
Abstract
Immunosuppression increases the risk of cancers that are associated with viral infection1. In particular, the risk of squamous cell carcinoma of the skin—which has been associated with beta human papillomavirus (β-HPV) infection—is increased by more than 100-fold in immunosuppressed patients2,3,4. Previous studies have not established a causative role for HPVs in driving the development of skin cancer. Here we show that T cell immunity against commensal papillomaviruses suppresses skin cancer in immunocompetent hosts, and the loss of this immunity—rather than the oncogenic effect of HPVs—causes the markedly increased risk of skin cancer in immunosuppressed patients. To investigate the effects of papillomavirus on carcinogen-driven skin cancer, we colonized several strains of immunocompetent mice with mouse papillomavirus type 1 (MmuPV1)5. Mice with natural immunity against MmuPV1 after colonization and acquired immunity through the transfer of T cells from immune mice or by MmuPV1 vaccination were protected against skin carcinogenesis induced by chemicals or by ultraviolet radiation in a manner dependent on CD8+ T cells. RNA and DNA in situ hybridization probes for 25 commensal β-HPVs revealed a significant reduction in viral activity and load in human skin cancer compared with the adjacent healthy skin, suggesting a strong immune selection against virus-positive malignant cells. Consistently, E7 peptides from β-HPVs activated CD8+ T cells from unaffected human skin. Our findings reveal a beneficial role for commensal viruses and establish a foundation for immune-based approaches that could block the development of skin cancer by boosting immunity against the commensal HPVs present in all of our skin.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others


Data availability
The data that support the findings of this study are available from the corresponding author on reasonable request. RNA-seq data have been deposited to the NCBI GEO (accession number GSE128476).
References
Grulich, A. E., van Leeuwen, M. T., Falster, M. O. & Vajdic, C. M. Incidence of cancers in people with HIV/AIDS compared with immunosuppressed transplant recipients: a meta-analysis. Lancet 370, 59–67 (2007).
Nehal, K. S. & Bichakjian, C. K. Update on keratinocyte carcinomas. N. Engl. J. Med. 379, 363–374 (2018).
Wang, J., Aldabagh, B., Yu, J. & Arron, S. T. Role of human papillomavirus in cutaneous squamous cell carcinoma: a meta-analysis. J. Am. Acad. Dermatol. 70, 621–629 (2014).
Chockalingam, R., Downing, C. & Tyring, S. K. Cutaneous squamous cell carcinomas in organ transplant recipients. J. Clin. Med. 4, 1229–1239 (2015).
Ingle, A. et al. Novel laboratory mouse papillomavirus (MusPV) infection. Vet. Pathol. 48, 500–505 (2011).
Howley, P. M. & Pfister, H. J. Beta genus papillomaviruses and skin cancer. Virology 479–480, 290–296 (2015).
Hufbauer, M. & Akgül, B. Molecular mechanisms of human papillomavirus induced skin carcinogenesis. Viruses 9, 187 (2017).
Wang, J. W. et al. Immunologic control of Mus musculus papillomavirus type 1. PLoS Pathog. 11, e1005243 (2015).
Uberoi, A., Yoshida, S., Frazer, I. H., Pitot, H. C. & Lambert, P. F. Role of ultraviolet radiation in papillomavirus-induced disease. PLoS Pathog. 12, e1005664 (2016).
Sands, A. T., Abuin, A., Sanchez, A., Conti, C. J. & Bradley, A. High susceptibility to ultraviolet-induced carcinogenesis in mice lacking XPC. Nature 377, 162–165 (1995).
Migden, M. R. et al. PD-1 blockade with cemiplimab in advanced cutaneous squamous-cell carcinoma. N. Engl. J. Med. 379, 341–351 (2018).
Tirosh, O. et al. Expanded skin virome in DOCK8-deficient patients. Nat. Med. 24, 1815–1821 (2018).
Joh, J. et al. Molecular diagnosis of a laboratory mouse papillomavirus (MusPV). Exp. Mol. Pathol. 93, 416–421 (2012).
Jiang, R. T. et al. Spontaneous and vaccine-induced clearance of Mus musculus papillomavirus 1 infection. J. Virol. 91, e00699-17 (2017).
Chen, B. J., Cui, X., Sempowski, G. D., Liu, C. & Chao, N. J. Transfer of allogeneic CD62L− memory T cells without graft-versus-host disease. Blood 103, 1534–1541 (2004).
Abel, E. L., Angel, J. M., Kiguchi, K. & DiGiovanni, J. Multi-stage chemical carcinogenesis in mouse skin: fundamentals and applications. Nat. Protocols 4, 1350–1362 (2009).
D’Orazio, J. A. et al. Topical drug rescue strategy and skin protection based on the role of Mc1r in UV-induced tanning. Nature 443, 340–344 (2006).
Technology Planning and Management Corporation. Report on Carcinogens Background Document for Broad-Spectrum Ultraviolet (UV) Radiation and UVA, and UVB, and UVC (Technology Planning and Management Corporation, 2000).
Joh, J. et al. MmuPV1 infection and tumor development of T cell-deficient mice is prevented by passively transferred hyperimmune sera from normal congenic mice immunized with MmuPV1 virus-like particles (VLPs).Exp. Mol. Pathol. 100, 212–219 (2016).
Nelson, M. A., Futscher, B. W., Kinsella, T., Wymer, J. & Bowden, G. T. Detection of mutant Ha-ras genes in chemically initiated mouse skin epidermis before the development of benign tumors. Proc. Natl Acad. Sci. USA 89, 6398–6402 (1992).
Joh, J., Jenson, A. B., Ingle, A., Sundberg, J. P. & Ghim, S. J. Searching for the initiating site of the major capsid protein to generate virus-like particles for a novel laboratory mouse papillomavirus. Exp. Mol. Pathol. 96, 155–161 (2014).
Wang, F. et al. RNAscope: a novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J. Mol. Diagn. 14, 22–29 (2012).
Dang, C. et al. E6/E7 expression of human papillomavirus types in cutaneous squamous cell dysplasia and carcinoma in immunosuppressed organ transplant recipients. Br. J. Dermatol. 155, 129–136 (2006).
Watanabe, R. et al. Human skin is protected by four functionally and phenotypically discrete populations of resident and recirculating memory T cells. Sci. Transl. Med. 7, 279ra39 (2015).
Acknowledgements
We thank D. G. DeNardo for providing Cd4−/−Cd8−/− mice. S.D. holds a Career Award for Medical Scientists from the Burroughs Wellcome Fund and was supported by a Cancer Research Institute CLIP Grant; T.H. was supported by Shiseido; J.D.S., J.L.M., T.H., D.T.H., P.A.B., K.H.N. and S.D. were supported by grants from the Burroughs Wellcome Fund, Sidney Kimmel Foundation, Cancer Research Institute and NIH (K08AR068619, DP5OD021353 and U01CA233097); H.W.N., A.B.J., J.J. and the mouse studies were supported by anonymous donations for Oral Cancer Research, the Elsa U. Pardee Foundation, the Helmsley Trust Foundation and the James Graham Brown Cancer Center, University of Louisville.
Author information
Authors and Affiliations
Contributions
S.D. conceived the study; J.D.S., J.L.M., M.E.A., T.H., J.J. and S.D. designed the experiments; J.D.S., J.L.M., T.L., D.T.H., H.W.N., P.A.B. and K.H.N. performed the mouse experiments including tumour studies and molecular and histological analyses; M.E.A. and T.L. designed and performed RNA ISH and DNA ISH assays on human tissues; T.L. and T.H. performed immunostaining experiments on human tissue samples; T.H. performed the ex vivo CD8+ T cell-activation assay; J.D.S., J.L.M., M.E.A., T.L., T.H., A.B.J., J.J. and S.D. analysed and interpreted the data; J.D.S., J.L.M. and S.D. wrote the manuscript; M.M.A., R.M.N. and V.A.N. contributed clinical samples; R.M.N. determined the histological diagnosis of the human skin cancers.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Peer review information Nature thanks Akiko Iwasaki, Thomas Tüting and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Extended data figures and tables
Extended Data Fig. 1 Infection of back skin with MmuPV1 in wild-type and T cell-deficient mice and the effect of MmuPV1 colonization on the outcomes of chemical carcinogenesis in wild-type C57BL/6J mice.
a, Wart burden in Cd4−/−Cd8−/− mice (right) compared with the absence of warts in wild-type mice (left) after infection of back skin with MmuPV1, at 10 weeks after infection. Note the confluent pattern of wart development in the T cell-deficient mouse. b, MmuPV1-induced wart in a Cd4−/−Cd8−/− mouse, stained with H&E (left), MmuPV1 L2 RNA ISH probe (middle) and negative-control RNA ISH probe (right). c, Left, representative images of the back skin of wild-type C57BL/6J mice on the day of MmuPV1 infection and 21 days after infection. Middle, MmuPV1 L1 PCR on 20 segments of the back skin. MmuPV1 L1 PCR bands are marked by arrows; PCR amplicon size, 339 bp. PCR primers, forward: GAGCTCTTTGTTACTGTTGTC; reverse: ATCCTCTCTTTCCTTGGGC. M, molecular-weight size marker; N, negative control; P1–P3, positive controls. Right, a typical wild-type C57BL/6J mouse five weeks after infection, highlighting the absence of warts, which was the case for 100% of the mice. d, Representative macroscopic images of wild-type C57BL/6J mice that were either infected with MmuPV1 on their back skin or sham-infected, and treated with DMBA–TPA. Papillomas and invasive skin cancers are highlighted with yellow and red circles, respectively. e, Left, Representative images of the back skin of wild-type FVB mice on the day of MmuPV1 infection and 31 days after infection, and MmuPV1 L1 PCR on 20 segments of the back skin. Mice were shaved for visualization of the skin and skin tumours. Scale bars, mouse, 1 cm (a, c–e); tissue, 1 mm (b).
Extended Data Fig. 2 T cells transferred from wild-type MmuPV1-colonized immune mice to T cell-deficient mice reduce the burden of warts in mice infected with MmuPV1, but have no effect on the growth of uninfected SCC cells.
a, Schematic of the T cell-transfer experiment. The inset shows the gating strategy for flow cytometry that was used to select memory T cells. T cell-donor mice received CD45–APC intravenously (IV) three minutes before the collection of lymph nodes to label and exclude the circulating immune cells. Note that the control experiment in which mice were vaccinated with parvovirus was done in parallel with the MmuPV1 challenge, and the SCC primary tumour growth experiment was done in parallel with the infection of back skin with MmuPV1. b, Right, representative images of the warts on the back skin of mice three weeks after infection with MmuPV1. Scale bar, 1 cm. Left, flow cytometry demonstrates the presence of CD4+ and CD8+ T cells in the peripheral blood of the recipient mice, indicating successful adoptive transfer of T cells (n = 4 per group). Wt, wild type. c, Growth of subcutaneously injected DMBA–TPA-induced primary SCC tumour cells in wild-type mice (n = 9), Cd4−/−Cd8−/− mice (n = 5) and Cd4−/−Cd8−/− mice that received T cells from MmuPV1-immune donors (Cd4−/−Cd8−/− + test T cells) (n = 4). Two-tailed Mann–Whitney U test; *P < 0.05 compared with the wild-type group. Data are mean + s.d.
Extended Data Fig. 3 Evidence of MmuPV1 colonization and T cells homing to the epidermis of MmuPV1-infected mice at the completion of the chemical carcinogenesis protocol.
a, b, MmuPV1 L1 PCR on DNA isolated from the skin of wild-type C57BL/6J (B6) (a) and the skin and tumour of wild-type FVB (b) mice more than 6 months after MmuPV1 infection. MmuPV1 L1 PCR bands are highlighted by arrows; PCR amplicon size: 339 bp. Plus sign indicates positive control; minus sign, negative control. c, d, Anti-MmuPV1 seroconversion in DMBA–TPA-treated cohorts of C57BL/6J mice (c; n = 5 per group) and FVB mice (d; n = 4 per group). Two-tailed Mann–Whitney U test; *P < 0.05, **P < 0.01, NS, not significant. e, Representative images of CD3/CD45-stained skin from MmuPV1/DMBA–TPA FVB mice compared with sham/DMBA–TPA controls at the completion of the chemical carcinogenesis protocol. Arrows indicate T cells in the epidermis; dashed lines highlight the epidermal basement membrane. f, CD45+ leukocytes quantified in skin sections of MmuPV1/DMBA–TPA and sham/DMBA–TPA FVB mice across ten randomly selected HPF images of normal skin per mouse and averaged across the mice in each group (two-tailed unpaired t-test; n = 8 per group). Each dot represents the leukocyte count in one high-power image. g, h, Homing of T cells to the epidermis in MmuPV1/DMBA–TPA skin compared with sham/DMBA–TPA control skin of wild-type FVB mice. g, Representative images of CD8/CD3- and CD4/CD3-stained skin sections. Arrows indicate epidermal CD8+ TRM cells; dashed lines highlight the epidermal basement membrane. h, The ratio of epidermal CD8+ TRM and CD4+ TRM cells to total CD3+ T cells in the skin per HPF image (two-tailed unpaired t-test). T cells in up to ten randomly selected HPF images of normal skin per mouse were counted. Each dot represents one high-power image. n = 10 (MmuPV1/DMBA–TPA); n = 9 (sham/DMBA–TPA). i, Representative skin tumours from MmuPV1/DMBA–TPA and sham/DMBA–TPA wild-type FVB mice stained with keratin 6 (K6; a marker for epidermal hyperplasia) and Ki67 (a proliferation marker). Dashed lines highlight the epidermal basement membrane in the skin. j, PCR amplification of the wild-type (A) and mutant (T) region of the Hras gene in DNA of MmuPV1/DMBA–TPA and sham/DMBA–TPA tumours and skin, and untreated skin from a wild-type FVB mouse (band size, 110 bp). The A-to-T mutation in Hras codon 61 highlights DMBA–TPA-induced skin tumours in MmuPV1/DMBA–TPA and sham/DMBA–TPA wild-type FVB cohorts. k, Matched H&E and MmuPV1 L2 RNA ISH images of a wart from an MmuPV1-infected Cd4−/−Cd8−/− mouse, and a skin tumour and normal skin from an MmuPV1/DMBA–TPA wild-type mouse. Note the dense and confluent RNA ISH signals in the wart from the T cell-deficient mouse. After the completion of DMBA–TPA treatment, positive MmuPV1 RNA ISH signals are detectable in the normal skin of the wild-type mouse. The skin tumour from the same mouse lacks a MmuPV1 RNA ISH signal. Stained cells were counted blindly. Data are mean ± s.d. (c, d) or mean + s.d. (f, h). Scale bars, 100 μm (e, g, i, k).
Extended Data Fig. 4 Immunization of MmuPV1-infected SKH-1 mice with MmuPV1 vaccine protects against UV-driven carcinogenesis.
a, Top, representative images of SKH-1 mice with no evidence of disease following infection (immune) and with visible warts after back-skin infection with MmuPV1 (non-immune). Bottom, MmuPV1 L2 RNA ISH of skin from an immune and a non-immune mouse, collected three weeks after infection with MmuPV1, to detect viral activity in the normal skin and the MmuPV1-driven wart. Insets highlight the active virus in the normal skin of the immune mouse and the wart of the non-immune mouse. b, Macroscopic images of the SKH-1 mice three months after MmuPV1 back-skin infection. SKH-1 mice with spontaneous immunity to the virus (no wart) were treated once with an immunosuppressive dose of UVB (300 mJ cm−2); images of the mice three weeks after UV treatment are shown. Arrows point to the newly developed warts on the UV-treated skin. c, Histological images of a wart (yellow circle), stained with H&E and MmuPV1 RNA ISH. The magnified inset highlights MmuPV1-induced cytopathic changes in the H&E image and confluent positive MmuPV1 RNA ISH signals in the wart. d, Macroscopic images of MmuPV1-infected SKH-1 mice that continued to have warts (yellow arrows) before MmuPV1 vaccination, four weeks after vaccination and at the completion of the UV carcinogenesis protocol. The nine wart-bearing mice were treated with MmuPV1 live virus particles intraperitoneally three times over two weeks. Four weeks later, the mice underwent the UV carcinogenesis protocol. Mice with acquired antiviral immunity (n = 5) are compared with non-immune mice that have persistent warts (n = 4). e, Skin tumour burden in vaccinated immune (n = 5) and non-immune (n = 4) mice treated with the UV carcinogenesis protocol. In mice with a confluent pattern of skin tumours, counts represent the individual lesions before their coalescence. Two-tailed Mann–Whitney U test; data are mean ± s.d. f, Representative images of CD3/CD45-stained skin from MmuPV1/DMBA–UV SKH-1 mice compared with sham/DMBA–UV controls at the completion of the UV carcinogenesis protocol. Arrows indicate T cells in the epidermis; dashed lines highlight the epidermal basement membrane. g–i, Skin-infiltrating total CD45+ leukocytes (g), CD3+CD45+ T cells (h) and CD3−CD45+ leukocytes (i) quantified in CD3/CD45-stained skin sections of MmuPV1/DMBA–UV (n = 10) and sham/DMBA–UV (n = 9) SKH-1 mice across ten randomly selected HPF images of each skin sample and averaged across the mice in each group. Each dot represents one high-power image. Note the trend towards an increase in T cells and a decrease in CD3− inflammatory cells in MmuPV1/DMBA–UV skin compared with sham/DMBA–UV control. j, Representative images of CD3/CD45-stained cells in the skin tumours of MmuPV1/DMBA–UV SKH-1 mice compared with sham/DMBA–UV controls at the completion of the UV carcinogenesis protocol. Magnified insets highlight the immune cells in the tumour parenchyma. k–m, Tumour-infiltrating total CD45+ leukocytes (k), CD3+CD45+ T cells (l) and CD3−CD45+ leukocytes (m) quantified in CD3/CD45-stained sections of MmuPV1/DMBA–UV and sham/DMBA–UV SKH-1 skin tumours across HPF images of each tumour and averaged across the mice in each group (n = 12 early skin tumours per group). Each dot represents one high-power image. Stained cells were counted blindly. Two-tailed unpaired t-test; data are mean + s.d. (g–i, k–m). Scale bars, mouse, 1 cm (a, b, d); tissue, 100 μm (a, c, f, j).
Extended Data Fig. 5 CD8+ T cell immunity is required to protect MmuPV1-colonized mice from UV carcinogenesis and MmuPV1 colonization protects Xpc−/− mice from UV carcinogenesis.
a, Representative images of CD8+ T cells in the skin tumours of MmuPV1/DMBA–UV SKH-1 mice compared with sham/DMBA–UV controls at the completion of the UV carcinogenesis protocol. Magnified insets highlight T cells in the tumour parenchyma. b–d, Tumour-infiltrating CD3+ (b), CD8+ (c) and CD4+ (d) T cells quantified in CD8/CD3- and CD4/CD3-stained tumour sections of MmuPV1/DMBA–UV and sham/DMBA–UV SKH-1 mice across HPF images of each tumour and averaged across the mice in each group (n = 12 early skin tumours per group). Each dot represents one high-power image. e, f, CD4+ T cell infiltrates in MmuPV1/DMBA–UV and sham/DMBA–UV SKH-1 skin. e, Representative images of the CD4/CD3-stained skin sections. Arrows indicate epidermal CD4+ TRM cells; dashed lines highlight the epidermal basement membrane, f, Quantification of CD4+ T cells per high-power image of the skin. Ten randomly selected HPF images of skin per mouse in each group are included. Each dot represents one high-power image. n = 10 (MmuPV1/DMBA–UV); n = 9 (sham/DMBA–UV). Two-tailed unpaired t-test; data are mean + s.d. (b–d, f). g, Schematic diagram of anti-CD8 or IgG antibody treatment combined with the UV carcinogenesis protocol. Four weeks after MmuPV1 or sham(VLP) infection, mice began treatment with anti-CD8 or IgG isotype control antibodies (red arrows). A day after the first treatment with antibodies, the back skin of SKH-1 mice was treated with 50 μg DMBA once (green triangle). Seven days later, mice began UVB treatment (100 mJ cm−2) three times a week (yellow triangles). h, Flow cytometry analysis of spleen and skin of MmuPV1/DMBA–UV mice treated with anti-CD8 or IgG antibodies to evaluate the efficiency of CD8+ T cell depletion at six weeks after treatment with DMBA. The percentage of CD8+ T cells is shown on each plot. i, Skin tumour burden in MmuPV1-colonized mice treated with IgG control (MmuPV1 + IgG; n = 10) or anti-CD8 antibody (MmuPV1 + anti-CD8; n = 10), and sham(VLP)-infected mice treated with IgG control (sham(VLP) + IgG; n = 7) or anti-CD8 antibody (sham(VLP) + anti-CD8; n = 7) after DMBA–UV treatment. Two-tailed Mann–Whitney U test; *P < 0.05, NS, not significant. Data are mean ± s.d. j, Representative images of mice in the four treatment groups. Owing to the large skin tumours in MmuPV1-colonized CD8+ T cell-depleted mice, the UV carcinogenesis study was terminated at 18 weeks after DMBA treatment. k, l, Xpc−/− (XPCKO) mice were infected with MmuPV1 on their back skin (n = 15) or sham-infected (n = 13) and subjected to the UV carcinogenesis protocol. Skin tumour outcomes are shown as the time to development of the first skin tumour (k) and time to development of the first invasive skin cancer (l) (log-rank test). Note that all Xpc−/− mice in the study were immune to MmuPV1 (that is, exhibited no wart development). m, Representative images of Xpc−/− mice at the completion of the 30-week UV carcinogenesis protocol. Premalignant tumours (papillomas) and invasive skin cancers are highlighted with yellow and red circles, respectively. Mice were shaved for UV treatments and the visualization of the skin tumours. n, Representative H&E-stained histological images of a papilloma in MmuPV1/DMBA–UV and invasive skin cancer in sham/DMBA–UV Xpc−/− mice. The inset shows the cellular atypia in the sham/DMBA–UV skin cancer (scale bar, 50 μm). Stained cells were counted blindly. Scale bars, mouse, 1 cm (j, m); tissue: 100 μm (a, e, n).
Extended Data Fig. 6 Validation of β-HPV RNA ISH using a wart as a positive control and qPCR on RNA ISH-positive and -negative human samples.
a, Binding site of β-HPV RNA ISH and DNA ISH probes, shown on the HPV9 genome. The RNA ISH and DNA ISH probe against each type of β-HPV comprised a pool of 20 double-Z probes that target a region of 1,000 bases (Advanced Cell Diagnostics). b, H&E and RNA ISH staining of a wart from a 63-year-old immunosuppressed female. Note the abundance of positive signals (red dots) throughout the wart. c, Top, β-HPV RNA ISH of a skin cancer from an 87-year-old immunosuppressed female, including the stains for the positive- and negative-control probes. The detection of β-HPV by RNA ISH correlates with qPCR positivity for transcripts of HPV5 and HPV9 E6 proteins in the same skin cancer. Bottom, β-HPV RNA ISH of a sample of normal skin from an 18-year-old immunocompetent African American female. The lack of RNA ISH signal (red) in this sample correlates with undetectable transcripts of HPV5, HPV9 or HPV15 E6 proteins in qPCR of the same sample. qPCR products were visualized using gel electrophoresis. PCR band sizes: HPV5 E6, 100 bp; HPV9 E6, 66 bp, HPV15 E6, 78 bp; keratin 14, 109 bp. Scale bars, 100 μm (b, c).
Extended Data Fig. 7 Immunosuppressed patients have greater β-HPV viral activity in their skin lesions compared to immunocompetent patients.
a, β-HPV RNA ISH signal counts in skin cancer cells from immunosuppressed (n = 38) and immunocompetent (n = 32) patients. b, Clinical image of a skin cancer surgical site showing the skin cancer (red arrow), its adjacent normal skin (green arrow) and the normal skin away from the cancer site (blue arrow). c, Quantification of β-HPV RNA ISH signals in high-power images across the immunosuppressed lesions, immunocompetent lesions and normal facial skin away from a cancer site. Skin lesions include β-HPV RNA ISH signal counts from skin cancer (red dots) and the adjacent normal skin (green dots). Thirty samples of normal facial skin (blue dots) from immunocompetent patients are included (18 males and 12 females; average age 71; age range 39–94). d, Representative low- and high-magnification images of β-HPV RNA ISH-stained normal skin samples from immunosuppressed and immunocompetent patients. Note the density and size of the apparent RNA ISH signals in basal-layer keratinocytes of an immunosuppressed patient. e, Density of β-HPV RNA ISH signals in basal-layer keratinocytes, quantified across 38 immunosuppressed and 31 immunocompetent skin samples. f, Right, β-HPV DNA ISH to detect β-HPV viral load in the skin. Compared to β-HPV RNA ISH, which marks viral transcripts, β-HPV DNA ISH detects viral load at a subcellular resolution in skin keratinocytes. Note the higher level of viral DNA ISH signals compared with RNA ISH (left), and the localization of the signals in the nucleus and cytoplasm of the keratinocytes. Two-tailed unpaired t-test; data are mean + s.d. (a, c, e). Scale bars, 50 μm (d, f).
Extended Data Fig. 8 β-HPV viral load is markedly reduced in skin cancer cells compared to their adjacent normal skin in immunocompetent patients.
a, Representative DNA ISH of a wart, hypertrophic actinic keratosis arising in association with a wart (HAK in verruca), SCC in immunosuppressed patients and an SCC in an immunocompetent patient. Scale bars, 100 μm. b, c, Quantification of β-HPV DNA ISH signals in paired samples of skin cancer and the adjacent normal skin from immunosuppressed patients (b; n = 10) and immunocompetent patients (c; n = 10) (two-tailed Wilcoxon matched-pairs signed-rank test).
Extended Data Fig. 9 Significantly fewer T and TRM cells infiltrate skin cancer and the adjacent normal skin in immunosuppressed compared to immunocompetent patients.
a, Representative images of CD3/CD103-stained SCC from immunosuppressed and immunocompetent patients (the same cancers are shown for β-HPV RNA ISH and DNA ISH stains in Fig. 3a and Extended Data Fig. 8a). Magnified insets highlight CD103+ TRM cells in the cancer and adjacent normal skin. Scale bars, 100 μm. b, c, CD3/CD8/CD103-stained sections of skin cancer were used to quantify tumour-infiltrating CD3+ T, CD103+CD3+ TRM, CD8+ T and CD103+CD8+ TRM cells infiltrating the skin cancer parenchyma (b), and CD3+ T, CD103+CD3+ TRM, CD8+ T and CD103+CD8+ TRM cells in the adjacent normal skin of immunosuppressed (S) versus immunocompetent (C) patients. Note that most T cells in the normal skin reside in the dermis. Stained cells were counted blindly in ten randomly selected HPF images of skin cancer and adjacent normal skin from each tissue specimen and averaged across the samples in each group; 37 immunosuppressed and 32 immunocompetent samples of skin cancer are included (skin cancer characteristics are listed in Supplementary Table 2). Each dot represents the average of the T cell counts in the high-power images from each sample. Two-tailed unpaired t-test; data are mean + s.d. d, Cytotoxic degranulation of CD8+ T lymphocytes after exposure to β-HPV peptides. T cells isolated from the normal facial skin of adults were exposed to β-HPV E7 peptides (far left), HPV16 E7 peptides (middle left), PMA/ionomycin (positive control; middle right) and medium (negative control; far right). Representative flow cytometry plots are shown. The percentage of CD107a+CD8+ T cells is shown on each plot. Data represent two independent sets of experiments with similar results.
Extended Data Fig. 10 DAMP molecules are upregulated during the development of warts and skin cancer.
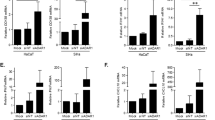
a, Principle component analysis (PCA) of gene-expression profiles obtained from MmuPV1-induced warts (n = 4; blue triangles), MmuPV1-infected skin (n = 4; pink squares) or sham-infected skin (n = 4; grey circles), and MmuPV1-infected tumours (n = 4; red squares) or sham-infected tumours (n = 4; black circles) of SKH-1 mice. Note that DMBA–UV-induced skin tumours from MmuPV1-infected mice are indistinguishable from skin tumours from sham-infected mice, whereas both have very distinct transcriptional profiles compared with MmuPV1-driven warts. b, c, Volcano plots of differentially expressed genes in MmuPV1- versus sham-infected skin (b; n = 4 per group), and skin tumours and warts (n = 12) versus MmuPV1- and sham-infected skin (c; n = 8). Gm5416 is also known as Csta3. P values were calculated using the DESeq2 R package (v.2 1.6.3), and the resulting P values were adjusted using the Benjamini–Hochberg method for controlling the false discovery rate. The 20 genes that were upregulated in skin tumours and warts compared with MmuPV1- and sham-infected skin are shown in the table on the left. d–f, Analysis of the expression of immune genes in human skin lesions on the basis of the mouse RNA-seq data. d, Representative macroscopic and H&E-stained histological images of SCC, wart, seborrheic keratosis (SK) and unaffected human skin. Scale bar, 500 μm. e, Relative gene expression in SCCs (n = 7) and warts (n = 5) compared with normal skin (n = 8). f, Normalized relative gene expression in SCCs (n = 7), warts (n = 5) and seborrheic keratosis (n = 5) compared for several DAMP genes. Average relative gene expression in the normal skin was used for normalization. GAPDH is used as the reference gene. Two-tailed Mann–Whitney U test; *P < 0.05, **P < 0.01, NS, not significant; data are mean + s.d. (e, f).
Supplementary information
Supplementary Tables
This file contains: Supplementary Table 1 β-HPV and MmuPV1 probes used for RNAish; Supplementary Table 2 Demographics of skin cancers used in β-HPV RNAish assays; Table 3 Amino acid sequences of β-HPV and HPV16 E7 peptides used for in vitro T cell stimulation assay and Supplementary Table 4 Antibodies (a) and primers (b) used for tissue analysis
Source data
Rights and permissions
About this article

Cite this article
Strickley, J.D., Messerschmidt, J.L., Awad, M.E. et al. Immunity to commensal papillomaviruses protects against skin cancer. Nature 575, 519–522 (2019). https://doi.org/10.1038/s41586-019-1719-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-019-1719-9
This article is cited by
-
Advances in cutaneous squamous cell carcinoma
Nature Reviews Cancer (2023)
-
Physicochemical investigations of clouding development and physicochemical properties of Triton X-100 and levofloxacin hemihydrate mixture: influence of sodium salts composition
Colloid and Polymer Science (2023)
-
Therapeutic Cancer Vaccines for Nonmelanoma Skin Cancer
Current Treatment Options in Oncology (2023)
-
Human papillomaviruses: diversity, infection and host interactions
Nature Reviews Microbiology (2022)
-
The precursors of CD8+ tissue resident memory T cells: from lymphoid organs to infected tissues
Nature Reviews Immunology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
