
Abstract
In this study, the impact of the apolipoprotein B mRNA-editing catalytic subunit-like (APOBEC) enzyme APOBEC3B (A3B) on epidermal growth factor receptor (EGFR)-driven lung cancer was assessed. A3B expression in EGFR mutant (EGFRmut) non-small-cell lung cancer (NSCLC) mouse models constrained tumorigenesis, while A3B expression in tumors treated with EGFR-targeted cancer therapy was associated with treatment resistance. Analyses of human NSCLC models treated with EGFR-targeted therapy showed upregulation of A3B and revealed therapy-induced activation of nuclear factor kappa B (NF-κB) as an inducer of A3B expression. Significantly reduced viability was observed with A3B deficiency, and A3B was required for the enrichment of APOBEC mutation signatures, in targeted therapy-treated human NSCLC preclinical models. Upregulation of A3B was confirmed in patients with NSCLC treated with EGFR-targeted therapy. This study uncovers the multifaceted roles of A3B in NSCLC and identifies A3B as a potential target for more durable responses to targeted cancer therapy.
Similar content being viewed by others

Main
Apolipoprotein B mRNA-editing catalytic subunit-like (APOBEC) enzymes are cytosine deaminases that have an important role in intrinsic responses to viral infection through deamination of deoxycytidine residues in viral single-stranded DNA1,2. APOBEC3 (A3) enzymes can act as potent host genome mutagens in multiple cancer types including non-small-cell lung cancer (NSCLC)3,4. In patients, both APOBEC3A (A3A)5 and A3B6 have been implicated to have a major role in NSCLC3. Earlier tumor genome sequencing studies revealed subclonal enrichment for mutations in an APOBEC substrate context, suggesting a possible role for this enzyme family in the acquisition of mutations later in tumor evolution7,8,9,10. Analysis of APOBEC3 family gene expression across multiple stages of lung adenocarcinoma revealed significantly elevated expression of A3B at multiple timepoints (adenocarcinoma in situ and invasive lung adenocarcinoma) compared to normal tissue4.
While mouse models have contributed to our understanding of cancer evolution and drug responses11,12,13,14, they lack the mutational heterogeneity observed in human tumors15,16,17. This may be due in part to the fact that mice encode only a single, cytoplasmic and nongenotoxic APOBEC3 enzyme18,19. To understand the role of A3B in tumor evolution and therapy resistance, several mouse strains incorporating a human A3B transgene were engineered to mimic clonal and subclonal induction of A3B in oncogene-driven NSCLC and human preclinical models and clinical specimens were studied.
Results
A3B restrains tumor initiation in an epidermal growth factor receptor mutant (EGFRmut) lung cancer mouse model
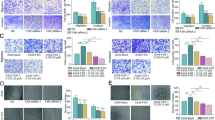
The role of A3B in tumor initiation was first investigated in a mouse strain combining a new loxP-STOP-loxP (LSL) inducible human A3B transgenic model (Rosa26LSL-A3Bi)20 with a Cre-inducible EGFRL858R-driven lung cancer mouse model (TetO-EGFRL858R; Rosa26LNL-tTA) to generate EA3B (TetO-EGFRL858R; Rosa26LNL-tTA/LSL-A3Bi)11,12,21 mice (Fig. 1a). The tumor number and total tumor volume per mouse at 3 months postinduction, and the fraction of mice with tumors was significantly lower in EA3B mice than in E (TetO-EGFRL858R; Rosa26LNL-tTA) control mice (Fig. 1b,c and Extended Data Fig. 1a). A significantly decreased number of EGFRL858R+ cells per lung area was also observed in EA3B mice versus E control mice (Fig. 1d). The programmed cell death marker caspase-3 was significantly higher in tumor cells of EA3B mice compared with E mice (Fig. 1e,f).

a, Tumorigenesis in E (TetO-EGFRL858R; Rosa26LNL-tTA) and EA3B (TetO-EGFRL858R; Rosa26LNL-tTA/LSL-A3Bi) mice was induced using the indicated viral titer. Tumor growth was assessed by micro-CT analysis. b, Total tumor volume per mouse at 3 months postinduction quantified by micro-CT analysis (E, n = 15; EA3B, n = 24; mean ± s.d., two-sided Mann–Whitney test, *P = 0.0163, each dot represents a mouse). c, Total tumor number per mouse at 3 months postinduction quantified by micro-CT analysis (E, n = 15; EA3B, n = 24, mean ± s.d., two-sided Mann–Whitney test, *P = 0.0236, each dot represents a mouse). d, Quantification of EGFRL858R+ cells per lung area (mm2) by IHC staining at 3 months postinduction (E, n = 9; EA3B, n = 10; mean ± s.d., two-sided Mann–Whitney test, *P = 0.0435, each dot represents a mouse). e, Quantification of caspase 3+ cells per mm2 of tumor at 3 months postinduction (E, n = 9; EA3B, n = 10; mean ± s.d., two-sided Mann–Whitney test, ****P < 0.0001, each dot represents a tumor). f, Representative IHC stainings of EGFRL858R, APOBEC3B and caspase-3 (scale bar = 20 µm, arrow indicates positive cell; E, n = 9; EA3B, n = 10 biological replicates). g, Percent chromosome missegregation errors at 3 months postinduction (two-sided Fisher’s exact test, *P = 0.016; E, n = 9; EA3B, n = 10). h, Tumorigenesis in E and E(CAG)A3BE255A mice was induced using the indicated viral titer (2.5 × 107 viral particles per mouse). i, Quantification of EGFRL858R+ cells per lung area (mm2) by IHC staining at 3 months postinduction (E, n = 12; E(CAG)A3BE255A, n = 12; mean ± s.d., each dot represents a mouse). j, Representative IHC staining of EGFRL858R and APOBEC3B (scale bar = 20 µm; E, n = 12; E(CAG)A3BE255A, n = 12). k, Tumor growth was assessed by micro-CT analysis in EP and EPA3B mice. l, Total tumor number per mouse at 3 months postinduction quantified by micro-CT analysis (EP, n = 21; EPA3B, n = 30; combined from two separate experiments). m, Survival curve of EP versus EPA3B mice (EP, n = 8; EPA3B, n = 7; each dot represents a mouse). NS, not significant.
We hypothesized that A3B expression at tumor initiation in EA3B mouse models might induce increased chromosomal instability (CIN), p53 pathway activation and tumor cell death based on previous work4. In our models, a significantly higher fraction of lagging chromosomes and chromatin bridges were observed in anaphase tumor cells of EA3B mice compared with E mice4 (Fig. 1g). There was also a significant increase in p53 nuclear positivity in tumors of EA3B mice compared with E mice that was not present at later stages (Extended Data Fig. 1b–d). No difference was observed in proliferation (Ki67) or DNA damage (γH2AX; Extended Data Fig. 1e,f). To assess if APOBEC activity contributes to increased tumor cell death at initiation, an EGFRL858R mouse model combined with a catalytically inactive form of A3B (E(CAG)A3BE255A)22,23 was generated (Fig. 1h). The decrease in EGFRL858R+ cells at 3 months postinduction observed with wildtype (WT) A3B was no longer observed in the enzyme inactive A3B mouse model (E(CAG)A3BE255A) compared with E control mice (Fig. 1h–j), suggesting that the increase in tumor cell death with A3B expression is at least in part due to the enzymatic activity of A3B.
We hypothesized that A3B expression could drive increased tumor cell death through enhanced immune surveillance in response to increased A3B activity24. A significant increase in both CD4 and CD8 T cells in EA3B mice was observed at 3 months postinduction (Extended Data Fig. 1g–i). Transplantation of an EPA3B mouse tumor cell line into WT C57BL/6J or EPA3B C57BL/6J transgenic mice resulted in the growth of EGFRL858R+ A3B+ tumors in EPA3B C57BL6/J transgenic mice but not WT C57BL/6J mice (Extended Data Fig. 1j–m), suggesting a level of immune tolerance to both the EGFRL858R and A3B transgenes.
Tumors were induced in an EGFRL858R p53-deficient mouse model either with or without A3B (EP and EPA3B; Fig. 1k and Extended Data Fig. 1j). No difference in the number of tumors at 3 months postinduction (Fig. 1l) or in overall survival (Fig. 1m) was observed in EP versus EPA3B mice, suggesting that A3B expression is tolerated in a p53-deficient model of EGFR-driven lung cancer. Thus, p53 in this model limits the tolerance of cancer cells to A3B expression at tumor initiation.
Next, CIN was assessed in systemic treatment-naïve (TN) patients with lung adenocarcinoma from the TRACERx421 (Tx421) cohort, confirming and expanding on previous findings from Tx100 (ref. 4). Tracking NSCLC evolution through therapy (TRACERx) is a prospective multicenter cancer study designed to delineate tumor evolution from diagnosis and surgical resection to either cure or disease recurrence. Tx100 was the analysis of the first 100 patients enrolled9, while Tx421 was the analysis of the first 421 patients enrolled25. We considered the following three orthogonal approaches to estimate the extent of CIN in tumors: chromosome missegregation errors captured during anaphase; the amount of somatic copy-number alteration (SCNA) intratumor heterogeneity (ITH) between tumor regions (SCNA ITH)25 and expression-based 70-gene CIN signature (CIN70)4,26. We observed a significant correlation between all three measures of CIN and A3B expression in both a subset of EGFRmut patients with lung adenocarcinoma in the Tx421 dataset (Fig. 2a–d) and patients with lung adenocarcinoma in the Tx421 dataset (Fig. 2e–h). Focusing on the genomic data, we observed a significant correlation between SCNA ITH and mutations in an APOBEC context (TCN/TCW C>T/G; Fig. 2i). These data together suggest that the increased CIN observed with A3B expression in EGFRmut mouse models is reflected in human NSCLCs in the Tx421 dataset.

a, Correlation between APOBEC3B (A3B) expression and percent missegregation errors calculated using patients with EGFRmut lung adenocarcinoma (n = 13 tumors; Spearman, R = 0.59; P = 0.038). b, Significant correlation between A3B expression and CIN70 GSEA score calculated using EGFRmut tumors from patients with lung adenocarcinoma (n = 19 tumors; Spearman, R = 0.59; P = 0.009). c, Significant correlation between A3B expression and CIN70 GSEA score calculated using EGFRmut tumor regions in patients with lung adenocarcinoma (n = 42 tumor regions; Spearman, R = 0.64; P < 9 × 10−6). d, Correlation between A3B expression and subclonal CIN fraction calculated in EGFRmut patients with lung adenocarcinoma (n = 19 tumors; bootstrapped Spearman, R = 0.5; P = 0.032). e, Significant correlation between percent missegregation errors (anaphase bridges (bridges) and lagging chromosomes (lagging)) and CIN70 score calculated using tumors from patients (n = 112 tumors; Spearman, R = 0.27; P = 0.0038). f, Significant correlation between A3B expression and CIN70 GSEA score calculated using tumors from patients with lung adenocarcinoma (n = 188 tumors; Spearman, R = 0.56; P < 2 × 10−16). g, Significant correlation between A3B expression and CIN70 GSEA score calculated using tumor regions in patients with lung adenocarcinoma (n = 466 tumor regions; Spearman, R = 0.54; P < 2 × 10−16). h, Correlation between A3B expression and subclonal CIN fraction calculated patients with lung adenocarcinoma in the Tx421 cohort (n = 168 tumors; bootstrapped Spearman, R = 0.26; P = 0.00087). i, Comparisons between C>T and C>G mutation counts at TCN and TCW trinucleotide context and percentage of genome altered subclonally (n = 25, two-sided Pearson, TCW R = 0.49, P = 0.015; TCN R = 0.52, P = 0.0092). j, Comparison of clonal and subclonal APOBEC-associated mutation signature (clonal APOBEC–subclonal APOBEC) in patients with EGFR driver mutations (1, 1a, exon 19 deletion). White bars indicate that the patient is TP53 WT or has a subclonal TP53 mutation. Red bars indicate that the patient has a clonal TP53 mutation (n = 23, one-sided Wilcoxon, P = 1 × 10−4). k, Number of APOBEC-associated mutations in patients with EGFR driver mutations (1, 1a, exon 19 deletion). Colors indicate clonal or subclonal APOBEC or non-APOBEC-associated mutations (n = 23). All analyses were performed on samples from the Tx421 cohort. GSEA, gene set enrichment analysis; NES, normalized enrichment score; TMM, trimmed mean of M values.
Subclonal A3B inhibits tumorigenesis
Analysis of TN patients in the Tx421 cohort revealed that APOBEC-mediated mutagenesis is enriched subclonally in EGFRmut disease (Fig. 2j,k) and the wider cohort9. Mice in which A3B expression could be temporally separated from EGFRL858R expression (EA3Bi), allowing for induction of A3B expression in a subset of tumor cells within the already proliferating EGFRmut tumor, were generated to mirror subclonal APOBEC induction and to assess if subclonal A3B expression decreased tumor cell death observed at initiation11,12,21,27 (Extended Data Fig. 2a). EA3Bi mice had significantly lower tumor nodules per lung section and tumor area per lung area compared with E control mice (Extended Data Fig. 2b,c) along with significantly higher survival (Extended Data Fig. 2d). These data suggest that subclonal A3B also inhibits tumor growth, confirming the phenotype previously observed when A3B was induced concomitantly with EGFRL858R (Fig. 1a). Both mouse models (Fig. 1a and Extended Data Fig. 2a) are p53 WT.
A3B promotes tyrosine kinase inhibitor (TKI) resistance
Next, the impact of A3B on tumor evolution with EGFR TKI therapy was examined. Subclonal expression of A3B in TKI-treated EA3Bi mice drove a significant increase in tumor grade, tumor nodules per lung section and tumor area per tissue area compared with TKI-treated Ei control mice (Fig. 3a–d). Heterogeneous A3B tumor positivity (Fig. 3e) and a significant increase in A3B positivity with TKI therapy compared to untreated EA3Bi mice were observed (Fig. 3f). In an additional experiment, tumor growth and progression with TKI treatment were associated with a significant increase in tumor nodules and a substantial increase in tumor grade in EA3Bi mice compared with Ei control mice (Fig. 3g–i). Based on previous work illustrating an important role for uracil DNA glycosylase (UNG) in repairing APOBEC-induced uracil lesions28, we evaluated UNG expression in A3B-expressing EA3Bi tumors. Staining for UNG revealed a significant decrease in UNG-positive cells per tumor in EA3Bi mice compared with Ei mice treated with TKI therapy (Fig. 3j,k). Taken together, these findings suggest that subclonal A3B expression with TKI therapy in conjunction with UNG downregulation contributes to increased tumor growth and TKI resistance.

a, TetO-EGFRL858R;CCSP-rtTA;R26LSL-APOBEC3B/Cre-ER(T2) mice with or without induction of subclonal APOBEC3B (A3B) with TKI therapy (erlotinib). b, Fraction of tumor grade, not present or hyperplasia only. Bronchioloalveolar adenoma or carcinoma at 5 months (Ei, n = 19; EA3Bi, n = 19; two-sided Fisher’s exact test, **P = 0.0044). c, Tumor nodules per lung section per mouse at 5 months (Ei, n = 19; EA3Bi, n = 19; two-sided Mann–Whitney test, *P = 0.0443). d, Tumor area per lung area per mouse at 5 months (Ei, n = 19; EA3Bi, n = 19; two-sided Mann–Whitney test, *P = 0.0212). e, Representative IHC staining of EGFRL858R and A3B (scale bar = 100 µm and 20 µm; Ei, n = 19; EA3Bi, n = 19 biological replicates). f, A3B+ cells per mm2 of tumor per mouse (EA3Bi −TKI = 151, EA3Bi +TKI = 52, two-sided Mann–Whitney test, ****P < 0.0001). g, Induction of subclonal A3B using TetO-EGFRL858R;CCSP-rtTA;R26Cre-ER(T2)/+ or TetO-EGFRL858R;CCSP-rtTA;R26LSL-APOBEC3B/Cre-ER(T2) mice with continuous TKI therapy (erlotinib). h, Tumor nodules per lung section per mouse (Ei, n = 13; EA3Bi, n = 17; two-sided Mann–Whitney test, **P = 0.0086). i, Fraction of tumor grade, not present or hyperplasia only. Bronchioloalveolar adenoma or carcinoma at 11 months (Ei, n = 13; EA3Bi, n = 17; two-sided Fisher’s exact test). j, Quantification of UNG+ cells per mm2 of tumor at 5 months postinduction (E, n = 10; EA3Bi, n = 10; two-tailed t test, *P = 0.0226, each dot represents a tumor). k, Representative IHC staining of EGFRL858R and UNG. Scale bar = 50 µm. l–n, CellTiter-Glo viability timecourse assays performed on A3B-deficient or A3B-proficient PC9 cells treated with 100 nM Osi (l, n = 3 biological replicates, mean ± s.d., two-sided t test, *P = 0.0439, *P = 0.0155, *P = 0.0168); HCC827 cells treated with 100 nM Osi (m, n = 3 biological replicates, mean ± s.d., two-sided t test, *P = 0.0377, **P = 0.0029, ****P = 0.0004, ****P = 0.00009); H3122 cells treated with 100 nM alectinib (n, n = 3 biological replicates, mean ± s.d., two-sided t test, *P = 0.0189, **P = 0.0044). Osi, osimertinib.
Next, whole-exome sequencing (WES) was performed on TN and matched TKI-resistant mouse tumor cell lines (Extended Data Fig. 3a,b and Supplementary Table 1). A significantly higher number of mutations, as well as mutations in an APOBEC context, were detected in TKI-resistant A3B-expressing EGFRmut tumor cell lines (EPA3B) compared with control TKI-resistant EGFRmut tumor cell lines (EP), and compared with both control (EP) and A3B-expressing TN EGFRmut tumor cell lines (Extended Data Fig. 3a,b). Two unique de novo putative loss-of-function mutations in the protein tyrosine phosphatase receptor type S (Ptprs) gene were identified in an APOBEC context (Extended Data Fig. 3c). Loss of PTPRS function through mutation or deletion has been shown to increase TKI resistance in multiple human preclinical cancer models and has been linked with worse overall survival and more rapid disease progression in patients with EGFR-driven lung cancer29,30,31. The equivalent of the A3B-driven mutation in humans (Ptprs_mut1, D138N; Extended Data Fig. 3c) was identified in tumors of patients with lung, colorectal and bladder cancer from The Cancer Genome Atlas (TCGA) and in one EGFRL858R TRACERx patient with NSCLC (Extended Data Fig. 3d).
To validate our findings from mouse models, long-term cell viability with targeted therapy was assessed in established human cell line models of oncogenic EGFRmut and echinoderm microtubule-associated protein-like 4-anaplastic lymphoma kinase (EML4-ALK) lung adenocarcinoma with CRISPR-mediated A3B depletion. Under EGFR TKI treatment (osimertinib), A3B-depleted PC9 and HCC827 lines (harboring EGFRexon19del; Extended Data Fig. 4a–d) showed significantly reduced cell viability compared to A3B-competent control lines (Fig. 3l,m). Similarly, a significant reduction in cell viability was observed in an A3B-knockout (KO) EML4-ALK cancer cell line (H3122; Extended Data Fig. 4e,f) treated with the Food and Drug Administration-approved ALK TKI alectinib (Fig. 3n). KO of A3B had no effect on cell viability in untreated PC9, HCC827 or H3122 cell lines (Extended Data Fig. 4g–i). These data suggest that A3B expression confers enhanced cell survival with targeted therapy.
Targeted therapy induces A3B expression and UNG downregulation
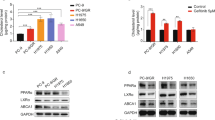
Our mouse lung cancer models demonstrated that A3B expression is associated with targeted therapy resistance. We hypothesized that targeted therapy may induce adaptations that increase the expression of A3 family members and decrease the expression of UNG in human models. Based on current literature4,5,32,33, mRNA expression levels of A3A, A3B, APOBEC3C (A3C) and APOBEC3F (A3F) were measured. In PC9 cells, a significant increase in all four members was observed with osimertinib, with A3A being the most significantly elevated (Fig. 4a). In HCC827 cells, A3A and A3B were the most significantly elevated, with both induced to similar levels with osimertinib (Fig. 4b). A significant increase in overall APOBEC activity (Fig. 4c,d) and A3B protein levels (Fig. 4e,f) were also observed. Each A3 gene was then silenced using small interfering RNAs (siRNAs) specific for each family member (Extended Data Fig. 5a–i), and APOBEC activity was assessed. Only knockdown of A3B resulted in a significant decrease in APOBEC activity with TKI therapy in PC9 and HCC827 cell lines (Fig. 4g). These data suggest that while several A3 family members likely contribute to the increased APOBEC activity observed with TKI therapy, A3B appears to be a major contributor.

a, RT–qPCR performed on PC9 cells treated with DMSO or 0.5 μM Osi for 18 h, measuring APOBEC3A (A3A), APOBEC3B (A3B), APOBEC3C (A3C) and APOBEC3F (A3F; n = 4 biological replicates, mean ± s.d., one-way ANOVA test, ***P = 0.0002, ****P < 0.0001). b, RT–qPCR analysis of HCC827 cells treated with DMSO or 0.5 μM Osi for 18 h (n = 3 biological replicates, mean ± s.d., one-way ANOVA test, *P = 0.0264, ***P = 0.0005, ***P = 0.0008, ****P < 0.0001). c, APOBEC activity assay performed using nuclear extracts of PC9 cells treated with DMSO or 2 μM Osi for 18 h (n = 3 biological replicates, mean ± s.d., two-tailed t test, ***P = 0.0002). d, APOBEC activity assay using nuclear extracts of HCC827 cells treated with DMSO or 0.4 µM Osi for 18 h (n = 3 biological replicates, mean ± s.d., two-tailed t test, *P = 0.0213). e, Western blot analysis of A3B protein levels in PC9 cells treated with DMSO or 0.5 μM Osi for 18 h with quantification (n = 3 biological replicates, mean ± s.d., two-tailed t test, *P = 0.0129). f, Western blot analysis for A3B protein levels in HCC827 cells treated with DMSO or 0.5 μM Osi for 18 h (n = 3 biological replicates, mean ± s.d., two-tailed unpaired t test, **P = 0.0082). g, APOBEC activity assay performed on lysates of PC9 or HCC827 cells treated with DMSO or 0.5 μM Osi for 18 h, with siRNA knockdown of APOBEC3A (siA3A), APOBEC3B (siA3B), APOBEC3C (siA3C) and APOBEC3F (siA3F) and nontargeting siRNA (siNTC), and quantification (PC9, n = 4 biological replicates, mean ± s.d., one-way ANOVA test (nonparametric), **P = 0.0017; HCC827, n = 3 biological replicates, mean ± s.d., one-way ANOVA test (nonparametric), **P = 0.0076). ANOVA, analysis of variance.
Targeted therapy-induced transcriptional changes of A3B and UNG were assessed in established human lung cancer cell line data from publicly available datasets (Gene Expression Omnibus (GEO) database, GEO2R). Treatment of EGFRmut cell lines (HCC827, PC9 and HCC4006 harboring EGFRL747-E749del,A750P) with the EGFR TKI erlotinib was associated with transcriptional upregulation of A3B both acutely (6-h to 1-d treatment) and at later timepoints (8-d treatment; Fig. 5a). These transcriptional changes were confirmed in an independent RNA-seq (RNA sequencing) dataset34 with a significant upregulation of A3B and downregulation of UNG following osimertinib treatment (Fig. 5b), suggesting a conserved effect of EGFR pharmacologic inhibition independent of the generation (evolution of targeted therapy development leading to more specific and effective molecules) of EGFR inhibitor.

a, GSEA of the indicated GEO2R datasets of EGFR-driven cellular models of human lung adenocarcinoma treated with erlotinib or a mitogen-activated protein kinase kinase (MAP2K or MEK1) inhibitor (AZD6244). b, RNA-seq analysis of gene expression changes in PC9 cells treated with 2 μM Osi for 9 d relative to DMSO-treated cells (n = 3 biological replicates, mean ± s.d., ANOVA test). c, RT–qPCR analysis of PC9 cells treated with DMSO or 2 μM Osi for 18 h (n = 4 biological replicates, mean ± s.d., one-way ANOVA test, *P = 0.0349, ****P < 0.0001). d, RT–qPCR analysis of HCC827 cells treated with DMSO or 0.4 μM osimertinib for 18 h (n = 4 biological replicates, mean ± s.d., one-way ANOVA test, ***P = 0.0008, **P = 0.0014). e, Western blot analysis of cells treated in a and b (CYTO, cytoplasmic extracts; H3, histone H3; NUC, nuclear extracts) with quantification of A3B levels in PC9 cells (n = 3 biological replicates, mean ± s.d., one-way ANOVA test, **P = 0.0012, **P = 0.0058) and HCC827 cells (n = 3 biological replicates, mean ± s.d., one-way ANOVA test, *P = 0.0186). f, RT–qPCR analysis of PC9 cells treated with nontargeting siRNA (siNTC) or EGFR siRNA (siEGFR) for 18 h and grown for 2 d (n = 4 biological replicates, mean ± s.d., two-sided t test, **P = 0.0075, ***P = 0.0002, **P = 0.0027). FC, fold change.
Transcriptional upregulation of A3B and downregulation of UNG were subsequently validated in multiple oncogenic EGFR-driven cellular models of lung adenocarcinoma at both the RNA (Fig. 5c,d) and protein levels (Fig. 5e). To rule out off-target pharmacological effects of EGFR TKIs, A3B expression was examined with siRNA-mediated silencing of EGFR and also led to A3B upregulation and UNG downregulation (Fig. 5f). Induction of A3B was also observed upon treatment with an inhibitor of mitogen-activated protein kinase kinase (MAP2K or MEK1 (selumetinib; Fig. 5a). The induction of A3B by different inhibitors of oncogenic receptor tyrosine kinases (RTKs) and their downstream signaling components, such as MEK1, indicates that upregulation of A3B is likely a consequence of oncogenic signaling inhibition, and not specific to EGFR TKIs.
Consistent with RNA and protein level changes, TKI treatment resulted in a significant increase in nuclear APOBEC activity35 and decrease in nuclear uracil excision capacity of UNG in multiple EGFR-driven cell line models, including EGFRexon19del cells (PC9 and HCC827) and EGFRL858R+T790M cells (H1975; Fig. 4c,d and Extended Data Fig. 6a–e). Increased A3B expression and APOBEC activity as well as decreased UNG expression and uracil excision activity were also observed in EML4-ALK-driven cellular models (H3122 and H2228) during ALK TKI treatment (Extended Data Fig. 6f–i).
A3B was then stably knocked down using small hairpin RNA (shRNA) in PC9 cells, and rescue experiments with expression vectors containing either WT A3B tagged with human influenza hemagglutinin (HA) (A3B WT-HA tagged) or catalytically inactive A3B tagged with HA (A3B E225A-HA tagged) were performed. APOBEC activity with A3B knockdown was significantly reduced with TKI treatment versus A3B-proficient lines with TKI treatment (Extended Data Fig. 6j). Expression of the WT catalytically active, but not the mutant catalytically inactive A3B, rescued the decline in nuclear APOBEC activity caused by A3B depletion (Extended Data Fig. 6j–l). While knockdown of A3B induced no off-target reductions in any other A3 family members, significant increases in A3A, A3G and A3H expression were detected (Extended Data Fig. 6m), corroborating previous reports in human breast and lymphoma cancer cell lines showing increased A3A expression with A3B loss36. These data suggest that A3B is a substantial contributor to the increased APOBEC activity observed with TKI treatment.
To exclude an indirect effect of targeted therapy on cell cycle arrest that might alter APOBEC enzyme expression, EGFRmut NSCLC PC9 cells were treated with the CDK4/6 cell cycle inhibitor palbociclib37. Palbociclib treatment-induced G0/G1 cell cycle arrest with a comparable arrest measured with osimertinib (Extended Data Fig. 6n). UNG expression decreased upon palbociclib treatment; however, there was a significant decline in A3B expression (Extended Data Fig. 6o), contrasting with the increased expression observed upon TKI therapy and suggesting that TKI-mediated induction of A3B is unlikely to be a consequence of TKI treatment-induced cell cycle inhibition.
A3B and UNG expression levels were then examined in multiple human tumor xenograft models. An increase in A3B and a decrease in UNG protein levels were detected in EGFR TKI-treated tumor tissues from three distinct oncogenic EGFR-driven CDX models of human lung adenocarcinoma (Extended Data Fig. 7a–f). Additionally, RNA-seq analyses from an EGFRL858R-harboring patient-derived xenograft (PDX) model of lung adenocarcinoma38 revealed a nonsignificant increase in A3B mRNA and a decrease in UNG mRNA levels upon treatment with erlotinib (Extended Data Fig. 7g), and significant increase in A3B and a nonsignificant decrease in UNG with osimertinib34 (Extended Data Fig. 7h). These findings support a model whereby EGFR oncoprotein inhibition induces increased A3B expression and decreased UNG expression.
Nuclear factor-kappa B (NF-κB) signaling contributes to TKI-induced A3B upregulation
Prior work from our group and others revealed that NF-κB signaling is activated upon EGFR oncogene inhibition in human lung cancer as a stress and survival response38. Previous data suggest that NF-κB signaling may be a prominent inducer of A3B gene expression39,40. We hypothesized that NF-κB signaling activation upon targeted therapy promotes A3B upregulation. To test this hypothesis, an established RNA-seq dataset generated from EGFR-driven human lung adenocarcinoma cells treated acutely with either erlotinib or an NF-κB inhibitor (PBS-1086) or both in combination was examined38. TKI treatment-induced transcriptional upregulation of A3B was attenuated by cotreatment with the NF-κB inhibitor38 (Extended Data Fig. 8a), suggesting that the NF-κB pathway induces A3B expression. To confirm this, the NF-κB pathway was activated with increasing concentrations of Tumor necrosis factor-α, which elevated nuclear RELA and RELB as well as nuclear A3B protein levels (Extended Data Fig. 8b) and cellular A3B mRNA expression (Extended Data Fig. 8c). Inhibition of the NF-κB pathway by simultaneous depletion of both RELA and RELB (Extended Data Fig. 8d) reduced TKI-induced A3B mRNA expression (Extended Data Fig. 8e) and A3B protein levels (Extended Data Fig. 8f). Co-inhibition of EGFR and NF-κB pathways blocked EGFR inhibition-induced A3B upregulation in oncogenic EGFR-driven NSCLC xenografts (Extended Data Fig. 7c,d). Codepletion of both NF-κB transcription factors RELA and RELB impaired TKI-induced nuclear APOBEC activity (Extended Data Fig. 8g). These data support NF-κB activation with EGFR TKI treatment as an inducer of A3B upregulation in response to therapy.
To investigate the clinical relevance of these findings, we examined single-cell RNA-seq data in an established dataset obtained from clinical specimens of NSCLC procured from patients at the following three timepoints: (1) treatment naïve before initiation of systemic targeted therapy (classified as TN), (2) while on targeted therapy when the tumor was regressing or at stable state as evaluated by standard clinical imaging (classified as residual disease (RD)) and (3) at clear progressive disease (PD, acquired resistance) as determined by standard clinical imaging (classified as PD). The classification of response was based on Response Evaluation Criteria in Solid Tumors (RECIST) criteria41. In total, 66 samples obtained from 30 patients with lung cancer pre-TKI or post-TKI therapy (erlotinib (EGFR), osimertinib (EGFR) and crizotinib (ALK) being the most frequent targeted therapies) were analyzed (Supplementary Table 2a). We observed that mRNA expression of A3B and NF-κB components RELA and RELB, as well as an NF-κB gene signature42, were significantly increased in tumors exposed to EGFR TKI treatment, in particular at tumor progression with therapy (Extended Data Fig. 8h–k).
UNG downregulation is associated with c-JUN suppression during TKI treatment
We next investigated the mechanism of UNG downregulation during targeted therapy. UNG gene promoter analysis (using PROMO)43 revealed the presence of predicted JUN consensus binding sites. RNA-seq data from EGFR TKI-treated PC9 cells indicated that like UNG, c-JUN was also transcriptionally downregulated upon treatment, which was validated using RT–qPCR (Extended Data Fig. 8l). This aligns with the expected downregulation of c-JUN upon inhibition of the mitogen-activated protein kinase (MAPK) pathway during EGFR inhibition by TKI treatment44. We hypothesized that TKI treatment-induced UNG downregulation could be caused by c-JUN downregulation. Silencing of c-JUN by siRNA was sufficient to suppress UNG expression, suggesting that UNG downregulation could be a consequence, in part, of the c-JUN suppression that occurs during TKI-mediated MAPK signaling suppression (Extended Data Fig. 8m).
A3B is required for APOBEC mutation signature accumulation during targeted therapy
To examine the role of A3B expression on mutagenesis during targeted therapy, A3B-deficient and A3B-proficient single-cell cloned PC9 cells (Extended Data Fig. 4a,b) were treated with osimertinib using a dose-escalation protocol to resistance (3 months; Fig. 6a). The mutations and proportion of APOBEC mutation signatures (SBS2 + SBS13) acquired were quantified following whole-genome sequencing (WGS; Fig. 6a–g, and Extended Data Fig. 9a). This revealed that only A3B-proficient lines gained APOBEC mutation signatures (SBS2 + SBS13) during TKI treatment (Fig. 6b,f,g and Supplementary Table 3). Examination of the fraction of mutations in an APOBEC context (TCW C>T/G) revealed a significant decrease in A3B-deficient lines (Fig. 6c). Examination of APOBEC pentanucleotide sequences6,32,36,45 in the osimertinib-treated A3B-deficient and A3B-proficient groups (Fig. 6d,e) revealed significant decreases in the fraction of APOBEC mutations in an A3B-preferred RTCW context in A3B-deficient clones, with no significant decrease in mutations in a A3A-preferred YTCW context (Fig. 6d,e). These data suggest that A3B is required for the accumulation of APOBEC mutations during TKI treatment.

a, Outline of WGS long-term TKI treatment experiment on APOBEC3B (A3B)-deficient and A3B-proficient PC9 single-cell clone lines. Figure created in BioRender.com. b, Focused plots showing APOBEC signature (SBS2 + SBS13) burden in the indicated A3B-deficient (A3B KO) and A3B-proficient (A3B WT) PC9 clones (A3B WT, n = 6 biological replicates; A3B KO, n = 6 biological replicates). c, Fraction of mutations in an APOBEC context (TCW C>T/G) of total mutations per replicate, of Osi-treated A3B WT and A3B KO cells (all data points shown, n = 6 biological replicates, mean ± s.d., two-tailed Mann–Whitney test, **P = 0.0043). d, Fraction of APOBEC mutations (RTCW C>T/G) of total mutations per replicate Osi-treated A3B WT and A3B KO cells (all data points shown, n = 6 biological replicates, two-tailed Mann–Whitney test, **P = 0.0022). e, Fraction of APOBEC mutations (YTCW C>T/G) of total mutations per replicate in Osi-treated A3B WT and A3B KO cells (all data points shown, n = 6 biological replicates, two-tailed Mann–Whitney test, P = 0.0931). f, Profiles of APOBEC-associated signatures SBS2 and SBS13 from the Catalogue of Somatic Mutations in Cancer (COSMIC) (cancer.sanger.ac.uk). g, Mutational profiles of A3B KO and A3B WT Osi-treated PC9 cell lines. Mutational profiles are plotted as the number of mutations (y axis) at cytosine or thymine bases classified into 96 possible trinucleotide sequence contexts (asterisk indicates cell lines that acquired APOBEC signature during TKI treatment timecourse (SBS2 + SBS13; A3B WT, n = 6 biological replicates; A3B KO, n = 6 biological replicates)).
To further explore this hypothesis, we analyzed sequencing data for potential TKI resistance mutations in A3B-proficient PC9 TKI-resistant clones and found an acquired early stop codon mutation in the tumor suppressor gene NRXN3 (Q54*)46,47 in an APOBEC-preferred context (T(C>T)A). The potential impact of this loss-of-function mutation was validated by depleting NRXN3 (given the early stop codon mutation detected, which is likely a loss-of-function event) in a naïve PC9 lung cancer cell line, which increased levels of phosphorylated AKT, a previously identified convergent feature of EGFR TKI resistance48, and conferred resistance to EGFR TKI treatment (Extended Data Fig. 9b–d).
A3B expression and APOBEC-associated mutations are elevated with targeted therapy in NSCLC
To verify the clinical relevance of our findings, A3B expression was examined in several NSCLC clinical datasets (Supplementary Table 2b)41,49,50,51,52. Bulk RNA-seq of 32 pre-TKI and 42 post-TKI treated (osimertinib/erlotinib/crizotinib/alectinib) clinical tumor samples revealed a significant increase of A3B expression post-TKI relative to pre-TKI samples (P = 0.011; Fig. 7a). A3B was the only A3 family member with significantly increased expression post-TKI treatment (Extended Data Fig. 10a). Stratification at TN, RD and PD timepoints revealed a significant expression increase from TN to RD (P = 0.02) and an increase approaching significance from TN to PD (P = 0.057; Extended Data Fig. 10b). Further validating these observations, single-cell RNA-seq data revealed that A3B expression, specifically in tumor cells isolated from clinical specimens, was significantly increased from TN to PD (P < 0.001) and from RD to PD (P < 0.001; Fig. 7b). Compared to the other A3 genes, A3B expression had the second highest effect scores of all A3 family members as calculated using Cohen’s d method (TN to PD, d = 1.048; RD to PD, d = 0.953; Extended Data Fig. 10c). A3C expression exhibited the highest effect scores; however, APOBEC activity assays revealed A3C did not contribute to overall activity with TKI treatment (Fig. 4g). Immunohistochemical (IHC) analyses, as performed previously4, on clinical samples also revealed a significant increase in A3B nuclear protein levels in EGFR TKI-treated tumor samples both at RD and PD timepoints (Fig. 7c,d and Supplementary Table 2c).

a, APOBEC3B (A3B) expression levels (batch-corrected transcripts per million (TPM)) measured using RNA-seq analysis in human NSCLC specimens driven by EGFR- and ALK-driver mutations obtained before TKI treatment (pre-TKI, n = 32 samples) or post-treatment (post-TKI, n = 42 samples; all data points shown, two-sided t test, *P = 0.02). b, APOBEC family member expression measured using single-cell RNA-seq obtained from human NSCLC before TKI treatment (TN), on-treatment at RD or at PD (all data points shown, n = 762, 553 and 988 cells per group, respectively, two-sided Wilcoxon test with a Holm correction, ****P < 2.22 × 10−16). c,d, Representative images of IHC analysis of A3B protein levels in patients with NSCLC at TN, RD and PD stages. Red arrows indicate positive stained cells (scale bar: 30 µM, c) with IHC quantification of human NSCLC samples pre-TKI (n = 16 samples) or post-TKI single agent (n = 15 samples; all data points shown, two-sided unpaired t test, *P = 0.0113, d). e, Total mutation burden (SNV count) in paired human NSCLC samples pre-TKI or post-TKI (n = 32, two-tailed Wilcoxon matched-pairs signed-rank test, **P = 0.0013). f, APOBEC-associated mutation count in paired human NSCLC samples pre-TKI or post-TKI (n = 32, two-tailed Wilcoxon matched-pairs signed-rank test, *P = 0.0155). g, Mutation signature associated with each putative de novo TKI resistance mutation detected in clinical samples analyzed post-TKI at PD. An asterisk denotes a sample from a patient who has received prior chemotherapy. Boxplots: middle line represents median; lower and upper hinges represent the first and third quartiles; lower and upper whiskers represent smallest and largest values within 1.5× interquartile range from hinges.
Demonstrating the clinical effect of TKI treatment on the proportion of mutational signatures, a recently published dataset shows that APOBEC-associated mutation signatures (SBS2 and SBS13) were dominant, defined as the mutational signature with the highest fraction of mutations, in a significantly higher number of osimertinib-resistant samples when compared with naïve samples53. To independently test this observation with our own data, WES was performed on paired pre- and post-TKI treated samples obtained from 32 patients (Supplementary Table 4) to quantify mutations acquired following TKI treatment in NSCLC EGFRmut (treated with erlotinib/osimertinib) and ALK fusion (treated with alectinib) clinical samples. This analysis revealed that both the overall mutation burden (SNV count; Fig. 7e) and number of APOBEC-associated mutations (C>T or C>G mutations in a TCN context; Fig. 7f) increased post-treatment.
Next, mutations in an APOBEC-preferred context were identified in genes previously associated with TKI resistance in tumors from patients who had progressed on or shown incomplete response to EGFR inhibitor therapy (Fig. 7g and Supplementary Table 5). These mutations include activating mutations in PIK3CA (E545K)54, WNT signaling-activating mutations in β-catenin at a glycogen synthase kinase-3β (GSK-3β) phosphorylation site55, MAPK pathway reactivating-mutations through inactivation of PP2A, a negative regulator of MAPK signaling56,57, an activating mutation in MET tyrosine kinase domain (H1095Y)53,58, as well as an ALK inhibitor desensitizing mutation in ALK (E1210K)59 in the tumors of some patients who had progressed on or shown incomplete response to EGFR or ALK inhibitor therapy. AKT, WNT and MAPK pathway activation have previously been shown to cause EGFR and ALK inhibitor resistance60,61,62,63,64,65. All but one of these APOBEC-associated putative resistance mutations were detected selectively post-treatment, suggesting not only that these mutations are induced by APOBEC (itself engaged) during targeted therapy but also that these variants could promote resistance. All samples containing these APOBEC-associated mutations, except for one, did not harbor a detectable EGFR T790M mutation, which has been reported to be present in ~50–60% of first- and second-generation EGFR TKI-resistant cases66,67 and arising from a non-APOBEC clock-like mutation signature (SBS1 (ref. 68); Fig. 7g). Altogether, of the resistance mutations in this cohort, 53% (8/15) of mutations were associated with clock-like mutation signature SBS1 and 46% (7/15) of mutations with the APOBEC signatures SBS2 + SBS13, with no other mutational signatures contributing to putative resistance mutations. In total, 8/32 tumors have APOBEC-associated putative resistance mutations. The observation that APOBEC-mediated mutations in resistance-associated genes detected in post-treatment samples and the EGFR T790M mutation appear to be mutually exclusive suggests that these APOBEC-mediated mutations could be the potential mechanism of resistance to targeted therapy in these patients. These data suggest that APOBEC signatures are a complementary route to acquired TKI therapy resistance, contributing to the diverse mechanisms of resistance that exist69,70,71.
Taken together, these data illustrate the diverse effects of A3B at different stages of tumor evolution with or without the selective pressure of therapy. The findings demonstrate multiple roles of A3B, as an inhibitor of tumor progression at initiation, an inducer of APOBEC mutations and a contributor to targeted therapy resistance (Fig. 8).

At tumor initiation, continuous APOBEC3B expression and activity induces CIN and p53 pathway activation, resulting in cell death. With targeted therapy, NF-κB induction leads to increased A3B expression, fueling TKI resistance. Figure created in BioRender.com.
Discussion
Our collective findings shed light on the important, context-specific roles of A3B on lung cancer pathogenesis and tumor evolution. Along with other recent findings in the field5, our data reinforce the concept that targeted therapies can induce adaptive changes that promote resistance72, including those that are APOBEC-mediated and that may involve multiple APOBEC family members. This A3 induction during therapy might contribute to the development of treatment resistance and appears to be clinically relevant based on our clinical datasets obtained from targeted therapy-treated patients. Additional clinical cohort analyses will be important to conduct as further human tumors obtained from patients on targeted therapy become available.
We demonstrate that the expression of A3 family members might contribute to resistance in preclinical human and mouse models of lung adenocarcinoma. Although we focus on oncogenic EGFR-driven lung adenocarcinomas, our findings appear to extend to other molecular subsets such as EML4-ALK-driven lung cancer (Fig. 3l and Extended Data Fig. 6b–d) and likely reflect a more general principle of targeted therapy-induced adaptability. While APOBEC has been implicated in drug resistance previously33,73, our study reveals a distinct mechanism by which targeted cancer therapy is actively responsible for the upregulation of APOBEC via NF-κB-mediated transcriptional induction in response to therapy. Our study further explains the enhanced efficacy of cotreatment with an NF-κB inhibitor compared to EGFR inhibition alone at preventing the emergence of resistance38.
There are however caveats to our findings (further discussion in Supplementary Note). The mouse models, although helpful for a deeper understanding of the biological effects of enforced A3B expression, are imperfect as A3B is expressed from a transgene promoter system. APOBEC3 enzyme expression has also been shown to occur episodically32, which differs from the constitutive expression of our mouse models. Future studies that reveal the upstream regulators of endogenous mouse APOBEC enzymes could help in the development of better models in future studies.
Our work expands upon prior studies suggesting a potential association between APOBEC-mediated mutagenesis and acquisition of putative resistance mutations in the APOBEC-preferred context during the treatment of EGFR-driven lung cancers74,75. Our data suggest that inhibition of APOBEC3 family members could suppress the emergence of one pathway to resistance and thereby improve response to targeted therapy, consistent with the work of others in the field that suggests that multiple APOBEC3 family members including A3B contribute to targeted therapy resistance5,32, with both A3A and A3B shown to be contributors of mutagenesis6,32,36,76. The role of A3B in promoting resistance to TKI is likely multifaceted, and our data do not discount the contribution of other possible parallel cytosine deaminase-independent mechanisms, such as induced CIN4,77, regulation of cell cycle22 and regulation of the DNA damage repair pathway78,79. Our evidence here and these emerging collective findings5,33,80,81 suggest that endogenous drivers of mutagenesis have diverse roles that are both detrimental and beneficial to tumor evolution depending on the context of tumor pathogenesis and treatment.
Methods
Cell line and growth assays
Cell lines were grown in Roswell Park Memorial Institute-1640 medium (RPMI-1640) with 1% penicillin–streptomycin (10,000 U ml−1) and 10% FBS or in Iscove’s modified Dulbecco’s medium (IMDM) with 1% penicillin–streptomycin (10,000 U ml−1), l-glutamine (200 mM) and 10% FBS in a humidified incubator with 5% CO2 maintained at 37 °C. Drugs used for treatment except PBS-1086 (ref. 38) were purchased from Selleck Chemicals or MedKoo Biosciences. For growth assays, cells were exposed to DMSO or the indicated drugs for indicated durations in six-well or 96-well plates and assayed using crystal violet staining or Celltiter-Glo luminescent viability assay (Promega) according to the manufacturer’s instructions.
Deriving clonal populations and generating APOBEC3B KO cells
Clonal cells were derived by sorting single cells into 96-well plates and expanding them over a few weeks. We then derived pools of one of the clones expressing either a green fluorescent protein (GFP)-targeting or A3B-targeting guide along with CRISPR/Cas9 by lentiviral transduction as done in a previously published study82. A3B gRNA target sequences, designed by the Zhang Lab83, were subcloned into the lentiCRISPR v2 plasmid (Addgene, 52961; a gift from F. Zhang)83 and the one that showed better A3B depletion was selected for further analysis.
Transductions and transfections
Hek293T cells were cotransfected with lentiviral packaging plasmids pCMVdr8 and pMD2.G plasmid, along with the plasmid of interest using FuGENE 6 Transfection Reagent (Promega). APOBEC3B shRNA was purchased from Sigma (TRCN0000142875). Cells were transduced with 1:1 diluted lentivirus for 1–2 d and selected with antibiotic marker (puromycin). siRNAs were purchased from GE Healthcare Dharmacon and transfected using Lipofectamine RNAi Max according to the manufacturer’s protocol, and the cells were collected within 48 h of transfection for subsequent assays.
RT–qPCR
Total RNA was extracted using GeneJet RNA purification kit (Thermo Fisher Scientific) or RNeasy Mini kit (Qiagen), and cDNA was synthesized from it using sensiFast cDNA Synthesis Kit or High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) in accordance with the manufacturer’s instructions. qPCR reactions were performed using PowerUP SYBR Green Master Mix (Applied Biosystems) or TaqMan Universal PCR Master Mix (Applied Biosystems) and previously validated primers84 (PrimerBank) on a QuantStudio. Glyceraldehyde-3-phosphatase dehydrogenase (GAPDH), actin, 18S RNA or β2-microglobulin were used as reference genes. The following primers were used for p53 pathway activation: actin: Mm02619580_g1, Bax: Mm00432051_m1, Cdkn1a/p21: Mm04205640_g1, Mdm2: Mm01233138_m1, Pmaip1/Noxa: Mm00451763_m1 and Sesn2: Mm00460679_m1. Data were analyzed using QuantStudio 12K Flex Software (v1.3) and GraphPad Prism.
Western blot assay
Whole-cell extracts were collected in RIPA buffer containing protease and phosphatase inhibitors followed by sonication and centrifugation for clarification of extracts. Nuclear-cytoplasmic extracts were collected as described previously with 0.1% nonidet P-40 (NP-40) in PBS85. Extracts were quantified using Lowry assay, run on 4–15% Criterion TGX Gels (Bio-Rad) and transferred to a nitrocellulose membrane with Trans-Blot Turbo RTA Midi Nitrocellulose Transfer Kit (Bio-Rad). Membranes were blocked in 3% milk in tris-buffered saline with 0.1% Tween 20 (TBST), incubated with primary antibody overnight followed by secondary antibody, either horse radish peroxidase (HRP)-conjugated or fluorescently labeled, for 1–2 h and imaged on a LI-COR imager or ImageQuant LAS 4000 (GE HealthCare). Anti-APOBEC3B (5210-87-13)86 and anti-UNG28 antibodies were kindly provided by R. Harris, and anti-GAPDH antibody (sc-59540) was purchased from Santa Cruz Biotechnology. Anti-EGFR (4267), anti-phospho-EGFR (Y1068, 3777 or 2236), anti-STAT3 (9139), anti-phospho-STAT3 (Y705, 9145), anti-AKT (2920), anti-phospho-AKT (S473, 4060), anti-phospho-ERK (T202, Y204; 4370 or 9106), anti-ERK (9102), anti-RELA (8242), anti-RELB (4922), anti-HSP90 (4874), anti-TUBB (2146) and anti-histone H3 (9715) were purchased from Cell Signaling Technology (CST). All primary antibodies were used at a dilution of 1:1,000.
Enzymatic assays
APOBEC assays were performed by incubating nuclear extracts from rapid efficient and practical (REAP) method58 or whole-cell extracts with the following DNA oligo substrates (Integrated DNA Technologies, IDT): 5′-ATT ATT ATT ATT CAA ATG GAT TTA TTT ATT TAT TTA TTT ATT T-FAM-3′ using established protocols28,35. Upon completion of the reactions, they were heated at 95 °C for 5 min after the addition of TBE-urea buffer (Novex) and immediately run on a 15% TBE-urea gel (Bio-Rad) and imaged using Cy2 filter on ImageQuant LAS 4000.
Subcutaneous tumor xenografts and PDX studies
All animal experiments were conducted under University of California, San Francisco (UCSF) Institutional Animal Care & Use Committee (IACUC)-approved animal protocols. PC9 and H1975 tumor xenografts were generated by injection of 1 million cells in a 1:1 mixture of matrigel and PBS into 6- to 8-week-old female non-obese diabetic/severe combined immunodeficiency disease (NOD/SCID) mice. Once the tumors grew to ∼100 mm3, the mice were treated with vehicle or 5 mg kg−1 osimertinib once daily by oral gavage and the tumors were collected on day 4 for western blot analysis. PDX was generated as indicated in a previous study38. Tumors were passaged in SCID mice, treated with 25 mg kg−1 erlotinib once daily by oral gavage once they reached ~400 mm3 and collected on day 2.
Mouse strains and tumor induction and treatment
The Cre-inducible Rosa26::LSL-APOBEC3Bi mice and Rosa26::CAG-LSL-APOBEC3Bi-E255A are described in refs. 20,23. The TetO-EGFRL858R;Rosa26LNL-tTA (E) and CCSP-rtTA;TetO-EGFRL858R;Rosa26CreER(T2) mice have been described in refs. 11,12,87,88. All mice were purified C57BL/6J mice, aged between 8 and 20 weeks, with a mixed sex ratio for each experiment (Supplementary Table 6). Tumors were initiated in E, EA3B, EP and EPA3B mice by intratracheal infection with adenoviral vectors expressing Cre recombinase as described89. Adenoviral-Cre (Ad-Cre-GFP) was from the University of Iowa Gene Transfer Core. Tumors were initiated in EA3Bi mice using chow containing doxycycline (625 ppm) obtained from Harlan-Teklad. All animal-regulated procedures were approved by the Francis Crick Institute BRF Strategic Oversight Committee that incorporates the Animal Welfare and Ethical Review Body and conformed with the UK Home Office guidelines and regulations under the Animals (Scientific Procedures) Act 1986 including Amendment Regulations 2012. To assess the recombination efficiency of the LSL allele upstream of APOBEC3B, PCR primers targeting the R26 site, the LSL cassette and the APOBEC3B transgene were used as described20. Erlotinib was purchased from Selleckchem (erlotinib, Osi-744), dissolved in 0.3% methylcellulose and administered intraperitoneally at 25 mg kg−1, 5 d a week. Tamoxifen was administered by oral gavage three times in 1 week at 2–4 d intervals (three injections total). Mice received tamoxifen at 150 mg kg−1 dissolved in sunflower oil.
Assessment of recombination efficiency
PCR was performed to assess the recombination of the LSL cassette upstream of the A3B allele in six tumors collected at progression. Five of six (5/6) of the tumors had a recombination efficiency above 90%, and one tumor of six was unrecombined. This rate of recombination aligns with the rate of recombination observed by IHC staining at 3 months and at termination and suggests that a lack of recombination of the LSL cassette upstream of the A3B transgene explains A3B-negative tumors.
Micro-computed tomography (micro-CT) imaging
Mice were anesthetized with isoflurane/oxygen for no more than an hour each and minimally restrained during imaging (~8 to 10 min). Mice were then observed and, if necessary, placed in cages in a recovery chamber/rack until they regained consciousness and started to feed. Tumor burden was quantified by calculating the volume of visible tumors using AnalyzeDirect.
Histological preparation and IHC staining
Tissues were fixed in 10% formalin overnight and transferred to 70% ethanol until paraffin embedding. IHC was performed using the following primary antibodies: EGFRL858R mutant specific (CST, 3197 and 43B2), APOBEC3B (5210-87-13)86, Ki67 (Abcam, Ab15580), Caspase 3 (R&D (Bio-Techne), AF835), p-Histone H2AX (Sigma-Aldrich, 05-636), Phospho-Histone H3 (Ser10; CST, 9706), CD4 (Abcam, ab183685; EPR19514), CD8 (Thermo Fisher Scientific, 14-0808-82; 4SM15) and UNG (Novus Biologicals, NB600-1031). Sections were developed with 3,3′-Diaminobenzidine (DAB) and counterstained with hematoxylin. Staining for p53 (Leica, NCL-L-p53-CM5p) was performed on a Dako Autostainer Link 48 (Agilent) as previously described90. The number of EGFRL858R, APOBEC3B, Ki67, Caspase 3 and gH2AX-positive cells were quantified using QuPath.
Evaluation of chromosome missegregation errors in hematoxylin and eosin (H&E)- and/or phospho-histone H3-stained samples
Lung sections were evaluated for anaphases with chromosome missegregation events using a ×100 objective light microscope. For E and EA3B mice at early and late timepoints, the percentage of missegregation errors was calculated and averaged across all mice using the harmonic mean. For EA3B mice, the percent error was normalized to an A3B recombination efficiency of 82% based on observed recombination efficiency observed (Extended Fig. 4). For E and EA3Bi mice with subclonal A3B expression, normalization for the recombination efficiency was not possible, so the percentage of missegregation errors was calculated based on the number of errors versus normal anaphases observed.
Mouse tumor processing
Frozen tumor tissue was cut into pieces and lysed in RLT Buffer with β-mercaptoethanol. TissueRuptor was used for disruption and homogenization of tissue. Lysate was added to a QIAshredder tube and centrifuged at full speed for 1 min. The homogenized solution was then added to AllPrep DNA spin columns (Qiagen AllPrep DNA/RNA Mini Kit, 80204).
Histopathological examination of mouse
Four micrometers thick, formalin-fixed, paraffin-embedded (FFPE) sections from lung lobes were stained with H&E and examined by two board-certified Veterinary Pathologists (A.S.B. and S.L.P.). Histopathological assessment was performed blind to experimental grouping using a light microscope (Olympus, BX43). Tissue sections were examined individually, and in case of discordance in diagnosis, a consensus was reached using a double-head microscope.
Proliferative lesions were diagnosed as alveolar hyperplasia, bronchioloalveolar adenoma and well-differentiated, moderately or poorly differentiated bronchioloalveolar adenocarcinoma. Sections were histopathologically assessed and graded for the presence and type of proliferative epithelial lung lesions using the International Harmonization of Nomenclature and Diagnostic Criteria for Lesions (INHAND) guide for nonproliferative and proliferative lesions of the respiratory tract of the mouse91.
WES—mouse data
WES was performed by the Advanced Sequencing Facility at the Francis Crick Institute using the Human Core Exome Kit (Twist BioScience) for library preparation and SureSelectXT Mouse All Exon, 16, Kit (Agilent) for library preparation, respectively. Sequencing was performed on HiSeq 4000 platforms.
RNA-seq—mouse data
RNA-seq was performed by the Advanced Sequencing Facility at the Francis Crick Institute using the KAPA mRNA HyperPrep Kit (KK8581—96 Libraries) and KAPA Dual-Indexed Adapters (Roche, KK8720). Sequencing was performed on HiSeq 4000 platforms. The processed FASTQ files were mapped to mm10 reference genome using the STAR (version 2.4) algorithm, and transcript expressions were quantified using the RSEM (version 1.2.29) algorithm with the default parameters. The read counts were used for downstream analysis.
Alignment—mouse
All samples were demultiplexed, and the resultant FASTQ files aligned to the mm10 mouse genome, using BWA-MEM (BWA, v0.7.15). Deduplication was performed using Picard (v2.1.1; http://broadinstitute.github.io/picard). Quality control metrics were collated using FASTQC (v0.10.1; http://www.bioinformatics.babraham.ac.uk/projects/fastqc/), Picard and GATK (v3.6). SAMtools (v1.3.1) was used to generate mpileup files from the resultant BAM files. Thresholds for base phred score and mapping quality were set at 20. A threshold of 50 was set for the coefficient of downgrading mapping quality, with the argument for base alignment quality calculation being deactivated. The median depth of coverage for all samples was 92× (range: 58–169×).
Variant detection and annotation—mouse
Variant calling was performed using VarScan2 (v2.4.1), MuTect (v1.1.7) and Scalpel (v0.5.4)92,93,94.
The following argument settings were used for variant detection using VarScan2:
--min-coverage 8 --min-coverage-normal 10 --min-coverage-tumor 6 --min-var-freq 0.01 --min-freq-for-hom 0.75 --normal-purity 1 --p-value 0.99 --somatic-p-value 0.05 --tumor-purity 0.5 --strand-filter 0
For MuTect, only ‘PASS’ variants were used for further analyses. Except for allowing variants to be detected down to a variant allele frequency (VAF) of 0.001, default settings were used for Scalpel insertion/deletion detection.
To minimize false positives, additional filtering was performed. For single-nucleotide variants (SNVs) or dinucleotides detected by VarScan2, a minimum tumor sequencing depth of 30, VAF of 5%, variant read count of 5 and a somatic P value < 0.01 were required to pass a variant. For variants detected by VarScan2 between 2% and 5% VAF, the mutation also needs to be detected by MuTect.
As for insertions/deletions (INDELs), variants need to be passed by both Scalpel (PASS) and VarScan2 (somatic P < 0.001). A minimum depth of 50×, 10 alt reads and VAF of 2% were required.
For all SNVs, INDELs and dinucleotides, any variant also detected in the paired germline sample with more than five alternative reads or a VAF greater than 1% was filtered out.
The detected variants were annotated using Annovar95.
Functional annotation of SNVs—mouse
Mouse gene mutation callings from WES were parsed with some modifications including genomic coordinates (removing ‘chr’ before chromosomal numbers, only ‘SNV’ was selected). The modified files were fed into Protein Variation Effect Analyzer (PROVEAN)96,97,98 software tool (http://provean.jcvi.org/index.php) to predict whether an amino acid substitution has an impact on the biological function of a protein (Sorting Intolerant From Tolerant, SIFT score). The predict files were merged with original files at gene level annotation using the R program.
Human EGFR transgene amplicon sequencing of mouse
FASTQ files were aligned to hg19 obtained from the GATK bundle (v2.8) using BWA-MEM (BWA, v0.7.15)99,100. Analyses were performed using R (v3.3.1) and deepSNV (v1.18.1)101. The median depth of coverage of sequenced EGFR exons (19,20,21) was 5290× (range: 2,238–8,040). Variants associated with resistance to EGFR TKIs were queried using deepSNV’s bam2R function, with the arguments q = 20 and s = 2. The variants explored include the following: T790M, D761Y, L861Q, G796X, G797X, L792X and L747S. L858R was identified in every sequenced sample.
Generation of EGFRL858R mutant mouse tumor cell lines
A portion of mouse lung tumor was dissected (1/3 to 1/2 of the original tumor depending on size) and cut into small pieces with scissors. Pieces were then digested for 30 min at 37 °C while rotating at full speed in digestion media (1,400 µl HBSS-free w/o Ca2+, 200 µl Collagenase IV and 40 U ml−1 DNase). Tumor cells were pelleted down in a centrifuge (1,100 r.p.m. for 4 min) and resuspended in IMDM supplemented with 1% penicillin–streptomycin solution (10,000 U ml−1), l-glutamine (200 mM) and 10% FBS. This cell suspension was then plated in a 10-cm plate and passaged over a period of 1–3 months until consistent growth was observed.
Generation of TKI-resistant mouse or human tumor cell lines
TKI naïve cell lines were cultured in increasing levels of erlotinib or osimertinib using a dose-escalation protocol from 100 nM to 1 µM when cells were growing with minimal cell death.
Mutational and SCNA ITH calculations for TRACERx data
SCNA ITH was calculated by dividing the percentage of the genome harboring heterogeneous SCNA events, that is, those events that were not present in every region, by the percentage of the genome involved in any SCNA event in each tumor25.
Cell line whole-genome mutational signature analysis
Sequences were aligned to the human genome (hg38) using the Burrows-Wheeler Aligner (version 0.7.17). PCR duplicates were removed using Picard (version 2.18.16). Reads were locally realigned around indels using GATK3 (version 3.6.0) tools RealignerTargetCreator to create intervals, followed by IndelRealigner on the aligned BAM files. MuTect2 from GATK3 (version 3.6.0) was used in tumor/normal mode to call mutations in test versus control cell lines. SNVs that passed the internal GATK3 filter with read depths over 30 reads at called positions, at least 4 reads in the alternate mutation call and an allele frequency greater than 0.05 were used for downstream analysis. Mutational profile plots in Fig. 6g were plotted using the deconstructSigs R package102.
DNA and RNA isolation from cell line models for sequencing
DNA or RNA were extracted from frozen cell pellets using Qiagen’s DNeasy Blood and Tissue Kit or Qiagen’s RNeasy MINI Kit, respectively, as per the manufacturer’s instructions. The isolated DNA or RNA was quantified and qualitatively assessed using a Qubit Fluorometer (Thermo Fisher Scientific) and a Bioanalyzer (Agilent), as per the manufacturer’s instructions. The DNA or RNA were then sent to BGI for WGS (30×) or Novogene for mRNA or WES.
Cell cycle analysis
Cell cycle analysis was performed by propidium iodide (PI) staining. Briefly, PC9 cells were treated for 24 h with DMSO, 2 µM osimertinib or 1 µM palbociclib and then fixed in ice-cold 70% ethanol and stained with a 50 µg ml−1 PI (MilliporeSigma, P4864) + 0.1% Triton X-100 (MilliporeSigma, X100) solution. PI fluorescence was then measured on a flow cytometer (BD FACSAria II).
Human participants
All patients gave informed written consent for the collection of clinical correlates, tissue collection and research testing under institutional review board (IRB)-approved protocols (CC13-6512 and CC17-658, NCT03433469). Patient demographics are listed in Supplementary Tables 2a–c, 4, and 5a,b. Patient studies were conducted according to the Declaration of Helsinki, the Belmont Report and the U.S. Common Rule.
Studies with specimens from patients with lung cancer
Frozen or FFPE tissues from patients with lung cancer for DNA or RNA sequencing (bulk and single cell) studies were processed and sequenced as described previously41,60. Classification of response was based on RECIST criteria. Some of these biopsies were subjected to WES at the QB3-Berkley Genomics for which library preparation was performed using IDT’s xGen exome panel. For additional specimens, tumor DNA from FFPE tissues and matched nontumor from blood aliquots or stored buffy coats were collected as part of the UCSF biospecimen resource program (BIOS) in accordance with UCSF’s IRB-approved protocol. DNA from blood aliquots was isolated at the BIOS. Other nontumor samples and FFPE tumor tissues were sent for extraction and assessment of quality and quantity to Novogene, and those meeting the required sample standards were subjected to WES at Novogene’s sequencing facility.
Mutation analysis
Paired-end reads were aligned to the hg19 human genome using the Picard pipeline (https://gatk.broadinstitute.org/). A modified version of the Broad Institute Getz Lab CGA WES Characterization pipeline (https://docs-google-com.ezp-prod1.hul.harvard.edu/document/d/1VO2kX_fgfUd0x3mBS9NjLUWGZu794WbTepBel3cBg08) was used to call, filter and annotate somatic mutations. Specifically, SNVs and other substitutions were called with MuTect (v1.1.6)93. Mutations were annotated using Oncotator103. MuTect mutation calls were filtered for 8-OxoG artifacts, and artifacts were introduced through the formalin fixation process (FFPE) of tumor tissues66. Indels were called with Strelka (v1.0.11). MuTect calls and Strelka calls were further filtered through a panel of normal samples (PoN) to remove artifacts generated by rare error modes and miscalled germline alterations93. To pass quality control, samples were required to have <5% cross-sample contamination as assessed with ContEst93; mean target coverage of at least 25× in the tumor sample and 20× in the corresponding normal as assessed using GATK3.7 DepthOfCoverage and a percentage of tumor-in-normal of <30% as determined by deTiN104. This pipeline was modified for analysis of cell lines rather than tumor-normal pairs as follows: indels were called through MuTect2 alone rather than Strelka; deTiN was not performed and a common variant filter was applied to exclude variants present in the Exome Aggregation Consortium if at least ten alleles containing the variant were present across any subpopulation, unless they appeared in a list of known somatic sites105,106.
Mutational signature analysis
Active mutational processes107 were determined using the deconstructSigs R package63, with a signature contribution cutoff of 6%. This cutoff was chosen because it was the minimum contribution value required to obtain a false-positive rate of 0.1% and a false-negative rate of 1.4% per the authors’ in silico analysis and is the recommended cutoff102. Samples with <10 mutations were excluded from analysis due to poor signature discrimination with only a few mutations, and a sample with less than 15 d of exposure to TKI therapy was excluded because it is too short a time to accumulate detectable mutations due to therapy. For TRACERx data analysis, data processing was performed in the R statistical environment version ≥3.3.1.
RNA-seq analyses
PDX tissue and mouse tumor cell line RNA extractions were carried out using an RNeasy Micro Kit (Qiagen). RNA-seq was performed on PDX tissue using replicate samples on the Illumina HiSeq 4000, paired-end 100-bp reads at the Center for Advanced Technology (UCSF). For the differential gene expression analysis, DESeq program was used to compare controls to erlotinib samples as previously described108.
RNA-seq samples from patients and cell lines were sequenced by Novogene (https://en.novogene.com/) with paired-end sequencing (150 bp in length). There were ~20 million reads for each sample. The processed FASTQ files were mapped to the hg19 reference genome using the STAR (version 2.4) algorithm, and transcript expressions were quantified using the RSEM (version 1.2.29) algorithm. The default parameters in the algorithms were used. The normalized transcript reads (TPM) were used for downstream analysis. Gene set enrichment analysis was performed using GSEA software109.
For single-cell RNA-seq analyses, the data from a previously published study (all cancer cells from patients with advanced lung cancer) were used and analyzed in a similar manner41. All cells used are identified as malignant by marker expression and CNV inference and originated in from various biopsy sites (adrenal, liver, lymph node, lung and pleura/pleural fluid). Nonparametric, pairwise comparisons (Wilcoxon rank-sum test) were used to determine the statistical significance of the pairwise comparisons of different timepoints for their average scaled expression.
Statistical analysis
One-way or two-way ANOVA test with Holm–Sidak correction for multiple comparisons (>2 groups) or two-tailed t test (2 groups) were used to determine the statistical significance of the differences between groups for RT–qPCR, growth and enzymatic assays and bulk RNA-seq analysis. Normality of IHC and micro-CT data was determined using multiple testing methods (Anderson–Darling test, D’Agostino–Pearson test, Shapiro–Wilk test and Kolmogorov–Smirnov test). A two-sided t test or two-sided Mann–Whitney test was used for IHC and micro-CT data depending on the normality tests to determine the statistical significance of the differences between groups. Analysis for these assays was done using GraphPad Prism.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The WES data and RNA-seq data (from the TRACERx study) used during this study have been deposited at the European Genome-phenome Archive (EGA), which is hosted by the European Bioinformatics Institute and the Center for Genomic Regulation under the accession codes EGAS00001006494 and EGAS00001006517, respectively, is under controlled access due to its nature and commercial licenses. Specifically, data are available through the Cancer Research UK and University College London Cancer Trials Center (ctc.tracerx@ucl.ac.uk) for academic noncommercial research purposes only and are subject to review of a project proposal by the TRACERx data access committee, entering into an appropriate data access agreement and subject to any applicable ethical approvals. A response to the request for access is typically provided within ten working days after the committee has received the relevant project proposal and all other required information.
The WES data of tumor-derived cell lines shown in Extended Data Fig. 3 are available at the European Nucleotide Archive (ENA) with the identifier PRJEB67640 (ERP152649). The WGS data of PC9 cell lines shown in Fig. 6 are available at the ENA with the identifier PRJEB67559 (ERP152586). For the single-cell RNA-seq analyses shown in Extended Data Fig. 10b,c, the data from a previously published study (all advanced lung cancer cell data) were used and analyzed in a similar manner41. These data are available in the National Center for Biotechnology Information (NCBI) BioProject ID PRJNA591860. The RNA-seq data for Extended Data Fig. 10a were from a previously published study38. These data are available at NCBI GEO under accession GSE65420. Clinical sample RNA-seq and WES sequencing data are available in NCBI BioProject ID PRJNA1029563. Source data are provided with this paper.
References
Cheng, A. Z. et al. APOBECs and herpesviruses. Viruses 13, 390 (2021).
Harris, R. S. & Liddament, M. T. Retroviral restriction by APOBEC proteins. Nat. Rev. Immunol. 4, 868–877 (2004).
Swanton, C., McGranahan, N., Starrett, G. J. & Harris, R. S. APOBEC enzymes: mutagenic fuel for cancer evolution and heterogeneity. Cancer Discov. 5, 704–712 (2015).
Venkatesan, S. et al. Induction of APOBEC3 exacerbates DNA replication stress and chromosomal instability in early breast and lung cancer evolution. Cancer Discov. 11, 2456–2473 (2021).
Isozaki, H. et al. Therapy-induced APOBEC3A drives evolution of persistent cancer cells. Nature 620, 393–401 (2023).
Jarvis, M. C. et al. Mutational impact of APOBEC3B and APOBEC3A in a human cell line. Preprint at bioRxiv 10.1101/2022.04.26.489523 (2022).
de Bruin, E. C. et al. Spatial and temporal diversity in genomic instability processes defines lung cancer evolution. Science 346, 251–256 (2014).
Chen, J. et al. Genomic landscape of lung adenocarcinoma in East Asians. Nat. Genet. 52, 177–186 (2020).
Jamal-Hanjani, M. et al. Tracking the evolution of non-small-cell lung cancer. N. Engl. J. Med. 376, 2109–2121 (2017).
Henderson, S., Chakravarthy, A., Su, X., Boshoff, C. & Fenton, T. R. APOBEC-mediated cytosine deamination links PIK3CA helical domain mutations to human papillomavirus-driven tumor development. Cell Rep. 7, 1833–1841 (2014).
Politi, K. et al. Lung adenocarcinomas induced in mice by mutant EGF receptors found in human lung cancers respond to a tyrosine kinase inhibitor or to down-regulation of the receptors. Genes Dev. 20, 1496–1510 (2006).
Politi, K., Fan, P.-D., Shen, R., Zakowski, M. & Varmus, H. Erlotinib resistance in mouse models of epidermal growth factor receptor-induced lung adenocarcinoma. Dis. Model. Mech. 3, 111–119 (2010).
Starrett, J. H. et al. Drug sensitivity and allele specificity of first-line osimertinib resistance EGFR mutations. Cancer Res. 80, 2017–2030 (2020).
De Bruin, E. C., McGranahan, N. & Swanton, C. Analysis of intratumor heterogeneity unravels lung cancer evolution. Mol. Cell. Oncol. 2, e985549 (2015).
Westcott, P. M. K. et al. The mutational landscapes of genetic and chemical models of Kras-driven lung cancer. Nature 517, 489–492 (2015).
McFadden, D. G. et al. Mutational landscape of EGFR-, MYC-, and Kras-driven genetically engineered mouse models of lung adenocarcinoma. Proc. Natl Acad. Sci. USA 113, E6409–E6417 (2016).
Chung, W.-J. et al. Kras mutant genetically engineered mouse models of human cancers are genomically heterogeneous. Proc. Natl Acad. Sci. USA 114, E10947–E10955 (2017).
Jónsson, S. R. et al. Evolutionarily conserved and non-conserved retrovirus restriction activities of artiodactyl APOBEC3F proteins. Nucleic Acids Res. 34, 5683–5694 (2006).
MacMillan, A. L., Kohli, R. M. & Ross, S. R. APOBEC3 inhibition of mouse mammary tumor virus infection: the role of cytidine deamination versus inhibition of reverse transcription. J. Virol. 87, 4808–4817 (2013).
Boumelha, J. et al. An immunogenic model of KRAS-mutant lung cancer for study of targeted therapy and immunotherapy combinations. Cancer Res. 82, 3435–3448 (2022).
Ji, H. et al. The impact of human EGFR kinase domain mutations on lung tumorigenesis and in vivo sensitivity to EGFR-targeted therapies. Cancer Cell 9, 485–495 (2006).
McCann, J. L. et al. The DNA deaminase APOBEC3B interacts with the cell-cycle protein CDK4 and disrupts CDK4-mediated nuclear import of cyclin D1. J. Biol. Chem. 294, 12099–12111 (2019).
Durfee, C. et al. APOBEC3B promotes tumor development in vivo including signature mutations and metastases. Cell. Rep. Med. 4, 101211 (2023).
DiMarco, A. V. et al. APOBEC mutagenesis inhibits breast cancer growth through induction of a T cell mediated antitumor immune response. Cancer Immunol. Res. 10, 70–86 (2022).
Frankell, A. M. et al. The evolution of lung cancer and impact of subclonal selection in TRACERx. Nature 616, 525–533 (2023).
Carter, S. L., Eklund, A. C., Kohane, I. S., Harris, L. N. & Szallasi, Z. A signature of chromosomal instability inferred from gene expression profiles predicts clinical outcome in multiple human cancers. Nat. Genet. 38, 1043–1048 (2006).
Seibler, J. Rapid generation of inducible mouse mutants. Nucleic Acids Res. 31, e12 (2003).
Serebrenik, A. A. et al. The deaminase APOBEC3B triggers the death of cells lacking uracil DNA glycosylase. Proc. Natl Acad. Sci. USA 116, 22158–22163 (2019).
Veeriah, S. et al. The tyrosine phosphatase PTPRD is a tumor suppressor that is frequently inactivated and mutated in glioblastoma and other human cancers. Proc. Natl Acad. Sci. USA 106, 9435–9440 (2009).
Ortiz, B. et al. Loss of the tyrosine phosphatase PTPRD leads to aberrant STAT3 activation and promotes gliomagenesis. Proc. Natl Acad. Sci. USA 111, 8149–8154 (2014).
Morris, L. G. et al. Genomic dissection of the epidermal growth factor receptor (EGFR)/PI3K pathway reveals frequent deletion of the EGFR phosphatase PTPRS in head and neck cancers. Proc. Natl Acad. Sci. USA 108, 19024–19029 (2011).
Petljak, M. et al. Characterizing mutational signatures in human cancer cell lines reveals episodic APOBEC mutagenesis. Cell 176, 1282–1294 (2019).
Law, E. K. et al. The DNA cytosine deaminase APOBEC3B promotes tamoxifen resistance in ER-positive breast cancer. Sci. Adv. 2, e1601737 (2016).
Haderk, F. et al. A focal adhesion kinase-YAP signaling axis drives drug tolerant persister cells and residual disease in lung cancer. Preprint at bioRxiv https://doi.org/10.1101/2021.10.23.465573 (2021).
Vieira, V. C. et al. Human papillomavirus E6 triggers upregulation of the antiviral and cancer genomic DNA deaminase APOBEC3B. mBio 5, e02234-14 (2014).
Petljak, M. et al. Mechanisms of APOBEC3 mutagenesis in human cancer cells. Nature 607, 799–807 (2022).
Bollard, J. et al. Palbociclib (PD-0332991), a selective CDK4/6 inhibitor, restricts tumour growth in preclinical models of hepatocellular carcinoma. Gut 66, 1286–1296 (2017).
Blakely, C. M. et al. NF-κB-activating complex engaged in response to EGFR oncogene inhibition drives tumor cell survival and residual disease in lung cancer. Cell Rep. 11, 98–110 (2015).
Leonard, B. et al. The PKC/NF-κB signaling pathway induces APOBEC3B expression in multiple human cancers. Cancer Res. 75, 4538–4547 (2015).
Maruyama, W. et al. Classical NF-κB pathway is responsible for APOBEC3B expression in cancer cells. Biochem. Biophys. Res. Commun. 478, 1466–1471 (2016).
Maynard, A. et al. Therapy-induced evolution of human lung cancer revealed by single-cell RNA sequencing. Cell 182, 1232–1251 (2020).
Gilmore, T. D. Introduction to NF-κB: players, pathways, perspectives. Oncogene 25, 6680–6684 (2006).
Farré, D. et al. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 31, 3651–3653 (2003).
Hollenhorst, P. C. RAS/ERK pathway transcriptional regulation through ETS/AP-1 binding sites. Small GTPases 3, 154–158 (2012).
Chan, K. et al. An APOBEC3A hypermutation signature is distinguishable from the signature of background mutagenesis by APOBEC3B in human cancers. Nat. Genet. 47, 1067–1072 (2015).
Sun, H. T., Cheng, S. X., Tu, Y., Li, X. H. & Zhang, S. FoxQ1 promotes glioma cells proliferation and migration by regulating NRXN3 expression. PLoS ONE 8, e55693 (2013).
Kusinska, R. et al. Influence of genomic variation in FTO at 16q12.2, MC4R at 18q22 and NRXN3 at 14q31 genes on breast cancer risk. Mol. Biol. Rep. 39, 2915–2919 (2012).
Jacobsen, K. et al. Convergent Akt activation drives acquired EGFR inhibitor resistance in lung cancer. Nat. Commun. 8, 410 (2017).
Jia, P. et al. Next-generation sequencing of paired tyrosine kinase inhibitor-sensitive and -resistant EGFR mutant lung cancer cell lines identifies spectrum of DNA changes associated with drug resistance. Genome Res. 23, 1434–1445 (2013).
Ramirez, M. et al. Diverse drug-resistance mechanisms can emerge from drug-tolerant cancer persister cells. Nat. Commun. 7, 10690 (2016).
Roper, N. et al. Clonal evolution and heterogeneity of osimertinib acquired resistance mechanisms in EGFR mutant lung cancer. Cell Rep. Med. 1, 100007 (2020).
Roper, N. et al. APOBEC mutagenesis and copy-number alterations are drivers of proteogenomic tumor evolution and heterogeneity in metastatic thoracic tumors. Cell Rep. 26, 2651–2666 (2019).
Selenica, P. et al. APOBEC mutagenesis, kataegis, chromothripsis in EGFR-mutant osimertinib-resistant lung adenocarcinomas. Ann. Oncol. 33, 1284–1295 (2022).
Samuels, Y. et al. Mutant PIK3CA promotes cell growth and invasion of human cancer cells. Cancer Cell 7, 561–573 (2005).
Kim, S. & Jeong, S. Mutation hotspots in the β-catenin gene: lessons from the human cancer genome databases. Mol. Cells 42, 8–16 (2019).
Ruvolo, P. P. The broken ‘off’ switch in cancer signaling: PP2A as a regulator of tumorigenesis, drug resistance, and immune surveillance. BBA Clin. 6, 87–99 (2016).
Silverstein, A. M., Barrow, C. A., Davis, A. J. & Mumby, M. C. Actions of PP2A on the MAP kinase pathway and apoptosis are mediated by distinct regulatory subunits. Proc. Natl Acad. Sci. USA 99, 4221–4226 (2002).
Yao, Y. et al. Mutations in the MET tyrosine kinase domain and resistance to tyrosine kinase inhibitors in non-small-cell lung cancer. Respir. Res. 24, 28 (2023).
Gainor, J. F. et al. Molecular mechanisms of resistance to first- and second-generation ALK inhibitors in ALK-rearranged lung cancer. Cancer Discov. 6, 1118–1133 (2016).
Blakely, C. M. et al. Evolution and clinical impact of co-occurring genetic alterations in advanced-stage EGFR-mutant lung cancers. Nat. Genet. 49, 1693–1704 (2017).
Shen, S. et al. Upregulation of miR-136 in human non-small cell lung cancer cells promotes Erk1/2 activation by targeting PPP2R2A. Tumour Biol. 35, 631–640 (2014).
Engelman, J. A. et al. Effective use of PI3K and MEK inhibitors to treat mutant Kras G12D and PIK3CA H1047R murine lung cancers. Nat. Med. 14, 1351–1356 (2008).
Ercan, D. et al. Reactivation of ERK signaling causes resistance to EGFR kinase inhibitors. Cancer Discov. 2, 934–947 (2012).
Hrustanovic, G. et al. RAS-MAPK dependence underlies a rational polytherapy strategy in EML4-ALK-positive lung cancer. Nat. Med. 21, 1038–1047 (2015).
Papadakis, A. I. et al. SMARCE1 suppresses EGFR expression and controls responses to MET and ALK inhibitors in lung cancer. Cell Res. 25, 445–458 (2015).
Yu, H. A. et al. Analysis of tumor specimens at the time of acquired resistance to EGFR-TKI therapy in 155 patients with EGFR-mutant lung cancers. Clin. Cancer Res. 19, 2240–2247 (2013).
Sequist, L. V. et al. Genotypic and histological evolution of lung cancers acquiring resistance to EGFR inhibitors. Sci. Transl. Med. 3, 75ra26 (2011).
Nik-Zainal, S. et al. Mutational processes molding the genomes of 21 breast cancers. Cell 149, 979–993 (2012).
Huang, L. & Fu, L. Mechanisms of resistance to EGFR tyrosine kinase inhibitors. Acta Pharm. Sin. B 5, 390–401 (2015).
Nagano, T., Tachihara, M. & Nishimura, Y. Mechanism of resistance to epidermal growth factor receptor-tyrosine kinase inhibitors and a potential treatment strategy. Cells 7, 212 (2018).
Leonetti, A. et al. Resistance mechanisms to osimertinib in EGFR-mutated non-small cell lung cancer. Br. J. Cancer 121, 725–737 (2019).
Noronha, A. et al. AXL and error-prone DNA replication confer drug resistance and offer strategies to treat EGFR-mutant lung cancer. Cancer Discov. 12, 2666–2683 (2022).
Sieuwerts, A. M. et al. Elevated APOBEC3B correlates with poor outcomes for estrogen-receptor-positive breast cancers. Horm. Cancer 5, 405–413 (2014).
Leshchiner, I. et al. Comprehensive analysis of tumour initiation, spatial and temporal progression under multiple lines of treatment. Preprint at bioRxiv 10.1101/508127 (2019).
Lee, J.-K. et al. Clonal history and genetic predictors of transformation into small-cell carcinomas from lung adenocarcinomas. J.Clin. Oncol. 35, 3065–3074 (2017).
Garcia, N. M. G. et al. APOBEC3 activity promotes the survival and evolution of drug-tolerant persister cells during acquired resistance to EGFR inhibitors in lung cancer. Preprint at bioRxiv 10.1101/2023.07.02.547443 (2023).
Maciejowski, J. et al. APOBEC3-dependent kataegis and TREX1-driven chromothripsis during telomere crisis. Nat. Genet. 52, 884–890 (2020).
Nikkila, J. et al. Elevated APOBEC3B expression drives a kataegic-like mutation signature and replication stress-related therapeutic vulnerabilities in p53-defective cells. Br. J. Cancer 117, 113–123 (2017).
Buisson, R., Lawrence, M. S., Benes, C. H. & Zou, L. APOBEC3A and APOBEC3B activities render cancer cells susceptible to ATR inhibition. Cancer Res. 77, 4567–4578 (2017).
Russo, M. et al. Adaptive mutability of colorectal cancers in response to targeted therapies. Science 366, 1473–1480 (2019).
Ma, W. et al. APOBEC3B promotes hepatocarcinogenesis and metastasis through novel deaminase-independent activity. Mol. Carcinog. 58, 643–653 (2019).
Xie, S., Cooley, A., Armendariz, D., Zhou, P. & Hon, G. C. Frequent sgRNA-barcode recombination in single-cell perturbation assays. PLoS ONE 13, e0198635 (2018).
Sanjana, N. E., Shalem, O. & Zhang, F. Improved vectors and genome-wide libraries for CRISPR screening. Nat. Methods 11, 783–784 (2014).
Burns, M. B. et al. APOBEC3B is an enzymatic source of mutation in breast cancer. Nature 494, 366–370 (2013).
Suzuki, K., Bose, P., Leong-Quong, R. Y., Fujita, D. J. & Riabowol, K. REAP: a two minute cell fractionation method. BMC Res. Notes 3, 294 (2010).
Brown, W. L. et al. A rabbit monoclonal antibody against the antiviral and cancer genomic DNA mutating enzyme APOBEC3B. Antibodies (Basel) 8, 47 (2019).
Murillo, M. M. et al. Disruption of the interaction of RAS with PI 3-kinase induces regression of EGFR-mutant-driven lung cancer. Cell Rep. 25, 3545–3553 (2018).
Tichelaar, J. W., Lu, W. & Whitsett, J. A. Conditional expression of fibroblast growth factor-7 in the developing and mature lung. J. Biol. Chem. 275, 11858–11864 (2000).
Dupage, M., Dooley, A. L. & Jacks, T. Conditional mouse lung cancer models using adenoviral or lentiviral delivery of Cre recombinase. Nat. Protoc. 4, 1064–1072 (2009).
Humpton, T. J. et al. A noninvasive iRFP713 p53 reporter reveals dynamic p53 activity in response to irradiation and liver regeneration in vivo. Sci. Signal 15, eabd9099 (2022).
Renne, R. et al. Proliferative and nonproliferative lesions of the rat and mouse respiratory tract. Toxicol. Pathol. 37, 5S–73S (2009).
Koboldt, D. C. et al. VarScan 2: somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 22, 568–576 (2012).
Cibulskis, K. et al. Sensitive detection of somatic point mutations in impure and heterogeneous cancer samples. Nat. Biotechnol. 31, 213–219 (2013).
Fang, H. et al. Indel variant analysis of short-read sequencing data with Scalpel. Nat. Protoc. 11, 2529–2548 (2016).
Wang, K., Li, M. & Hakonarson, H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 38, e164 (2010).
Choi, Y., Sims, G. E., Murphy, S., Miller, J. R. & Chan, A. P. Predicting the functional effect of amino acid substitutions and indels. PLoS ONE 7, e46688 (2012).
Choi, Y. Fast computation of pairwise sequence alignment scores between a protein and a set of single-locus variants of another protein. Proceedings of the ACM Conference on Bioinformatics, Computational Biology and Biomedicine (BCB' 12) pp. 414–417 (Association for Computing Machinery, 2012).
Choi, Y. & Chan, A. P. PROVEAN web server: a tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 31, 2745–2747 (2015).
McKenna, A. et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303 (2010).
Li, H. et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Gerstung, M. et al. Reliable detection of subclonal single-nucleotide variants in tumour cell populations. Nat. Commun. 3, 811 (2012).
Rosenthal, R., McGranahan, N., Herrero, J., Taylor, B. S. & Swanton, C. DeconstructSigs: delineating mutational processes in single tumors distinguishes DNA repair deficiencies and patterns of carcinoma evolution. Genome Biol. 17, 31–11 (2016).
Ramos, A. H. et al. Oncotator: cancer variant annotation tool. Hum. Mutat. 36, E2423–E2429 (2015).
Taylor-Weiner, A. et al. DeTiN: overcoming tumor-in-normal contamination. Nat. Methods 15, 531–534 (2018).
Lek, M. et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 536, 285–291 (2016).
AACR Project GENIE Consortium AACR project GENIE: powering precision medicine through an international consortium. Cancer Discov. 7, 818–831 (2017).
Alexandrov, L. B. et al. Signatures of mutational processes in human cancer. Nature 500, 415–421 (2013).
Anders, S. & Huber, W. Differential expression analysis for sequence count data. Genome Biol. 11, R106–R112 (2010).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Acknowledgements
C.S. is a Royal Society Napier Research Professor (RSRP\R\210001). His work is supported by the Francis Crick Institute which receives its core funding from Cancer Research UK (CRUK) (CC2041), the UK Medical Research Council (CC2041) and the Wellcome Trust (CC2041). For Open Access, the author has applied a CC BY public copyright license to any Author Accepted Manuscript version arising from this submission. C.S. is funded by Cancer Research UK (TRACERx (C11496/A17786), PEACE (C416/A21999) and CRUK Cancer Immunotherapy Catalyst Network); Cancer Research UK Lung Cancer Center of Excellence (C11496/A30025); the Rosetrees Trust, Butterfield and Stoneygate Trusts; Novo Nordisk Foundation (ID16584); Royal Society Professorship Enhancement Award (RP/EA/180007); National Institute for Health Research (NIHR) University College London Hospitals Biomedical Research Center; the Cancer Research UK-University College London Center; Experimental Cancer Medicine Center; the Breast Cancer Research Foundation (United States, BCRF-22-157); Cancer Research UK Early Detection and Diagnosis Primer Award (grant EDDPMA-Nov21/100034) and the Mark Foundation for Cancer Research Aspire Award (grant 21-029-ASP). This work was supported by a Stand Up To Cancer‐LUNGevity-American Lung Association Lung Cancer Interception Dream Team Translational Research Grant (grants SU2C-AACR-DT23-17 to S.M. Dubinett and A.E. Spira). Stand Up To Cancer is a division of the Entertainment Industry Foundation. Research grants are administered by the American Association for Cancer Research, the Scientific Partner of SU2C. C.S. is in receipt of an ERC Advanced Grant (PROTEUS) from the European Research Council under the European Union’s Horizon 2020 research and innovation program (grant 835297). This project is supported by the NIH/NCI U54CA224081, R01CA169338, R01CA211052, R01CA204302, U01CA217882 and the Chan-Zuckerberg Biohub (to T.G.B.), Pfizer, as well as the University of California Cancer League (to C.E.M.), AstraZeneca, the Damon Runyon Cancer Research Foundation P0528804, Doris Duke Charitable Foundation P2018110, V Foundation P0530519 and NIH/NCI R01CA227807 (to C.M.B.). F.H. was supported by the Mildred Scheel postdoctoral fellowship from the German Cancer Aid. E.A.Y. is supported by T32 HL007185 from the NHLBI. Cancer studies in the Harris Lab are supported in part by the National Cancer Institute (P01-CA234228). R.S.H. is the Ewing Halsell President’s Council Distinguished Chair at the University of Texas San Antonio and an Investigator of the Howard Hughes Medical Institute. D.R.C. was supported by the Francis Crick Institute receives its core funding from Cancer Research UK (FC001169), the UK Medical Research Council (FC002269) and the Wellcome Trust (FC001169), as well as an NC3Rs training fellowship (NC/S001832/1). J.S.R.-F. is funded in part by the Breast Cancer Research Foundation, by a Susan G. Komen Leadership grant and by the NIH/NCI (grant P50 CA247749 01). H.Y. is funded in part by NIH/NCI (grant P50 CAS247749 01) and 1R01CA264078-01. M.J.-H. has received funding from CRUK, NIH National Cancer Institute, International Association for the Study of Lung Cancer (IASLC) International Lung Cancer Foundation, Lung Cancer Research Foundation, Rosetrees Trust, UK and Ireland Neuroendocrine Tumour Society (UKI NETs) and NIHR. Special thanks to the Biological Research Facility at the Francis Crick Institute, specifically to A. Adekoya, J. Cormack, A. Horwood and S. Lighterness for their hard work and support. Special thanks also to the Experimental Histopathology Laboratory at the Francis Crick Institute, specifically to E. Nye, B. Almeida, M. Green and R. Stone for their help and support. Special thanks to all the members of the Bivona Laboratory (former and current), D. Gordenin, A. Sweet-Cordero, S. Bandyopadhyay, M. Breese, S. Kaushik, B. Leonard, S. Raju and K. Descamp for their insights and support and S. Elmes, A. Maynard, D.V. Allegakoen and A. Tambe for their technical support.
Author information
Authors and Affiliations
Contributions
D.R.C., P.G., M.K.M., E.K.L., N.K., R.S.H., J.D., T.G.B. and C.S. conceived and designed the study. D.R.C., P.G., M.K.M., J.B., J.D.D.P.V., F.H., B.G., T.M., W.T., T.A., P.A., S.N., C.G., E.G., M.A.B., A.N., F.G.V., W.H., W.T.L., B.A., M.G., C.M., J.P., E.G., C.Z., S.L., J.C., B.R., W.B., A.R., B.A., R.I.V., M.M., N.J.T., T.J.H., C.E.W., N.K., S.V., K.V., S.H., V.O., D.B., M.T., S.D.C.T., R.V., V.B., X.Z. and Y.J. conducted data acquisition for cell line and animal studies. C.M.R., M.D., M.A., C.B., O.P., B.B., C.E.M., J.R.M.B., C.M.B., D.L.K., J.K.R., A.M., J.R.F., P.S., H.Y., M.J.H., P.A., E.A.Y. and L.T. performed data acquisition for clinical studies. C.B., O.P., B.B., M.D., M.A., N.I.V., N.A.T., W.W., L.C., E.M.V.A., J.Y. and J.B. conducted mutational signature analysis and/or other computational analyses. D.R.C, P.G., M.K.M., N.I.V., T.G.B., C.S., E.K.L, R.S.H., W.L.B., L.K.L., C.D., P.P.A., J.P., T.M., M.A.B., A.N., M.D., C.M.R., S.F.B., S.K.C., S.L.P., A.S.B., N.M., C.M., B.R., B.B., W.W., K.H.V., D.L.K., F.H., C.B., O.P., B.B., N.K., N.A.T., U.G. and N.R. were involved in the analysis and interpretation of data. D.R.C., P.G., M.K.M., M.D., M.A., C.B., O.P., K.H.V., N.M., E.M.V.A., N.K., R.S.H., J.D., T.G.B. and C.S. were responsible for drafting and revising the manuscript.
Corresponding authors
Ethics declarations
Competing interests
T.G.B. is an advisor to Novartis, AstraZeneca, Revolution Medicines, Array/Pfizer, Springworks, Strategia, Relay, Jazz, Rain, Engine, Granule Therapeutics and EcoR1 and receives research funding from Novartis and Revolution Medicines, Kinnate, Verastem and Strategia. N.I.V. served on an advisory board for Sanofi Genzyme. C.S. acknowledges grants from AstraZeneca, Boehringer-Ingelheim, Bristol Myers Squibb, Pfizer, Roche-Ventana, Invitae (previously Archer Dx—collaboration in minimal RD sequencing technologies), Ono Pharmaceutical, and Personalis. He is the chief investigator for the AZ MeRmaiD 1 and 2 clinical trials and is the Steering Committee Chair. He is also co-chief investigator of the NHS Galleri trial funded by GRAIL and a paid member of GRAIL’s Scientific Advisory Board (SAB). He receives consultant fees from Achilles Therapeutics (also an SAB member), Bicycle Therapeutics (also an SAB member), Genentech, Medicxi, China Innovation Center of Roche (CICoR) formerly Roche Innovation Center—Shanghai, Metabomed (until July 2022), Relay Therapeutics and the Sarah Cannon Research Institute. C.S. has received honoraria from Amgen, AstraZeneca, Bristol Myers Squibb, GlaxoSmithKline, Illumina, MSD, Novartis, Pfizer and Roche-Ventana; has previously held stock options in Apogen Biotechnologies and GRAIL; currently has stock options in Epic Bioscience and Bicycle Therapeutics and has stock options and is a cofounder of Achilles Therapeutics. C.S. declares a patent application (PCT/US2017/028013) for methods to lung cancer; targeting neoantigens (PCT/EP2016/059401); identifying patent response to immune checkpoint blockade (PCT/EP2016/071471), determining HLA LOH (PCT/GB2018/052004); predicting survival rates of patients with cancer (PCT/GB2020/050221), identifying patients who respond to cancer treatment (PCT/GB2018/051912); methods for lung cancer detection (US20190106751A1). He is an inventor on a European patent application (PCT/GB2017/053289) relating to assay technology to detect tumor recurrence. This patent has been licensed to a commercial entity, and under their terms of employment, C.S. is due a revenue share of any revenue generated from such license(s). E.M.V.A. is a consultant for Tango Therapeutics, Genome Medical, Invitae, Enara Bio, Janssen, Manifold Bio, Monte Rosa; receives research funding from Novartis, BMS; has equity in Tango Therapeutics, Genome Medical, Syapse, Enara Bio, Manifold Bio, Microsoft and Monte Rosa; has received travel reimbursement from Roche/Genentech and own institutional patents filed on chromatin mutations and immunotherapy response, and methods for clinical interpretation. C.E.M. is on the advisory board of Genentech; receives honoraria from Novartis, Guardant, Research and receives funding from Novartis, Revolution Medicines. C.M.B. is a consultant for Amgen, Foundation Medicine, Blueprint Medicines and Revolution Medicines; receives research funding from Novartis, AstraZeneca and Takeda and receives institutional research funding from Mirati, Spectrum, MedImmune and Roche. J.S.R.-F. reports receiving personal/consultancy fees from Goldman Sachs, Bain Capital, REPARE Therapeutics, Saga Diagnostics and Paige.AI, membership of the SAB of VolitionRx, REPARE Therapeutics and Paige.AI, membership of the Board of Directors (BOD) of Grupo Oncoclinicas, and ad hoc SAB of Astrazeneca, Merck, Daiichi Sankyo, Roche Tissue Diagnostics and Personalis, outside the scope of this study. H.Y. receives consulting fees from AstraZeneca, Daiichi, Taiho, Janssen, AbbVie, Blueprint, Black Diamond Research funding to my institution from AstraZeneca, Daiichi, Cullinan, Janssen, Blueprint, Black Diamond, Novartis, Pfizer, ERASCA. S.F.B. owns equity in, receives compensation from, serves as a consultant for and serves on the SAB and BOD of Volastra Therapeutics. He serves on the scientific advisory board of Meliora Therapeutics. M.J.-H. has consulted for, and is a member of, the Achilles Therapeutics Scientific Advisory Board and Steering Committee; has received speaker honoraria from Pfizer, Astex Pharmaceuticals, Oslo Cancer Cluster and Bristol Myers Squibb and is listed as a co-inventor on a European patent application relating to methods to detect lung cancer (PCT/US2017/028013). This patent has been licensed to commercial entities and, under terms of employment, M.J.-H. is due a share of any revenue generated from such license(s). The other authors have no competing interests to declare.
Peer review
Peer review information
Nature Genetics thanks the anonymous reviewers for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 APOBEC3B is detrimental for tumorigenesis in an EA3B mouse model of lung cancer.
a, Two by two contingency table of the number of mice with visible tumors (VT) or no visible tumors (NVT) by microCT at 3 months (two-sided Fisher’s exact test, *P = 0.0236). b, Representative images of p53 nuclear IHC staining (scale bar=10 µm, arrows indicate positive cells, E n = 5, EA3B n = 5 biological replicates). c, Quantification of p53 positive cells per lung area by IHC staining at 3 months post-induction (E n = 5, EA3B n = 5, mean ± SD, two-sided Mann-Whitney test, *P = 0.0159). d, Quantification of p53 positive cells per lung area by IHC staining at late timepoint (termination) (E n = 8, EA3B n = 8, mean ± SD, two-sided Mann-Whitney test). e, Quantification of Ki67-positive cells per mm2 of tumor at 3 months post-induction (E n = 9, EA3B n = 10, each dot represents a tumor, mean ± SD, two-sided unpaired t-test). f, Quantification of γH2AX-positive cells per mm2 of tumor at 3 months post-induction (E n = 9, EA3B n = 10, each dot represents a tumor, mean ± SD, two-sided Mann-Whitney test). g, Quantification of CD4+ cells per mm2 of tumor at 3 months post-induction (E n = 8, EA3B n = 7, each dot represents a tumor, mean ± SD, two-sided Mann-Whitney test, **P = 0.0086). h, Quantification of CD8+ cells per mm2 of tumor at 3 months post-induction (E = 8, EA3B = 8, each dot represents a tumor, mean ± SD, two-sided Mann-Whitney test, ***P = 0.0003). i, Representative IHC stainings of EGFRL858R, APOBEC3B, and CD4 and CD8 T cells (scale bar=50 µm, EGFRL858R E n = 9, EA3B n = 10, A3B E n = 9, EA3B n = 10, p53fl/fl E n = 5, EA3B n = 5, CD4 E n = 8, EA3B n = 7, CD8 E n = 8, EA3B n = 8). j, Intravenous transplantation using an EGFRL858R; p53fl/fl;APOBEC3B (EPA3B) mouse tumor cell line injected into a wildtype C57BL/6J mouse or a C57BL/6J EPA3B GEMM mouse. k, Quantification of EGFRL858R positive tumors in C57BL/6 wildtype versus EPA3B mice at 4 weeks (mean ± SD, two-sided Mann-Whitney test, n = 4, *P = 0.0286, each dot represents a mouse, C57BL/6 wildtype n = 4, C57BL/6J EPA3B GEMM n = 4). l, Quantification of EGFRL858R positive tumors in C57BL/6 wildtype versus EPA3B mice at 12 weeks (mean ± SD, two-sided Mann-Whitney test, n = 3, *P = 0.0286, each dot represents a mouse, C57BL/6 wildtype n = 4, C57BL/6J EPA3B GEMM n = 3). m, Representative IHC staining of EGFRL858R and APOBEC3B (scale bar=50 µm, 4 weeks C57BL/6 wildtype n = 4, C57BL/6J EPA3B GEMM n = 4, 12 weeks C57BL/6 wildtype n = 4, C57BL/6J EPA3B GEMM n = 3).
Extended Data Fig. 2 Subclonal A3B expression in treatment naive mice inhibits tumor growth.
a, Experimental set up of induction of subclonal APOBEC3B using TetO-EGFRL858R;CCSP-rtTA;Rosa26LSL-APOBEC3B/Cre-ER(T2)(EA3Bi) or TetO-EGFRL858R;CCSP-rtTA;Rosa26Cre-ER(T2)/+(Ei) mice. b, Tumor nodules per lung section per mouse at termination (Ei n = 10, EA3Bi n = 10, two-sided Mann-Whitney test, *P = 0.0494). c, Tumor area per lung area at termination (Ei n = 10, EA3Bi n = 10, two-sided Mann-Whitney test, *P = 0.0216). d, Survival curve of Ei versus EA3Bi mice (Ei n = 14, EA3Bi n = 17, each dot represents a mouse, Log-rank (Mantel-Cox) test, *P = 0.0358).
Extended Data Fig. 3 Putative resistance mutations in genes previously associated with TKI resistance in mouse tumor cell lines.
a, Comparison of EP and EPA3B mutation burdens in TKI naive and TKI resistant mouse lung cancer cell lines (mean ± SD, one-way ANOVA test, *P = 0.0135, *P = 0.0346, **P = 0.0039). b, Comparison of EP and EPA3B APOBEC driven mutations (TCN, C > T or C > G SNVs) in TKI naive and TKI resistant mouse lung cancer cell lines (mean ± SD, one-way ANOVA test, *P = 0.0333, *P = 0.0333, **P = 0.0012). c, Functional annotation of TCN mutations in potential TKI resistance genes with change in variant allele frequency shown (x=TCN, Red square=deleterious mutation, yellow square=mixed (neutral and deleterious), orange square=neutral). d, Significant subclonal enrichment of the APOBEC-associated mutation signature in the TRACERx patient with A3B driven D129N mutation in the type IIa PTP PTPRD (equivalent to D138N mutation in PTPRS ***P = 0.0002, two-sided one-sample Wilcoxon test).
Extended Data Fig. 4 APOBEC3 family member mRNA and protein levels in control and A3B knockout cell lines.
a, Immunoblot for APOBEC3B (A3B) protein levels in PC9 control (sgGFP) and A3B knockout (sgA3B) cell lines, (n = 3 biological replicates, 2 independent experiments). b, mRNA expression levels of APOBEC3 family members in control (sgGFP) and A3B knockout (sgA3B) PC9 cell lines (n = 3 biological replicates, mean ± SD, one-way ANOVA test, ***P = 0.0001). c, Immunoblot for A3B protein levels in HCC827 control (sgGFP) and A3B knockout (sgA3B) cell lines (n = 3 biological replicates, 2 independent experiments). d, mRNA expression levels of APOBEC3 family members in control (sgGFP) and A3B knockout (sgA3B) HCC827 cell lines (n = 3 biological replicates, mean ± SD, one-way ANOVA test, ***P = 0.0001). e, Immunoblot for A3B protein levels in H3122 control (sgCtrl) or A3B knockout (sgA3B) cell line (n = 1 biological replicate, 2 independent experiments). f, mRNA expression levels of APOBEC3 family members in control (sgGFP) and A3B knockout (sgA3B) H3122 cell lines (n = 2 biological replicates, mean ± SD, one-way ANOVA test, ****P < 0.0001). g, CellTiter-Glo (CTG) viability assay performed on A3B-deficient or A3B-proficient PC9 cells treated with DMSO for 7 days (n = 3 biological replicates, mean ± SD, two-sided t-test). h, CTG viability assay performed on A3B-deficient or A3B-proficient HCC827 cells treated with DMSO for 7 days (n = 3 biological replicates, mean ± SD, two-sided t-test). i, CTG viability assay performed on A3B-deficient or A3B-proficient H3122 cells treated with DMSO for 7 days (n = 3 biological replicates, mean ± SD, two-sided t-test, *P = 0.0293).
Extended Data Fig. 5 Knockdown of APOBEC3 family members under TKI treatment.
a, Western blot analyses for pEGFR and pERK1/2 to confirm loss with osimertinib treatment in PC9 and HCC827 cells treated with DMSO or 0.5 μM osimertinib (Osi) for 18 hours (PC9 n = 4 independent experiments, HCC827 n = 1 independent experiment). b–e, RT-qPCR analysis of APOBEC3 family members expression in PC9 cells treated with DMSO or 0.5 μM osimertinib for 18 hours, with siRNA knockdown of APOBEC3A (A3A), APOBEC3B (A3B), APOBEC3C (A3C) or APOBEC3F (A3F): A3A expression (b, n = 3 biological replicates, mean ± SD, one-way ANOVA test ****P < 0.0001); A3B expression (c, n = 3 biological replicates, mean ± SD, one-way ANOVA test, ****P < 0.001); A3C expression (d, n = 3 biological replicates, mean ± SD, one-way ANOVA test, **P = 0.0049, ****P < 0.0001); A3F expression (e, n = 3 biological replicates, mean ± SD, one-way ANOVA test ****P = < 0.001). f–i, RT-qPCR analysis of APOBEC3 family members expression in HCC827 cells treated with DMSO or 0.5 μM osimertinib for 18 hours, with siRNA knockdown of A3A, A3B, A3C or A3F: A3A expression (f, n = 3 biological replicates, mean ± SD, one-way ANOVA test, ***P = 0.0003); A3B expression (g, n = 3 biological replicates, mean ± SD, one-way ANOVA test, **P = 0.0011); A3C expression (h, n = 3 biological replicates, mean ± SD, one-way ANOVA test, ***P = 0.0002, **P = 0.0040); A3F expression (i, n = 3 biological replicates, mean ± SD, one-way ANOVA test, ****P < 0.0001).
Extended Data Fig. 6 TKI treatment induces increased A3B and decreased UNG expression and activity in pre-clinical models of lung adenocarcinoma.
a, Uracil excision capacity assay (UEC) using PC9 nuclear extracts treated with DMSO or 2 μM osimertinib (Osi) (n = 3 biological replicates, mean ± SD, two-tailed t-test, *P = 0.0275). b, UEC in HCC827 treated with DMSO or 0.4 µM osi (n = 3 biological replicates, mean ± SD, two-tailed t-test, ****P < 0.0001). c, Western blot (WB) from H1975 treated with DMSO, 0.1 µM or 0.5 μM crizotinib (CYTO: cytoplasmic; NUC: nuclear; H3: Histone H3; TUBB: beta-tubulin) (n = 3 biological replicates). d, APOBEC activity assay (AAA) using H1975 treated with DMSO or 1 µM osi (n = 3 biological replicates, mean ± SD, two-tailed t-test, **P = 0.0084). e, UEC in H1975 treated with DMSO or 1 uM osi (n = 3 biological replicates, mean ± SD, two-tailed t-test, **P = 0.0054). f, WB from H3122 treated with DMSO or 1 μM crizotinib (n = 3 biological replicates). g, AAA from H3122 treated with DMSO or 0.5 μM crizotinib (n = 3 biological replicates, mean ± SD, two-tailed t-test, *P = 0.0204). h, UEC in H3122 treated with DMSO or 0.5 μM crizotinib (n = 3 biological replicates, mean ± SD, two-tailed t-test, *P = 0.0123). i, WB of H2228 treated with DMSO or 0.5 μM alectinib for (n = 3 biological replicates). j, AAA from PC9 transduced with empty vector (shEV) or shRNA against A3B (shA3B-1) and treated with DMSO or 1 μM erlotinib (n = 3 biological replicates). k, WB from nuclear extracts of PC9 transduced with shEV or shA3B-1 alone or together with wild-type HA-tagged A3B or HA-tagged catalyticaly-inactive A3B mutant (E255A) expression plasmid (n = 3 biological replicates). l, AAA from PC9 as in panel k, in the absence of RNase A (n = 3 biological replicates). m, mRNA expression levels of APOBEC3 family members in control (shEV) and A3B knockdown (shA3B) PC9 (n = 3 biological replicates, mean ± SD, one-way ANOVA test, **P = 0.0059, ****P < 0.0001). n, Cell cycle analysis of PC9 treated with DMSO, 2 μM osimertinib or 1 μM palbociclib (Palbo) (n = 4 biological replicates, mean ± SD, two-tailed t-tests, *P = 0.012, **P = 0.0032, **P = 0.0071, **P = 0.0084, *P = 0.0105). o, RT-qPCR analysis of PC9 cells treated as in panel a, (n = 2 or 3 biological replicates, mean ± SD, one-way ANOVA test, ****P < 0.0001, *P = 0.0215, **P = 0.0018). Panels a–i, n: treatment for 18 hours.
Extended Data Fig. 7 EGFR inhibition induces A3B upregulation and UNG downregulation in xenograft models.
a, Western blot analysis using extracts of EGFR-mutant H1975 human NSCLC xenografts harvested after 4 days of treatment with vehicle or the indicated doses of osimertinib (TUBB: Tubulin Beta Class I) (n = 1 biological replicate). b, Western blot analyses of extracts of PC9 tumor xenografts treated with vehicle or 5 mg/kg osimertinib (n = 2 biological replicates). c, Representative images of IHC analysis of APOBEC3B (A3B) protein levels in 11-18 xenografts treated with vehicle, 12.5 mg/kg/day erlotinib, 7.5 mg/kg/day NF-κB inhibitor (NF-κBi, PBS-1086) or combination (Erlotinib + NF-κBi) for 2 months (scale: 60 µM, n = 2 biological replicates)17. d, Quantification of immunohistochemical staining for A3B in 11-18 xenografts treated with vehicle, erlotinib (Erl), NF-κB inhibitor (NF-κBi, PBS-1086) or combination (Erl + NF-κBi) for 2 months (n = 2 biological replicates). e, Representative images of IHC analysis of UNG protein levels in 11-18 xenografts treated with vehicle or 12.5 mg/kg/day erlotinib for 2 months (n = 2 biological replicates). f, Quantification of immunohistochemical staining for UNG in 11-18 xenografts treated with vehicle or erlotinib for 2 months (n = 2 biological replicates). g, RNA-Seq analysis upon treatment of a PDX model of human EGFR-driven lung adenocarcinoma with vehicle or erlotinib (2 days, 25 mg/kg) (n = 2 biological replicates). h, RNA-Seq analysis upon treatment of a PDX model of human EGFR-driven lung adenocarcinoma with vehicle or osimertinib (6 days, 10 mg/kg) (n = 3 biological replicates, mean ± SD, two-sided t-test, *P = 0.0267).
Extended Data Fig. 8 NF-κB signaling contributes to TKI-induced A3B upregulation, and expression of c-Jun and UNG are decreased upon TKI treatment.
a, RNA-Seq analysis of EGFR-mutant 11-18 cells treated with DMSO, 100 µM erlotinib (erl), 5 µM NF-κB inhibitor (NF-κBi, PBS-1086) or combination (Erl+NF-κBi) (n = 3 biological replicates, mean ± SEM, one-way ANOVA test, ****P < 0.0001). b, Western blot analysis of extracts from PC9 treated with DMSO or with TNFα for 8.5 hours (n = 3 biological replicates). c, RT-qPCR analysis of TNFα-treated PC9 (n = 3 biological replicates, mean ± SD, two-tailed t-test, *P = 0.0406, *P = 0.0299, **P = 0.0024). d, RT-qPCR validation of RELA and RELB knockdown in PC9 with non-targeting vector or combination of shRELA-1+shRELB-1 (mix1) or shRELA-2+shRELB-2 (mix2) (n = 3 biological replicates; mean ± SD, one-way ANOVA test, ****P < 0.0001). e, RT-qPCR analysis of APOBEC3B (A3B) in PC9 with non-targeting vector or mix1 or mix2, treated with DMSO or 500 nM osi for 1 day (n = 3 biological replicates; mean ± SD, two-tailed t-test, *P = 0.0465, **P = 0.0026). f, Western blot analysis of PC9 used in e (n = 3 biological replicates). g, APOBEC activity assay of PC9 used in f (n = 3 biological replicates). h–j, Single-cell RNA-Seq expression in lung cancer cells from patient tumors at treatment naïve (TN, 762 cells), residual disease (RD, 553 cells) and progressive disease (PD, 988 cells) of: A3B (h), RelA (i) and RelB (j) (all data points shown, two-sided Wilcoxon test with Holm correction, ****P < 2.22e-16). k, Single-cell RNA-Seq analysis of NF-κB signature (from Gilmore_Core_NFκB_Pathway, GSEA, C2) in tumors from panels h–j (mean ± SD, two-sided Wilcoxon test with Holm correction, ****P < 2.22e-16). l, RT-qPCR analysis of c-JUN in PC9 treated with DMSO or 2 μM osimertinib for 9 days (n = 3 biological replicates, mean ± SEM, two-tailed t-test, ***P = 0.0009). m, RT-qPCR analysis of PC9 with non-targeting (siNTC) or c-JUN siRNA, treated with DMSO or 2 μM osimertinib for 18 hours (n = 3 biological replicates, mean ± SD, one-way ANOVA test, ****P < 0.0001). Boxplots: middle line=median, lower and upper hinges=first and third quartiles, lower and upper whiskers=smallest and largest values within 1.5×inter-quartile range from hinges.
Extended Data Fig. 9 Mutation burden and putative resistance mutations in genes previously associated with TKI resistance in PC9 TKI resistant cell line.
a, Mutation burden quantified in APOBEC3B (A3B)-deficient (A3B KO), and A3B-proficient (A3B WT) single cell cloned PC9 cells treated with osimertinib for 3 months (n = 6 biological replicates, mean ± SD, two-tailed Mann-Whitney test). b, Western blot analysis of PC9 cells treated with non-targeting (siNTC) or NRXN3-targeting (siNRXN3) siRNA and treated with DMSO or 500 nM osimertinib for 2 days (n = 3 biological replicates). c, RT-qPCR-based validation of NRXN3 knockdown in cells shown in a (n = 3 technical replicates, mean ± SD, two-sided t-test performed on ΔCt values shown, ***P = 0.0007). d, IC50 analysis of PC9 siNTC or siNRXN3 after 3-day treatment (n = 5 biological replicates for each of the following doses of osimertinib: 0 nM, 5 nM, 50 nM, 100 nM, 500 nM and 5000 nM, mean ± SD, two-sided t-test, ***P = 0.0004).
Extended Data Fig. 10 Expression of APOBEC3 enzymes in clinical samples upon targeted therapy treatment.
a, Comparison of APOBEC3B (A3B) expression levels (Exp: batch corrected TPM) measured using RNA-Seq analysis in human NSCLC specimens driven by EGFR and ALK driver mutations obtained before treatment (Pre-TKI, 32 samples), or post-treatment (Post-TKI, 42 samples) (all data points shown, two-sided t-test, *P = 0.011). b, Comparison of APOBEC3 (A3) family member expression levels (Exp: batch corrected Log (TPM + 1) measured using RNA-seq analysis in human NSCLC specimens obtained at treatment naïve (TN), residual disease (RD) or progressive disease (PD) with TKI (all data points shown, 762, 553, and 988 cells per group respectively, two-sided Wilcoxon test with Holm correction, *P = 0.02). c, Boxplot of normalized A3 family member expression measured using scRNA-seq obtained from the same samples as b (all data points shown, 762, 553, and 988 cells per group respectively, two-sided Wilcoxon test with Holm correction, *P < 0.05, **P < 0.01, ****P < 0.001, d=effect size calculated using a Cohen test). Boxplots: middle line=median, lower and upper hinges=first and third quartiles, lower and upper whiskers=smallest and largest values within 1.5×inter-quartile range from hinges.
Supplementary information
Supplementary Information
Supplementary Note.
Supplementary Tables
Supplementary Table 1: Mouse tumor cell line WES. Supplementary Table 2a: Metadata of patient tumor samples analyzed using single-cell RNA-seq analysis. Supplementary Table 2b: Metadata of patient tumor samples analyzed using bulk RNA-seq analysis. Supplementary Table 2c: Metadata of human biopsies stained for APOBEC3B via immunohistochemistry. Supplementary Table 3: Signature exposures calculated from WGS of A3B-proficient and A3B-deficient single-cell clones PC9 cell lines, treated with osimertinib or DMSO for 3 months until resistant. Supplementary Table 4: Metadata of patient tumor samples processed for WES and subsequent mutational signature analysis. Supplementary Table 5a: Mutations observed in EGFR- and ALK-driven patients with lung cancer. Supplementary Table 5b: Selective putative resistance mutations observed in EGFR- and ALK-driven patients with lung cancer. Supplementary Table 6: Metadata of mice in animal studies.
Source data
Source Data Fig. 1
Unprocessed western blots and/or gels.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Caswell, D.R., Gui, P., Mayekar, M.K. et al. The role of APOBEC3B in lung tumor evolution and targeted cancer therapy resistance. Nat Genet 56, 60–73 (2024). https://doi.org/10.1038/s41588-023-01592-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-023-01592-8
