
Abstract
Fish populations can be threatened by distorted sex ratios that arise during sex differentiation. Here we describe sex differentiation in a wild grayling (Thymallus thymallus) population that suffers from distorted sex ratios. We verified that sex determination is linked to the sex determining locus (sdY) of salmonids. This allowed us to study sex-specific gene expression and gonadal development. Sex-specific gene expression could be observed during embryogenesis and was strong around hatching. About half of the fish showed immature testes around eleven weeks after fertilization. This phenotype was mostly replaced by the “testis-to-ovary” or “ovaries” phenotypes during development. The gonads of the remaining fish stayed undifferentiated until six months after fertilization. Genetic sexing revealed that fish with undifferentiated gonads were all males, who grew larger than the genetic females during the observational period. Only 12% of the genetic males showed testicular tissue six months after fertilization. We conclude that sex differentiation starts before hatching, goes through an all-male stage for both sexes (which represents a rare case of “undifferentiated” gonochoristic species that usually go through an all-female stage), and is delayed in males. During these juvenile stages males grow faster than females instead of developing their gonads.
Similar content being viewed by others
Introduction
Fishes show a great diversity of gonadal development and differentiation that can be classified into five categories: (i) gonochoristic species with individuals developing either testes or ovaries, (ii) sequential hermaphrodites that mature as males (protandrous) or (iii) as females (protogynous) and may change sex later in life, (iv) simultaneous hermaphrodites, and (v) all-female species that reproduce gynogenetically1,2. In gonochoristic species, primordial germ cells are typically formed during embryonic or early larval development and subsequently differentiate into male or female gonads under the influence of genetic mechanisms and/or endocrine, environmental, or behavioural signals1. This process can be direct, as in the so-called “differentiated” gonochoristic species3 where primordial germ cells develop into testicular or ovarian tissues. Examples of such development can be found in Arctic charr (Salvelinus alpinus)4 and some cyprinid fishes5,6. However, functional sex reversal can sometimes be induced after gonad formation, for example, with aromatase inhibitors in medaka, Oryzias latipes, and Nile tilapia, Oreochromis niloticus7. In “undifferentiated” gonochoristic species, the typical pattern is that individuals first develop ovarian tissues that may subsequently degenerate, followed by a masculinization of the gonads that finally leads to normal testes, as in zebrafish (Danio rerio) and some other cyprinids8,9. Other species, so-called “secondary” gonochoristics10, seem to first develop into simultaneous hermaphrodites before most individuals mature as only females or males, as in many eel populations Anguilla sp.11,12. We know of no example of an “undifferentiated” gonochoristic species where all individuals first develop immature testicular tissue followed by a feminization of the gonads that finally leads to normal ovaries in females.
Fishes also show a great diversity in sex determination systems that range from purely genetic to purely environmental, and different types of environmentally induced sex reversals have been described in many different orders of the teleosts1,13. The fact that sex determination in fish can be very labile has several practical consequences: it can (i) be exploited in aquaculture where one-sex cultures can sometimes be more profitable14, (ii) be used to control problem species, e.g. invasive species15,16, (iii) be used to boost population growth in the wild17, and (iv) cause some species to be sensitive to environmental changes, especially to different types of endocrine-disrupting pollutants18,19. It is therefore important to understand the diversity of sex determination and sex differentiation among fishes.
We study a population of European grayling (Thymallus thymallus L.) that uses the pre-alpine lake Thun as its feeding habitat and spawns in spring at the outlet of this lake. A yearly monitoring program initiated in the 1940s revealed significantly distorted adult sex ratios (an excess of males) in this population coinciding with an abrupt temperature change in Europe20,21, potentially contributing to the continuous decline of the population22. Grayling belong to the family Salmonidae that include the three subfamilies Salmoninae (e.g. Pacific and Atlantic salmon, trout, char), Thymallinae (grayling), and Coregoninae (whitefish). Salmonids are usually keystone species of their respective habitat and of considerable economic and cultural importance for local communities. Sex determination seems mostly genetic in Salmoninae and Thymallinae, where it may be driven by a master sex-determining gene23. With regard to sex differentiation, much variation is observed among the Salmoninae1, and little is known about the Thymallinae, except that environmental temperature does not seem to have an effect on sex determination in the European grayling (Thymallus thymallus)24.
Here we first verified that the sdY locus that Yano et al.23 tested on 54 graylings from a fish farm in France can be used to predict phenotypic sex in our wild study population. We searched for genotype-phenotype mismatches in (i) in wild-caught breeders, (ii) F1 progeny that were raised to maturity in captivity, and (iii) juvenile fish that were raised in the laboratory under warm or cold conditions. We then experimentally produced half-sib groups and raised the embryos first individually in 24-well plates, then group-wise during their larval and juvenile stages. To detect sex-specific gene expression patterns, we extracted mRNA from embryos, hatchlings, and early larvae. These time periods correspond to the onset of sexual differentiation in another salmonid, the rainbow trout (Oncorhynchus mykiss)25. We also used histological techniques to determine sex differentiation in relation to genetic sex markers over a period of several months.
Methods
Ethical approval
All methods were carried out in accordance with the relevant guidelines and regulations. The project was approved by the veterinary authorities of the cantons of Bern (BE118/14) and Vaud (VD2710; VD2956) and by the fishery inspectorate of the Bern canton.
Verification of genetic sex determination and genetic sexing of larvae
For determining the sex of larvae and juveniles, genomic DNA was extracted from tissue samples (tails or fins) using the DNEasy Blood and Tissue Kit (Qiagen, Hombrechtikon, Switzerland), following manufacturer’s instructions. To verify the efficacy of the sex typing primers, fin clips were taken from a total of 192 adults that were either caught from the study population22 (92 males and 29 females) or F1s that were raised in captivity until sexual maturity (20 males and 49 females). All adults showed the typical sexual dimorphism of this species. They were stripped for their gametes, i.e. phenotypic sex could be verified without dissection. For adult tissue, DNA was extracted using a BioSprint 96 robot tissue extraction kit (Qiagen).
Polymerase chain reaction (PCR) was conducted following protocols by Yano et al.23 with modifications. Briefly, each 15 μl PCR contained 1.5 μL 10X PCR Buffer, 210 μM dNTPs, 1.5 mM MgCl2, 0.3 μM each of primers sdY E1S1 and sdY E2AS4 (male-specific amplification23), 0.075 μM each of primers 18SS and 18S AS ribosomal RNA (positive amplification control23), 0.75 units of Taq Polymerase (Qiagen or Promega GoTaq), and 100 ng of DNA. Thermal cycling consisted of denaturing for 3 min at 95 °C followed by 40 amplification cycles of 94 °C/30 s, 62 °C/30 s, and 72 °C/30 s, with a final extension of 10 min at 72 °C. PCR products were visualized on a 1.5% agarose gel at 100 V for 50 min (Supplementary Fig. S1).
A multiplex reaction with three microsatellite markers previously used to characterize population differentiation in grayling: BFRO005, BFRO006 26 and Ogo2 27 and with ThySex225 was used to analyze the tissue samples of the adults. We designed a small amplicon (~225 bps) of the sdY locus from sequences specific to grayling23 using Primer328 (ThySex225: 5′-AGCCCAGCACTCTTTTCTTATCTC-3′; genbank probe DB accession #: Pr032825786). The 5′ end of the ThySex225 was labelled with a fluorescent label (ATTO532) to aid viewing using capillary gel electrophoresis and to multiplex with microsatellites for high-throughput genotyping. We used the reverse primer sdY E2AS4 of Yano et al.23. Each polymerase chain reaction (PCR) was accomplished in a10µl reaction volume containing 1.5 µl water, 5 µl of Qiagen Hotstar Taq Mix (final concentration 0.5 units of HotStarTaq DNA Polymerase, 1XPCR buffer, 1.5 mM MgCl2 and 200uM of each dNTP), 0.3 µl of each primer (10 µM), and 2 µl of template DNA (1–5ng/µl). The thermal cycling profile consisted of an initial denaturation for 15 min at 95 °C followed by 35 cycles of 94 °C (30 sec), 56 °C reannealing temperature (90 sec) an extension phase at 72 °C (60 sec), and a final extension at 72 °C for 30 min. Fragment lengths were visualized on an Applied Biosystems® (ABI, Life Technologies GmbH, Darmstadt, Germany) 3730 capillary sequencer and alleles were scored using GeneMarker Version 2.6.4 software (SoftGenetics, LLC, State College, PA, USA).
Breeding experiments
Three sets of breeding experiments contribute to the present analyses. For the first breeding experiment, gametes were stripped from 16 wild genitors (8 males and 8 females) and used for block-wise full-factorial breeding with two males and two females per block. The embryos of these 16 half-sib groups were distributed to 24-well plates (Falcon, Becton-Dickinson), with two eggs per well that had been filled with 2 mL of chemically standardized water29. After hatching, in total 532 fish were about equally distributed to eight 200 L aquaria filled with filtered lake water (closed system). Fish were randomly assigned to 4 aquaria at 12 °C and 4 aquaria at 18 °C in separate climate chambers. At 174 days post-fertilization (dpf, i.e. day 145 after peak hatching), two of the aquaria of each temperature treatment were exposed to the parasite Tetracapsuloides bryosalmonae in the course of another study on temperature-dependent pathogen resistance (Uppal et al., unpublished manuscript). Exposure to T. bryosalmonae and the temperature treatment had no effect on any analysis presented here.
Fish were fed ad libitum, initially on a live zooplankton and then on dry food (Skretting, Nutra Brut 3.0, 2.0, T-1.1). At 63–71 days post-exposure, all fish were euthanized with an overdose of Koi Med Sleep (Ethylenglycolmonophenylether) and sexed by visual inspection of the gonads or via the sdY genotype. In total, 60 individuals were both morphologically and genetically sexed to test for possible genotype-phenotype mismatches at this stage.
The second breeding experiment was performed similarly to the first breeding experiment, with the following modifications. The genitors were sampled from a captive breeding stock (F1 progeny of the wild population), and their gametes stripped and used in two full-factorial breeding blocks with 4 females crossed with 5 males each, resulting in 40 different half-sib groups. The embryos were raised individually in 2 mL wells of 24-well plates at 7 °C. At 14 dpf, embryos were exposed either to 1 ng/L 17α-ethinylestradiol (“EE2”), to Pseudomonas fluorescens (“PF”; 106 bacterial cells per well), simultaneously to EE2 and P. fluorescens (“EPF”), or sham-treated (“control”) in the course of a parallel study on EE2-effects on gene expression at different developmental stages30.
In order to induce and synchronize hatching, the temperature was raised to 10 °C at 27 dpf and to 11.5 °C the next day. At 40 dpf, i.e. 11 days after peak hatching, a random sample per treatment was transferred to 8 tanks filled with 200 L lake water. Two of the 8 tanks each were stocked with fish that had been either exposed to EE2, P. fluorescens, EE2 and P. fluorescens, or nothing, respectively. Once per week, 40 L of water per tank was replaced with either filtered lake water (groups “controls” and “PF”) or with filtered lake water to which EE2 had been added to reach a concentration of 1ng/L (groups “EE2” and “EPF”). The larvae were fed with live Artemia, then live copepods and later also dry food as in the first breeding experiment. Temperature was gradually increased to 18 °C towards the end of the study at 163 dpf (in order to simulate the increase of mean temperature in the wild). The treatment with P. fluorescens during embryogenesis did not show any significant effects during larval stages while treatment with EE2 delayed sex differentiation30. EE2-treated individuals were therefore excluded from the present analyses.
The third breeding experiment was accomplished like the second one with a new sample of eggs and sperm collected from the same captive breeding stock, but one year after the second breeding experiment and without any experimental treatments during embryogenesis and larval stages. Its purpose here was to verify an important result of the second breeding study, namely that sex differentiation is more likely to start in genetic females than in genetic males at day 79 dpf.
Larval length was determined by digital analysis of photos taken from freshly killed fish (first and third breeding experiment) or from the fixed and stained sections that were used for histological analyses (second breeding experiment; see below) using ImageJ31.
Sampling and preparations for gene expression analyses
A subset of 5 half-sib groups (1 female crossed with 5 males) from the treatment groups “controls” and “EE2” of the second breeding experiment was used to also study gene expression during early developmental stages. The first samples (12 embryos per family and treatment, i.e. 120 in total) were taken at 21 dpf (Embryo stage). Embryos were immediately transferred to RNAlater (Thermo Scientific, Reinach, Switzerland) and snap frozen at −80 °C. The second sampling occurred the day of hatching (31 dpf; Hatching stage) with 8 larvae per family and treatment (i.e. 80 in total). The third sampling was at day 52 dpf (i.e. 21 days after hatching; First feeding stage) with 5 larvae per family and treatment (40 in total). Larvae of the second and third sampling were euthanized with KoiMed (0.5 mL/L for five minutes) and then decapitated. The heads were immediately stored in RNAlater at −80 °C for later analyses. We sampled heads because the neuroendocrine system is known to play a crucial role in the sexual differentiation1. For example, sex differentiation of the brain is strongly dependent on the local action of estrogenic compounds32.
RNA extractions were performed using the QIAgen 96 RNeasy Universal Tissue Kit (Qiagen) following the manufacturer instructions, except that the centrifugation was done at half of the protocol speed for double the amount of the time (Eppendorf 5804 R centrifuge with an A-2-DWP rotor; Eppendorf, Schönenbuch, Switzerland). In total, three distinct runs of extractions (up to 96 samples each) were performed. Samples from the same treatment, the same family, and collected at the same developmental stage were assigned to the same run of extraction each. RNA was extracted from the whole egg for the first sampling date and from heads only in subsequent samples. Samples were eluted in 100 µL of RNase free water.
The RNA extraction protocol we used did not include a DNase treatment. Therefore, we amplified DNA traces inside the RNA samples to determine the sdY genotype of eggs and fry. We used two amplification protocols using the 18 s gene as an internal control. The first method was used in multiplex for samples with a high amount of DNA. For the samples with low DNA content, the second PCR protocol was used in single reactions with half the amounts of the respective primers each. After genetic sexing, one female and one male per maternal half-sib group and time point was haphazardly chosen for sequencing. In one family, two females were used for the second time point because no male was found in the respective family. In another family, two males were used for the third time point each because no female could be found in the respective family.
The 60 samples designed for sequencing were checked for quality (absorbance ratios and RNA-Quality-Number, RQN) and concentration using both Nanodrop (Thermo Scientific, Reinach, Switzerland) and a Fragment Analyser (Advanced Analytical, Ankeny, USA). All samples were provided for library preparation in an equimolar concentration of 6 ng/µL in 100 µL of RNAse-free water. For each library, 50 µL were used (i.e. 300 ng of RNA). The libraries were prepared in one batch on a robot using the Truseq Stranded RNA protocol (Illumina, Part# 15026495 Rev. A) and multiplexing adaptors. Libraries were then sequenced on an Illumina HiSeq. 2500 machine to produce 2*100 bp paired-end reads. The 60 samples were sequenced in ten lanes with six samples per lane. Both the library preparation and sequencing steps were performed at the Genomic Technologies Facility at the University of Lausanne. See Supplementary Tables S1 and S2 for further details.
Bioinformatics RNA-seq processing
Pairs of reads of 100 bps in length were quality trimmed using fastq-mcf (ea-utils, version 1.1.2; Aronesty, 2013). Low quality reads (mean Phred quality score < 20) and reads containing adapter sequences were trimmed. All the retained reads were truncated and returned with a length of exactly 2*90 bps. An additional quality check using FastQC (version 0.11.2; http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) showed abnormal k-mer frequencies, so we removed ten additional bps at the 5′ end of reads, returning 2*80 bps long reads. This last step was performed with a custom Python script (all scripts available at https://github.com/Oselmoni/GSD_MS), which also corrected the headers of the sequence names in order to allow compatibility with downstream tools. After these processing steps, the quality of each library was rechecked using FastQC.
Twenty-four libraries, i.e. two individuals randomly chosen across each of all 12 possible combinations of developmental stage, sex and treatment, were chosen (Table S3), and duplicated reads were removed with fastq-mcf. The 24 de-duplicated read sets were then merged and used for assembly with Trinity (version 2.0.3; Grabherr et al. 2011). To speed up the assembly, we ran Trinity with the read coverage normalization option set to 50. The assembled transcriptome statistics were provided by Trinity and custom Python scripts.
To exclude spurious transcripts from the assembly, we blasted the assembled sequences against the UniProtKB/Swiss-Prot database (version of October the 16th 2015)33 using the blastx command line application (version 2.2.26)34. For each transcript, only the best hit was kept using a cut-off E-value set at 10−6. The transcripts that did not have any match were filtered out of the assembly. In total, 228,417 transcripts were kept, distributed across 52,353 genes. A similarity search34 (E-value again < 10−6) showed that 99.4% of the coding genes predicted from our assembly aligned against the recently published grayling reference genome (that identified 48,753 coding genes)35.
We used Kallisto (version 0.42)36 to pseudo-map reads to the transcriptome. Next, we summed the estimated counts of the isoforms of each gene by using a custom Python script. The estimated counts per gene were used for differential expression analysis.
Normalization factors for each library were calculated using the Trimmed Mean Method (TMM)37 of the EdgeR package (version 3.12.0)38. A log transformation of count-per-million (log2(cpm)) was then applied to raw expression values. Log transformed count-per-million values were used for a Principal Component Analysis (PCA) of samples39. Identified outlier samples (s24, s25) were filtered out. Additionally, visual inspection of the distribution of the normalized expression values showed that one sample (s58) did not follow the trend of the other samples and was therefore filtered out.
To process the remaining 57 samples, only genes showing an expression of at least 2 cpm in at least five samples were kept (in total, 35,348 genes). Two more PCAs were performed: one to investigate the role of experimental factors (developmental stage, sex, sibgroup, sequencing lane, batch of RNA extraction) in the expression measures of control individuals, and a second one applied to the whole expression matrix (i.e. with both control and EE2 treated individuals). This last PCA allowed us to select the most important factors to include in our model. We considered developmental stage, sex and treatment as a combined variable (with twelve possible levels) and the sib-group as an independent variable. This model included sufficient replication for each comparison: including sib-group inside the “combined block” would not have allowed comparisons between the different levels of this block. For this same reason, technical factors (sequencing lane and batch of RNA purification) were not included in the model.
The differential gene expression analysis was performed using the Bioconductor limma-voom package (version 3.26.3)40,41. We used the voomWithQualityWeight, which calculated sample quality weights in addition to the observational weights of limma-voom42. We used a modified version of the voomWithQualityWeight function performing the cpm transformation using the cpm function from the edgeR package, resulting in a reduced range of minimal expression values across samples (since the minimal cpm values for each sample depend on the size of the library). voomWithQualityWeight was run with a cyclic-loess normalization step and the distribution of the expression values was manually checked (Fig. S2) before proceeding to the differential gene expression analysis.
A linear model was fit for each gene, and coefficients and standard errors were computed for all contrasts of interest using limma. At each developmental stage, we obtained p-values for each gene for the comparison between control males and control females. Q-values43 were obtained from the vector of p-values. A threshold of q = 0.15 was applied to select differentially expressed genes, i.e. a 15% false discovery rate.
In order to have a biologically meaningful annotation of Gene Ontology (GO; Ashburner et al., 2000) terms, we performed a similarity search via blastx command line application (version 2.2.26; National Center for Biotechnology Information, 2008) of the remaining transcripts against a customized database. This customized database contained all the protein sequences from Uniprot (version of December the 15th 2015; Boeackmann et al., 2003) associated to Zebrafish. According to the transcript identifiers of the Trinity assembly, we were able to identify isoforms from the same gene and to keep all the associated Uniprot entries. This information was then processed by a custom Python script: for each gene, we retrieved a list of all the GO terms associated to all blast hits of the respective isoforms. Our annotation concerned only the Biological Function domain of the GO term classifications (Ashburner et al., 2000). The enrichment analysis of GO terms was performed using the goseq package (version 1.22.0; Young et al., 2010) of the R Bioconductor (version 1.20.1; Huber et al., 2015). The list of genes differentially expressed in a contrast were checked for enrichment of GO terms using a Wallenius hypergeometric distribution inspired method (Young et al., 2010). A key feature of goseq is that it takes into account the length of the gene to calculate the enrichment scores (Young et al., 2010). Length of genes was measured by a custom Python script. When a gene showed multiple isoforms, the median isoform length was used as the length of the gene. For each GO term, goseq returns a p- value associated to the score of the enrichment test. These p-values were used to filter and rank GO terms significantly overrepresented in the gene list of interest. The results of the enrichment analysis were then visualized with REVIGO (Supek et al., 2011). For each GO terms enrichment analysis, the REVIGO visualization concerned only the GO terms with a p < 0.05. To assure output readability, if the number of terms having a p < 0.05 was above 150, only the 150 GO terms with the lowest p-value were kept for the REVIGO visualization.
Gonadal development
In total 124 fish were randomly sampled from the second breeding experiment at 51, 79, 107, 135, and 159–163 dpf, i.e. about monthly over the course of 5 months. A further 24 fish were randomly sampled from the third breeding experiment at 79 dpf. These fish were euthanized with an overdose of KoiMed Sleep, the heads stored in RNAlater at −80 °C for further analyses (second breeding experiment only), and the rest of the bodies fixated for two weeks in Davidson solution (AppliChem product No. A3200) for histology. They were transferred to embedding cassettes and dehydrated for 48 h using a Leica TP1020 tissue processor (Leica, Tempe, USA). Dehydrated tissues were flowed in hot paraffin wax (Histoplast P, Serva, Heidelberg, Germany) using a paraffin dispenser embedding (Leica EG1150H, Leica, Tempe, USA) and paraffin wax was finally cooled to obtain a solid paraffin block. Sections were cut at 4 µm ventrally, from anterior to posterior, floated in a water-bath, and collected onto glass slides. Sections were stained with standard Mayer’s haematoxylin and eosin staining (HE-stain44) and cover slipped to be conserved. Fish sections were analysed by light microscopy using a Leitz Aristoplan microscope (Leitz, Wetzlar, FRG) and analysed with an associated digital camera (Color View, Soft Imaging Systems, Münster, FRG) supported by “Analysis software” (Soft Imaging Systems, Münster, FRG).
Data availability
The gene expression data can be downloaded from the NCBI server (Bioproject Accession: PRJNA388031). All other data can be downloaded from the Dryad server (doi:10.5061/dryad.bp12s).
Results
Verification of genetic sex determination
Using the microsatellite multiplex with the Thysex225 locus, we assigned sex to 121 wild-caught adults and to 71 adults from the captive breeding stock. Males have a single peak at 228 bps at this locus whereas females only showed signals for the microsatellite loci (Fig. S1). There was perfect alignment of sex assigned in the field based on morphology and on the presence of eggs and sperm with the multiplex protocol, i.e. there was no ambiguous assignment of sex based on this procedure.
Genetic sexing of 237–245 dpf old juveniles (first breeding experiment) based on the multiplex PCR using sdY primers, and 18 S primers as a positive amplification control, also provided a perfect alignment between gonad morphology and the multiplex protocol with 60 individuals.
Gene expression
RNA quality check, RNA sequencing reads quality check, transcriptome assembly statistics and data preparation for differential gene expression analysis are given in the Supplementary Material. Figure 1 shows the results of a principal component analysis (PCA) on the normalized expression levels matrix of the 28 samples. The strong biological clustering provides a quality control of our measure of expression data, with 79.4% of the variance explained by the first two principal components. Gene expression clustered first by developmental stage, and second by maternal half-sibgroups within each developmental stage (Fig. 1). Inside each developmental stage, sex appeared to best explain the variance in gene expression in the first two principal components in hatchlings (Fig. 1a). This is confirmed in Table 1 that reports the number of genes differentially expressed between males and females at each developmental stages. While only 15 genes were differentially expressed at the embryo stage (Fig. S3), a strong increase of differently expressed genes could be observed on the day of hatching (72% of total genes, Fig. S4). The sex difference remained important at the first feeding stage (3% of total genes, Fig. S5).

Principal component analysis of the gene expression matrix. (a) The first two principal components, explaining 35.0% and 34.7% of the observed variance, respectively, and (b) the next two principle components, explaining 9.5% and 3.3% of the variance, respectively. Round symbols represent females, squared symbols represent males, the colours represent the 5 maternal half-sib families. Ellipses and the grey area in panel b emphasize the clustering by developmental stage.
The GO enrichment analysis of the genes differentially expressed between males and females are shown in the Supplementary Figs S6–S9. At hatching day (Figs S6, S7), the genes identified with sex-specific expression are related to development of the nervous system (e.g. neural crest cell migration, semaphorin-plexin signalling pathway involved in axon guidance, axoneme assembly, etc.). For example, we found that the sexes significantly differed in their expression levels for genes annotated as Serotonin receptor 1B (q < 0.05, logFC* = −1.85) or Neural Cell Adhesion Molecule (q < 0.05, logFC* = 1.78). These annotations were confirmed when aligning these transcripts to the recently published grayling reference genome35. The same analysis at first feeding stage (Figs S8, S9) shows a more heterogeneous landscape of biological processes associated to the genes differentially expressed. We find, for example, GO terms as melanocyte differentiation, cartilage development, cellular response to estrogen stimulus. According to these results, we can hypothesize that a dissimilarity in the development of the nervous system exists between sexes and that it is particularly strong at hatching. However, it is still unclear whether the decrease of this differentiation at first feeding stage is due to a biological reason (e.g., the brain development could converge between sexes) or to a technical one (e.g., other tissues of the head could develop differentially and hide gene expression differences associated to the brain45).
While we should be careful in interpreting single gene patterns, given the draft quality of our transcriptome assembly and the many paralogs due to genome duplications in teleosts and in salmonids46,47, we checked the expression changes between untreated males and females for known sex marker genes. For two such genes, GSDF and DMRT1, we were not able to reconstruct any transcript in our data. One of the transcripts of androgen receptor is 1.5 times more expressed in hatching males than females (p = 0.0052; q = 0.029). Similarly, one of the transcripts of aromatase is marginally more expressed in hatching females than males (1.3 fold-change; p = 0.027; q = 0.066). Most striking, 15 transcripts of cytochrome P450 have some level of differential expression between sexes (uncorrected p < 0.05), of which 12 with a q-value < 0.10, i.e. 11 are probably true positives. Almost all of these are at hatching, although one transcript is 8.3 times more expressed in males than females at first feeding (p = 0.0013; q = 0.087). At hatching, most significant (q < 0.1) P450 transcripts are more expressed in females than males (1.3 to 5.3 fold), although two transcripts are 1.4 and 1.5 fold more expressed in males. Thus for those genes which we are able to detect in our data, changes of expression are consistent with expectations of sex differentiation in fish.
Gonadal development
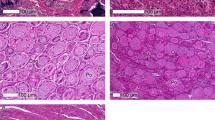
Figure 2 gives an overview of sex differentiation over the 5 sampling periods of the second breeding experiment. The figure also gives the frequency of genetic males and females in the first and the second breeding experiment. All gonads were undifferentiated at the first sampling period (51 dpf). The percent of undifferentiated gonads dropped to 30.4% at the second sampling period (79 dpf). The other fish first showed early testicular tissues while there were no signs of ovarian tissue. From the second to the third sampling day (107 dpf), the percent of fish with testes only dropped from 69.6% to 20.8%, while 45.8% of the fish showed testis-to-ovary or ovaries only. From then until the fifth sampling, the percent of fish with undifferentiated gonads did not decline further, and there was no clear change in the percent of fish with different types of gonadal tissues. All fish that remained undifferentiated at the fourth and fifth sampling period turned out to have the male genotype, while all except three of the remaining fish showed the females genotype (Table 2). See Fig. 3 for examples of the various phenotypes.

Sex differentiation in grayling. Frequencies of fish with undifferentiated gonads (grey bars), gonads with testicular tissue only (blue bars), the testis-to-ovary phenotype (orange bars), and ovaries only (red bars) for the second experiment (sampling periods between 51–163 dpf) when phenotypes were determined by histology, and in the first experiment (sampling period at 237–245 dpf) when phenotypes were determined by morphology. The numbers in the boxes give the total sample sizes for the second experiment, and the number of fish that were both phenotypically and genetically sexed for the first experiment. The blue and red background colors indicate the overall frequencies of genetic males and females, respectively, for each of the two experiments. Phenotype and genotype matched perfectly for the first experiment (sampling period 237–245 dpf). At the last two sampling periods of the second experiment (135 dpf and 159–163 dpf), all fish with undifferentiated gonads had the male genotype, 3 of 5 individuals with testes had the male genotype, all other individuals had the female phenotype. See Table 2 for the match between phenotype and genotype at sampling periods after 135 dpf and Fig. 3 for examples of the various developmental stages.

Representative examples of (a) undifferentiated gonad at fifth sampling period. Gonad consists entirely of undifferentiated cells. (b) Testis in late but still immature stage at third sampling period consists of only few spermatogonia (asterisks) while the majority of germ cells are spermatocytes (SPC). Numerous Sertoli cells are located in the periphery of the spermatocysts (arrowheads). (c) Testis-to-ovary at fourth sampling period. Perinuclear follicles (PNF) are loosely scattered within the testicular tissue. Testis is dominated by spermatocytes (SPC) while spermatogonia (red arrows) are scarce. Some speratids (green asterisk) can be observed within the spermatocysts. (d) Ovary at fifth sampling period: ovary consists completely of perinucleolar follicles (PNF) and a few oogonia (asterisk).
In the third breeding experiment, the frequency of undifferentiated fish at 79 dpf was 79.1%. Their genetic sex ratio was 54.2% males and again not significantly different from a sex ratio of equality (χ2 = 0.2, p = 0.68). The five fish that clearly started to differentiate all showed early testicular tissues. As expected from the second breeding experiment, they all had the female genotype (sex effect on differentiation: χ2 = 9.4, p = 0.002).
Sex difference in growth
In the first breeding experiment (sampling between days 237 and 245 dpf, i.e. 208–216 post hatching peak), males were larger than females (on average 56.5 mm vs 54.9 mm; 95% CI = ±1.8 each, Ntotal = 436), while rearing temperature had no significant effect on size (effects of sex: t = 2.4, p = 0.016; of rearing temperature: t = -0.8, p = 0.45). Males were also heavier than females (t = 1.9, p = 0.05). These findings were confirmed in the second experiment where genetic males were significantly larger than genetic females at the last two sampling periods: male trunks were on average 40.6 mm (95% CI = ±5.3) long, while female trunks were on average 35.3 mm (95% CI = ±5.8) (t = 2.7, p = 0.008). In the third breeding experiment, when fish were only sampled at 79 dpf, there was no sex difference in length or weight (t < 0.96, p > 0.33).
Discussion
It is important to understand sex determination and sex differentiation in fish that play key roles in their respective environment48, that are, or could potentially be, important in aquaculture14, or that already suffer from distorted sex ratios as repeatedly observed in river-dwelling salmonids49. All of this is true for the European grayling, and especially for the population we study22.
As a rule, sex determination is more diverse and also more labile in ray-finned fishes and amphibians than it is in birds and mammals50,51. Sometimes variation in sex determination can be found within a species or a genus52,53. Therefore, even though Yano et al.23 found genetic sex determination in 54 grayling sampled from a fish farm in France, it was necessary to verify their finding in a geographically distinct wild population. We first tested whether sex determination in a wild and in a related captive population is indeed genetic. Our PCR protocols resulted in perfect alignment of sex phenotype and genotype in 192 adults of wild and captive origin, and in 60 juveniles that had been raised at cold and warm temperatures for 8 months. This demonstrates that sex determination has a strong genetic basis in our study population. It also supports Yano et al.’s23 hypothesis that the sdY locus is conserved among two of the three subfamilies of the Salmonidae, namely the Salmoninae and the Thymallinae, and it supports the conclusion of Pompini et al.24 that distorted sex ratios in grayling are not due to temperature-induced sex reversal under ecologically relevant conditions. Because of the clear pattern we found, we were able to use the sdY locus to study sex-specific gene expression at embryonic stages, and sex-specific gonadal development at early juvenile stages.
Sex differentiation is expected to be largely controlled by steroids and gonadotropins, produced mainly by the brain in early life, and later also by gonads1. We found that only a few genes show sex-specific expression in late embryogenesis. However, we cannot exclude the possibility that, by sampling whole embryos instead of heads only, we diluted, and therefore potentially missed, some sex-specific gene expression in the brain. At the time of hatching and when sampling heads only, we found that a very high number of genes showed sex-specific expression. The number of differentially expressed genes dropped towards first feeding but was still high around that developmental stage. Our findings support Baroiller et al.54 who concluded for another salmonid, the coho salmon (O. kisutch), that the maximum sensitivity to an exogenous estrogenic treatment was around hatching time.
We analyzed gene expression in five different paternal half-sib groups that had been experimentally bred and raised to differ only in their paternal contribution, i.e. they differed only genetically (they all shared the same mother and were raised in the same environment). As we sampled only one female and five male breeders, a reliable quantification of the additive genetic effects on gene expression is not yet possible. However, our principle component analysis suggests that family effects are important. This suggests that there is significant heritability in gene expression around hatching. It remains to be tested whether rapid evolution in response to anthropogenic changes of the environment is therefore possible.
The first microscopically detectable characteristic of gonadal differentiation is the migration of the primordial germ cell55. At the time of hatching, the number of primordial germ cells in salmonid gonads is small55. When we analyzed gonadal tissue three weeks after hatching, all gonads were still undifferentiated. Four weeks later, about half of the juveniles showed testicular tissues while the remaining fish showed undifferentiated gonads. The percent of undifferentiated fish remained approximately constant over the next three sampling periods up to about the 19th week after hatching, i.e. the percent of differentiated fish remained about constant, too. Surprisingly, however, the male phenotype was increasingly replaced by female phenotypes among the differentiated fish over the 14 weeks that were covered by the second to the fifth sampling periods. Our third breeding experiment and the sampling of additional male and female juveniles around 7 weeks after hatching confirmed that sex differentiation starts in females and that they first develop immature testes.
Within the Salmoninae, differentiated gonochorism where primordial germ cells develop directly into testis or ovarian tissues has been described in, for example, Arctic charr (Salvelinus alpinus)4, brook charr (Salvelinus fontinalis)55, brown trout (Salmo trutta)56, rainbow trout (O. mykiss)57, and coho salmon (O. kisutch) 58. In contrast, the European grayling, as representative of the Thymallinae, shows a rare form of undifferentiated gonochorism, since undifferentiated gonochoristic species usually go through an all-female stage before they differentiate into testis and ovaries1. We conclude from our observations that the European grayling goes through an all-male stage before developing mature testis and ovaries. Further studies will be necessary to clarify the various processes of morphological sex differentiation in this species.
By the end of the observational period, all fish that still showed undifferentiated gonads had the male genotype and grew faster, i.e. genetic males differentiated later and reached larger body lengths and body weights than genetic females. Delayed male gonad development was also observed in sea trout (Salmo trutta morpha trutta L.), but sex-specific growth was not observed during the early juvenile stages of this species56. As we experimentally raised fish at warm and cold temperatures in our first experiment, we could also test for an interaction between sex-specific growth and water temperature. Such an interaction would potentially help explain the sex-specific juvenile mortality that seems to contribute to the observed correlation between increased water temperatures and population sex ratio22,24. However, we could not find any such effects of temperature on sex ratio under our laboratory conditions. We found no significant sex-specific differences in growth before the onset of sex differentiation (our third breeding experiment with early sampling of juvenile). This suggests that the sex-specific growth patterns are linked to the sex-specific timing of gonad formation.
Among the 5 individuals with testis at the last two sampling times, two individuals were genetic females and would possibly have developed ovaries later. All fish with female gonads (testis-to-ovaries and ovaries) were genetic females. No genetic male showed female gonadal tissue. As mentioned above, the breeding stock we used for the present study derived from a wild population that suffers from male-biased population sex ratios that may contribute to the continuous decline of the population22. Similar observations have been made in other salmonid populations59. Pollution of aquatic systems by endocrine-disrupting compounds (e.g. EE2) is probably ubiquitous wherever humans live. Lake Thun and the grayling spawning ground that is located within the city of Thun should not be an exception. Various cities, villages, and different types of industries discharge their sewage into the lake or into rivers that feed the lake, normally after treatment in sewage treatment facilities that may not be fully effective. Moreover, some of the lake’s whitefish populations (Coregonus sp.) were observed to suffer from increased prevalence of gonadal misdevelopments60,61 that may have been due to anthropogenic disturbances of the ecosystem. However, we found no phenotype-genotype mismatch in any of the adult fish that had been sampled in the wild and in the F1 breeding stock. This suggests that environmental sex reversal and its possible effects on the next generation18 do not, by themselves, explain the distorted population sex ratios.
In conclusion, the sex-specific growth patterns and the differences in the timing of gonadal differentiation that we observed here demonstrate sex-specific life histories of juvenile grayling during their first year of life. Such significant sex differences may promote sex-specific juvenile mortality in response to various types of environmental changes. The male-biased sex-ratio distortions of the study population22 suggest that juvenile mortality has become significantly sex-specific over a period of at least two decades from 1988 on, i.e. after a cascade of abrupt environmental changes that seem to have led to the regime shift described by Reid et al.21.
References
Devlin, R. H. & Nagahama, Y. Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental influences. Aquaculture 208, 191–364, https://doi.org/10.1016/s0044-8486(02)00057-1 (2002).
Wootton, R. J. & Smith, C. Reproductive biology of teleost fishes. (Wiley-Blackwell, 2014).
Yamamoto, T. in Fish physiology (eds W. Hoar & D. Randall) 117–175 (Academic Press, 1969).
Chiasson, M. & Benfey, T. J. Gonadal differentiation and hormonal sex reversal in arctic charr (Salvelinus alpinus). Journal of Experimental Zoology Part a-Ecological Genetics and Physiology 307A, 527–534, https://doi.org/10.1002/jez.407 (2007).
Komen, J., Yamashita, M. & Nagahama, Y. Testicular development induced by a recessive mutation during gonadal differentiation of female common carp (Cyprinus carpio L.). Development Growth & Differentiation 34, 535–544 (1992).
Talikina, M. G. Sex differentiation and gonad development during the first years of life in the bream Abramis brama from the Rybinsk water reservoir. Voprosy Ikhtiologii 35, 114–119 (1995).
Paul-Prasanth, B. et al. Estrogen oversees the maintenance of the female genetic program in terminally differentiated gonochorists. Sci. Rep. 3, https://doi.org/10.1038/srep02862 (2013).
Takahashi, H. Juvenile hermaphroditism in the zebrafish Brachydanio rerio. Bulletin of the Faculty of Fisheries Hokkaido University 28, 57–65 (1977).
Takahashi, H. & Shimizu, M. Juvenile inter sexuality in a cyprinid fish the Sumatra barb Barbus tetrazona tetrazona. Bulletin of the Faculty of Fisheries Hokkaido University 34, 69–78 (1983).
Atz, J. W. in Intersexuality in vertebrates including man (eds C. N. Armstrong & A. J. Marshall) 145–232 (Academic Press, 1964).
Beullens, K. et al. Gonadal differentiation, intersexuality and sex ratios of European eel (Anguilla anguilla L) maintained in captivity. Aquaculture 153, 135–150, https://doi.org/10.1016/s0044-8486(97)00018-5 (1997).
Colombo, G. & Grandi, G. Histological study of the development and sex differentiation of the gonad in the European eel. J. Fish Biol. 48, 493–512, https://doi.org/10.1111/j.1095-8649.1996.tb01443.x (1996).
Stelkens, R. B. & Wedekind, C. Environmental sex reversal, Trojan sex genes, and sex ratio adjustment: conditions and population consequences. Mol. Ecol. 19, 627–646, https://doi.org/10.1111/j.1365-294X.2010.04526.x (2010).
Piferrer, F. Endocrine sex control strategies for the feminization of teleost fish. Aquaculture 197, 229–281, https://doi.org/10.1016/s0044-8486(01)00589-0 (2001).
Gutierrez, J. B. & Teem, J. L. A model describing the effect of sex-reversed YY fish in an established wild population: the use of a Trojan Y chromosome to cause extinction of an introduced exotic species. J. theor. Biol. 241, 333–341 (2006).
Cotton, S. & Wedekind, C. Control of introduced species using Trojan sex chromosomes. Trends Ecol. Evol. 22, 441–443 (2007).
Cotton, S. & Wedekind, C. Introduction of Trojan sex chromosomes to boost population growth. J. theor. Biol. 249, 153–161, https://doi.org/10.1016/j.jtbi.2007.07.016 (2007).
Cotton, S. & Wedekind, C. Population consequences of environmental sex reversal. Cons. Biol. 23, 196–206, https://doi.org/10.1111/j.1523-1739.2008.01053.x (2009).
Lange, A., Paull, G. C., Hamilton, P. B., Iguchi, T. & Tyler, C. R. Implications of persistent exposure to treated wastewater effluent for breeding in wild roach (Rutilus rutilus) populations. Environ. Sci. Technol. 45, 1673–1679, https://doi.org/10.1021/es103232q (2011).
Wedekind, C. & Küng, C. Shift of spawning season and effects of climate warming on developmental stages of a grayling (Salmonidae). Cons. Biol. 24, 1418–1423 (2010).
Reid, P. C. et al. Global impacts of the 1980s regime shift. Global Change Biology 22, 682–703, https://doi.org/10.1111/gcb.13106 (2016).
Wedekind, C. et al. Persistent unequal sex ratio in a population of grayling (Salmonidae) and possible role of temperature increase. Cons. Biol. 27, 229–234, https://doi.org/10.1111/j.1523-1739.2012.01909.x (2013).
Yano, A. et al. The sexually dimorphic on the Y-chromosome gene (sdY) is a conserved male-specific Y-chromosome sequence in many salmonids. Evol. Appl. 6, 486–496, https://doi.org/10.1111/eva.12032 (2013).
Pompini, M. et al. Temperature-induced sex reversal is not responsible for sex-ratio distortions in grayling Thymallus thymallus or brown trout Salmo trutta. J. Fish Biol. 83, 404–411, https://doi.org/10.1111/jfb.12174 (2013).
Krisfalusi, M. & Cloud, J. G. Gonadal sex reversal in triploid rainbow trout (Oncorhynchus mykiss). Journal of Experimental Zoology 284, 466–472, https://doi.org/10.1002/(sici)1097-010x(19990901)284:4<466::aid-jez13>3.0.co;2-g (1999).
Sušnik, S., Snoj, A. & Dovč, P. Microsatellites in grayling (Thymallus thymallus): comparison of two geographically remote populations from the Danubian and Adriatic river basin in Slovenia. Mol. Ecol. 8, 1756–1758 (1999).
Olsen, J. B., Bentzen, P. & Seeb, J. E. Characterization of seven microsatellite loci derived from pink salmon. Mol. Ecol. 7, 1087–1089 (1998).
Untergasser, A. et al. Primer3 - new capabilities and interfaces. Nucleic Acids Res. 40, e115 (2012).
OECD. OECD guideline for the testing of chemicals 203 (fish acute toxicity test), Annex 2. 9., http://www.oecd.org/ (1992).
Selmoni, O. M. et al. Sex-specific changes in gene expression and delayed sex differentiation in response to estrogen pollution in grayling (Salmonidae). bioRxiv, 135210, https://doi.org/10.1101/135210 (2017).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9, 671–675 (2012).
Diotel, N. et al. Aromatase in the brain of teleost fish: Expression, regulation and putative functions. Front. Neuroendocrinol. 31, 172–192, https://doi.org/10.1016/j.yfrne.2010.01.003 (2010).
Boeckmann, B. The SWISS-PROT protein knowledgebase and its supplement TrEMBL in 2003. Nucleic Acids Res. 31, 365–370, https://doi.org/10.1093/nar/gkg095 (2003).
Altschul, S. F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402, https://doi.org/10.1093/nar/25.17.3389 (1997).
Varadharajan, S. et al. The grayling genome reveals selection on gene expression regulation after whole genome duplication. bioRxiv, https://doi.org/10.1101/153270 (2017).
Bray, N., Pimentel, H., Melsted, P. & Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 34, 525–527, https://doi.org/10.1038/nbt.3519 (2016).
Robinson, M. D. & Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 11, R25, https://doi.org/10.1186/gb-2010-11-3-r25 (2010).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics (Oxford, England) 26, 139–140, https://doi.org/10.1093/bioinformatics/btp616 (2010).
Yeung, K. Y. & Ruzzo, W. L. Principal component analysis for clustering gene expression data. Bioinformatics 17, 763–774, https://doi.org/10.1093/bioinformatics/17.9.763 (2001).
Law, C. W., Chen, Y., Shi, W. & Smyth, G. K. voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 15, R29, https://doi.org/10.1186/gb-2014-15-2-r29 (2014).
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47, https://doi.org/10.1093/nar/gkv007 (2015).
Liu, R. et al. Why weight? Modelling sample and observational level variability improves power in RNA-seq analyses. Nucleic Acids Res. 43, e97, https://doi.org/10.1093/nar/gkv412 (2015).
Storey, J. D. The positive false discovery rate: a Bayesian interpretation and the q-value. The Annals of Statistics 31, 2013–2035 (2003).
Mulisch, M. & Welsch, U. Romeis - Mikroskopische Technik, 18th Edition. (Spektrum Akademischer Verlag, 2010).
Pantalacci, S. & Semon, M. Transcriptomics of developing embryos and organs: A raising tool for evo-devo. Journal of Experimental Zoology Part B-Molecular and Developmental Evolution 324, 363–371, https://doi.org/10.1002/jez.b.22595 (2015).
Jaillon, O. et al. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 431, 946–957, https://doi.org/10.1038/nature03025 (2004).
Lien, S. et al. The Atlantic salmon genome provides insights into rediploidization. Nature 533, 200–205, https://doi.org/10.1038/nature17164 (2016).
Wedekind, C. Demographic and genetic consequences of disturbed sex determination. Phil. Trans. R. Soc. B 372, 20160326, https://doi.org/10.1098/rstb.2016.0326 (2017).
Thompson, N. F. et al. Sex reversal, selection against hatchery females or wild males does not explain differences in sex ratio between first generation hatchery and wild steelhead. Oncorhynchus mykiss. Environmental Biology of Fishes 98, 113–120, https://doi.org/10.1007/s10641-014-0240-0 (2015).
Beukeboom, L. W. & Perrin, N. The evolution of sex determination. (Oxford University Press, 2014).
Perrin, N. Random sex determination: When developmental noise tips the sex balance. Bioessays 38, 1218–1226, https://doi.org/10.1002/bies.201600093 (2016).
Rodrigues, N., Vuille, Y., Loman, J. & Perrin, N. Sex-chromosome differentiation and ‘sex races’ in the common frog (Rana temporaria). Proc. R. Soc. B Biol. Sci. 282, https://doi.org/10.1098/rspb.2014.2726 (2015).
Geffroy, B. & Bardonnet, A. Sex differentiation and sex determination in eels: consequences for management. Fish Fish. 17, 375–398, https://doi.org/10.1111/faf.12113 (2016).
Baroiller, J. F., Guiguen, Y. & Fostier, A. Endocrine and environmental aspects of sex differentiation in fish. Cell. Mol. Life Sci. 55, 910, https://doi.org/10.1007/s000180050344 (1999).
Sacobie, C. F. D. & Benfey, T. J. Sex differentiation and early gonadal development in brook trout. N. Am. J. Aquac. 67, 181–186, https://doi.org/10.1577/a04-059.1 (2005).
Dziewulska, K. & Domagala, J. Testicular development in the sea trout (Salmo trutta morpha trutta L.) after sex differentiation, with a reference to precocious maturation. Journal of Applied Ichthyology 20, 282–289, https://doi.org/10.1111/j.1439-0426.2004.00553.x (2004).
Xu, G. et al. Morphology, sex steroid level and gene expression analysis in gonadal sex reversal of triploid female (XXX) rainbow trout (Oncorhynchus mykiss). Fish Physiology and Biochemistry 42, 193–202, https://doi.org/10.1007/s10695-015-0129-7 (2016).
Piferrer, F. & Donaldson, E. M. Gonadal differentiation in Coho salmon, Oncorhynchus kisutch, after a single treatment with androgen or estrogen at different stages during ontogenesis. Aquaculture 77, 251–262 (1989).
Clews, E., Durance, I., Vaughan, I. P. & Ormerod, S. J. Juvenile salmonid populations in a temperate river system track synoptic trends in climate. Global Change Biology 16, 3271–3283, https://doi.org/10.1111/j.1365-2486.2010.02211.x (2010).
Bernet, D., Wahli, T., Küng, C. & Segner, H. Frequent and unexplained gonadal abnormalities in whitefish (central alpine Coregonus sp.) from an alpine oligotrophic lake in Switzerland. Diseases of Aquatic Organisms 61, 137–148 (2004).
Urbach, D. et al. Gonadal alterations in male whitefish Coregonus fatioi: no evidence for genetic damage reducing viability in early life stages. Diseases of Aquatic Organisms 81, 119–125, https://doi.org/10.3354/dao01952 (2008).
Acknowledgements
We thank B. Bracher, U. Gutmann, C. Küng, R. Mani, M. Schmid, and T. Vuille from the Fishery Inspectorate Bern for permissions and provision of fish for tissue sampling and experimental fertilizations, the Pachtverein Thun for catching the wild fish, the Saint-Sulpice pump station for zooplankton supplies, T. Braunbeck for access to the histology laboratory, C. Berney, T. Bösch, J. Buser, E. Clark, B. des Monstiers, M. dos Santos, J. Kast, C. Luca, Y. Marendaz, D. Nusbaumer, E. Pereira Alvarez, M. Pompini, A.-L. Roulin, V. Sentchilo, R. Sermier, E. Vermeirssen, B. von Siebenthal, D. Zeugin, and the staff at Ecogenics GmbH (Switzerland) for assistance in the field or the laboratory, and two reviewers for helpful comments. This project was financially supported by the Swiss Federal Office for the Environment and the Swiss National Science Foundation (31003A_159579).
Author information
Authors and Affiliations
Contributions
M.R.R. and C.W. initiated the project. D.M., O.S., A.U., L.M.C., L.W., I.C., and C.W. sampled the adult fish, did the experimental in vitro fertilizations, and prepared the embryos for experimental rearing in the laboratory. All further manipulations on the embryos and the larvae were done by D.M., O.S., A.U., L.M.C., I.C., and L.W. The RNA-seq data were analyzed by O.S., J.R., and M.R.R., the histological analyses were done by D.M., supervised by S.K., and the molecular genetic sexing was performed by D.M., O.S., A.U., and K.B.M. D.M., O.S., and C.W. performed the remaining statistical analyses and wrote the first version of the manuscript that was then critically revised by all other authors.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Maitre, D., Selmoni, O.M., Uppal, A. et al. Sex differentiation in grayling (Salmonidae) goes through an all-male stage and is delayed in genetic males who instead grow faster. Sci Rep 7, 15024 (2017). https://doi.org/10.1038/s41598-017-14905-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-14905-9
This article is cited by
-
Activation of stress response axis as a key process in environment-induced sex plasticity in fish
Cellular and Molecular Life Sciences (2020)
-
Sex-specific changes in gene expression in response to estrogen pollution around the onset of sex differentiation in grayling (Salmonidae)
BMC Genomics (2019)
-
Low adaptive potential for tolerance to ethynylestradiol, but also low toxicity, in a grayling population (Thymallus thymallus)
BMC Evolutionary Biology (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
