
Abstract
Glioblastoma (GBM) tumors consist of multiple cell populations, including self-renewing glioblastoma stem cells (GSCs) and immunosuppressive microglia. Here we identified Kunitz-type protease inhibitor TFPI2 as a critical factor connecting these cell populations and their associated GBM hallmarks of stemness and immunosuppression. TFPI2 promotes GSC self-renewal and tumor growth via activation of the c-Jun N-terminal kinase–signal transducer and activator of transcription (STAT)3 pathway. Secreted TFPI2 interacts with its functional receptor CD51 on microglia to trigger the infiltration and immunosuppressive polarization of microglia through activation of STAT6 signaling. Inhibition of the TFPI2–CD51–STAT6 signaling axis activates T cells and synergizes with anti-PD1 therapy in GBM mouse models. In human GBM, TFPI2 correlates positively with stemness, microglia abundance, immunosuppression and poor prognosis. Our study identifies a function for TFPI2 and supports therapeutic targeting of TFPI2 as an effective strategy for GBM.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
Proteomic data generated by IP–MS have been deposited in the Mass spectrometry Interactive Virtual Environment under dataset accession number of MSV000090624 (https://massive.ucsd.edu/ProteoSAFe/dataset.jsp?accession = MSV000090624). RNA-seq data have been desposited in the Gene Expression Omnibus under the accession code is GSE232486 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE232486). The TCGA GBM datasets, TCGA GBMLGG merged datasets, Brain Tumor Immune Micro Environment dataset (https://joycelab.shinyapps.io/braintime/) and published GEO datasets EGAS00001004422 (https://ega-archive.org/datasets/EGAD00001006206) and GSE131928 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE131928) were also accessed. All other data are available within the article and supplementary information. Source data are provided with this paper.
References
Prager, B. C., Bhargava, S., Mahadev, V., Hubert, C. G. & Rich, J. N. Glioblastoma stem cells: driving resilience through chaos. Trends Cancer 6, 223–235 (2020).
Couturier, C. P. et al. Single-cell RNA-seq reveals that glioblastoma recapitulates a normal neurodevelopmental hierarchy. Nat. Commun. 11, 3406 (2020).
Guilhamon, P. et al. Single-cell chromatin accessibility profiling of glioblastoma identifies an invasive cancer stem cell population associated with lower survival. eLife 10, e64090 (2021).
Chen, P., Hsu, W. H., Han, J., Xia, Y. & DePinho, R. A. Cancer stemness meets immunity: from mechanism to therapy. Cell Rep. 34, 108597 (2021).
Xuan, W., Lesniak, M. S., James, C. D., Heimberger, A. B. & Chen, P. Context-dependent glioblastoma-macrophage/microglia symbiosis and associated mechanisms. Trends Immunol. 42, 280–292 (2021).
Khan, F. et al. Macrophages and microglia in glioblastoma: heterogeneity, plasticity, and therapy. J. Clin. Invest. 133, e163446 (2023).
Zhou, W. et al. Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat. Cell Biol. 17, 170–182 (2015).
Shi, Y. et al. Tumour-associated macrophages secrete pleiotrophin to promote PTPRZ1 signalling in glioblastoma stem cells for tumour growth. Nat. Commun. 8, 15080 (2017).
Xuan, W. et al. Circadian regulator CLOCK drives immunosuppression in glioblastoma. Cancer Immunol. Res. 10, 770–784 (2022).
Chen, P. et al. Circadian regulator CLOCK recruits immune-suppressive microglia into the GBM tumor microenvironment. Cancer Discov. 10, 371–381 (2020).
Sprecher, C. A., Kisiel, W., Mathewes, S. & Foster, D. C. Molecular cloning, expression, and partial characterization of a second human tissue-factor-pathway inhibitor. Proc. Natl Acad. Sci. USA 91, 3353–3357 (1994).
Herman, M. P. et al. Tissue factor pathway inhibitor-2 is a novel inhibitor of matrix metalloproteinases with implications for atherosclerosis. J. Clin. Investig. 107, 1117–1126 (2001).
Winkler, J., Abisoye-Ogunniyan, A., Metcalf, K. J. & Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 11, 5120 (2020).
Rollin, J. et al. Expression and methylation status of tissue factor pathway inhibitor-2 gene in non-small-cell lung cancer. Br. J. Cancer 92, 775–783 (2005).
Sato, N. et al. Epigenetic inactivation of TFPI-2 as a common mechanism associated with growth and invasion of pancreatic ductal adenocarcinoma. Oncogene 24, 850–858 (2005).
Wang, G. L. et al. TFPI-2 suppresses breast cancer cell proliferation and invasion through regulation of ERK signaling and interaction with actinin-4 and myosin-9. Sci. Rep. 8, 14402 (2018).
Neaud, V. et al. Paradoxical pro-invasive effect of the serine proteinase inhibitor tissue factor pathway inhibitor-2 on human hepatocellular carcinoma cells. J. Biol. Chem. 275, 35565–35569 (2000).
Ruf, W. et al. Differential role of tissue factor pathway inhibitors 1 and 2 in melanoma vasculogenic mimicry. Cancer Res. 63, 5381–5389 (2003).
Mo, J. et al. TFPI2 promotes perivascular migration in an angiotropism model of melanoma. Front. Oncol. 11, 662434 (2021).
Ota, Y. et al. Tissue factor pathway inhibitor‑2 is specifically expressed in ovarian clear cell carcinoma tissues in the nucleus, cytoplasm and extracellular matrix. Oncol. Rep. 45, 1023–1032 (2021).
George, J., Gondi, C. S., Dinh, D. H., Gujrati, M. & Rao, J. S. Restoration of tissue factor pathway inhibitor-2 in a human glioblastoma cell line triggers caspase-mediated pathway and apoptosis. Clin. Cancer Res. 13, 3507–3517 (2007).
Gessler, F., Voss, V., Seifert, V., Gerlach, R. & Kogel, D. Knockdown of TFPI-2 promotes migration and invasion of glioma cells. Neurosci. Lett. 497, 49–54 (2011).
Rao, C. N. et al. Expression of tissue factor pathway inhibitor 2 inversely correlates during the progression of human gliomas. Clin. Cancer Res. 7, 570–576 (2001).
Miranda, A. et al. Cancer stemness, intratumoral heterogeneity, and immune response across cancers. Proc. Natl Acad. Sci. USA 116, 9020–9029 (2019).
Yoshihara, K. et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 4, 2612 (2013).
Chai, R. et al. A novel gene signature based on five glioblastoma stem-like cell relevant genes predicts the survival of primary glioblastoma. J. Cancer Res. Clin. Oncol. 144, 439–447 (2018).
Uhlen, M. et al. The human secretome. Sci. Signal 12, eaaz0274 (2019).
Klemm, F. et al. Interrogation of the microenvironmental landscape in brain tumors reveals disease-specific alterations of immune cells. Cell 181, 1643–1660 e1617 (2020).
Neftel, C. et al. An integrative model of cellular states, plasticity, and genetics for glioblastoma. Cell 178, 835–849 e821 (2019).
Shingu, T. et al. Qki deficiency maintains sternness of glioma stem cells in suboptimal environment by downregulating endolysosomal degradation. Nat. Genet. 49, 75–86 (2017).
Saha, D., Martuza, R. L. & Rabkin, S. D. Macrophage polarization contributes to glioblastoma eradication by combination immunovirotherapy and immune checkpoint blockade. Cancer Cell 32, 253–267 e255 (2017).
Yoon, C. H. et al. c-Jun N-terminal kinase has a pivotal role in the maintenance of self-renewal and tumorigenicity in glioma stem-like cells. Oncogene 31, 4655–4666 (2012).
Gu, C. Y. et al. Tumor-specific activation of the C-JUN/MELK pathway regulates glioma stem cell growth in a p53-dependent manner. Stem Cells 31, 870–881 (2013).
Shi, Y. et al. Ibrutinib inactivates BMX-STAT3 in glioma stem cells to impair malignant growth and radioresistance. Sci. Transl. Med. 10, eaah6816 (2018).
Stechishin, O. D. et al. On-target JAK2/STAT3 inhibition slows disease progression in orthotopic xenografts of human glioblastoma brain tumor stem cells. Neuro Oncol. 15, 198–207 (2013).
Deng, Y. et al. Identifying mutual exclusivity across cancer genomes: computational approaches to discover genetic interaction and reveal tumor vulnerability. Brief. Bioinform. 20, 254–266 (2019).
Guo, L. et al. Postmortem adult human microglia proliferate in culture to high passage and maintain their response to amyloid-β. J. Alzheimers Dis. 54, 1157–1167 (2016).
Nagashima, S. et al.Synthesis and evaluation of 2-{[2-(4-hydroxyphenyl)-ethyl] amino} pyrimidine-5-carboxamide derivatives as novel STAT6 inhibitors. Bioorgan. Med. Chem 15, 1044–1055 (2007).
Uhlen, M. et al. Tissue-based map of the human proteome. Science 347, 1260419 (2015).
Chen, J. et al. CCL18 from tumor-associated macrophages promotes breast cancer metastasis via PITPNM3. Cancer Cell 19, 541–555 (2011).
Chen, C. H. et al. Novel murine glioblastoma models that reflect the immunotherapy resistance profile of human disease. Neuro Oncol. 27, noad025 (2023).
Bloch, O. et al. Gliomas promote immunosuppression through induction of B7-H1 expression in tumor-associated macrophages. Clin. Cancer Res. 19, 3165–3175 (2013).
Gimple, R. C., Yang, K. L., Halbert, M. E., Agnihotri, S. & Rich, J. N. Brain cancer stem cells: resilience through adaptive plasticity and hierarchical heterogeneity. Nat. Rev. Cancer 22, 497–514 (2022).
Wang, X. et al. Sequential fate-switches in stem-like cells drive the tumorigenic trajectory from human neural stem cells to malignant glioma. Cell Res 31, 684–702 (2021).
Bachoo, R. M. et al. Epidermal growth factor receptor and Ink4a/Arf: convergent mechanisms governing terminal differentiation and transformation along the neural stem cell to astrocyte axis. Cancer Cell 1, 269–277 (2002).
Dong, Z. et al. Targeting glioblastoma stem cells through disruption of the circadian clock. Cancer Discov. 9, 1556–1573 (2019).
Dirkse, A. et al. Stem cell-associated heterogeneity in glioblastoma results from intrinsic tumor plasticity shaped by the microenvironment. Nat. Commun. 10, 1787 (2019).
Schaettler, M. O. et al. Characterization of the genomic and immunologic diversity of malignant brain tumors through multisector analysis. Cancer Discov. 12, 154–171 (2021).
Konduri, S. D. et al. A novel function of tissue factor pathway inhibitor-2 (TFPI-2) in human glioma invasion. Oncogene 20, 6938–6945 (2001).
Jacob, F. et al. A patient-derived glioblastoma organoid model and biobank recapitulates inter- and intra-tumoral heterogeneity. Cell 180, 188–204.e122 (2020).
LeBlanc, V. G. et al. Single-cell landscapes of primary glioblastomas and matched explants and cell lines show variable retention of inter- and intratumor heterogeneity. Cancer Cell 40, 379–392.e379 (2022).
Bayik, D. & Lathia, J. D. Cancer stem cell–immune cell crosstalk in tumour progression. Nat. Rev. Cancer 21, 526–536 (2021).
Pang, L., Khan, F., Dunterman, M. & Chen, P. Pharmacological targeting of the tumor–immune symbiosis in glioblastoma. Trends Pharmacol. Sci. 43, 686–700 (2022).
Brennan, C. W. et al. The somatic genomic landscape of glioblastoma. Cell 155, 462–477 (2013).
Parsa, A. T. et al. Loss of tumor suppressor PTEN function increases B7-H1 expression and immunoresistance in glioma. Nat. Med. 13, 84–88 (2007).
Chen, P. et al. Symbiotic macrophage–glioma cell interactions reveal synthetic lethality in PTEN-null glioma. Cancer Cell 35, 868–884 e866 (2019).
Liu, H., Wang, G., Hao, D., Wang, C. & Zhang, M. Antimicrobial and immunoregulatory activities of TS40, a derived peptide of a TFPI-2 homologue from black rockfish (Sebastes schlegelii). Mar. Drugs 20, 353 (2022).
Cai, W. et al. STAT6/Arg1 promotes microglia/macrophage efferocytosis and inflammation resolution in stroke mice. JCI Insight 4, e131355 (2019).
Wang, N., Liang, H. & Zen, K. Molecular mechanisms that influence the macrophage m1–m2 polarization balance. Front. Immunol. 5, 614 (2014).
Pang, L., Khan, F., Heimberger, A. B. & Chen, P. Mechanism and therapeutic potential of tumor–immune symbiosis in glioblastoma. Trends Cancer 8, 839–854 (2022).
Zhang, B. et al. Macrophage-expressed CD51 promotes cancer stem cell properties via the TGF-β1/smad2/3 axis in pancreatic cancer. Cancer Lett. 459, 204–215 (2019).
Chen, P. & Dey, P. Co-dependencies in the tumor immune microenvironment. Oncogene 41, 3821–3829 (2022).
Chamberlain, M. C. & Kim, B. T. Nivolumab for patients with recurrent glioblastoma progressing on bevacizumab: a retrospective case series. J. Neurooncol. 133, 561–569 (2017).
Reiss, S., Yerram, P., Modelevsky, L. & Grommes, C. Retrospective review of safety and efficacy of checkpoint inhibition in refractory high-grade gliomas. J. Clin. Oncol. 35, no. 15_suppl. 2033 (2017).
Reardona, D. A. et al. Randomized phase 3 study evaluating the efficacy and safety of nivolumab vs bevacizumab in patients with recurrent glioblastoma: Checkmate 143. Neuro Oncol. 19, 21 (2017).
Zhao, J. et al. Immune and genomic correlates of response to anti-PD-1 immunotherapy in glioblastoma. Nat. Med. 25, 462–469 (2019).
Arrieta, V. A. et al. ERK1/2 phosphorylation predicts survival following anti-PD-1 immunotherapy in recurrent glioblastoma. Nat. Cancer 2, 1372–1386 (2021).
Seyfried, T. N., El-Abbadi, M., Ecsedy, J. A., Bai, H. W. & Yohe, H. C. Influence of host cell infiltration on the glycolipid content of mouse brain tumors. J. Neurochem. 66, 2026–2033 (1996).
Chen, P. et al. Collagen VI regulates peripheral nerve regeneration by modulating macrophage recruitment and polarization. Acta Neuropathol. 129, 97–113 (2015).
Ershov, D. et al. TrackMate 7: integrating state-of-the-art segmentation algorithms into tracking pipelines. Nat. Methods 19, 829–832 (2022).
Tinevez, J. Y. et al. TrackMate: an open and extensible platform for single-particle tracking. Methods 115, 80–90 (2017).
Chen, P. et al. Gpr132 sensing of lactate mediates tumor-macrophage interplay to promote breast cancer metastasis. Proc. Natl Acad. Sci. USA 114, 580–585 (2017).
Varghese, F., Bukhari, A. B., Malhotra, R. & De, A. IHC Profiler: an open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 9, e96801 (2014).
Barth, N. D. et al. A fluorogenic cyclic peptide for imaging and quantification of drug-induced apoptosis. Nat. Commun. 11, 4027 (2020).
Ong Derrick Sek, T. et al. PAF promotes stemness and radioresistance of glioma stem cells. Proc. Natl Acad. Sci. USA 114, E9086–E9095 (2017).
Galaxy, C. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2022 update. Nucleic Acids Res. 50, W345–W351 (2022).
Couturier, C. P. et al. Single-cell RNA-seq reveals that glioblastoma recapitulates a normal neurodevelopmental hierarchy. Nat. Commun. 11, 3406 (2020).
Acknowledgements
We thank S. D. Rabkin, F. Lang, J. Hu and H. Okada for providing 005 GSCs, patient-derived GSCs, QPP7 GSC and SB28, respectively. This work was supported in part by NIH R00 CA240896 (P.C.), NIH R01 NS124594 (P.C.), DoD Career Development Award W81XWH-21-1-0380 (P.C.), Cancer Research Foundation Young Investigator Award (P.C.), Lynn Sage Scholar Award (P.C.), American Cancer Society Institutional Research Grant IRG-21-144-27 (P.C.), NIH P50 CA221747 (to P.C., Brain Cancer SPORE CEP Award), philanthropic donation from Mindy Jacobson and the Bill Bass Foundation (P.C.), Northwestern University start-up funds (P.C.), the Northwestern Medicine Malnati Brain Tumor Insitute of the Robert H. Lurie Comprehensive Cancer Center (P.C.), NIH R24 NS104160 (C.G.) and NIH P30 AG072977 (C.G.). W.H.H. is funded by the CPRIT Training Program (RP210028). Imaging work was performed at the Northwestern University Center for Advanced Microscopy generously supported by NCI CCSG P30 CA060553. Proteomics experiments were performed at the Northwestern Proteomics Core Facility, which is generously supported by NCI CCSG P30 CA060553, instrumentation award (S10OD025194) from NIH Office of Director, and P41 GM108569. Incucyte analysis was performed in the Analytical bioNanoTechnology Core (ANTEC) Facility supported by the Soft and Hybrid Nanotechnology Experimental (SHyNE) Resource (NSF ECCS-2025633).
Author information
Authors and Affiliations
Contributions
L.P. performed most of the experiments. M.D. performed some in vitro and in vivo studies. S.G. provided help for IHC staining. F.K. performed the kinase array profiling and provided assistance for tumor implantation. Y.L. provided help for migration assay and IP. E.T., A.B. and C.G. isolated and cultured human primary microglia. W.-H.H. provided help for scRNA-seq data analysis. C.H. and C.D.J. provided help with human patient samples and commented the study. P.C. designed the project and analyzed data. P.C., L.P. and M.D. wrote the paper. All authors participated in editing the paper.
Corresponding author
Ethics declarations
Competing interests
L.P. and P.C. are listed as inventors on a patent related to targeting the TFPI2–CD51–STAT6 signaling axis combining with or without anti-PD1 therapy. All other authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks the anonymous reviewers for their contribution to the peer review of this work. N. Bernard was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 TFPI2 promotes GSC self-renewal.
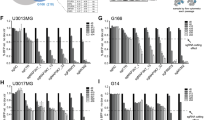
a, Correlation between TFPI2 or LOXL2 expression and IDH1-WT GBM survival in TCGA datasets. P values are shown. b, c, Correlation between stemness (b) or GSC signature (c) and TFPI2 or LOXL2 expression in TCGA GBM dataset. R and P values were determined by Pearson correlation. d, e, Expression of TFPI2 (d) and LOXL2 (e) in CD45− cells and immune cells of non-tumor tissues and GBM tumors. n = 5 and 17 for CD45− cells, 8 and 15 for microglia (MG), 7 and 13 for macrophages (MDM), 6 and 13 for neutrophils, 6 and 11 for CD4+ T cells, as well as 7 and 6 for CD8+ T cells of non-tumor tissues and GBM, respectively. f, TFPI2 copy number correlates with TFPI2 in HG-U133A TCGA GBM tumors. n = 6, 88, 405 and 9 for Hetloss, Diploid, Gain and Amp, respectively. g, TFPI2 expression in glioma tumors at different stages. n = 256, 244, and 150 for stage II, III and IV, respectively. h, TFPI2 expression in different types of brain tumors. n = 191, 130, 194 and 152 for oligodendroglioma (OG), oligoastrocytoma (OA), astrocytoma and GBM, respectively. i, TFPI2 expression in different subtypes of GBM tumors. n = 182, 156 and 151 for classical, mesenchymal and proneural, respectively. j, k, High-resolution UMAP dimensional reduction of all cells (j) and malignant cells (k) from GBM patient tumors. l, m, UMAP dimensional reduction of the basis of the expression pattern of hallmark stemness signature (l) and TFPI2 (m) in malignant cells. n, Quantification of hallmark stemness signature in TFPI2-low and TFPI2-high tumors. o, Immunoblots for CD133 and SOX2 in cell lysates from GSC20 treated with TFPI2 protein at indicated concentrations. p–r, Representative (p) and quantification (q,r) of flow cytometry analysis of apoptosis in GSC262 and GSC23. n = 3 biological replicates. Error bars indicate mean ± SD (f–i) or SEM (q, r). One-way ANOVA test (f–i, q, r) and two-tailed Student’s t-test (n).
Extended Data Fig. 2 TFPI2 regulates GSC self-renewal, proliferation, and apoptosis.
a, b, Representative of tumorspheres of QPP7 (a) and CT2A (b) cells expressing control shRNA (shC) and Tfpi2 shRNAs (shTfpi2). Scale bar, 200 µm. c, d, Representative of proliferation in shC and shTfpi2 QPP7 (c) and CT2A (d) cells. e–j, Representative and quantification of flow cytometry for apoptosis in GSC272 (e, f), QPP7 GSCs (g, h) and CT2A cells (i, j) expressing shC and shTFPI2. k, Immunoblots for cleaved caspase 3 (CC3) in cell lysates from GSC272, QPP7, and CT2A cells expressing shC and shTFPI2. l, Immunoblots for CD133 and SOX2 in cell lysates from QPP7 GSCs expressing shTfpi2 with or without reexpression of shRNA-resistant TFPI2 cDNA (OE). m, Representative of tumorspheres in QPP7 GSCs expressing shTfpi2 with or without TFPI2 OE. Scale bar, 400 µm. n, Immunoblots for TFPI2 in cell lysates from TFPI2 wild-type (WT) and different clones of CRISPR knockout (KO) GSC272. o, Immunoblots for CD133 and SOX2 in cell lysates from TFPI2-WT and TFPI2-GSC272. p, Representative of tumorspheres in TFPI2-WT and TFPI2-KO GSC272. Scale bar, 400 µm. q, r, Representative (q) and quantification (r) of immunofluorescence staining of SOX2 in size-matched tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells. Scale bar, 50 µm. s, t, Representative (s) and quantification (t) of immunochemistry staining of CD133 in tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells. Scale bar, 200 µm. u–x, Representative and quantification of immunofluorescence staining of Ki67 (u,v) and CC3 (w,x) in tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells. Scale bar, 50 µm. In (f, h, j, r, t, v, x), n = 3 biological replicates. Error bars indicate mean ± SEM. One-way ANOVA test.
Extended Data Fig. 3 TFPI2 regulates GSC proliferation and apoptosis via JNK and STAT3 pathways.
a, b, Representative (a) and quantification (b) of proliferation in GSC23 expressing control and TFPI2 overexpression (OE) plasmid treated with or without JNK inhibitor JNK-IN-8 (10 nM). n = 3 biological replicates. c, d, Representative (c) and quantification (d) of proliferation in GSC262 expressing control and TFPI2 OE plasmid treated with or without JNK-IN-8 (10 nM). n = 3 biological replicates. e, f, Representative (e) and quantification (f) of apoptosis in GSC23 expressing control and TFPI2 OE plasmid treated with or without JNK-IN-8 (10 nM). n = 3 biological replicates. g, h, Representative (g) and quantification (h) of apoptosis in GSC262 expressing control and TFPI2 OE plasmid treated with or without JNK-IN-8 (10 nM). n = 3 biological replicates. i, j, Representative (i) and quantification (j) of proliferation in GSC23 expressing control and TFPI2 OE plasmid treated with or without STAT3 inhibitor WP1066 (20 nM). n = 3 biological replicates. k, l, Representative (k) and quantification (l) of proliferation in GSC262 expressing control and TFPI2 OE plasmid treated with or without WP1066 (20 nM). n = 3 biological replicates. m, n, Representative (m) and quantification (n) of apoptosis in GSC23 expressing control and TFPI2 OE plasmid treated with or without WP1066 (20 nM). n = 3 biological replicates. o, p, Representative (o) and quantification (p) of apoptosis in GSC262 expressing control and TFPI2 OE plasmid treated with or without WP1066 (20 nM). n = 3 biological replicates. Error bars indicate mean ± SEM. One-way ANOVA test.
Extended Data Fig. 4 TFPI2 promotes microglia migration via activating STAT6 signaling.
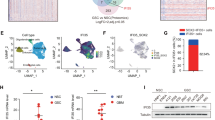
a, Amplification pattern of TFPI2 and CLOCK in GBM and low-grade glioma (LGG) patient tumors. b, GSEA analysis shows the top 10 enriched hallmark pathways in TFPI2-high GBM patient tumors compared to TFPI2-low tumors. Blue highlighted pathways relate to immune response. c, Representative of transwell migration of PrhMG following stimulation with conditioned media (CM) from GSC262 or GSC23 expressing control and TFPI2 overexpression (OE) plasmid. Scale bar, 400 µm. d, Representative of transwell migration of PrhMG, HMC3 and SIM-A9 microglia following stimulation with CM from GSC272 (for PrhMG and HMC3) or QPP7 cells (for SIM-A9) expressing control shRNA (shC) and TFPI2 shRNAs (shTFPI2). Scale bars, 400 µm for PrhMG; 200 µm for HMC3 and SIM-A9. e, f, Representative (e) and quantification (f) of flow cytometry for the percentage of CD45lowCD11b+ and CD45highCD11b+ cells in size matched tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells. n = 4 and 3 biological replicates for shC and shTfpi2, respectively. g, h, Representative (g) and quantification (h) of flow cytometry for the percentage of CD45highCD11b+CD68+ macrophages in size matched tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells. n = 4 and 3 biological replicates for shC and shTfpi2, respectively. i, Correlation between microglia signature and STAT6, AKT1, AKT2 or AKT3 in TCGA GBM dataset. R and P values were determined by Pearson correlation. j, Representative of transwell migration of mouse primary microglia isolated from wild-type (WT) and STAT6 knockout (KO) mice treated with or without TFPI2 recombinant protein (20 ng/ml). Scale bar, 200 µm. k, Representative of transwell migration of PrhMG, HMC3 and SIM-A9 microglia following stimulation with TFPI2 protein (20 ng/ml) in the presence or absence of STAT6 inhibitor AS1517499 at indicated concentrations. Scale bars, 400 µm for PrhMG; 200 µm for HMC3 and SIM-A9. Error bars indicate mean ± SEM. One-way ANOVA test.
Extended Data Fig. 5 TFPI2-STAT6 axis regulates microglia polarization.
a, Representative of flow cytometry for CD206 in PrhMG treated with conditioned media (CM) from GSC23 expressing control and TFPI2 overexpression (OE) plasmid. b, Representative of flow cytometry for CD206 in HMC3 microglia treated with CM from GSC272 expressing shRNA control (shC) and TFPI2 shRNAs (shTFPI2). c, Representative of flow cytometry for CD206 in SIM-A9 microglia treated with CM from QPP7 GSCs expressing shC and shTfpi2 with or without reexpression of shRNA-resistant TFPI2 cDNA (OE). d, e, Representative (d) and quantification (e) of flow cytometry for the percentage of CD45hiCD11b+CD68+CD206+ macrophages in size matched tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells. n = 4 and 3 for shC and shTfpi2, respectively. f–i, Immunofluorescence and quantification of CX3CR1+CD206+ (f,g) and CX3CR1+CD163+ (h,i) cells in tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells. Scale bar, 25 μm. n = 3 (g) or 6 (i) biological replicates. j–m, Immunofluorescence and quantification of CX3CR1+CD206+ (j,k) and CX3CR1+CD163+ (l,m) cells in tumors from nude mice intracranially implanted with shC and shTFPI2 GSC272. Scale bar, 25 μm. n = 6 biological replicates. n, Representative of CD206 positive microglia isolated from wild-type (WT) and STAT6 knockout (KO) mice and treated with or without TFPI2 recombinant protein (20 ng/ml). o, p, Representative of flow cytometry for CD206 (o) and ARG1 (p) in SIM-A9 microglia treated with or without TFPI2 protein (20 ng/ml) and STAT6 inhibitor (STAT6i) AS1517499 (25 nM). q, r, Representative of flow cytometry for CD206 (q) and ARG1 (r) in HMC3 microglia treated with or without TFPI2 protein (20 ng/ml) and AS1517499 (25 nM). s, Relative mRNA expression of IL6, TNFA, LFNG, and HLA-DR in HMC3 microglia treated with or without TFPI2 protein (20 ng/ml) and AS1517499 (25 nM). n = 3 biological replicates. Error bars indicate mean ± SEM. A one-way ANOVA followed by a Tukey test was performed to compare more than two groups.
Extended Data Fig. 6 CD51 is a receptor for TFPI2 activities on microglia.
a, CD51 expression level in human GBM samples from the TIME dataset. n = 17, 15, 13, 13, 11, and 6 for CD45− cells, microglia (MG), macrophages (MDM), neutrophils, CD4+ T cells and CD8+ T cells, respectively. b, CD51 expression in different cell populations in mouse CT2A tumor tissues. c, d, Immunoprecipitation with TFPI2 antibody (TFPI2 Ab) and immunoblotting for the interaction between TFPI2 and CD51 in SIM-A9 (c) and HMC3 (d) microglia. e, f, Calcium mobilization triggered by TFPI2 protein in HMC3 microglia (e) and PrhMG (f) treated with or without CD51 inhibitor MK-0429 at indicated concentrations. g, Immunoblots for CD51 in cell lysates of SIM-A9 microglia expressing shRNA control (shC) and Itgav shRNAs (shItgav). h, Immunoblots for P-PKCζ and PKCζ in cell lysates of shC and shItgav SIM-A9 microglia treated with or without TFPI2 protein. i, Immunoblots for P-STAT6 and STAT6 in cell lysates from SIM-A9 microglia and PrhMG treated with TFPI2 protein and MK-0429. j, Representative of transwell migration of PrhMG human HMC3, and mouse SIM-A9 microglia treated with or without TFPI2 protein and MK-0429 at indicated concentrations. Scale bars, 200 µm for HMC3 and SIM-A9, and 400 µm for PrhMG. k–n, Representative and quantification of flow cytometry for the percentage of CD206+ (k,l) and ARG1+ (m,n) SIM-A9 microglia treated with or without TFPI2 protein and MK-0429. n = 3 biological replicates. o–r, Representative (o) and quantification (p) of flow cytometry for the percentage of CD206+ (o,p) and ARG1+ (q,r) HMC3 microglia treated with or without TFPI2 recombinant protein and MK-0429 (50 nM). n = 3 biological replicates. s, t, Representative of flow cytometry for the percentage of CD206+ (s) and ARG1+ (t) PrhMG treated with or without TFPI2 protein and MK-0429. TFPI2 protein at 20 ng/ml and MK-0429 at 50 nM were used unless indicated. Error bars indicate mean ± SEM. One-way ANOVA test.
Extended Data Fig. 7 TFPI2-STAT6 signaling mediates microglia-induced T cell suppression.
a, b, Survival curves of nude mice implanted with QPP7 cells (2x104 cells; a) or CT2A cells (2x104 cells; b) expressing control shRNA (shC) and Tfpi2 shRNAs (shTfpi2). n = 8 biological replicates. c, d, Immunofluorescence (c) and quantification (d) of CD8 in size matched tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 CT2A cells expressing. Scale bar, 50 μm. n = 3 biological replicates. e, f, Immunofluorescence (e) and quantification (f) of CD4 in size matched tumors from C57BL/6 mice intracranially implanted with shC and shTfpi2 QPP7 GSCs. Scale bar, 50 μm. n = 3 biological replicates. g, Representative of flow cytometry analysis for CD45+CD3+CD8+PD1high and CD45+CD3+CD4+PD1high T cells in the spleens of C57BL/6 mice bearing size matched shC and shTfpi2 CT2A tumors. h, Representative of flow cytometry for IFNγ+CD8+ T cells cocultured with SIM-A9 mouse microglia treated with or without TFPI2 recombinant protein (20 ng/ml) and STAT6 inhibitor (STAT6i) AS1517499 (25 nM). i, Representative of flow cytometry for fixable viability dye-labeled JURKAT T cells cocultured with PrhMG treated with or without TFPI2 protein (20 ng/ml) and AS1517499 (25 nM). Percentage of live cells and dead cells are indicated. j, k, Representative (j) and quantification (k) of flow cytometry for the percentage of HLA-DR+ cells out of total JURKAT T cells cocultured with PrhMG treated with TFPI2 protein (20 ng/ml) in the presence or absence of AS1517499 (25 nM). n = 3 biological replicates. l, Representative of flow cytometry for CD69+ JURKAT T cells cocultured with PrhMG treated with or without TFPI2 protein (20 ng/ml) and AS1517499 (25 nM). m, Representative of flow cytometry analysis for IFNγ+ JURKAT T cells cocultured with PrhMG treated with TFPI2 protein (20 ng/ml) in the presence or absence of AS1517499 (25 nM). Error bars indicate mean ± SEM. One-way ANOVA test. In (a,b), log-rank test.
Extended Data Fig. 8 TFPI2-CD51-STAT6 signaling mediates microglia-induced T cell suppression.
a, b, CT2A cancer cell cytotoxicity induced by activated mouse primary CD8+ T cells cocultured with SIM-A9 microglia treated with TFPI2 recombinant protein (20 ng/ml) in the presence or absence of STAT6 inhibitor (STAT6i) AS1517499 (25 nM). The ratios between CD8+ T cells and CT2A cells are indicated. n = 10 biological replicates. c, GSC272 cytotoxicity induced by activated JURKAT T cells cocultured with PrhMG treated with TFPI2 protein (20 ng/ml) in the presence or absence of AS1517499 (25 nM). The ratios between CD8+ T cells and CT2A cells are indicated. n = 10 biological replicates. d, Correlation between CD274 (PD-L1) and TFPI2, STAT6, or ITGAV (CD51) in TCGA GBM dataset. e, Correlation between PDCD1LG2 (PD-L2) and TFPI2, STAT6, or ITGAV (CD51) in TCGA GBM dataset. In (d, e), R and P values were determined by Pearson correlation. Error bars indicate mean ± SEM. One-way ANOVA test.
Extended Data Fig. 9 TFPI2 correlates with proliferation and apoptosis markers in GBM patient tumors.
a, Representative images show low, medium, and high expression of TFPI2, Ki67, and cleaved caspase 3 (CC3) in human GBM tumor samples based on immunohistochemistry staining. Scale bar, 100 µm. b,c, Quantification of immunohistochemistry staining showing strong positive correlation between TFPI2 and Ki67 (b) and negative correlation between TFPI2 and CC3 (c) in human GBM tumor samples (n = 24). R and P values were determined by Pearson correlation. d-f, The plasma TFPI2 level in newly diagnosed (n = 38) and recurrent (n = 17) (d), female (n = 26) and male (n = 29) (e), as well as old (>65) (n = 29) and young (<65) (n = 26) GBM patients (f). Error bars indicate mean ± SEM. Student’s t-test.
Extended Data Fig. 10 Working model.
a, Illustration shows a proposed mechanism for the TFPI2 and its associated signaling axis in regulating self-renewal of glioma stem cells (GSCs) and GSC-microglia symbiosis in GBM. TFPI2 is amplified in GBM and essential for tumor progression. Upon secreting from GSCs, TFPI2 promotes GSC self-renewal by activating the JNK-STAT3 signaling pathway. Secreted TFPI2 binds to CD51 on microglia to promote microglia infiltration and immunosuppressive polarization via activation of STAT6 signaling. Consequently, these activated microglia inhibit the infiltration and activation of T cells and damage their tumor killing potential. b, Inhibition of TFPI2-CD51-STAT6 signaling axis impairs tumor progression by reducing immunosuppressive microglia and activating T cell-mediated antitumor immunity and synergizes with anti-PD1 therapy in GBM. This image was created with BioRender.com.
Supplementary information
Supplementary Information
Supplementary Figs. 1–13, uncropped blots of supplementary figures, and Supplementary Tables 1–3.
Supplementary Video 1
Supplementary Videos 1 and 2.
Supplementary Data 1
Source data for supplementary figures.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Statistical source data.
Source Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 1 (unprocessed western blots)
Unprocessed western blots.
Source Data Extended Data Fig. 2 (unprocessed western blots)
Unprocessed western blots.
Source Data Extended Data Fig. 6 (unprocessed western blots)
Unprocessed western blots.
Source Data Fig. 1 (unprocessed western blots)
Unprocessed western blots.
Source Data Fig. 2 (unprocessed western blots)
Unprocessed western blots.
Source Data Fig. 3 (unprocessed western blots)
Unprocessed western blots.
Source Data Fig. 4 (unprocessed western blots)
Unprocessed western blots.
Source Data Fig. 6 (unprocessed western blots)
Unprocessed western blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Pang, L., Dunterman, M., Guo, S. et al. Kunitz-type protease inhibitor TFPI2 remodels stemness and immunosuppressive tumor microenvironment in glioblastoma. Nat Immunol 24, 1654–1670 (2023). https://doi.org/10.1038/s41590-023-01605-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-023-01605-y
This article is cited by
-
Lactate dehydrogenase A regulates tumor-macrophage symbiosis to promote glioblastoma progression
Nature Communications (2024)
