
Abstract
Criminal investigations of suspected murder cases require estimating the post-mortem interval (PMI, or time after death) which is challenging for long PMIs. Here we present the case of human remains found in a Swiss forest. We have used a multidisciplinary approach involving the analysis of bones and soil samples collected beneath the remains of the head, upper and lower body and “control” samples taken a few meters away. We analysed soil chemical characteristics, mites and nematodes (by microscopy) and micro-eukaryotes (by Illumina high throughput sequencing). The PMI estimate on hair 14C-data via bomb peak radiocarbon dating gave a time range of 1 to 3 years before the discovery of the remains. Cluster analyses for soil chemical constituents, nematodes, mites and micro-eukaryotes revealed two clusters 1) head and upper body and 2) lower body and controls. From mite evidence, we conclude that the body was probably brought to the site after death. However, chemical analyses, nematode community analyses and the analyses of micro-eukaryotes indicate that decomposition took place at least partly on site. This study illustrates the usefulness of combining several lines of evidence for the study of homicide cases to better calibrate PMI inference tools.
Similar content being viewed by others


Introduction
The estimation of a post-mortem interval (PMI) or the time since death has been a research priority in forensic science for over a century since the pioneering work of Mégnin1 (1894), who defined the decomposition stages of corpses for the first time. Currently the minimum post-mortem interval (PMImin) is mainly estimated based on a medical assessment that relies on the physical changes of the dead body occurring in the first hours up to days and/or entomological evidence, a well-established method applied to periods of up to several weeks or months2. Although insects can be related to all decomposition stages3,4, the accuracy of PMImin estimation decreases over time5,6. Additionally, methods based on tissue chemistry such as the citrate content of bones or radiocarbon dating of different human tissue can be useful7,8. However, until now there is no accurate PMI estimation method for human remains that have already reached the dry and remains stages. But each year a number of corpses in very advanced decomposition stages are found; obtaining a reliable PMI for these can be especially crucial if other forensic evidence is scarce. For example, in the Institute of Legal Medicine in Frankfurt am Main alone, 51 corpses with a long post-mortem interval were checked using entomological evidence in the years 2014–2016 and 20% originated from outdoor environments (V. Bernhardt personal communication). Although not very frequent, the discovery of old decomposed human corpses is a reality and new methods are therefore required for long PMI estimations.
Cadaver decomposition can be seen as a continuum of several stages: fresh, bloated, active decay, advanced decay, dry and remains9. The duration of each stage depends mainly on temperature, humidity and scavenger access to the cadaver10,11,12. A still limited, but increasing number of studies have focused on the effects of cadaver decomposition on the underlying soil as new venues for gathering forensic evidence. These studies monitored the changes in soil chemistry13,14,15,16 and the community structure of soil micro-organisms17,18, bacteria19,20,21, fungi20,22,23, testate amoebae24,25, nematodes26 and micro-arthropods27,28,29,30,31,32. While the relevance of mites for criminal investigations is well-established27,30, studies of other potential forensic indicators are still rare and comparative studies are lacking. Soil arthropods such as springtails (Collembola)28,29 and ants33 were also proven useful for PMI estimation. More recently, high-throughput sequencing (HTS) of soil organisms has been used to develop new forensic tools24,34,35. These methods may each provide complementary information and PMI estimations should thus become more robust when several methods are combined. Inferring forensic evidence from a decomposing cadaver is challenging and the whole picture may only appear clear when several independent lines of evidence are combined.
In the present work, five different approaches were applied to a case study, aiming to reconstruct the crime scene and the PMI. These different approaches were also combined and analysed for correlation via Multi-Factor Analysis (MFA). Human bones were found in a forest area in the Swiss midlands. Bones were examined morphologically and biochemically to determine sex and biological age of the individual and to estimate PMI. Soil samples were collected and analysed for selected chemical markers, for nematode and mite diversity (based on morphology), and micro-eukaryotes (using high throughput DNA amplicon sequencing, i.e. metabarcoding) to assess possible differences in community structure. Due to legal reasons, some details about the finding must be kept under nondisclosure.
Results
Bone and hair analyses
The preservation and the representation of the identifiable skeletal elements were low due to signs of thermal destruction caused by burning (Fig. 1). Particularly affected by the fire were the limbs including hands and feet. Parts of the pelvis and skull were present allowing a sex determination as male. An age at death was estimated to be between 18 to 25 years. The 14C in the collagen of the femur was integrated approximately between 10–12 years before (1 sigma) the remains were found. The 14C in the hair was integrated approximately in the last 3 years before (1 sigma) the investigation.

Identified bones from human remains (burnt fragments are not shown) found in a forest area in Switzerland.
Chemical analyses
Soil bioavailable P content was higher beneath the head (192.54 µg/g) and upper body (147.99 µg/g) than under the lower body (LB) and control samples (ranging from 17.52 µg/g in control West to 65.87 µg/g in control South, Table 1). Total N was at least twice as high under the head compared to other samples (Table 1). Other soil chemical markers (pH, NH4 +, NO3 −, Mg2+, Ca2+ and C content) did not differ between the control and cadaver samples (Table 1). The cluster analysis separated the samples into two clusters: 1) head and upper body and 2) controls and lower body (Fig. 2a). In the PCA the sample from beneath the head sample was separated from the other samples (Fig. 2b).

Multiple factor analysis (MFA) on chemical variables and community matrices of nematodes, mites and of micro-eukaryotes from soil samples taken beneath human remains and controls in a forest area in Switzerland. On the left part the projections of the samples according to the clustering (a,c,e,g) and the PCA (b,d,f,h) calculated on chemical variables (a,b), nematode families (c,d), mites taxa (e,f) and micro-eukaryotic OTUs (g,h) are represented. On the right part of the figure the projection of the samples according to the global analysis (i) and the correlation circle (j) of the most characteristic variable (P < 0.05) of the first and second dimensions are shown. The four variable types are represented with bold italic font and dashed arrows for chemical variables, bold font and dotted arrows for nematode families, italic font and dot dash arrows for mite taxa and normal font and plain arrows for micro-eukaryotic OTUs. In order to improve readability, only names of variables with a distance between the origin and their coordinate higher than 0.05 were shown. The correlation circle is shown on a log scale because of the difference between coordinates from eukaryotic and chemical data.
Nematodes
Nematode density under the cadaver was highest beneath the head (2488 ind./100 g dry soil), lower under the upper body (1653 ind./100 g) and lowest under the lower body (1192 ind./100 g), the latter values were within the range of the control samples (982–1402 ind./100 g). In total 19 nematode families were identified (Table 2). Diversity was highest under the lower body and in the control samples (12–14 families), nine of which being present in all of these samples (Table 2). By contrast, only five and six families were present beneath the head and the upper body respectively (Table 2). Bacterial feeding Cephalobidae, Rhabditidae and Plectidae were the three most abundant families and occurred in all samples (Table 2).
The cluster analysis of the community data separated the samples into two groups: 1) head and upper body and 2) controls and lower body (Fig. 2c). In the PCA the sample from beneath the head was separated from all other samples (Fig. 2d). Bacterial feeding Cephalobidae and Myolaimidae were most abundant in the sample beneath the head. The Myolaimidae were only found beneath the head and were further identified as Myolaimus sp. (Fig. 2i,j). Tylenchidae (herbivorous), Qudsianematidae (predators, omnivorous) and Prismatolaimidae (bacterivorous) were associated with the controls North and South and the lower body samples (Fig. 2i,j). Control West and East were associated with Teratocephalidae (bacterivorous) and Leptonchidae (fungivorous) (Fig. 2i,j). Control West was associated with the bacterivorous Alaimidae. Four nematode families (Alaimidae, Prismatolaimidae, Qudsianematidae, Tylenchidae) were considered as indicators for controls and lower body (Fig. 3).

Abundance of the bioindicators of head and upper body (H/UB), and controls and lower body (C/LB) from a criminal case investigation in Switzerland. For each group the eukaryotic bioindicators are sorted according to their taxonomic assignation followed by nematodes families and mite taxa.
Mites
A total of 391 mites belonging to four Acari orders (Astigmata, Mesostigmata, Prostigmata and Oribatida) were identified from the seven sampling sites. The most diverse and numerous were the Mesostigmata (N = 130; 13 species) followed by the Astigmata (N = 126; five species). A number of soil mites found in the control samples, such as Oribatida and Prostigmata were only discerned as morphotypes (Table 3).
Most mites were found in the adult stage, with the exception of members of the Astigmata. The Acaridae Sancassania berlesei (=Caloglyphus berlesei), S. oudemansi (=Caloglyphus oudemansi), and the Histiostomatidae unidentified sp. 1, 123 specimens in total, were sampled in their immature, phoretic hypopial form. Interestingly, Astigmata were extracted only from the sample sites associated with bones i.e. from the soil beneath the head, upper body and lower body (Table 3). These samples also contained the greatest richness of Mesostigmata.
Considering all seven samples, three Mesostigmata species frequently found in forest soil were the most numerous, the detritivorous Uropodidae unidentified sp. 1 (N = 55), Prozercon traegardhi (N = 13) and the predaceous Vulgarogamasus sp. (N = 23). A particular soil mite morphospecies of the Oribatida, Oppiidae, Oppiella was highly abundant and 100% prevalent.
In terms of the singularity of sample diversity, sample sites and mite species, mites aggregated into three defined clusters. Upper body and head were different from each other and from the remaining five samples, which group together (Fig. 2f). The clustering further indicated that the upper body and head were the two most distinctive samples (Fig. 2e). These two samples contained rare species, which are not typical inhabitants of forest soil. The taxa found in these samples rather correspond to poultry farm or granary species such as S. berlesei (hypopi), which is also a predominant species in corpses (especially in advanced decay or mummification stages), accompanied by a specialised predator of these unique farming environments, Macrocheles matrius (Fig. 2i,j).
Micro-eukaryotes
The metabarcoding analysis revealed 386 OTUs from a total of 648′344 sequences, that excluding Metazoa and Embryophyceae (animals and land plants), and rare OTUs (i.e. <0.0005% of the Hellinger transformed community matrix). Globally, the soil micro-eukaryotic community was dominated by fungi, with a majority of Basidiomycota in the control/lower body samples versus a large dominance of Mucoromycotina in the head/upper body samples (Supplementary Figure 1). Likewise, taxa containing many small bacterivores (Heterolobosea, Chrysophyceae, Tubulinea, Cercozoa) were relatively more abundant in head and upper body samples. Variosea (Amoebozoa) as well as the parasitic Apicomplexa were in turn more abundant under the control/lower body samples.
Multiple Factor Analysis (MFA), hierarchical clustering and Indicator Value
The first dimension showed a clear dichotomy between the head/upper body and control/lower body sample groups (Fig. 2i). This pattern is in line with the separate analyses of the three community matrices (Fig. 2d,f,h) and by the cluster analyses (Fig. 2a,c,e,g). This similarity among results is also shown by the RV coefficients which reveal a significant correlation among the three matrices (Table 4). By contrast, the chemical variables were only correlated with the nematodes data.
A total of 97 variables (three chemical variables, 79 eukaryotic OTUs, seven mite taxa and eight nematode families) were characteristic of the first dimension of the MFA ordination (Fig. 2j). From the 97 variables characteristic of the first dimension, the majority were well associated with the head/upper body samples (3/3 chemical variables, 61% of micro-eukaryotic OTUs, 2/8 nematodes families and 86% of mite taxa).
The IndVal analyses revealed 18 eukaryotic OTUs and the mite species Sancassania berlesei as head/upper body indicators, while indicators for controls/lower body indicators, included 53 eukaryotic OTUs, four nematode families (Alaimidae, Prismatolaimidae, Qudsianematidae, Tylenchidae) and two mite families (Uropodidae, Oppiidae) (Fig. 3).
Discussion
The analyses of this case were divided into two parts: the examination of the human remains alone and the analyses of the soil samples beneath the remains.
The PMI estimation via bomb peak radiocarbon dating resulted in a time range of up to 3 years (hair F14C-data). From the anthropological investigation, it was also assumed that the bones could not have been lying outside for less than one year due to the lack of fatty appearance, no filling of the medullary cavity of the femur, no smell and no soft tissue36 except of hair, which was found right beneath the occipital bone (Os occipetale). Although changes in taphonomy, in particular a faster decomposition due to the thermal destruction (fire) cannot be excluded. In conclusion, radiocarbon dating and anthropological analysis suggest that the individual must have been died between one to three years before the remains were found.
When interpreting the F14C-data of the collagen, the turnover rates of human bones were based on previous studies8,37,38,39. Since this individual was a late adolescence or young adult male at the time of death (based on anthropological bone features), a turnover rate of 10–30% per year (between the age of 10 and 15 years) and 3–1.5% per year (after the age of 20 years) has to be considered38. Therefore, collagen data are always mixed values containing the carbon signal of the last years of life. Nevertheless, the collagen of the femur has its core area between 9–12 years before the remains were found. Assuming that most of the incorporated carbon derives from the two first life decades the data are compatible with a young adult.
The individual was subsequently identified via a DNA match in the EDNAIS database, Switzerland. A witness statement showed that the individual was last seen 22 months before the remains were found (summer/date under nondisclosure). All data are thus concordant with this information.
The analyses of soil samples beneath decomposing cadavers has generally observed a sharp increase of soil nutrient content and pH during the active and advanced decay phases of cadaver decomposition40. This is when the most rapid breakdown of a cadaver takes place14. In this study, bones were found at the stage of dry remains, which does not mean that the concentration of nutrients have returned to basal levels14. According to the marker categories we have developed41, pH and NH4 + are described as early peak markers (EPMs), showing significant increases at the beginning of the decomposition process (Szelecz et al. submitted). The absence of these early peak markers in the samples beneath the remains suggests two interpretations, either the timespan since death was sufficient for EPMs to return to basal levels or the body had started decomposing elsewhere and was transported to the find site when it had already reached a later decomposition stage i.e. advanced decomposition.
At later decomposition stages, Ca2+ and other additional late peak markers (LPMs) such as NO3 − increase15,42. NO3 − levels were significantly elevated beneath carcasses one year post-mortem42 which was confirmed by our studies41. The lack of higher NO3 − concentration under the cadaver in this study suggests that the PMI was >1 year. The absence of LPMs such as Ca2+ might indicate an upper limit for the PMI as shown by Melis et al.15 who observed elevated calcium levels at carcass sites from three to six years post-mortem probably released from the bones. However, patterns of soil nutrient response to decomposing cadavers vary among studies15 and thus it is currently not possible to develop a precise PMI estimation method from soil chemical characteristics alone.
The presence of elevated levels for P (head and upper body) and N (head) indicate that the time elapsed since peak decomposition was not long enough for these markers to return to basal levels. Significantly elevated P levels were described one and three years post-mortem16. N levels beneath large ungulate carcasses were significantly higher two years post-mortem and 10 times higher than in the surrounding soil even three years post-mortem (not significant)16. Nevertheless, elevated levels of P beneath the head and upper body, plus a higher N concentration beneath the head and the clustering of head and upper body samples suggests that the decomposition process had at least partly taken place on site. In contrast, the lower body part groups with the controls indicate that decomposition may have been hindered or not taken place in this area. This is in accordance with the findings that the lower body parts were burnt much more than the upper body parts (excluding arms). However, this interpretation should be taken cautiously given the sample size of our study.
Nematode density in all samples was within the range reported elsewhere for terrestrial nematodes26,43 and the three most abundant families that occurred in all samples i.e. Cephalobidae, Rhabditidae and Plectidae are common in soil44. The clustering grouped nematode communities from controls and lower body samples together and revealed four indicators: Alaimidae, Prismatolaimidae, Qudsianematidae and Tylenchidae. Nematodes can be classified according to their feeding habits and life-history characteristics43,45. Among the indicators Alaimidae and Qudsianematidae are typical K-strategists (persisters) with a high sensitivity to disturbances46. Tylenchidae are very tolerant to disturbances, they are r-strategists (colonizers) and frequently found in soils44. Prismatolaimidae can be considered as intermediate between these two groups being more sensitive than Tylenchidae46. The presence of nematodes that are sensitive to disturbances in line with a higher family richness indicates that the control and lower body samples might not have been exposed to stress26.
By contrast, head and upper body samples were characterized by a low family richness and were dominated by r-strategists i.e. bacterial feeding nematodes that are tolerant to pollutants and organic matter decomposition46. Especially Myolaimidae which only occur in the head samples were shown to be enrichment opportunists47. Myolaimidae were further determined as Myolaimus sp., which is rarely recorded in samples from terrestrial habitats48. During the composting process Myolaimus sp. was restricted to the last phase of the composting process (maturation) indicating that it might have special requirements49. Therefore, Myolaimus sp. might be an indicator for a late decomposition stage suggesting that only part of the decomposition process took place in-situ and bones and some flesh remained after the fire to decompose. Nevertheless, in a study on the effect of decomposing pig cadavers on soil nematode communities in the same general region (albeit in a different forest and soil type) Myolaimidae did not occur at all during the whole decomposition process within a one-year post-mortem period26. In that study, it was also shown that family richness was still significantly lower 263 days post-mortem26. Here further studies are necessary. Bearing in mind that comparable data are sparse, a PMI of at least 8–9 months but more likely >1 year is suggested based on the nematode data.
Of the seven samples analysed in this study two stand out due to their unique mite fauna composition, which is exogenous to the forest soil and comprised species of forensic importance (previously found on corpses) and used as trace evidence. Communities from the upper body, followed by head are unique in that they contain the only and highest number of Sancassania berlesei, and the only specimens of Macrocheles matrius. Both, S. berlesei and its main predator, M. matrius are inhabitants of more synanthropic habitats, particularly those related to granaries, farms (poultry and pig hay-beddings) and less often agricultural lands (cereal fields). In Europe, these species are found in compost, poultry litter, decayed bulbs and tubers, especially deep litter broiler houses, and have been originally found in stored grains or food products50,51,52. Interestingly, S. oudemansi (found in the head sample) is even more restricted to synanthropic habitats, poultry settlings or granaries, wherever wheat or other grains abound51,52,53,54.
The large numbers of Sancassania all found in hypopial stages, in upper (the body and head) samples indicate: 1) that the human remains and their associated Sancassania (and Macrocheles) mites have originated in one of the aforementioned habitats (e.g. a farm); 2) that a massive Sancassania population was living from the remains; and 3) that this well-developed population was suddenly exposed to very unfavourable environmental conditions (e.g. drought, fire). These facts are strong indicators of movement or relocation of the remains. From the data of the two key sites (upper body and head) it is possible to infer that the original massive mite population very likely built up from (early) active to advance decay, and that at this stage the remains were transported and deposited in their final resting place, the forest soil, more precisely on a limited space or position where the upper body and head bones were later found. This is also explained by the lack of any other forensic markers, characteristic of decay on forest soil. Indeed, there is no acarological evidence indicative of decomposition having taken place on the forest soil.
Sancassania berlesei, like the majority of stored-product mites (Astigmata) are sensitive to humidity; exposure to low humidity levels leads either to death or to moulting into more resistant immature stages commonly known as hypopi51,54,55. Optimal conditions imply very high humidity levels; water saturated habitats or exceptionally damp conditions, such as enclosed or sealed habitats. A good example would be a human corpse wrapped, concealed in clothes, inside bags or inside any sort of sealed container able to maintain the damp conditions needed for the mites to thrive1,27,30,56,57,58,59,60. This might explain the well-developed colony of S. berlesei found only in a reduced area or particular patch of soil (upper body and head sites). Soil under the lower body bones did not contain any forensically relevant mite species. Mites in this area are the same as those found in controls, lower body and clustered together with most controls. This suggests that the lower body site was not the place or location of the ‘container’ or ‘bag’ with human remains, and perhaps the bones observed on this area were the product of the movement of the bones by forest scavengers (foxes, dogs, birds, etc.). Under unfavourable conditions such as drought and lack of food, or food lacking nutritional content, S. berlesei will always produce hypopi and most of these hypopi are active forms. Hypopi are second instar nymphs able to disperse mainly (not exclusive) as phoretic on other animals to new, optimal environmental conditions30,51,61,62.
Only a few insects have been described as hosts of S. berlesei hypopi, particularly Scarabaeidae (chaffers) and Tenebrioniidae (stored product beetles)62,63,64. The unique association with the flour beetle, Tenebrio molitor, confirms its habitat specificity to granaries or stored grain facilities. Termination of hypopial forms will occur when both high humidity and food are restored, moulting to the next stage or trinonymph; otherwise they will die as hypopi55. Experiments designed to produce hypopi considered exposure to dry cultures and lack of food55. All 121 S. berlesei collected in this case were hypopi (Fig. 4), no adult was found. It means that the massive population went through a bottleneck of unfavourable conditions, like exposure to extreme drought or lack of food. In addition to mites, the two sample sites upper body and head contained the majority of charcoal particles, indicative of fire and the site where the remains might have been burnt. The remains must have been burned exactly in that patch of forest soil. The fire most likely consumed the external layers of the wrapped remains killing most mites of the outer parts. The thickness of the massive population of perhaps thousands to millions of mites, which probably reached several cm, protected the most internal part of the colony from fire. Those hundreds of S. berlesei that survived the fire were then exposed to a foreign environment with no food, not enough humidity, produced resistant offspring or hypopi while surrounded by soil predators.

A hypopus of Sancassania berlesei from the head sample site (H) from a criminal case investigation in Switzerland.
Remarkably, the presence of a second mite species, a strict inhabitant of farm/granary habitats affirms the origin of the fauna associated with upper body and head. Macrocheles matrius is a predator, a foreign species to the forest soil and to carcasses/corpses, but specialized in hunting and consuming the acarids of granaries or poultry litter50,54,65. In terms of PMI estimation, the massive colony of mites inhabiting the original human remains that reached the forest soil are similar to that of a concealed corpse in its later stage of decomposition, very likely a mummified corpse. Judging by the size of the surviving population of Sancassania hypopi, and the time it takes to mummification of a human adult inside a sealed enclosure, it is possible that death and concealment of the body happened at least 8–12 months before the remains were disposed on the forest soil. This is due to the life-span of these hypopi which might have struggled to survive in the new habitat and conditions. Mite traces are good indicators of what might have happened from the moment of death until decomposition reached advanced stages, in this case indicating the corpse location on the forest soil, perhaps its origin (location of death) and confirming the burning process.
The presence of a decomposing corpse has clearly modified the micro-eukaryote communities. The combination of our two statistical analyses (IndVal and MFA) allowed us to determine a limited number of OTUs that were either typical for (1) head and upper body (H/UB) samples or (2) controls and lower body (C/LB).
General communities differ considerably between head/upper body samples and control/lower body. Basidiomycetes, which dominated the control and lower body samples, are virtually replaced by Mucoromycota in the head and upper body samples. While the first group includes many saprotrophs and mycorrhizae, the second includes typically r-strategists such as Mortierella spp. This shift has been observed also in the case of perturbations such as disruption of connections between the root and the fungus66. The higher incidence of mainly bacterivorous taxa is associated with higher values of P and N, which are expected to increase bacterial densities.
Soil fungi are the best-studied group of soil micro-eukaryotes and thus their community patterns are easy to interpret. For instance, OUT X242 is affiliated with genus Trichosporon, a fungus which belongs to the normal skin microbiota67. Members of this genus have been found associated with late stages of decomposition of human bodies68. OUT X246 (Rhopalomyces) is an exclusive parasite of nematode eggs69; its abundance in the head and upper body samples is thus consistent with the high number of nematodes found in these samples. Likewise, the prevalent presence of ectomycorrhizal fungi among the controls and lower body (C/LB) indicators (i.e. OTUs X3, representing several tree mycorrhizae in forests and X362, a Pezizomycotina affiliated to genus Stomiopeltis) was to be expected as these organisms are ubiquitous in forest soils.
Several species of amoeboid protists, known to occur only in stable systems, were found amongst the indicator OTUs of the controls and lower body samples. Amongst them, Variosea (Amoebozoa) from the Ischnamoeba/Darbyshirella clade (X109, X503) are reticulate organisms with very thin and delicate pseudopodia which are typical slow-growing organisms70. Rhogostoma (X273), like many other testate amoebae are also typically K-selected organisms that are supposed to perish underneath cadavers24,25. On the other hand, the presence of OUT X152, affiliated to the amoebozoan genus Mycamoeba suggests a beginning of recovery, as these organisms were shown to be negatively influenced by the presence of a cadaver in an experimental setup containing three pig corpses laid on forest soils71. The presence of this particular OTU therefore suggests a PMI of over one year71.
Conclusions
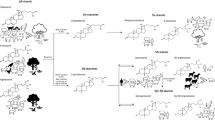
Following the analysis of five evidential components the following course of events can be proposed for this case: Human remains were found in a Swiss Plateau forest. The examination of the bones and the hair revealed that the remains were from a young-adult human male who must have died 1–2 years before the discovery. This person was subsequently identified and was last seen 22 months before his remains were found. The bones showed signs of scavenging and exposure to fire, likely the remains were burnt in-situ. The analyses of chemical constituents, nematodes and micro-eukaryotes revealed that the decomposing remains had probably been on the site for at least 8–9 months and more likely more than one year. This means at least part of the decomposition process had taken place on site.
Mite evidence, however, suggests that the corpse was translocated from its original crime scene to the forest soil. It first decomposed in-situ, reaching late stages of decomposition before being moved to the forest. According to survival rates of Sancassania hypopi, the remains were likely burnt in a small patch on the forest soil, just a few months before discovery.
If the decomposing remains were transported to the site at least 12 months before the finding, the person must have died between summer and spring before that. Bearing in mind that decomposition is accelerated by temperature and insect access to the body10,72 no tissue would have been left if the person had started to decompose at a different location in summer. Instead, our findings suggest that the person was killed probably in autumn or winter, the corpse started to decompose in a confined environment, possibly a farm (or similar), and was perhaps brought to the final or finding site in the early spring of the following year where it was partially burned (Fig. 5).

Reconstruction of the body posture and soil sampling area.
Although some of the methods shown are still in the process of being developed (e.g. nematodes as forensic indicators for PMI estimation), others are already well-established (e.g. forensic anthropology). Nevertheless, the interpretation of a crime scene gathered from the simultaneous study and combined analysis of independent lines of evidence illustrates the added value of this multiproxy approach (Fig. 5). The main aim of this work was to provide a strong incentive for case work as well as experimental studies to further develop a comprehensive toolbox for forensic or crime scene investigations.
Material and Methods
Case history and sampling
Human bones were found in a forest in the Swiss Plateau at a height of approx. 400–450 m above sea level. The ground was covered with dry leaves and the bones were partly covered with leaves and small branches. After their removal, the bones showed signs of thermal destruction. Some burned tree trunks with a diameter of approx. 20–30 cm and bigger branches were arranged in a square approximately 2 × 2.5 m around the bones. The branches originally covering the bones showed signs of charring and had been removed by the finder. These burnt branches were partially overgrown by moss. Due to the position of the skeletal elements in comparison with their biological anatomy, the location of the body before decomposition and burning, could be reconstructed (Fig. 6). Beside the bones, some scalp hair appeared below the Os occipetale during the recovery and residues of different fabrics, a key, some coins and jewellery were found.

Summary timeline of all suggested PMIs from a criminal case investigation in Switzerland.
Soil samples (soil cores of 10 cm in diameter by 10 cm depth) were taken from the area where the bones were found i.e. beneath the head (H), upper body (UB) and lower body (LB) and from control areas not impacted by the cadaver or the fire in the four cardinal directions, 4–6.5 m from the body: North (CN), South (CS), East (CE) and West (CW) (Fig. 6). The surrounding area was searched for further bones and everything was transferred to the Department of Physical Anthropology, Institute of Forensic Medicine, University of Bern, for examination.
Bone and hair analyses
The human bones which did not show severe thermal destruction were investigated anthropologically according to Acsádi and Nemeskéri73 (1970), Buikstra and Ubelaker74 (1994) and Rösing et al.75.
A 5.1 gram sample of the femoral diaphysis without any visible thermal destruction was sent for radiocarbon dating to CEZ Archäometie gGmbH, Mannheim, Germany. Additionally, 2.0 gram of hair was also sent for radiocarbon dating in order to perform a bomb peak application8,39. Bone collagen was extracted76,77 and the hair was carefully cleaned, both were measured via MICADAS-accelerator. The calibration was performed with CALIBomb78.
Soil chemical analyses
Soil pH was measured in the laboratory with a pH metre after diluting the sample in water in a 1:2.5 proportion. Ammonium and nitrate analyses were performed directly after sampling using colorimetric determination79. Total carbon and nitrogen was determined using a CHN analyser (Thermo Finnigan Flash EA 1112) on dry, ground soil. Bioavailable phosphorus content was determined by colorimetric analysis according to the Olsen method80. The concentrations of Mg2+ and Ca2+ were determined using inductively coupled plasma optical emission spectrometry (Perkin-Elmer Optima 3300 DV ICP-OES) preceded by a cation exchange capacity extraction (CEC, Cobaltihexamine method). Potential chemical soil markers were classified into (1) early peak markers (EPM) that show the highest concentrations early in decomposition until advanced decay following degradation to (2) late peak markers (LPM), during the dry and remains stage. Some of them might show (3) late elevated levels (LEL)41.
Nematodes
Nematodes were extracted from 100 g of soil using a modified Baermann funnel technique according to the protocol of Brown and Boag81. Nematodes were counted alive using a dissecting microscope (Olympus SZ51) and then fixed with heated formaldehyde (4%) and heat-killed at 65 °C for 3 minutes. One hundred randomly chosen nematodes per sample were identified to family level using an upright light microscope at 400x magnification (Axio Lab.A1, Zeiss). Identification was based on the identification guides of Bongers82 and Scholze and Sudhaus83. All nematode densities are given in individuals per 100 gdw.
Mites
A 500 mL soil sample was collected from each sampling site and placed on Tullgreen-Berlese funnels to extract moving mites into a collector jar with 70% ethanol (following the method used by Saloña et al.60). Mites were cleaned from the collector jars, cleared for permanent mounting in Hoyer84 and identified using a variety of keys and descriptions51,65,85,86,87,88,89. A voucher number was assigned to all microscope slides and they were deposited in the Forensic Acarology Reference Collection, Acarology Lab, University of Reading.
Soil micro-eukaryotes
Total soil DNA was extracted using the MoBio PowerSoil isolation kit following the manufacturer’s instruction. To screen the eukaryotic diversity of the seven soil samples, the V9 fragments of the small sub-unit ribosomal RNA (SSU rRNA) was amplified following the protocol of Amaral-Zettler et al.90 and the amplicons sequenced with an Illumina HiSeq. 2000® sequencer (Fasteris, Geneva, Switzerland). The resulting V9 sequences were then processed as follows: Sequences were removed when the average phred score of a 50 nucleotides window was below 20. Chimera were identified and discarded using the program Uchime (v. 7.0.1090)91 by comparing sequences among them and to the PR2 database92. The sequences were then clustered into Operational Taxonomic Units (OTUs) using the software Swarm (v. 1.2.12)93 with the default parameters. The OTUs were finally taxonomically assigned by aligning the dominant sequence of each OTU to the PR2 database using the program Ggsearch36 (Fasta package v. 36.3.6 http://faculty.virginia.edu/wrpearson/fasta/CURRENT/) with the default parameters. OTUs assigned to prokaryotes were removed from the analysis as well as those assigned to Metazoa or Embryophyceae in order to avoid bias which would be caused by a piece of those macro-organisms in the soil sample.
Numerical analysis
Chemical variables were scaled and community matrices (Eukaryotes, nematodes and mites) were Hellinger-transformed94. As metabarcoding data are prone to yield a huge proportion of rare OTUs, we discarded OTUs representing less than 0.5‰ of the Hellinger-transformed matrix. A multiple factor analysis (MFA) (package FactoMineR v. 1.31.495), was then computed with the four data matrices to assess the correlative structure of the data. A hierarchical clustering was also performed with Euclidean distances on the chemical variables and with Bray-Curtis distances on each of the three community matrices.
The bioindicator value of each taxon or OTU in each of the three community data sets was assessed using an indicator species analysis (function indval; package labdsv 1.6–1)96. For this analysis, we used two groups based on the results of the four clusterings and the first dimension of the MFA: (1) head and upper body, and (2) lower body and controls. OTUs were selected as bioindicators if the p-value of their indicator value was under 0.05 after 10000 iterations. As the number of OTUs was high, many were significant in the IndVal analysis. To keep the number manageable, we only retained those that had both a significant Indval and showed a clear pattern in the first dimension of the MFA.
Data availability statement
Due to legal reasons (criminal case investigation), the availability of the data is restricted and data must be kept under nondisclosure.
References
Mégnin, J. P. La faune des cadavres-application de l’entomologie a la médecine légale. (Gauthier-Villars et fils, 1894).
Amendt, J. et al. Best practice in forensic entomology—standards and guidelines. Int. J. Legal Med. 121, 90–104 (2007).
Anderson, G. S. & VanLaerhoven, S. L. Initial studies on insect succession on carrion in southwestern British Columbia. J. Forensic Sci. 41, 617–625 (1996).
Sharanowski, B. J., Walker, E. G. & Anderson, G. S. Insect succession and decomposition patterns on shaded and sunlit carrion in Saskatchewan in three different seasons. Forensic Sci. Int. 179, 219–240 (2008).
Amendt, J., Richards, C. S., Campobasso, C. P., Zehner, R. & Hall, M. J. R. Forensic entomology: applications and limitations. Forensic Sci. Med. Pathol. 7, 379–392 (2011).
Greenberg, B. & Kunich, J. C. Entomology and the law: flies as forensic indicators. (Cambridge University Press, 2002).
Schwarcz, H. P., Agur, K. & Jantz, L. M. A new method for determination of postmortem interval: citrate content of bone. J. Forensic Sci. 55, 1516–1522 (2010).
Ubelaker, D. H., Buchholz, B. A. & Stewart, J. E. B. Analysis of artificial radiocarbon in different skeletal and dental tissue types to evaluate date of death. J. Forensic Sci. 51, 484–488 (2006).
Payne, J. A. A summer carrion study of the baby pig Sus Scrofa Linnaeus. Ecology 46, 592–602 (1965).
Campobasso, C. P., Di Vella, G. & Introna, F. Factors affecting decomposition and Diptera colonization. Forensic Sci. Int. 120, 18–27 (2001).
Mann, R. W., Bass, W. M. & Meadows, L. Time since death and decomposition of the human body: variables and observations in case and field studies. J. Forensic Sci. 35, 103–111 (1990).
Megyesi, M. S., Nawrocki, S. P. & Haskell, N. H. Using accumulated degree-days to estimate the postmortem interval from decomposed human remains. J. Forensic Sci. 50, 1–9 (2005).
Benninger, L. A., Carter, D. O. & Forbes, S. L. The biochemical alteration of soil beneath a decomposing carcass. Forensic Sci. Int. 180, 70–75 (2008).
Carter, D. O., Yellowlees, D. & Tibbett, M. Cadaver decomposition in terrestrial ecosystems. Naturwissenschaften 94, 12–24 (2007).
Melis, C. et al. Soil and vegetation nutrient response to bison carcasses in Białowieża Primeval Forest, Poland. Ecol. Res. 22, 807–813 (2007).
Towne, E. G. Prairie vegetation and soil nutrient responses to ungulate carcasses. Oecologia 122, 232–239 (2000).
Metcalf, J. L. et al. Microbial community assembly and metabolic function during mammalian corpse decomposition. Science 351, 158–162 (2016).
Pechal, J. L. et al. Microbial community functional change during vertebrate carrion decomposition. PLoS ONE 8, e79035 (2013).
Cobaugh, K. L., Schaeffer, S. M. & DeBruyn, J. M. Functional and structural succession of soil microbial communities below decomposing human cadavers. PLOS ONE 10, e0130201 (2015).
Parkinson, R. A. et al. Microbial community analysis of human decomposition on soil in Criminal and environmental soil forensics (eds Ritz, K., Dawson, L., Miller, D.) 379–394 (Springer, 2009).
Pechal, J. L. et al. The potential use of bacterial community succession in forensics as described by high throughput metagenomic sequencing. Int. J. Legal Med. 128, 193–205 (2014).
Carter, D. O. & Tibbett, M. Taphonomic Mycota: fungi with forensic potential. J. Forensic Sci. 48, 2002169 (2003).
Hawksworth, D. L. & Wiltshire, P. E. J. Forensic mycology: the use of fungi in criminal investigations. Forensic Sci. Int. 206, 1–11 (2011).
Seppey, C. V. W. et al. Response of forest soil euglyphid testate amoebae (Rhizaria: Cercozoa) to pig cadavers assessed by high-throughput sequencing. Int. J. Legal Med. 130, 551–562 (2016).
Szelecz, I., Fournier, B., Seppey, C., Amendt, J. & Mitchell, E. Can soil testate amoebae be used for estimating the time since death? A field experiment in a deciduous forest. Forensic Sci. Int. 236, 90–98 (2014).
Szelecz, I. et al. Effects of decomposing cadavers on soil nematode communities over a one-year period. Soil Biol. Biochem. 103, 405–416 (2016).
Braig, H. R. & Perotti, M. A. Carcases and mites. Exp. Appl. Acarol. 49, 45–84 (2009).
Merritt, R. W. et al. Collembola of the grave: A cold case history involving arthropods 28 years after death. J. Forensic Sci. 52, 1359–1361 (2007).
Perez, A. E., Haskell, N. H. & Wells, J. D. Evaluating the utility of hexapod species for calculating a confidence interval about a succession based postmortem interval estimate. Forensic Sci. Int. 241, 91–95 (2014).
Perotti, M. A. & Braig, H. R. Phoretic mites associated with animal and human decomposition. Exp. Appl. Acarol. 49, 85–124 (2009).
Perotti, M. A., Braig, H. R. & Goff, M. L. Phoretic mites and carcasses.in Current Concepts in Forensic Entomology: Novel Arthropods, Environments and Geographical Regions (eds Amendt, J., Campobasso, C. P., Grassberger, M., Goff, M. L.) 69–92 (Springer, Amsterdam, 2010).
Alejandra Perotti, M., Lee Goff, M., Baker, A. S., Turner, B. D. & Braig, H. R. Forensic acarology: an introduction. Exp. Appl. Acarol. 49, 3–13 (2009).
Moretti, T., de, C. & Ribeiro, O. B. Cephalotes clypeatus Fabricius (Hymenoptera: Formicidae): nesting habits and occurrence in animal carcass. Neotrop. Entomol. 35, 412–415 (2006).
Finley, S. J., Benbow, M. E. & Javan, G. T. Potential applications of soil microbial ecology and next-generation sequencing in criminal investigations. Appl. Soil Ecol. 88, 69–78 (2015).
Metcalf, J. L. et al. A microbial clock provides an accurate estimate of the postmortem interval in a mouse model system. eLife 2, e01104 (2013).
Berg, S. & Protsch von Zieten, R. Datierung von Skelettfunden in Identifikation unbekannter Toter: Interdisziplinäre Methodik, Forensische Osteologie Arbeitsmethoden der Medizinischen und Naturwissenschaftlichen Kriminalistik(ed. Leopold, D.)107–162 (1998).
Geyh, M. A. Bomb radiocarbon dating of animal tissues and hair. Radiocarbon 43, 723–730 (2001).
Hedges, R. E. M., Clement, J. G., Thomas, C. D. L. & O’Connell, T. C. Collagen turnover in the adult femoral mid-shaft: modeled from anthropogenic radiocarbon tracer measurements. Am. J. Phys. Anthropol. 133, 808–816 (2007).
Wild, E. et al. 14C dating with the bomb peak: An application to forensic medicine. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 172, 944–950 (2000).
Tibbett, M. & Carter, D. O. Research in forensic taphonomy: a soil-based perspective.in Criminal and environmental soil forensics (eds Ritz, K., Dawson, L., Miller, D.) 317–331 (Springer, 2009).
Szelecz, I. Impact of vertebrate cadavers on soil communities and soil chemistry to develop new forensic indicators for estimating the time after death (unpublished doctoral thesis). (University of Neuchâtel, 2016).
Anderson, B., Meyer, J. & Carter, D. O. Dynamics of ninhydrin-reactive nitrogen and pH in gravesoil during the extended postmortem interval. J. Forensic Sci. 58, 1348–1352 (2013).
Bongers, T. & Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 10, 239–251 (1998).
Yeates, G. W. Nematodes as soil indicators: functional and biodiversity aspects. Biol. Fertil. Soils 37, 199–210 (2003).
Yeates, G. W., Bongers, T., De Goede, R. G. M., Freckman, D. W. & Georgieva, S. S. Feeding habits in soil nematode families and genera—an outline for soil ecologists. J. Nematol. 25(3), 315–331 (1993).
Ferris, H. & Bongers, T. Indices developed specifically for analysis of nematode assemblages in Nematodes as Environmental Indicators (eds Wilson, M. J. & Khakouli-Duarte, T.) 124–145 (CABI, 2009).
Bongers, T., De Goede, R. G. N., Korthals, G. W. & Yeates, G. W. Proposed changes of c-p classification for nematodes. Russ. J. Nematol. 3, 61–62 (1995).
Bärmann, E. V., Fürst van Lieven, A. & Sudhaus, W. Revision and phylogeny of Myolaimus Cobb, 1920 (Secernentea, Nematoda) with the description of four new species. J. Nematode Morphol. Syst. 12, 145–168 (2009).
Steel, H. et al. Survival and colonization of nematodes in a composting process. Invertebr. Biol. 132, 108–119 (2013).
Emberson, R. M. Macrochelid mites in N.Z. (Acarina: Mesostigmata: Macrochelidae). N. Z. Entomol. 5, 118–127 (1972).
Hughes, A. M. The mites of stored food and houses. (Her Majesty’s Stationary Office, 1976).
O’Connor, B. M. Astigmatid mites (Acari: Sarcoptiformes) of forensic interest. Exp. Appl. Acarol. 49, 125–133 (2009).
Hubert, J., Műnzbergová, Z., Kučerová, Z. & Stejskal, V. Comparison of communities of stored product mites in grain mass and grain residues in the Czech Republic. Exp. Appl. Acarol. 39, 149–158 (2006).
White, N. D. G. Insects, mites and insecticides in stored-grain ecosystems. in Stored-grain ecosystems (eds Jayas, D. S., White, D. G., Muir, W. E.) 123–168 (Marcel Dekker, 1995).
Timms, S., Ferro, D. N. & Emberson, R. M. General biology and nomenclature of Sancassania berlesei (Michael). Acarologia 22, 385–390 (1982).
Early, M. & Goff, M. L. Arthropod succession patterns in exposed carrion on the island of O’ahu, Hawaiian islands, USA. J. Med. Entomol. 23, 520–531 (1986).
Goff, M. L., Early, M., Odom, C. B. & Tullis, K. A preliminary checklist of arthropods associated with exposed carrion in the Hawaiian islands, USA. Proc. Hawaii. Entomol. Soc. 26, 53–58 (1986).
Leclercq, M. & Verstraeten, C. Entomologie et médecine légale. L’entomofaune des cadavres humains: sa succession par son interprétation, ses résultats, ses perspectives. J. Médecine Légale Droit Méd. 36, 205–222 (1993).
Leclercq, M. & Verstraeten, C. Entomologie et médecine légale. Datation de la mort. Acariens trouvés sur des cadavres humains. Bull. Ann. Société R. Belge D’Entomologie 124, 195–200 (1988).
Saloña, M. I. et al. Searching the soil: forensic importance of edaphic fauna after the removal of a corpse. Forensic Sci. 55, 1652–1655 (2010).
Houck, M. A. & O´Connor, B. M. Ecological and evolutionary significance of phoresy in the Astigmata. Annu. Rev. Entomol. 36, 611–636 (1991).
O’Connor, B. M. Evolutionary ecology of astigmatid mites. Annu. Rev. Entomol. 27, 385–409 (1982).
Sarwar, Z. M., Bashir, M. H., Khan, B. S. & Khan, M. A. Description of two new species of genus Caloglyphus Berlese (Acari: Acaridae) from Pakistan. Pak. J. Zool. 45, 101–106 (2013).
Zakhvatkin, A. A. Fauna of U.S.S.R. Arachnoidea Tyroglyphoidea (Acari). (Zoological Institute of the Academy of Sciences of the U.S.S.R., 1941).
Evans, G. O. & Browning, E. British mites of the subfamily Macrochelinae Trägårdh (Gamasina, Macrochelidae). Bull. Br. Mus. Nat. Hist. Zool. 4, 1–55 (1956).
Lindahl, B. D., de Boer, W. & Finlay, R. D. Disruption of root carbon transport into forest humus stimulates fungal opportunists at the expense of mycorrhizal fungi. ISME J. 4, 872–881 (2010).
Middelhoven, W. J. The yeast flora of some decaying mushrooms on trunks of living trees. Folia Microbiol. Praha 49(5), 569–573 (2004).
Martínez-Ramírez, J. A. et al. Studies on drug metabolism by fungi colonizing decomposing human cadavers. Part I: DNA sequence-based identification of fungi isolated from postmortem material. Anal. Bioanal. Chem. 405, 8443–8450 (2013).
Barron, G. L. Nematophagous fungi: Rhopalomyces elegans. Can. J. Bot.- Rev. Can. Bot. 51, 2505–2507 (1973).
Berney, C. et al. Expansion of the ‘reticulosphere’: diversity of novel branching and network-forming amoebae helps to define Variosea (Amoebozoa). Protist 166, 271–295 (2015).
Blandenier, Q. et al. Mycamoeba gemmipara nov. gen., nov. sp., the first cultured member of the environmental Dermamoebidae Clade LKM74 and its unusual life cycle. J. Eukaryot. Microbiol. https://doi.org/10.1111/jeu.12357 (2016).
Carter, D. O., Yellowlees, D. & Tibbett, M. Temperature affects microbial decomposition of cadavers (Rattus rattus) in contrasting soils. Appl. Soil Ecol. 40, 129–137 (2008).
Acsádi, G. & Nemeskéri, J. History of human lifespan and mortality. (Akadémiai Kiadó, 1970).
Buikstra, J. E. & Ubelaker, D. H. Standards for data collection from human skeletal remains. (Arkansas Archeological Survey Research Series No. 44, 1994).
Rösing, F. W. et al. Recommendations for the forensic diagnosis of sex and age from skeletons. HOMO - J. Comp. Hum. Biol. 58, 75–89 (2007).
Ambrose, S. H. Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17, 431–451 (1990).
Longin, R. New method of collagen extraction for radiocarbon dating. Nature 230(5291), 241–242 (1971).
Reimer, P. J., Brown, T. A. & Reimer, R. W. Discussion: reporting and calibration of post-bomb C-14 data. Radiocarbon 46(3), 1299–1304 (2004).
Scheiner, J. D. Spéciation du carbone, de l’azote et du phosphore de différentes boues de stations d’épuration au cours de leurs incubations contrôlées dans deux types de sol, thèse pour obtenir le diplôme de docteur de l’2005. (2005).
Olsen, S. R., Cole, C. V., Watanabe, F. S. & Dean, L. A. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Circ. No 939, 1–19 (1954).
Brown, D. J. F. & Boag, B. An examination of methods used to extract virus-vector nematodes (Nematoda: Longiridae and Trichodoridae) from soil samples. Nematol. Mediterranea 16, 93–99 (1988).
Bongers, T. De nematoden van Nederland: een identificatietabel voor de in Nederland aangetroffen zoetwater- en bodembewonende nematoden. (Stichting Uitgeverij Koninklijke Nederlandse Natuurhistorische Vereniging, 1994).
Scholze, V. S. & Sudhaus, W. A pictorial key to current genus groups of ‘Rhabditidae’. J. Nematode Morphol. Syst. 14, 105–112 (2011).
Krantz, G. W. A manual of acarology. (Oregon State University Book Stores, 1978).
Bregetova, N. G. et al. A key to the soil-inhabiting mites of the Mesostigmata. Leningrad (1977).
Evans, G. O., Sheals, J. G. & MacFarlane, D. The terrestrial Acari of the British Isles: an introduction to their morphology, biology and classification. (British Museum, 1961).
Evans, G. O. & Till, W. M. Mesostigmatic mites of Britain and Ireland (Chelicerata: Acari-Parasitiformes): An introduction to their external morphology and classification. Trans. Zool. Soc. Lond. 35, 139–262 (1979).
Hyatt, K. H. Mites associated with terrestrial beetles in the British Isles. Entomol. Mon. Mag. 126, 1512–1515 (1990).
Hyatt, K. H. Mites of the subfamily Parasitinae (Mesostigmata: Parasitidae) in the British Isles. Bull. Br. Mus. Nat. Hist. Zool. Ser. 38, 237–378 (1980).
Amaral-Zettler, L. A., McCliment, E. A., Ducklow, H. W. & Huse, S. M. A method for studying protistan diversity using massively parallel sequencing of V9 hypervariable regions of small-subunit ribosomal RNA genes. PLoS ONE 4, e6372 (2009).
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. & Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200 (2011).
Guillou, L. et al. The protist ribosomal reference database (PR2): a catalog of unicellular eukaryote small sub-unit rRNA sequences with curated taxonomy. Nucleic Acids Res. 41, D597–D604 (2013).
Mahé, F., Rognes, T., Quince, C., de Vargas, C. & Dunthorn, M. Swarm: robust and fast clustering method for amplicon-based studies. PeerJ 2, e593 (2014).
Legendre, P. & Gallagher, E. Ecologically meaningful transformations for ordination of species data. Oecologia 129, 271–280 (2001).
Husson, F., Josse, J., Lê, S. & Mazet, J. FactoMineR: Multivariate exploratory data analysis and data mining, 2015. http://CRAN.R-project.org/package=FactoMineR. R package version 1.31.4 (2015).
Dufrene, M., Legendre, P., Dufrene, M. & Legendre, P. 1997. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 67(3), 345–366 (1997).
Acknowledgements
This work was funded by research grants to E. Mitchell (Swiss SNF project no. 31003A_163431). We thank N. Moghaddam from the Institute of Forensic Medicine Bern, the team of the Laboratory of Soil Biodiversity Neuchâtel for their help with the analyses, Erik Barr for his comments and the responsible public prosecutor’s department for their cooperation.
Author information
Authors and Affiliations
Contributions
The authors contributed to the different parts of this manuscript as follows: I.S. sampling, soil chemistry, nematodes and coordination of the analyses and manuscript, S.L. bones and hair analyses, C.V.W.S. metabarcoding, numerical analyses, E.L. and D.S. metabarcoding, F.S. nematodes, J.T. sampling and coordination of analyses, M.A.P. mites and revision of the manuscript, E.A.D.M. numerical analyses and revision of the manuscript. All authors checked and commented on the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Szelecz, I., Lösch, S., Seppey, C.V.W. et al. Comparative analysis of bones, mites, soil chemistry, nematodes and soil micro-eukaryotes from a suspected homicide to estimate the post-mortem interval. Sci Rep 8, 25 (2018). https://doi.org/10.1038/s41598-017-18179-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-18179-z
This article is cited by
-
Microbial community composition of food waste before anaerobic digestion
Scientific Reports (2023)
-
A pilot study about the development and characterization of a Roux en Y gastric bypass model in obese Yucatan minipigs
Scientific Reports (2021)
-
Acari community in association with delayed pig carrion decomposition
Experimental and Applied Acarology (2021)
-
Assemblages of Acari in shallow burials: mites as markers of the burial environment, of the stage of decay and of body-cadaver regions
Experimental and Applied Acarology (2021)
-
Potential use of high-throughput sequencing of bacterial communities for postmortem submersion interval estimation
Brazilian Journal of Microbiology (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
