
Abstract
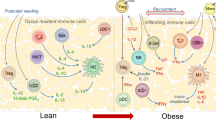
Visceral adipose tissue (VAT) is an energy store and endocrine organ critical for metabolic homeostasis. Regulatory T (Treg) cells restrain inflammation to preserve VAT homeostasis and glucose tolerance. Here, we show that the VAT harbors two distinct Treg cell populations: prototypical serum stimulation 2-positive (ST2+) Treg cells that are enriched in males and a previously uncharacterized population of C–X–C motif chemokine receptor 3-positive (CXCR3+) Treg cells that are enriched in females. We show that the transcription factors GATA-binding protein 3 and peroxisome proliferator-activated receptor-γ, together with the cytokine interleukin-33, promote the differentiation of ST2+ VAT Treg cells but repress CXCR3+ Treg cells. Conversely, the differentiation of CXCR3+ Treg cells is mediated by the cytokine interferon-γ and the transcription factor T-bet, which also antagonize ST2+ Treg cells. Finally, we demonstrate that ST2+ Treg cells preserve glucose homeostasis, whereas CXCR3+ Treg cells restrain inflammation in lean VAT and prevent glucose intolerance under high-fat diet conditions. Overall, this study defines two molecularly and developmentally distinct VAT Treg cell types with unique context- and sex-specific functions.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
Sequencing data generated for this study have been deposited in the Gene Expression Omnibus (GEO) database with accession number GSE205427. All other data and materials are available upon request.
References
Rosen, E. D. & Spiegelman, B. M. What we talk about when we talk about fat. Cell 156, 20–44 (2014).
Nawrocki, A. R. & Scherer, P. E. The delicate balance between fat and muscle: adipokines in metabolic disease and musculoskeletal inflammation. Curr. Opin. Pharmacol. 4, 281–289 (2004).
Ouchi, N., Parker, J. L., Lugus, J. J. & Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 11, 85–97 (2011).
Man, K., Kallies, A. & Vasanthakumar, A. Resident and migratory adipose immune cells control systemic metabolism and thermogenesis. Cell. Mol. Immunol. 19, 421–431 (2022).
Feuerer, M. et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med. 15, 930–939 (2009).
Cipolletta, D. et al. PPAR-γ is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature 486, 549–553 (2012).
Cipolletta, D., Cohen, P., Spiegelman, B. M., Benoist, C. & Mathis, D. Appearance and disappearance of the mRNA signature characteristic of Treg cells in visceral adipose tissue: age, diet, and PPARγ effects. Proc. Natl Acad. Sci. USA 112, 482–487 (2015).
Bapat, S. P. et al. Depletion of fat-resident Treg cells prevents age-associated insulin resistance. Nature 528, 137–141 (2015).
Beppu, L. Y. et al. Tregs facilitate obesity and insulin resistance via a Blimp-1/IL-10 axis. JCI Insight https://doi.org/10.1172/jci.insight.140644 (2021).
Wu, D. et al. T reg-specific insulin receptor deletion prevents diet-induced and age-associated metabolic syndrome. J. Exp. Med. https://doi.org/10.1084/jem.20191542 (2020).
Vasanthakumar, A. et al. The transcriptional regulators IRF4, BATF and IL-33 orchestrate development and maintenance of adipose tissue-resident regulatory T cells. Nat. Immunol. 16, 276–285 (2015).
Vasanthakumar, A. et al. Sex-specific adipose tissue imprinting of regulatory T cells. Nature 579, 581–585 (2020).
Molofsky, A. B. et al. Interleukin-33 and interferon-γ counter-regulate group 2 innate lymphoid cell activation during immune perturbation. Immunity 43, 161–174 (2015).
Kolodin, D. et al. Antigen- and cytokine-driven accumulation of regulatory T cells in visceral adipose tissue of lean mice. Cell Metab. 21, 543–557 (2015).
Han, J. M. et al. IL-33 reverses an obesity-induced deficit in visceral adipose tissue ST2+ T regulatory cells and ameliorates adipose tissue inflammation and insulin resistance. J. Immunol. 194, 4777–4783 (2015).
Li, C. et al. TCR transgenic mice reveal stepwise, multi-site acquisition of the distinctive fat-Treg phenotype. Cell 174, 285–299 (2018).
Sidwell, T. et al. Attenuation of TCR-induced transcription by Bach2 controls regulatory T cell differentiation and homeostasis. Nat. Commun. 11, 252 (2020).
Delacher, M. et al. Precursors for nonlymphoid-tissue Treg cells reside in secondary lymphoid organs and are programmed by the transcription factor BATF. Immunity 52, 295–312 (2020).
Vasanthakumar, A. & Kallies, A. The regulatory T cell: jack-of-all-trades. Trends Immunol. 36, 756–758 (2015).
Li, Y. et al. Insulin signaling establishes a developmental trajectory of adipose regulatory T cells. Nat. Immunol. 22, 1175–1185 (2021).
Mathis, D. Immunological goings-on in visceral adipose tissue. Cell Metab. 17, 851–859 (2013).
Wensveen, F. M. et al. NK cells link obesity-induced adipose stress to inflammation and insulin resistance. Nat. Immunol. 16, 376–385 (2015).
Wang, H. et al. Adipose group 1 innate lymphoid cells promote adipose tissue fibrosis and diabetes in obesity. Nat. Commun. 10, 3254 (2019).
O’Sullivan, T. E. et al. Adipose-resident group 1 innate lymphoid cells promote obesity-associated insulin resistance. Immunity 45, 428–441 (2016).
Spallanzani, R. G. et al. Distinct immunocyte-promoting and adipocyte-generating stromal components coordinate adipose tissue immune and metabolic tenors. Sci. Immunol. https://doi.org/10.1126/sciimmunol.aaw3658 (2019).
Burzyn, D., Benoist, C. & Mathis, D. Regulatory T cells in nonlymphoid tissues. Nat. Immunol. 14, 1007–1013 (2013).
Wohlfert, E. A. et al. GATA3 controls Foxp3+ regulatory T cell fate during inflammation in mice. J. Clin. Invest. 121, 4503–4515 (2011).
Kalekar, L. A. et al. Regulatory T cells in skin are uniquely poised to suppress profibrotic immune responses. Sci. Immunol. https://doi.org/10.1126/sciimmunol.aaw2910 (2019).
Hayakawa, M. et al. T-helper type 2 cell-specific expression of the ST2 gene is regulated by transcription factor GATA-3. Biochim. Biophys. Acta 1728, 53–64 (2005).
Schiering, C. et al. The alarmin IL-33 promotes regulatory T-cell function in the intestine. Nature 513, 564–568 (2014).
Wei, G. et al. Genome-wide analyses of transcription factor GATA3-mediated gene regulation in distinct T cell types. Immunity 35, 299–311 (2011).
Koch, M. A. et al. The transcription factor T-bet controls regulatory T cell homeostasis and function during type 1 inflammation. Nat. Immunol. 10, 595–602 (2009).
Yu, F., Sharma, S., Edwards, J., Feigenbaum, L. & Zhu, J. Dynamic expression of transcription factors T-bet and GATA-3 by regulatory T cells maintains immunotolerance. Nat. Immunol. 16, 197–206 (2015).
Levine, A. G. et al. Stability and function of regulatory T cells expressing the transcription factor T-bet. Nature 546, 421–425 (2017).
Tan, T. G., Mathis, D. & Benoist, C. Singular role for T-BET+CXCR3+ regulatory T cells in protection from autoimmune diabetes. Proc. Natl Acad. Sci. USA 113, 14103–14108 (2016).
Hall, A. O. et al. The cytokines interleukin 27 and interferon-γ promote distinct Treg cell populations required to limit infection-induced pathology. Immunity 37, 511–523 (2012).
Stolarczyk, E. et al. Improved insulin sensitivity despite increased visceral adiposity in mice deficient for the immune cell transcription factor T-bet. Cell Metab. 17, 520–533 (2013).
Kim, J. M., Rasmussen, J. P. & Rudensky, A. Y. Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice. Nat. Immunol. 8, 191–197 (2007).
Caton, M. L., Smith-Raska, M. R. & Reizis, B. Notch–RBP-J signaling controls the homeostasis of CD8− dendritic cells in the spleen. J. Exp. Med. 204, 1653–1664 (2007).
Outtz, H. H., Tattersall, I. W., Kofler, N. M., Steinbach, N. & Kitajewski, J. Notch1 controls macrophage recruitment and Notch signaling is activated at sites of endothelial cell anastomosis during retinal angiogenesis in mice. Blood 118, 3436–3439 (2011).
Lo, C.-M. et al. Cholecystokinin knockout mice are resistant to high-fat diet-induced obesity. Gastroenterology 138, 1997–2005 (2010).
Lima, W. G., Martins-Santos, M. E. S. & Chaves, V. E. Uric acid as a modulator of glucose and lipid metabolism. Biochimie 116, 17–23 (2015).
Park, W. Y. et al. PEX13 is required for thermogenesis of white adipose tissue in cold-exposed mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1867, 159046 (2022).
Wang, Y. et al. METTL3 is essential for postnatal development of brown adipose tissue and energy expenditure in mice. Nat. Commun. 11, 1648 (2020).
Arpaia, N. et al. A distinct function of regulatory T cells in tissue protection. Cell 162, 1078–1089 (2015).
Pierson, W. et al. Antiapoptotic Mcl-1 is critical for the survival and niche-filling capacity of Foxp3+ regulatory T cells. Nat. Immunol. 14, 959–965 (2013).
Hwang, E. S., Szabo, S. J., Schwartzberg, P. L. & Glimcher, L. H. T helper cell fate specified by kinase-mediated interaction of T-bet with GATA-3. Science 307, 430–433 (2005).
Groom, J. R. et al. CXCR3 chemokine receptor–ligand interactions in the lymph node optimize CD4+ T helper 1 cell differentiation. Immunity 37, 1091–1103 (2012).
Oboki, K. et al. IL-33 is a crucial amplifier of innate rather than acquired immunity. Proc. Natl Acad. Sci. USA 107, 18581–18586 (2010).
Zhu, J. et al. The transcription factor T-bet is induced by multiple pathways and prevents an endogenous Th2 cell program during Th1 cell responses. Immunity 37, 660–673 (2012).
Oghumu, S. et al. Distinct populations of innate CD8+ T cells revealed in a CXCR3 reporter mouse. J. Immunol. 190, 2229–2240 (2013).
Shi, C. et al. Bone marrow mesenchymal stem and progenitor cells induce monocyte emigration in response to circulating toll-like receptor ligands. Immunity 34, 590–601 (2011).
Notini, A. J., Davey, R. A., McManus, J. F., Bate, K. L. & Zajac, J. D. Genomic actions of the androgen receptor are required for normal male sexual differentiation in a mouse model. J. Mol. Endocrinol. 35, 547–555 (2005).
Roediger, B. et al. Cutaneous immunosurveillance and regulation of inflammation by group 2 innate lymphoid cells. Nat. Immunol. 14, 564–573 (2013).
Anderson, K. G. et al. Intravascular staining for discrimination of vascular and tissue leukocytes. Nat. Protoc. 9, 209–222 (2014).
Liao, Y., Smyth, G. K. & Shi, W. The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 41, e108 (2013).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Law, C. W., Chen, Y., Shi, W. & Smyth, G. K. voom: precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 15, R29 (2014).
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015).
McCarthy, D. J. & Smyth, G. K. Testing significance relative to a fold-change threshold is a TREAT. Bioinformatics 25, 765–771 (2009).
Young, M. D., Wakefield, M. J., Smyth, G. K. & Oshlack, A. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 11, R14 (2010).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Lun, A. T. L. et al. EmptyDrops: distinguishing cells from empty droplets in droplet-based single-cell RNA sequencing data. Genome Biol. 20, 63 (2019).
Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888–1902 (2019).
Hafemeister, C. & Satija, R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 20, 296 (2019).
van den Brink, S. C. et al. Single-cell sequencing reveals dissociation-induced gene expression in tissue subpopulations. Nat. Methods 14, 935–936 (2017).
Soh, Y. Q. S. et al. Sequencing the mouse Y chromosome reveals convergent gene acquisition and amplification on both sex chromosomes. Cell 159, 800–813 (2014).
Cao, J. et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 566, 496–502 (2019).
Ji, Z. & Ji, H. TSCAN: pseudo-time reconstruction and evaluation in single-cell RNA-seq analysis. Nucleic Acids Res. 44, e117 (2016).
Bunis, D. G., Andrews, J., Fragiadakis, G. K., Burt, T. D. & Sirota, M. dittoSeq: universal user-friendly single-cell and bulk RNA sequencing visualization toolkit. Bioinformatics https://doi.org/10.1093/bioinformatics/btaa1011 (2020).
Acknowledgements
This work was funded by the National Health and Medical Research Council Ideas Grants and Investigator Grants to A.K., A.V. (GNT2012119) and J.R.G. (GNT2007812) and the Australia Research Council Discovery Grant to A.K. S.V.T. and D.M. acknowledge the University of Melbourne for providing a Melbourne Research Scholarship to support the conduct of this study. M.B. and T.E. were supported by the Deutsche Forschungsgemeinschaft (German Research Foundation, IRTG2168 272482170, SFB 1454 TP16 432325352). We would like to thank S. Turner (Monash University), D. Strugnell (Peter Doherty Institute) and N. Wang (Peter Doherty Institute) for mice and S. Wilcox, J. Bayliss and M. Köhne for technical help. We acknowledge the Melbourne Cytometry Platform for the provision of flow cytometry services. This work was supported by infrastructure and technical assistance from the Melbourne Mouse Metabolic Phenotyping Platform at the University of Melbourne.
Author information
Authors and Affiliations
Contributions
A.K. and A.V. designed the project, interpreted results and wrote the manuscript. S.V.T. and K.M. performed most of the work, analyzed data, edited the manuscript and made figure panels. T.E. performed scRNAseq and analyzed the data, with assistance from M. Beyer. M. Becker, M.H.S. and D.H. J.W. and B.P. performed RNAseq with low cell input. D.C., Y.L. and W.S. analyzed all the bulk RNAseq and ATACseq datasets. A.T. performed flow cytometry, and D.M. performed HFD experiments. G.L.G. and M.P. analyzed multiple reporter mouse strains. J.R.G. provided reagents (mice). S.R.M. and C.A.A. analyzed the expression of various chemokine receptors. V.H. and M.J.W. performed metabolic experiments and interpreted the results. M.B. interpreted the scRNAseq data and edited the manuscript. T.M., C.T. and K.B. provided help with multiple experiments.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks Dirk Brenner and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available. Primary Handling Editor: N. Bernard, in collaboration with the Nature Immunology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Heterogeneity of Treg cells in male and female VAT.
a, Gating strategy to identify and isolate CD4+ T cells in the VAT. b, Single cell RNA sequencing (scRNAseq) heatmap showing expression of signature transcripts for VAT Treg cell clusters 1-5 (yellow denotes upregulated genes). c, Heat map showing expression of selected genes in clusters 1, 2 and 3-6 (red - high, blue - low expression). d, Bar graph showing percentages of cell clusters in the Treg cell compartment in male and female mice identified by scRNAseq. e, Flow cytometric analysis of 25-week-old male and female Cxcr3CIBER mice showing ST2 and CXCR3 expression in VAT Treg cells. f, Volcano plot shows differential expression of genes between ST2+ and CXCR3+ VAT Treg cells (FDR 0.2). g, Graphs show frequencies of ST2+ (left) and CXCR3+ (right) VAT Treg cells in different adipose tissue depots of male (VAT – visceral adipose tissue, sAT – subcutaneous adipose tissue, pAT – perinephral adipose tissue) (n = 10 VAT, n = 6 sAT, n = 5 pAT) and female (n = 9 VAT, n = 8 sAT and pAT each). Symbols represent individual mice; horizontal lines indicate means. Data are representative (e) or pooled (g) from at least two independent experiments. Error bars indicate the standard deviation. Statistical analyses were performed using two-way ANOVA (g).
Extended Data Fig. 2 Impact of age and physiology on ST2+ and CXCR3+ VAT Treg cells.
a, VAT Treg cells from 7-week-old male and female mice were analysed. ST2 and CXCR3 expression in VAT Treg cells (left). Frequencies (centre) and numbers (right) of ST2+ and CXCR3+ cells (n = 10 mice of each sex). P (****) < 0.0001. b, Frequencies of VAT Treg cells from female (red) and male (blue) mice at different ages. Females n = 94; males n = 94. c, d Correlation between body weight (top) or VAT weight (bottom) and ST2+ (c) and CXCR3+ (d) VAT Treg cell numbers from 25–30-week-old mice. (c) Females n = 24 (top), 21 (bottom); males n = 38 (top), 33 (bottom). (d) Females n = 13 (top), 30 (bottom); males n = 15 (top), 32 (bottom). e, Graphs show frequencies (top) and numbers (bottom) of total VAT Treg cells from normal diet (ND) and high fat diet (HFD) fed females and males (n = 10 mice each sex) f, Flow cytometry histograms show CD73 expression in DN, CXCR3+ and ST2+ VAT Treg cells from male and female mice (left) and quantification (right). Females n = 9; males n = 10. g, Circulatory cells in male mice were stained with an intravascular (i.v.) CD45 antibody. ST2 and CXCR3 expression versus i.v. CD45 expression in VAT Treg cells (left). Frequencies of intravascularly labelled (i.v.+) and non-labelled (i.v.-) DN, CXCR3+ and ST2+ VAT Treg cells (right). h, Flow cytometry plots show CD73 co-expression with CD62L, CXCR3 and ST2 in donor VAT Treg cells adoptively transferred to Rag1-/- mice. i, Quantification of VAT Treg cells in male Ar-/- and wildtype control mice (n = 8 control, n = 10 Ar-/-). Quantification of VAT Treg cells in female Era-/- and wildtype control mice (n = 4 control, n = 7 Era-/-). Symbols represent individual mice (a, c-f, i, j) or mean of a minimum of 5 mice (b). Error bars indicate standard deviation; horizontal lines indicate means. Statistical analyses performed using an unpaired, two-tailed Student’s t-test (b, j), two-way ANOVA (a, e, f) or a linear regression model (c, d). Data pooled from at least two independent experiments (a - g, i, j); flow cytometry (FACS) plots representative (a, f, h).
Extended Data Fig. 3 GATA3 controls ST2+ VAT Treg cell differentiation.
a, FACS plots show ST2 and GATA3 expression in VAT Treg cells of male and female mice. b, GATA3 expression in CD4+ Foxp3−and Foxp3+ cells of Gata3fl/flFoxp3Cre and control mice. c, Graphs show frequencies of total (left) and activated (right) splenic Treg cells of Gata3fl/flFoxp3Cre (left n = 4 females, n = 6 males; right n = 8 females, n = 13 males) and control (left n = 8 females, n = 11 males; right n = 8 females, n = 7 males) mice. d, Treg cell frequencies and numbers in small intestine lamina propria (n = 13 control, n = 6 Gata3fl/flFoxp3Cre) and lung (n = 10 per genotype). e, Numbers of ST2+ and CXCR3+ VAT Treg cells in Gata3fl/flFoxp3Cre (n = 6 each sex) and control (n = 8 females, n = 7 males) mice. f, Frequencies (control n = 12 each sex; Gata3fl/flFoxp3Cre n = 10 females, n = 11 males) and numbers (control n = 12 females, n = 11 males; Gata3fl/flFoxp3Cre n = 10 each sex) of VAT Treg cells. g, Numbers of TNF+ (control n = 9 females, n = 15 males; Gata3fl/flFoxp3Cre n = 4 females, n = 12 males), IL-5+ (control n = 8 females, n = 10 males; Gata3fl/flFoxp3Cre n = 5 females, n = 7 males) and IFN-γ+ (control n = 8 females, n = 10 male; Gata3fl/flFoxp3Cre n = 5 females, n = 7 males) cells. h, Body (control n = 13 females, n = 15 males; Gata3fl/flFoxp3Cre n = 11 females, n = 10 males) and VAT (control n = 8 females, n = 14 males; Gata3fl/flFoxp3Cre n = 10 females, n = 15 males) weights of male and female Gata3fl/flFoxp3Cre and control mice. i, Body composition of male and female Gata3fl/flFoxp3Cre and control mice (control n = 4 females, n = 5 male; Gata3fl/flFoxp3Cre n = 5 females, n = 4 males). Data representative (a, b) or pooled (c-i) from at least two independent experiments. Symbols represent individual mice, error bars indicate standard deviation, horizontal lines are means. Statistical analyses performed using two-way ANOVA or unpaired, two-tailed Student’s t-test (d).
Extended Data Fig. 4 GATA3 and T-bet control the differentiation of VAT Treg cells.
a, b, Volcano plots from bulk RNAseq analysis showing differential expression of genes in female (a) and male (b) Gata3fl/flFoxp3Cre and control mice. c, In silico foot printing analysis of open chromatin regions in ST2+ VAT Treg cells show binding sites for the transcription factors BATF, BACH2, GATA3 and PPARγ. d, ATAC-seq tracks from ST2+ and CXCR3+ VAT Treg cells showing differential chromatin accessibility in the Pparg locus. Accessible region in ST2+ Treg cells show putative GATA3 binding site. e, Bar graph showing scRNAseq UMAP cluster frequency of CD4 T cells from normal diet (ND) and high fat diet (HFD) fed male and female mice. f, FACS plots showing expression of CXCR3 and Foxp3 in splenic CD4+ T cells from Tbx21fl/flFoxp3Cre (n = 6 females, n = 7 males) and control mice (n = 9 females, n = 14 males). g, Frequencies of Treg cells (Foxp3+) among splenic CD4+TCRβ+ cells in Tbx21fl/flFoxp3Cre and control mice. h, Proportions (left) and numbers (right) of VAT Treg cells in Tbx21fl/flFoxp3Cre and control mice (control n = 6 females, n = 14 males; Gata3fl/flFoxp3Cre n = 9 females, n = 10 males). i, j, Volcano plots from bulk VAT Treg cell RNAseq analysis showing differential expression of genes in female (i) and male (j) Tbx21fl/flFoxp3Cre and control mice. Symbols represent individual mice; horizontal lines indicate means. Data are representative (f) or pooled (g, h) from at least two independent experiments. Error bars indicate the standard deviation. Statistical analyses were performed using two-way ANOVA.
Extended Data Fig. 5 Distinct cytokine profiles of female and male VAT.
a, FACS plots show TCRβ versus IL-5 or IL-13 expression in CD45+ VAT cells of female (top) and male (bottom) C57BL/6 mice. b, Il4AmCyan (n = 5 each sex) and Il5TdTom (n = 7 females, n = 5 males) expression in female (top) and male (bottom) VAT CD45+ VAT cells. Quantification on right. c, d, FACS plots show TCRβ versus IFN-γ or TNF expression in VAT CD45+ cells (c) and Foxp3 versus IFN-γ or TNF expression in VAT CD4+ cells of C57BL/6 female (d, top) and male (d, bottom) mice. Frequencies of IFN-γ+ (n = 8 females, n = 10 males) or TNF+ (n = 5 females, n = 9 males) among CD4+Foxp3−cells and amount (mean fluorescence index, MFI) of IFN-γ (n = 9 females, n = 10 males) or TNF (n = 8 females, n = 14 males). e, FACS plots show TCRβ versus Ifng expression in CD45+ VAT cells of female (top) and male (bottom) IfngeYFP mice. f, Frequencies of Ifng+ among CD45+ cells (left, n = 9 females, n = 11 males) and amount (MFI) in VAT TCRβ+ and TCRβ- cells (right, n = 5 females, n = 6 males) of IfngeYFP mice. g, Numbers of ST2+ (control n = 6 each sex; Ifng-/- n = 4 females, n = 9 males) and CXCR3+ (control n = 7 females, n = 6 males; Ifng-/- n = 7 females, n = 9 males) VAT Treg cells in female and male Ifng-/- mice. h, FACS plots show expression CD4 and Foxp3 in T cells from spleen (top) and VAT (bottom) in control (PBS) and single-dose diphtheria toxin (DT) treated Foxp3DTR mice. i, Numbers of TCRβ+ cells in control (PBS) and Foxp3DTR mice 10-days-post DT administration. j, Proportions of CD62L+, CXCR3+ and ST2+ VAT Treg cells in female control (PBS) or Foxp3DTR mice 10-days-post DT administration. Symbols represent individual mice, horizontal lines mean error bars standard deviation. Data pooled from at least two independent experiments (b, d, f, g, i, j); FACS plots representative (a - e, h). Unpaired, two-tailed Student’s t-test (b, d, f) or two-way ANOVA (g, i, j).
Extended Data Fig. 6 ST2+ and CXCR3+ VAT Treg cells have distinct functions.
a, Graphs show numbers of CD8 (control n = 22 females, n = 32 males; Ppargfl/flFoxp3Cre n = 17 each sex; Tbx21fl/flFoxp3Cre n = 11 each sex) and CD4 T cells (control n = 22 females, n = 32 males; Ppargfl/flFoxp3Cre n = 17 each sex; Tbx21fl/flFoxp3Cre n = 8 each sex) and TNF (control n = 21 females, n = 26 males; Ppargfl/flFoxp3Cre n = 9 females, n = 5 males; Tbx21fl/flFoxp3Cre n = 4 females, n = 10 males) or IFNγ (control n = 21 females, n = 28 males; Ppargfl/flFoxp3Cre n = 9 females, n = 5 males; Tbx21fl/flFoxp3Cre n = 8 females, n = 12 males) producing TCRβ+ cells in the VAT of male and female Tbx21fl/flFoxp3Cre, Ppargfl/flFoxp3Cre and control mice. b, VAT Treg cell quantification in male and female Tbx21fl/flFoxp3Cre, Ppargfl/flFoxp3Cre and control mice on HFD (control n = 10 females, n = 10 males; Tbx21fl/flFoxp3Cre n = 9 females, n = 8 males; Ppargfl/flFoxp3Cre n = 9 females, n = 14 males). c, d, Insulin tolerance was assessed in Ppargfl/flFoxp3Cre mice (c) and Tbx21fl/flFoxp3Cre mice (d) under conditions of normal diet (ND) and high fat-diet (HFD) feeding. ND on top and HFD bottom, females on left, males on right. Symbols represent individual mice (a, b) or mean of 3−5 mice (c, d). Data are pooled from 2−3 independent experiments. Error bars indicate the standard deviation (a, b) or s.e.m. (c, d). Statistical analyses were performed using two-way ANOVA (a, b) or unpaired, two-tailed Student’s t-test (c, d).
Extended Data Fig. 7 ST2+ and CXCR3+ Treg cells interact with distinct cell types.
a, FACS histogram overlays showing expression of GzmB, CTLA-4, Areg, CD25, IL-10 and CD39 in ST2+ and CXCR3+ VAT Treg cells. b, Graph showing frequency of IL-10-GFP expressing ST2+ VAT Treg cells from male (n = 16) and female (n = 9) mice. c, Dot plot shows reciprocal interaction between ST2+ and CXCR3+ VAT Treg cells and the VAT microenvironment. d, FACS gating scheme used to identify different myeloid cells in the VAT. e, Graph shows Ccl2-RFP mean fluorescence index (MFI) in VAT dendritic cells, macrophage subsets and neutrophils from male (n = 8) and female (n = 10) mice. f, FACS plots show frequency of Ccl2-RFP+ cells in male and female VAT resident and inflammatory macrophages. Quantification on the right. Symbols represent individual mice; horizontal lines indicate means. Data are pooled from at least two independent experiments (b, e, f); FACS plots are representative (a, d, f). Error bars indicate the standard deviation. Statistical analyses were performed using two-way ANOVA (e, f) or unpaired, two-tailed Student’s t-test (b).
Extended Data Fig. 8 ST2 and CXCR3 + VAT Treg cells regulate distinct inflammatory pathways.
a, b, Distribution of dendritic cell and macrophage populations in the VAT of male Tbx21fl/flFoxp3Cre (n = 5), Ppargfl/flFoxp3Cre (n = 8) and control mice under normal diet (ND, n = 4) and high fat diet (HFD, n = 5) conditions. (a) Macrophage frequencies (left) and numbers on right (b) Dendritic cell frequencies (left) and numbers on right. c, Representative flow cytometry plot showing expression of IL-5 by ST2+ Treg cells in male mice. d-f, (d) Representative FACS plots showing Cxcl9 and Cxcl10 reporter expression in male and female REX3 reporter mice. (e) Frequencies (left) and numbers (right) of Cxcl9+ cells in the VAT CD45+ compartment of male (n = 4) and female (n = 6) REX3 mice. (f) Frequencies (left) and numbers (right) of Cxcl10+ cells in the VAT CD45+ compartment of male (n = 4) and female (n = 6) REX3 mice. g, Gating strategy for macrophage and dendritic cells in the VAT. h, Graph showing frequency of Cxcl10 (left) and Cxcl9 (right) expressing macrophages and dendritic cells in male (n = 4) and female (n = 6) REX3 mice. Symbols represent individual mice; horizontal lines indicate means. Data are pooled from at least two independent experiments (a, b, e, f, h); FACS plots are representative (c, d, g). Error bars indicate the standard deviation. Statistical analyses were performed using two-way ANOVA (a, b, h) or unpaired, two-tailed Student’s t-test (e, f).
Extended Data Fig. 9 Altered VAT transcriptional profiles after Treg-cell-specific deletion of T-bet and PPARγ.
a-e, Volcano plots shows genes differentially expressed between total VAT of female vs male control mice (a), female vs male Ppargfl/flFoxp3Cre (b), female vs male Tbx21fl/flFoxp3Cre (c), female Tbx21fl/flFoxp3Cre vs control (d) and male Tbx21fl/flFoxp3Cre vs control (e) mice. f, KEGG (top) and GO (bottom) pathway enrichment analysis for differentially expressed genes in female VAT of Tbx21fl/flFoxp3Cre vs control mice. Positive values refer to pathways upregulated (red), negative values refer to pathways downregulated (blue) in the VAT. g, Flow cytometry plots showing glucose uptake measured by 2-NBDG staining in ST2+ and CXCR3+ male and female VAT Treg cells. Quantification on the right (n = 3 females, n = 6 males). h, i, Volcano plots showing genes differentially expressed between total VAT of female Ppargfl/flFoxp3Cre vs control (i) and male Ppargfl/flFoxp3Cre vs control (j) mice. Each dot in volcano plot represents a gene; genes highlighted in red are up-regulated and genes highlighted in blue are down-regulated.
Supplementary information
Supplementary Table 1
scRNAseq cluster marker genes.
Supplementary Table 2
ST2 and CXCR3 signatures.
Supplementary Table 3
GATA3 versus WT Treg RNAseq DE genes.
Supplementary Table 4
scRNAseq: ND versus HFD (all clusters).
Supplementary Table 5
T-bet versus WT Treg RNAseq DE genes.
Supplementary Table 6
Adipose tissue DE genes.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Torres, S.V., Man, K., Elmzzahi, T. et al. Two regulatory T cell populations in the visceral adipose tissue shape systemic metabolism. Nat Immunol 25, 496–511 (2024). https://doi.org/10.1038/s41590-024-01753-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-024-01753-9
This article is cited by
-
Adipose Treg cells in charge of metabolism
Nature Immunology (2024)
-
A Treg cell duo for VAT control
Nature Reviews Immunology (2024)
