variability. An R programming and statistical environment was used for statistical analyses (www.R-project.org). The general linear mixed model was analysed with the lme4 package41. Correlations among all parameters were tested with the Hmisc package (R package version 4.1-1) using non-parametric Spearmans rank correlations. Correlations with correlation coefficient r > 0.7 or <−0.7 were considered important.
References
Majtan, J., Majtanova, L., Bohova, J. & Majtan, V. Honeydew honey as a potent antibacterial agent in eradication of multi-drug resistant Stenotrophomonas maltophilia isolates from cancer patients. Phytother. Res. 25, 584–587 (2011).
Kus, P. M., Jerkovic, I., Marijanovic, Z. & Tuberoso, C. I. G. Screening of Polish fir honeydew honey using GC/MS, HPLC-DAD, and physical-chemical parameters: benzene derivatives and terpenes as chemical markers. Chem. Biodivers. 14, e1700179 (2017).
Grego, E. et al. Evaluation of antimicrobial activity of Italian honey for wound healing application in veterinary medicine. Schweiz. Arch. Tierheilkd. 158, 521–527 (2016).
Salonen, A., Virjamo, V., Tammela, P., Fauch, L. & Julkunen-Tiitto, R. Screening bioactivity and bioactive constituents of Nordic unifloral honeys. Food Chem. 237, 214–224 (2017).
Sojka, M., Valachova, I., Bucekova, M. & Majtan, J. Antibiofilm efficacy of honey and bee-derived defensin-1 on multispecies wound biofilm. J. Med. Microbiol. 65, 337–344 (2016).
Majtan, J. et al. Fir honeydew honey flavonoids inhibit TNF-α-induced MMP-9 expression in human keratinocytes: a new action of honey in wound healing. Arch. Dermatol. Res. 305, 619–627 (2013).
Shantal Rodríguez Flores, M., Escuredo, O. & Carmen Seijo, M. Assessment of physicochemical and antioxidant characteristics of Quercus pyrenaica honeydew honeys. Food Chem. 166, 101–106 (2015).
Martinotti, S., Calabrese, G. & Ranzato, E. Honeydew honey: biological effects on skin cells. Mol. Cell. Biochem. 435, 185–192 (2017).
Vlcekova, P. et al. Alternative treatment of gluteofemoral fistulas using honey: a case report. Int. Wound J. 9, 100–103 (2012).
Mayer, A., Slezak, V., Takac, P., Olejnik, J. & Majtan, J. Treatment of non-healing leg ulcers with honeydew honey. J. Tissue Viability 23, 94–97 (2014).
Majtanova, N. et al. Complementary treatment of contact lens-induced corneal ulcer using honey: a case report. Contact Lens Anterior Eye 38, 61–63 (2015).
Cernak, M., Majtanova, N., Cernak, A. & Majtan, J. Honey prophylaxis reduces the risk of endophthalmitis during perioperative period of eye surgery. Phytother. Res. 26, 613–616 (2012).
Kwakman, P. H., Te Velde, A. A., De Boer, L., Vandenbroucke-Grauls, C. M. & Zaat, S. A. Two major medicinal honeys have different mechanisms of bactericidal activity. PLoS ONE 6, e17709 (2011).
Brudzynski, K., Abubaker, A., St-Martin, L. & Castle, A. Re-examining the role of hydrogen peroxide in bacteriostatic and bactericidal activities of honey. Front. Microbiol. 2, 1–9 (2011).
Bucekova, M. et al. Honeybee glucose oxidase - its expression in honeybee workers and comparative analyses of its content and H2O2-mediated antibacterial activity in natural honeys. Naturwissenschaften 101, 661–670 (2014).
Alvarez-Suarez, J. M., Giampieri, F. & Battino, M. Honey as a source of dietary antioxidants: structures, bioavailability and evidence of protective effects against human chronic diseases. Curr. Med. Chem. 20, 621–638 (2013).
Akagawa, M., Shigemitsu, T. & Suyama, K. Production of hydrogen peroxide by polyphenols and polyphenol-rich beverages under quasi-physiological conditions. Biosci. Biotechnol. Biochem. 67, 2632–2640 (2003).
Long, L. H., Hoi, A. & Halliwell, B. Instability of, and generation of hydrogen peroxide by, phenolic compounds in cell culture media. Arch. Biochem. Biophys. 501, 162–169 (2010).
Fujiwara, S. et al. A potent antibacterial protein in royal jelly. Purification and determination of the primary structure of royalisin. J. Biol. Chem. 265, 11333–11337 (1990).
Bachanova, K., Klaudiny, J., Kopernicky, J. & Simuth, J. Identification of honeybee peptide active against Paenibacillus larvae larvae through bacterial growth-inhibition assay on polyacrylamide gel. Apidologie 33, 259–269 (2002).
Shen, L. et al. Mechanism of action of recombinant acc-royalisin from royal jelly of Asian honeybee against gram-positive bacteria. PLoS ONE 2012 7(10), e47194 (2012).
Tseng, J. M. et al. Facilitative production of an antimicrobial peptide royalisin and its antibody via an artificial oil-body system. Biotechnol. Prog. 27, 153–161 (2011).
Bílikova, K., Huang, S. C., Lin, I. P., Šimuth, J. & Peng, C. C. Structure and antimicrobial activity relationship of royalisin, an antimicrobial peptide from royal jelly of Apis mellifera. Peptides 68, 90–96 (2015).
Valachova, I., Bucekova, M. & Majtan, J. Quantification of bee-derived defensin-1 in honey by competitive enzyme-linked immunosorbent assay, a new approach in honey quality control. Czech J. Food Sci. 34, 233–243 (2016).
Bucekova, M. et al. Bee-derived antibacterial peptide, defensin-1, promotes wound re-epithelialisation in vitro and in vivo. Scientific Reports 7(1), 7340 (2017).
White, J. W., Subers, M. H. & Schepartz, A. I. The identification of inhibine, the antibacterial factor in honey, as hydrogenperoxide and its origin in a honey glucose-oxidase system. Biochim. Biophys. Acta 73, 57–70 (1963).
Brudzynski, K. Effect of hydrogen peroxide on antibacterial activities of Canadian honeys. Can. J. Microbiol. 52, 1228–1237 (2006).
Chen, C., Campbell, L. T., Blair, S. E. & Carter, D. A. The effect of standard heat and filtration processing procedures on antimicrobial activity and hydrogen peroxide levels in honey. Front. Microbiol. 3, 1–8 (2012).
Sowa, P., Grabek-Lejko, D., Wesołowska, M., Swacha, S. & Dżugan, M. Hydrogen peroxide-dependent antibacterial action of Melilotus albus honey. Lett. Appl. Microbiol. 65, 82–89 (2017).
Roshan, N., Rippers, T., Locher, C. & Hammer, K. A. Antibacterial activity and chemical characteristics of several Western Australian honeys compared to manuka honey and pasture honey. Arch. Microbiol. 199, 347–355 (2017).
de Graft-Johnson, J. & Nowak, D. Effect of selected plant phenolics on Fe2+-EDTA-H2O2 system mediated deoxyribose oxidation: molecular structure-derived relationships of anti- and pro-oxidant actions. Molecules 22, 59, https://doi.org/10.3390/molecules22010059 (2017).
Liu, X. et al. Green tea polyphenols function as prooxidants to inhibit Pseudomonas aeruginosa and induce the expression of oxidative stress-related genes. Folia Microbiol. 58, 211–217 (2013).
Brudzynski, K., Abubaker, K. & Miotto, D. Unraveling a mechanism of honey antibacterial action: polyphenol/H2O2-induced oxidative effect on bacterial cell growth and on DNA degradation. Food Chem. 133, 329–336 (2012).
Lachman, J., Orsak, M., Hejtmankova, A. & Kovarova, E. Evaluation of antioxidant activity and total phenolics of selected Czech honeys. LWT-Food Sci. Technol 43, 52–58 (2010).
Maksimovic, Z. & Nedic, N. In vitro antioxidant activity of honeydew and multifloral types of honey from Serbia. Acta Period. Technol. 43, 269–277 (2013).
Oroian, M., Ropciuc, S., Buculei, A., Paduret, S. & Todosi, E. Phenolic profile of honeydew honeys from the Nort-East part ofRomania. Bulletin UASVM Food Science and Technology 73, 105–110 (2016).
Seraglio, S. K. T. et al. Development and validation of a LC-ESI-MS/MS method for the determination of phenolic compounds in honeydew honeys with the diluted-and-shoot approach. Food Res. Int. 87, 60–67 (2016).
Bag, A. & Chattopadghyay, R. R. Synergistic antibacterial and antibiofilm efficacy of nisin in combination with p-coumaric acid against food-borne bacteria Bacillus cereus and Salmonella typhimurium. Lett. Appl. Microbiol. 65, 366–372 (2017).
Xiong, L.-G. et al. Tea polyphenol epigallocatechin gallate inhibits Escherichia coli by increasing endogenous oxidative stress. Food Chem. 217, 196–204 (2017).
Blair, S. E., Cokcetin, N. N., Harry, E. J. & Carter, D. A. The unusual antibacterial activity of medical-grade Leptospermum honey: antibacterial spectrum, resistance and transcriptome analysis. Eur. J. Clin. Microbiol. Infect. Dis. 28, 1199–1208 (2009).
Bates, D., Maechler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Statist. Softw. 67, 1–48 (2015).
Acknowledgements
We would like to thank Slovak beekeepers and Slovak beekeeping association for providing honeydew honey samples. This research was supported by the Scientific Grant Agency of the Ministry of Education of the Slovak Republic and the Slovak Academy of Sciences VEGA 2/0004/18.
Author information
Authors and Affiliations
Contributions
J.M. and M. Buc. designed the research, M. Buc., M. Bur. and J.M. conducted the experiments, M. Buc., J.M. and L.P. analysed the results, M. Buc., V.M., and J.M. wrote the manuscript, all authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bucekova, M., Buriova, M., Pekarik, L. et al. Phytochemicals-mediated production of hydrogen peroxide is crucial for high antibacterial activity of honeydew honey. Sci Rep 8, 9061 (2018). https://doi.org/10.1038/s41598-018-27449-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-27449-3
This article is cited by
-
Glucose oxidase as an important yet overlooked factor determining the antibacterial activity of bee pollen and bee bread
European Food Research and Technology (2022)
-
Mutual influence of polyphenols and Lactobacillus spp. bacteria in food: a review
European Food Research and Technology (2021)
-
Use of a model to understand the synergies underlying the antibacterial mechanism of H2O2-producing honeys
Scientific Reports (2020)
-
Anti-biofilm effects and characterisation of the hydrogen peroxide activity of a range of Western Australian honeys compared to Manuka and multifloral honeys
Scientific Reports (2019)
-
The role of hydrogen peroxide in the antibacterial activity of different floral honeys
European Food Research and Technology (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
