
Abstract
Harnessing the potential of human embryonic stem cells to mimic normal and aberrant development with standardized models is a pressing challenge. Here we use micropattern technology to recapitulate early human neurulation in large numbers of nearly identical structures called neuruloids. Dual-SMAD inhibition followed by bone morphogenic protein 4 stimulation induced self-organization of neuruloids harboring neural progenitors, neural crest, sensory placode and epidermis. Single-cell transcriptomics unveiled the precise identities and timing of fate specification. Investigation of the molecular mechanism of neuruloid self-organization revealed a pulse of pSMAD1 at the edge that induced epidermis, whose juxtaposition to central neural fates specifies neural crest and placodes, modulated by fibroblast growth factor and Wnt. Neuruloids provide a unique opportunity to study the developmental aspects of human diseases. Using isogenic Huntington’s disease human embryonic stem cells and deep neural network analysis, we show how specific phenotypic signatures arise in our model of early human development as a consequence of mutant huntingtin protein, outlining an approach for phenotypic drug screening.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
Data availability
The accession number for the single-cell RNA-seq data reported in this paper is GSE118682.
Code availability
The code is available upon request or directly from the GitHub submission: https://github.com/jjmetzger/neural_seg.
References
Ozair, M. Z., Kintner, C. & Brivanlou, A. H. Neural induction and early patterning in vertebrates. Wiley Interdiscip. Rev. Dev. Biol. 2, 479–498 (2013).
Chambers, S. M. et al. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nat. Biotechnol. 27, 275–280 (2009).
Ozair, M. Z., Noggle, S., Warmflash, A., Krzyspiak, J. E. & Brivanlou, A. H. SMAD7 directly converts human embryonic stem cells to telencephalic fate by a default mechanism. Stem Cells 31, 35–47 (2013).
Dincer, Z. et al. Specification of functional cranial placode derivatives from human pluripotent stem cells. Cell Rep. 5, 1387–1402 (2013).
Tchieu, J. et al. A modular platform for differentiation of human PSCs into all major ectodermal lineages. Cell Stem Cell 21, 399–410 e397 (2017).
Hemmati-Brivanlou, A. & Melton, D. A. Inhibition of activin receptor signaling promotes neuralization in Xenopus. Cell 77, 273–281 (1994).
Munoz-Sanjuan, I. & Brivanlou, A. H. Neural induction, the default model and embryonic stem cells. Nat. Rev. Neurosci. 3, 271–280 (2002).
Wilson, P. A., Lagna, G., Suzuki, A. & Hemmati-Brivanlou, A. Concentration-dependent patterning of the Xenopus ectoderm by BMP4 and its signal transducer Smad1. Development 124, 3177–3184 (1997).
Bhattacharyya, S. & Bronner-Fraser, M. Hierarchy of regulatory events in sensory placode development. Curr. Opin. Genet. Dev. 14, 520–526 (2004).
Litsiou, A., Hanson, S. & Streit, A. A balance of FGF, BMP and WNT signalling positions the future placode territory in the head. Development 132, 4051–4062 (2005).
Sauka-Spengler, T. & Bronner-Fraser, M. A gene regulatory network orchestrates neural crest formation. Nat. Rev. Mol. Cell Biol. 9, 557–568 (2008).
Betancur, P., Bronner-Fraser, M. & Sauka-Spengler, T. Genomic code for Sox10 activation reveals a key regulatory enhancer for cranial neural crest. Proc. Natl Acad. Sci. USA 107, 3570–3575 (2010).
Kwon, H. J., Bhat, N., Sweet, E. M., Cornell, R. A. & Riley, B. B. Identification of early requirements for preplacodal ectoderm and sensory organ development. PLoS Genet. 6, e1001133 (2010).
Schlosser, G. Early embryonic specification of vertebrate cranial placodes. Wiley Interdiscip. Rev. Dev. Biol. 3, 349–363 (2014).
Stuhlmiller, T. J. & Garcia-Castro, M. I. FGF/MAPK signaling is required in the gastrula epiblast for avian neural crest induction. Development 139, 289–300 (2012).
Reichert, S., Randall, R. A. & Hill, C. S. A BMP regulatory network controls ectodermal cell fate decisions at the neural plate border. Development 140, 4435–4444 (2013).
Patthey, C. & Gunhaga, L. Signaling pathways regulating ectodermal cell fate choices. Exp. Cell Res. 321, 11–16 (2014).
Leung, A. W. et al. WNT/beta-catenin signaling mediates human neural crest induction via a pre-neural border intermediate. Development 143, 398–410 (2016).
Greenberg, F. DiGeorge syndrome: an historical review of clinical and cytogenetic features. J. Med. Genet. 30, 803–806 (1993).
Roizen, N. J. & Patterson, D. Down’s syndrome. Lancet 361, 1281–1289 (2003).
Sarkozy, A., Digilio, M. C. & Dallapiccola, B. Leopard syndrome. Orphanet J. Rare Dis. 3, 13 (2008).
Ferner, R. E. Neurofibromatosis 1. Eur. J. Hum. Genet. 15, 131–138 (2007).
Greene, N. D. & Copp, A. J. Neural tube defects. Ann. Rev. Neurosci. 37, 221–242 (2014).
Elkabetz, Y. et al. Human ES cell-derived neural rosettes reveal a functionally distinct early neural stem cell stage. Genes Dev. 22, 152–165 (2008).
Ozair, M. Z. et al. hPSC modeling reveals that fate selection of cortical deep projection neurons occurs in the subplate. Cell Stem Cell 23, 60–73 e66 (2018).
Warmflash, A., Sorre, B., Etoc, F., Siggia, E. D. & Brivanlou, A. H. A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat. Methods 11, 847–854 (2014).
Deglincerti, A., Etoc, F., Ozair, M. Z. & Brivanlou, A. H. Self-organization of spatial patterning in human embryonic stem cells. Curr. Top. Dev. Biol. 116, 99–113 (2016).
Etoc, F. et al. A balance between secreted inhibitors and edge sensing controls gastruloid self-organization. Dev. Cell. 39, 302–315 (2016).
Knight, G. T. et al. Engineering induction of singular rosette emergence within hPSC-derived tissues. eLife 7, e37549 (2018).
Broccoli, V., Colombo, E. & Cossu, G. Dmbx1 is a paired-box containing gene specifically expressed in the caudal most brain structures. Mech. Dev. 114, 219–223 (2002).
Grenningloh, G., Soehrman, S., Bondallaz, P., Ruchti, E. & Cadas, H. Role of the microtubule destabilizing proteins SCG10 and stathmin in neuronal growth. J. Neurobiol. 58, 60–69 (2004).
Mallika, C., Guo, Q. & Li, J. Y. Gbx2 is essential for maintaining thalamic neuron identity and repressing habenular characters in the developing thalamus. Dev. Biol. 407, 26–39 (2015).
Quina, L. A., Wang, S., Ng, L. & Turner, E. E. Brn3a and Nurr1 mediate a gene regulatory pathway for habenula development. J. Neurosci. 29, 14309–14322 (2009).
Inoue, T., Chisaka, O., Matsunami, H. & Takeichi, M. Cadherin-6 expression transiently delineates specific rhombomeres, other neural tube subdivisions, and neural crest subpopulations in mouse embryos. Dev. Biol. 183, 183–194 (1997).
Van de Putte, T. et al. Mice lacking ZFHX1B, the gene that codes for Smad-interacting protein-1, reveal a role for multiple neural crest cell defects in the etiology of hirschsprung disease-mental retardation syndrome. Am. J. Hum. Genet. 72, 465–470 (2003).
Bolos, V. et al. The transcription factor slug represses E-cadherin expression and induces epithelial to mesenchymal transitions: a comparison with snail and E47 repressors. J. Cell Sci. 116, 499–511 (2003).
Huang, M. et al. Generating trunk neural crest from human pluripotent stem cells. Sci. Rep. 6, 19727 (2016).
Bhatt, S., Diaz, R. & Trainor, P. A. Signals and switches in mammalian neural crest cell differentiation. Cold Spring Harbor Perspect. Biol. 5, pii: a008326 (2013).
Ma, Q., Chen, Z., del Barco Barrantes, I., de la Pompa, J. L. & Anderson, D. J. neurogenin1 is essential for the determination of neuronal precursors for proximal cranial sensory ganglia. Neuron 20, 469–482 (1998).
Bae, S., Bessho, Y., Hojo, M. & Kageyama, R. The bHLH gene Hes6, an inhibitor of Hes1, promotes neuronal differentiation. Development 127, 2933–2943 (2000).
Byrne, C., Tainsky, M. & Fuchs, E. Programming gene expression in developing epidermis. Development 120, 2369–2383 (1994).
Moll, R., Divo, M. & Langbein, L. The human keratins: biology and pathology. Histochem. Cell Biol. 129, 705–733 (2008).
Lavery, D. L., Davenport, I. R., Turnbull, Y. D., Wheeler, G. N. & Hoppler, S. Wnt6 expression in epidermis and epithelial tissues during Xenopus organogenesis. Dev. Dyn. 237, 768–779 (2008).
Arabzadeh, A., Troy, T. C. & Turksen, K. Role of the Cldn6 cytoplasmic tail domain in membrane targeting and epidermal differentiation in vivo. Mol. Cell. Biol. 26, 5876–5887 (2006).
Kaufman, C. K. et al. GATA-3: an unexpected regulator of cell lineage determination in skin. Genes Dev. 17, 2108–2122 (2003).
Onorati, M. et al. Molecular and functional definition of the developing human striatum. Nat. Neurosci. 17, 1804–1815 (2014).
Selleck, M. A. & Bronner-Fraser, M. Origins of the avian neural crest: the role of neural plate-epidermal interactions. Development 121, 525–538 (1995).
Lo Sardo, V. et al. An evolutionary recent neuroepithelial cell adhesion function of huntingtin implicates ADAM10-N-cadherin. Nat. Neurosci. 15, 713–721 (2012).
Conforti, P. et al. Faulty neuronal determination and cell polarization are reverted by modulating HD early phenotypes. Proc. Natl Acad. Sci. USA 115, E762–E771 (2018).
Ruzo, A. et al. Chromosomal instability during neurogenesis in Huntington’s disease. Development 145, pii: dev156844 (2018).
Molero, A. E. et al. Selective expression of mutant huntingtin during development recapitulates characteristic features of Huntington’s disease. Proc. Natl Acad. Sci. USA 113, 5736–5741 (2016).
Arteaga-Bracho, E. E. et al. Postnatal and adult consequences of loss of huntingtin during development: implications for Huntington’s disease. Neurobiol. Dis. 96, 144–155 (2016).
Siebzehnrubl, F. A. et al. Early postnatal behavioral, cellular, and molecular changes in models of Huntington disease are reversible by HDAC inhibition. Proc. Natl Acad. Sci. USA 115, E8765–E8774 (2018).
Shelhamer, E., Long, J. & Darrell, T. Fully convolutional networks for semantic segmentation. IEEE Trans. Pattern Anal. Mach. Intell. 39, 640–651 (2017).
Barnat, M., Le Friec, J., Benstaali, C. & Humbert, S. Huntingtin-mediated multipolar-bipolar transition of newborn cortical neurons is critical for their postnatal neuronal morphology. Neuron 93, 99–114 (2017).
Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420 (2018).
Labadorf, A. et al. RNA sequence analysis of human Huntington disease brain reveals an extensive increase in inflammatory and developmental gene expression. PloS ONE 10, e0143563 (2015).
Lancaster, M. A. & Knoblich, J. A. Organogenesis in a dish: modeling development and disease using organoid technologies. Science 345, 1247125 (2014).
Lancaster, M. A. et al. Cerebral organoids model human brain development and microcephaly. Nature 501, 373–379 (2013).
Xue, X. et al. Mechanics-guided embryonic patterning of neuroectoderm tissue from human pluripotent stem cells. Nat. Mater. 17, 633–641 (2018).
Hikosaka, O. The habenula: from stress evasion to value-based decision-making. Nat. Rev. Neurosci. 11, 503–513 (2010).
Labbadia, J. & Morimoto, R. I. Huntington’s disease: underlying molecular mechanisms and emerging concepts. Trends Biochem. Sci. 38, 378–385 (2013).
Landles, C. & Bates, G. P. Huntingtin and the molecular pathogenesis of Huntington’s disease. fourth in molecular medicine review series. EMBO Rep. 5, 958–963 (2004).
Ruzo, A. et al. Discovery of novel isoforms of huntingtin reveals a new hominid-specific exon. PloS ONE 10, e0127687 (2015).
Tourette, C. et al. A large-scale huntingtin protein interaction network implicates Rho GTPase signaling pathways in Huntington disease. J. Biol. Chem. 289, 6709–6726 (2014).
Caviston, J. P. & Holzbaur, E. L. Huntingtin as an essential integrator of intracellular vesicular trafficking. Trends Cell Biol. 19, 147–155 (2009).
Mehler, M. F. et al. Loss-of-huntingtin in medial and lateral ganglionic lineages differentially disrupts regional interneuron and projection neuron subtypes and promotes Huntington’s disease-associated behavioral, cellular and pathological hallmarks. J. Neurosci. 39, 1892–1909 (2019).
Paulsen, J. S. et al. Clinical and biomarker changes in premanifest Huntington disease show trial feasibility: a decade of the PREDICT-HD study. Front. Aging Neurosci. 6, 78 (2014).
Ciarochi, J. A. et al. Patterns of co-occurring gray matter concentration loss across the Huntington disease prodrome. Front. Neurol. 7, 147 (2016).
Harrington, D. L. et al. Network topology and functional connectivity disturbances precede the onset of Huntington’s disease. Brain 138, 2332–2346 (2015).
Paulsen, J. S. et al. Detection of Huntington’s disease decades before diagnosis: the Predict-HD study. J. Neurol. Neurosurg. Psychiatry 79, 874–880 (2008).
Ernst, A. et al. Neurogenesis in the striatum of the adult human brain. Cell 156, 1072–1083 (2014).
Shi, Y., Kirwan, P. & Livesey, F. J. Directed differentiation of human pluripotent stem cells to cerebral cortex neurons and neural networks. Nat. Protoc. 7, 1836–1846 (2012).
Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888–1902.e21 (2019).
Sommer, C. et al. Ilastik: Interactive learning and segmentation toolkit. In Proc. 8th IEEE International Symposium on Biomedical Imaging (ed Sommer, C. et al.) 230–233 (ISBI, 2011).
Jegou, S. et al. The one hundred layers tiramisu: fully convolutional DenseNets for semantic segmentation. Preprint at https://arXiv.org/abs/1611.09326 (2016).
Kingma, D. et al. Adam A method for stochastic optimization. In Proc. 3rd International Conference on Learning Representations (ed Kingma, D. P. & Ba, J.) (ICLR, 2015).
Acknowledgements
We thank J. Auerbach for technical assistance, as well as J. Naftaly and C. Lara. We would like to thank C. Zhao and the Genomics Resource Center at the Rockefeller University for help and advice regarding single-cell RNA sequencing. We thank E. Siggia for constructive criticism and the members of the Brivanlou-Siggia laboratories for critical discussions and comments on the manuscript. We appreciated discussions and comments from D. Felsenfeld, T. Vogt and I. Muñoz-Sanjuán (CHDI Foundation). This work was supported by the CHDI Foundation (A-9423), an SRA from RUMI Scientific and private funding from the Rockefeller University.
Author information
Authors and Affiliations
Contributions
T.H. performed experiments. J.J.M. developed the image analysis pipeline. T.R. performed experiments and analyzed single-cell RNA-seq data. Z.O. created and validated transgenic hESC lines used for live imaging. F.E. directed the project. A.H.B. conceived and designed the project. F.E. and A.H.B. wrote the manuscript with input from all authors.
Corresponding author
Ethics declarations
Competing interests
A.H.B. is the co-founder of RUMI Scientific. Both A.H.B. and F.E. are shareholders of RUMI Scientific.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 Analysis of apical markers and seeding density for neural rosettes on micropatterned colonies.
(A) Day 7 micropatterned culture after 7days with SB+LDN. Colonies of the following diameters are shown: 80, 200, 500, 800 and 1000 μm. Samples were stained with DAPI (gray), the neural differentiation marker PAX6 (green) and neural adhesion protein N-cadherin (N-CAD, orange). (B) Representative immunofluorescence images of PAX6 and N-CAD at day 7 of 200 and 500 μm micropatterned culture with multiple z from bottom to top. (C) Representative immunofluorescence images of atypical protein kinase C (aPKC), partitioning defective 3 (PAR3) at day 7 of 200 and 500 μm micropatterned culture with multiple z from bottom to top. Experiments in panels A-C have been repeated more than three times. (D) Representative immunofluorescence images of 80, 200, 500 and 1000 μm of day 7 micropatterned colonies by PAX6 (green) and N-CAD (orange) with 3 different initial cell seeding number: 0.38, 0.50 and 0.64 million cells. (Right) Quantification of lumen size at different initial seeding densities. N-CAD+ lumen sizes of 200 μm colonies are shown as a ratio of inner lumen area to colony area (n>80). Data are mean ± standard deviation. Data from panel D are from a single experiment with n>80 colonies for each condition. Scale bars: 50 μm except (A) 800 μm.
Supplementary Figure 2 Detailed characterization of neural rosettes and self-organized neuruloids on micropatterns.
(A) Representative immunofluorescence images of day 7 micropatterned colonies of 200 and 500 μm stained with N-CAD. Stars indicate the micropatterns with incomplete lumen. Representative images of the complete (above) and the incomplete (bottom) lumen in the 500 μm colonies are shown. Representative images of the satellite N-CAD+ loci are also shown. (B) Quantification of the proportion of closed lumens in 200 and 500 μm colonies, as defined by the continuity of the N-CAD ring at the center of the rosette as shown in panel (A). (C) Representative immunofluorescence images of the monolayer differentiation protocol from day 1 to 6 with PAX6 (green), OCT4 (red) and N-CAD (orange). (D) Quantitative PCR of various lineage-specific genes from day 1 to 7 of monolayer culture. Data were averaged from three technical replicates and error bars quantify standard deviations (E) Side view by resliced z-stack around N-CAD+ center region of day 7 neuruloid micropatterned colonies (BMP4 50 ng/ml) of 80, 200, 500 and 1000 μm. Samples were stained by DAPI (gray), PAX6 (green) and N-CAD (orange). The 1000 um diameter sample is shown from center to edge only. (F) Quantification of the area occupied by each marker across 145 neuruloid colonies from 3 independent experiments. Violin plots of the area positive for different markers (n=145). The violins are kernel density estimates of the underlying distribution. (G) Immunofluorescence analysis of 500 μm micropatterned colonies differentiated by BMP4 (50 ng/ml) as well as BMP4 (3 ng/ml) protocol. Samples were stained with DAPI (gray), PAX6 (green), neural crest marker SOX10 (red) and cranial placode marker SIX1 (orange). (H) 500 μm neuruloids at day 7 with different BMP4 concentrations: 0, 3, 13 and 50 ng/ml. SIX1 expression is shown within 25 patterns for comparison of the four different conditions. (I) Radially averaged marker expression from 3 different colonies show the TFAP2A+ only cells covering the full surface of the colony. Experiments in panels C,G,H and I have been repeated more than three times. Scale bars: (A) left 200 μm, middle 500 μm, right 100 μm; (C) 50 μm; (E) 50 μm except “80 μm” 10 μm; (G) 50 μm; (H) 500 μm; (I) 50 μm.
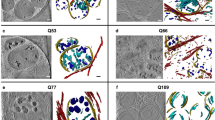
Supplementary Figure 3 Single-cell RNA-seq of neuruloid and validation of additional markers: neural populations.
(A) Cell numbers of the two single-cell RNA-seq RUES2 replicate transcriptomes produced in this study performed with Cell Ranger software and Seurat. Only the cells that passed QC were used in the analyses presented (see Methods). (B) UMAP plot showing the cell distributions of the two replicates. (C) Bar plot with the proportion of cells per cluster in both RUES2 replicates showing a difference of at most ~20% in the neural crest population. (D) UMAP graphs of: an overview of the clusters unbiasedly called using Louvain algorithm; OTX2 and PAX3 expression defining a dorsal anterior fate; EMX2 and LHX5 enriched in the caudal forebrain cluster NE1 and IRX3 and EN1 enriched in the midbrain (including the diencephalon) cluster NE2. (E) Expression region of forebrain and midbrain markers Pax6, Otx2, Emx2 and Dmbx1 from www.emouseatlas.org (Right) Scheme of markers in human embryo at 26 days post conception (d.p.c.). (F) 500 μm neuruloid (BMP4 3 ng/ml) at day 7 stained with DAPI, PAX6, LHX5. Zoomed and side views of the PAX6+ center region reveal a PAX6+LHX5+ population (forebrain) and a PAX6+LHX5- and PAX6-LHX5- populations corresponding to midbrain cells. (G) Characterization of the early-born neurons in the neuruloid. Battery of marker genes enriched in the neurons cluster. Zoomed immunofluorescence image of 500 μm neuruloid (BMP4 3 ng/ml) at day 7. Sample was stained with OTX2 and STMN2, a regulator of microtubule stability characteristic of early neurons. (H) UMAP plot and corresponding day 7 neuruloid stain of EBF3, specifically enriched in the diencephalic neurons cluster and not in the placode derived neurons, and STMN2. Immunofluorescence experiments in panels F and H have been repeated more than three times. Scale bars: 50 μm.
Supplementary Figure 4 Single-cell RNA-seq of neuruloid and validation of additional markers: neural crest, placodal and prospective epidermis.
(A) Single-cell gene expression UMAP plots with a battery of markers used to determine the anterior cranial nature of the neural crest population in the neuruloid. (B) Venn diagram with the overlap between isolated chick cranial neural crest genes (Simoes-Costa et al. 2014) and our collection of 149 NC marker genes (C) Immunofluorescence analysis of the 500 μm neuruloid (BMP4 3 ng/ml) NC at day 7. Sample was stained with DAPI, PAX6, SOX10, ETS1 and SNAIL. (D) Placodal cells express ISL1 and SIX1. (E) UMAP plot of POU4F1(BRN3A) and corresponding day 7 neuruloid stain of POU4F1(BRN3A), STMN2 and SIX1, marking specifically the sensory neurons coming from the trigeminal placode. (F) 500 μm neuruloid (BMP4 3 ng/ml) at day 7 stained with DAPI, PAX6, SOX10 and KRT18. Side view of the entire colony cross section was also shown. Immunofluorescence experiments in panels C, D, E and F have been repeated more than three times. Scale bars: 50 μm.
Supplementary Figure 5 Early human embryonic gene markers for each of the main cell types present in the neuruloid.
Marker genes enriched in the NE1, NE2, Neuron, Skin, Neural Crest and Placode populations. The complete marker list for each population can be found in Supplementary Table 1 and the associated statistical analysis is described in the method section. For each population, 5 markers are shown together with their expression on the UMAP plot.
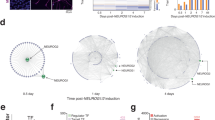
Supplementary Figure 6 Reproducibility of the neuruloid in different cell lines.
(A) Day7 neuruloid showing a similar structure to Fig. 3f when using a different hESC line (RUES1) and the iPSC line BJ-1. A typical colony is stained with PAX6 (green), SOX10 (red) and SIX1 (orange). Immunofluorescence experiment in panels A has been repeated more than three times. (B) qPCR data for 10 marker genes of NE, NC, Placode, Skin and Neurons clusters. Expression levels are shown for 3 independent experiments across all the three lines. Data are plotted with a bar centered on the mean value and standard deviations (C) UMAP plots of MKI67 and TOP2A which are enriched in cluster U1 (see also Supplementary Table 1) suggesting this cluster still represents a population of dividing cells. Scale bars: 50 μm.
Supplementary Figure 7 Performance of our neural network analysis in neural rosettes and neuruloid HD phenotypes.
(A) Comparison of lumen segmentation by the filter-based machine-learning framework Ilastik and a deep neural network trained on the same images and applied to previously unseen data. (Left) Examples in which the two approaches have similar performance, (Right) examples where the neural network performs significantly better. The neural network almost perfectly segments the lumen in a wide range of conditions, even when the filter-based classifier fails due to the reasons indicated on the side of the right column. (B) Immunofluorescence analysis of 500 μm micropatterned colonies of RUES2, 20CAG and 56CAG by PAX6, SOX10 at day 7 of SB+BMP4 (3 ng/ml) protocol. (Right) Quantification of PAX6+ area. N>20 colonies for every condition. Data are centered around the mean, with standard deviations. One-way anova test was used to quantify p-values. Scale bars: 50 μm.
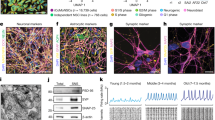
Supplementary Figure 8 Differentially expressed genes in the HD neuruloid (56CAG) and detailed analysis of the HD phenotype.
(A) Cell numbers and QC measures of the 56CAG single-cell RNA-seq. Only the cells that passed QC and were not proliferating were kept for subsequent analyses. Bar plot shows the proportion of cells allocated to each neuruloid cell population in both RUES2 replicates and 56CAG. (B) Gene ontology analyses using DAVID (https://david.ncifcrf.gov) for NE- and NC-specific DEG between RUES2 and 56CAG, the associated statistical analysis is described in the method section. (C) Violin plots showing the distribution of normalized gene expression values for the genes in Fig. 9, f. The violins are kernel density estimates of the underlying distribution. (D) qPCR validation of the down-regulation of selected WNT/PCP and cytoskeleton-associated genes in the 56CAG neuruloid versus WT RUES2. (E) Representative image of a day 7 neuruloid treated with Blebbistatin (5 μM) and stained for DAPI, N-CAD, PAX6 and COL4. (Top) side view, (Bottom) top view. Profile plot of N-CAD and PAX6 is also shown. Experiment in panels E F have been repeated more than three times. Scale bars: 50 μm.
Supplementary Figure 9
Full Western Blot images for Fig. 7b.
Supplementary Information
Supplementary Information
Supplementary Figs. 1–9.
Supplementary Table 1
Neuruloid single-cell RNA-seq gene markers for the main cell populations and DEGs in HD neuruloid phenotype (56CAG).
Supplementary Video 1
Three-dimensional structure of the neuruloid immunofluorescence z-stack image of the neuruloid stained with PAX6 (Green), SOX10 (Red), SIX1 (Orange) and TFAP2A (Cyan). The movie starts from the bottom of the neuruloid to the top.
Supplementary Video 2
Live-cell imaging of the neuruloid formation. Time-lapse microscopy video from days 3–6 of neuruloid formation using a live reporter cell line (PAX6::H2B-Citrin and SOX10::H2B-tdTomato). PAX6 is colored in green and SOX10 is red.
Rights and permissions
About this article

Cite this article
Haremaki, T., Metzger, J.J., Rito, T. et al. Self-organizing neuruloids model developmental aspects of Huntington’s disease in the ectodermal compartment. Nat Biotechnol 37, 1198–1208 (2019). https://doi.org/10.1038/s41587-019-0237-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41587-019-0237-5
This article is cited by
-
Competence for neural crest induction is controlled by hydrostatic pressure through Yap
Nature Cell Biology (2024)
-
A patterned human neural tube model using microfluidic gradients
Nature (2024)
-
Mechanically enhanced biogenesis of gut spheroids with instability-driven morphomechanics
Nature Communications (2023)
-
Primate gastrulation and early organogenesis at single-cell resolution
Nature (2022)
-
A Progressive Loss of phosphoSer138-Profilin Aligns with Symptomatic Course in the R6/2 Mouse Model of Huntington’s Disease: Possible Sex-Dependent Signaling
Cellular and Molecular Neurobiology (2022)
