
Abstract
SIRT6 acts as a longevity protein in rodents1,2. However, its biological function in primates remains largely unknown. Here we generate a SIRT6-null cynomolgus monkey (Macaca fascicularis) model using a CRISPR–Cas9-based approach. SIRT6-deficient monkeys die hours after birth and exhibit severe prenatal developmental retardation. SIRT6 loss delays neuronal differentiation by transcriptionally activating the long non-coding RNA H19 (a developmental repressor), and we were able to recapitulate this process in a human neural progenitor cell differentiation system. SIRT6 deficiency results in histone hyperacetylation at the imprinting control region of H19, CTCF recruitment and upregulation of H19. Our results suggest that SIRT6 is involved in regulating development in non-human primates, and may provide mechanistic insight into human perinatal lethality syndrome.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
References
Houtkooper, R. H., Pirinen, E. & Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 13, 225–238 (2012).
Kanfi, Y. et al. The sirtuin SIRT6 regulates lifespan in male mice. Nature 483, 218–221 (2012).
Niu, Y. et al. Generation of gene-modified cynomolgus monkey via Cas9/RNA-mediated gene targeting in one-cell embryos. Cell 156, 836–843 (2014).
Wan, H. et al. One-step generation of p53 gene biallelic mutant cynomolgus monkey via the CRISPR/Cas system. Cell Res. 25, 258–261 (2015).
Zuo, E. et al. One-step generation of complete gene knockout mice and monkeys by CRISPR/Cas9-mediated gene editing with multiple sgRNAs. Cell Res. 27, 933–945 (2017).
Chalkiadaki, A. & Guarente, L. Sirtuins mediate mammalian metabolic responses to nutrient availability. Nat. Rev. Endocrinol. 8, 287–296 (2012).
Kugel, S. et al. SIRT6 suppresses pancreatic cancer through control of Lin28b. Cell 165, 1401–1415 (2016)
Schiaffino, S. & Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 91, 1447–1531 (2011).
Kugel, S. & Mostoslavsky, R. Chromatin and beyond: the multitasking roles for SIRT6. Trends Biochem. Sci. 39, 72–81 (2014).
Giblin, W., Skinner, M. E. & Lombard, D. B. Sirtuins: guardians of mammalian healthspan. Trends Genet. 30, 271–286 (2014).
Kernohan, K. D. et al. ATRX partners with cohesion and MeCP3 and contributes to developmental silencing of imprinted genes in the brain. Dev. Brain 18, 191−202 (2010).
Gabory, A., Jammes, H. & Dandolo, L. The H19 locus: role of an imprinted non-coding RNA in growth and development. BioEssays 32, 473–480 (2010).
Wakeling, E. L. et al. Diagnosis and management of Silver–Russell syndrome: first international consensus statement. Nat. Rev. Endocrinol. 13, 105–124 (2016).
Pan, H. et al. SIRT6 safeguards human mesenchymal stem cells from oxidative stress by coactivating NRF2. Cell Res. 26, 190–205 (2016).
Liu, G. H. et al. Progressive degeneration of human neural stem cells caused by pathogenic LRRK2. Nature 491, 603–607 (2012).
Bell, A. C. & Felsenfeld, G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature 405, 482–485 (2000).
Kurukuti, S. et al. CTCF binding at the H19 imprinting control region mediates maternally inherited higher-order chromatin conformation to restrict enhancer access to Igf2. Proc. Natl Acad. Sci. USA 103, 10684–10689 (2006).
Fedoriw, A. M., Stein, P., Svoboda, P., Schultz, R. M. & Bartolomei, M. S. Transgenic RNAi reveals essential function for CTCF in H19 gene imprinting. Science 303, 238–240 (2004).
Schwer, B. et al. Neural sirtuin 6 (Sirt6) ablation attenuates somatic growth and causes obesity. Proc. Natl Acad. Sci. USA 107, 21790–21794 (2010).
Ferrer, C. M. et al. An inactivating mutation in the histone deacetylase SIRT6 causes human perinatal lethality. Genes Dev. 32, 373–388 (2018).
Yang, S. et al. Effects of rhFSH dose on ovarian follicular response, oocyte recovery and embryo development in rhesus monkeys. Theriogenology 67, 1194–1201 (2007).
Niu, Y. et al. Transgenic rhesus monkeys produced by gene transfer into early-cleavage-stage embryos using a simian immunodeficiency virus-based vector. Proc. Natl Acad. Sci. USA 107, 17663–17667 (2010).
Wang, H. et al. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 153, 910–918 (2013).
Pyzocha, N. K., Ran, F. A., Hsu, P. D. & Zhang, F. RNA-guided genome editing of mammalian cells. Methods Mol. Biol. 1114, 269–277 (2014).
Zhang, W. et al. A Werner syndrome stem cell model unveils heterochromatin alterations as a driver of human aging. Science 348, 1160–1163 (2015).
Robinson, J. T. et al. Integrative genomics viewer. Nat. Biotechnol. 29, 24–26 (2011).
Veres, A. et al. Low incidence of off-target mutations in individual CRISPR-Cas9 and TALEN targeted human stem cell clones detected by whole-genome sequencing. Cell Stem Cell 15, 27–30 (2014)
Bae, S., Park, J. & Kim, J. S. Cas-OFFinder: a fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 30, 1473–1475 (2014).
Mao, P. et al. Homologous recombination-dependent repair of telomeric DSBs in proliferating human cells. Nat. Commun. 7, 12154 (2016).
Kim, D., Langmead, B. & Salzberg, S. L. HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12, 357–360 (2015).
Trapnell, C. et al. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 28, 511–515 (2010).
Huang, D. W., Sherman, B. T. & Lempicki, R. A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protocols 4, 44–57 (2009).
Huang, D. W., Sherman, B. T. & Lempicki, R. A. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 37, 1–13 (2009).
Trapnell, C. et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protocols 7, 562–578 (2012).
Ocampo, A. et al. In vivo amelioration of age-associated hallmarks by partial reprogramming. Cell 167, 1719–1733.e12 (2016).
Chalkiadaki, A., Igarashi, M., Nasamu, A. S., Knezevic, J. & Guarente, L. Muscle-specific SIRT1 gain-of-function increases slow-twitch fibers and ameliorates pathophysiology in a mouse model of duchenne muscular dystrophy. PLoS Genet. 10, e1004490 (2014).
Nativio, R. et al. Cohesin is required for higher-order chromatin conformation at the imprinted IGF2-H19 locus. PLoS Genet. 5, e1000739 (2009).
Kim, H. S. et al. The hsSsu72 phosphatase is a cohesin-binding protein that regulates the resolution of sister chromatid arm cohesion. EMBO J. 29, 3544–3557 (2010).
Acknowledgements
The authors acknowledge X. Wang, Y. Fu, L. Zhao, J. An and L. Wei for brain tissue analysis, S. Duan for western blotting, L. Sun from the Center for Biological Imaging for electron microscopy, R. Bai and P. Wang for technical assistance, L. Bai for administrative assistance, X. Liu for image processing using Vectra Automated Quantitative Pathology Imaging System, and P. Zhang for fluorescence in situ hybridization analysis. This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA16010100), the National Key Research and Development Program of China (2015CB964800, 2017YFA0103304, 2017YFA0102802, 2014CB910503, 2014CB964600, 2018YFA0107203, 2016YFA0101403), the National Natural Science Foundation of China (91749202, 31471394, 31671429, 91749123, 81625009, 81330008, 81371342, 81471414, 81422017, 81601233, 81671377, 31601109, 31601158, 81771515, 81701388, 31571533, 31621004, 81422014), Program of Beijing Municipal Science and Technology Commission (Z181100001818002, Z151100003915072), Key Research Program of the Chinese Academy of Sciences (ZDRW-ZS-2017-5, KJZDEW-TZ-L05), Beijing Municipal Commission of Health and Family Planning (PXM2018_026283_000002) and Advanced Innovation Center for Human Brain Protection (117212).
Reviewer information
Nature thanks H. Cohen, H. Yang and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
G.-H.L., W.L., B.H. and Q.Z. conceptualized the work and supervised overall experiments; W.Z. performed the phenotypic and mechanistic analyses; H.W. generated gene-edited monkeys; G.F. performed bioinformatics analyses; J.Q. guided cell culture, differentiation and data analysis; Y.J. performed molecular experiments; R.R. performed brain biopsy; Z.C. and S.W. performed brain immunochemistry; Y.Y. and Z.W. performed muscle analysis; Y.Z. performed fluorescence in situ hybridization; L.Z. designed sgRNAs; Z.L. and L.Z. performed genotyping; J.W. performed plasmid construction; G.-H.L., W.L., B.H., Q.Z., W.Z., H.W., G.F. and J.Q. performed data analysis and wrote the manuscript. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
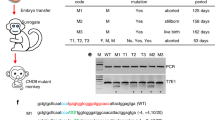
Extended Data Fig. 1 Generation and genotyping of SIRT6−/− monkeys.
a, Schematic showing the optimization of the CRISPR–Cas9-based targeting system for SIRT6 in monkeys. b, T7 endonuclease I assay for determining the indel rates of the six sgRNAs that target the monkey SIRT6 gene. The white arrows indicate the cleavage bands. M, marker. c, Schematic of the mosaic mutations in the SIRT6 loci. The red blocks identify the nucleotide sequences deleted in the SIRT6 gene. E, exon. d, Types of deletions identified by Sanger sequencing of the PCR-amplified SIRT6 cDNA from organs of the three SIRT6−/− monkeys. e, PCR of the SIRT6 gene from the wild-type and SIRT6−/− monkey genomes using primer pairs P1 and P2 or P3 and P4 (Supplementary Table 4). PCRs of the p53 locus were used as a control. f, Whole-genome sequencing data did not reveal mutations in the potential off-target sites predicted on the basis of sequences similar to the sgRNA sequence. The presumed PAM sequences and genomic locations of these sites are shown. g, Scatter plots showing the coverage scores for targeting sites within the SIRT6 gene and predicted potential off-target sites for the two sgRNAs. Three hundred and seventy-four predicted potential off-target sites and two SIRT6 sgRNA-target sequences located in the CDS regions are presented. The methods for calculating the coverage score are described in Methods. h, Venn diagram showing the lack of consistency of mutations in CDS regions detected by whole-genome sequencing among all three SIRT6−/− monkey samples. All the possible off-target sites were identified by whole-genome sequencing regardless of the similarity to sgRNA sequences, as described in Methods.
Extended Data Fig. 2 Characterization of SIRT6−/− monkeys.
a, Western blots showing the absence of the SIRT6 protein and the alterations in H3K56ac levels in the kidney, lung, liver and skin of SIRT6−/− monkeys. b, Immunohistochemical staining for H3K56ac in tissues from SIRT6−/− and wild-type monkeys. The red dashed lines show the upper boundaries of the cortices. Scale bar, 100 μm. n ≥ 54 glomeruli, kidney; n = 18 images, lung; n = 18 images, liver; n = 18 images, skin; n = 9 images, brain. Data are mean ± s.e.m.; P values were determined by one-way ANOVA followed by Holm–Sidak’s multiple comparisons test. For uncropped gels, see Supplementary Fig. 1.
Extended Data Fig. 3 SIRT6 deficiency does not cause genomic and epigenomic instabilities in newborn monkeys.
a, b, The relative telomere lengths of chromosomes from the monkey brain and liver were measured by qPCR. n = 3 monkeys. c, Whole-genome sequencing of copy number variations in brain samples from SIRT6+/+ and SIRT6−/− monkeys. Each point represents a 500-kb genomic region of each chromosome. d, The distribution of different types of repetitive sequences in the monkey genome. Each type of repeat is marked in the same colour: LINE type 1 (LINE-1), SINEs, LTRs (including ERV1, ERV2, EVR3 and other types of LTR), rRNAs and other types of repeat (such as satellite, small nuclear RNA and ‘other’, as annotated in Repbase). The distributions of repeat reads were not significantly different in the wild-type and SIRT6−/− monkeys, according to the two sided Wilcoxon signed-rank paired test. e, Whole-genome sequencing of SNVs using cortical samples from SIRT6+/+ and SIRT6−/− monkeys. Sites with a heterozygosity percentage ranging between 0% and 30% were considered as SNV sites, whereas sites with a heterozygosity of >30% were considered single nucleotide polymorphisms. f, Schematic and morphology of primary mesenchymal stem cells isolated from monkey bone marrow. Scale bar, 50 μm. g, Western blots showing the absence of the SIRT6 protein and the alterations in H3K56ac levels in primary mesenchymal stem cells from SIRT6−/− monkeys. h, Karyotyping analysis of primary mesenchymal stem cells. i, Metaphase spread and fluorescence in situ hybridization showing telomeres in primary mesenchymal stem cells from wild-type and SIRT6−/− monkeys (left). Quantification of the numbers of fluorescence in situ hybridization-labelled telomeres in each nucleus (right). n ≥ 21 metaphases per monkey. Scale bar, 25 μm. j, The relative telomere lengths of chromosomes from primary monkey mesenchymal stem cells were measured by qPCR. n = 6 qRT–PCR repeats for each monkey. k, Western blots of heterochromatin-related proteins in the cortex, muscle and liver of the wild-type and SIRT6−/− monkeys. Data are mean ± s.e.m., P values were determined by two-sided Student’s t-test (a, b) or one-way ANOVA followed by Holm–Sidak’s multiple comparisons test (i, j). For uncropped gels, see Supplementary Fig. 1.
Extended Data Fig. 4 SIRT6 deficiency resulted in a pan-tissue developmental delay.
a, Micro-computed tomography of the hand (left), distal femoral trabecular (middle) bones and relative bone density (right) of wild-type and SIRT6−/− newborn, and wild-type 2- and 4-month-old fetal, monkeys. Arrows point to the missing bone connections in the hands of the SIRT6−/− monkeys and wild-type fetuses. Scale bars, 2.5 mm. n = 3 monkeys. b, Haematoxylin and eosin staining of the superficial intestinal epithelia from a wild-type newborn and 4-month-old fetus, and a SIRT6−/− newborn monkey. Arrows point to regions of immature intestine epithelium in the tissues of the fetus and the SIRT6−/− monkey. Scale bar, 75 μm. n = 3 images (Vectra automated quantitative pathology imaging system) per monkey. c, Haematoxylin and eosin staining shows the absence of subcutaneous fat in a newborn SIRT6−/− monkey and wild-type 4-month-old fetus. In the tissue of the wild-type infant, the subcutaneous fat is circled with a dashed black line. Scale bar, 100 μm. n = 3 images (Vectra automated quantitative pathology imaging system) per monkey. d–f, Haematoxylin and eosin staining of the indicated tissues in newborn SIRT6−/− monkeys and 2-month-old fetal, 4-month-old fetal and newborn wild-type monkeys. d, Kidney, n ≥ 80 glomeruli per monkey; e, liver, n ≥ 9 images per monkey; f, lung, n = 9 images per monkey. In the bar graph, the grey dashed lines represent the average value of the wild-type fetuses. Scale bar, 100 μm. Data are mean ± s.e.m.; P values were determined by two-sided Student’s t-test (a), one-way ANOVA followed by Holm–Sidak’s multiple comparisons test (b–f).
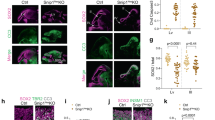
Extended Data Fig. 5 SIRT6 deficiency resulted in a delay in brain development (sagittal sections).
a, Representative photographs (left) and quantification (right) showing the sizes of the brains from wild-type newborn, wild-type 2-month-old and 4-month-old fetuses, and SIRT6−/− monkeys. The cerebella are depicted in blue; the temporal and occipital lobes are depicted in red; and the frontal lobes are depicted in green. n = 3 monkeys. Scale bar, 0.5 cm. b, Representative photographs of sagittal sections from the indicated monkey. Scale bar, 0.5 cm. c, Left, images of immunofluorescence staining for H3K56ac and the neuronal nuclear antigen (NeuN) in the cortices of newborn wild-type and SIRT6−/− monkeys. Right, cell density calculation. Numbers on the images represent percentages of H3K56ac-positive cells in the cortices. The white dashed lines identify the boundaries of the cerebral cortex. n = 4 images per monkey. d, Left, images of immunofluorescence staining for SATB2 and BRN2 in the cortices of the newborn SIRT6−/− and wild-type monkeys. Right, the proportion of SATB2+ cells in the cortical layers were quantified in brain sections. The white dashed lines identify the boundaries of the cortical layers. n = 16 images per monkey. e, Images of immunofluorescence staining for HOPX in SIRT6−/− and wild-type cortical tissue sections. f, Images of immunostaining (left) for PCP2, a marker of cerebellar Purkinje neurons, and quantification of the thickness of the molecular layer of the cerebellum (right). PCP2+ cells were not detected in the cerebellum of SIRT6−/− monkeys. The white dashed lines identify the boundaries of the cerebellum. Green lines indicate the thickness of the molecular layer of the cerebellum. Scale bar, 50 μm (c–e). n = 6 images per monkey. Data are mean ± s.e.m.; P values were determined by two-sided Student’s t-test.
Extended Data Fig. 6 SIRT6 deficiency resulted in a delay in brain development (coronal sections).
a, Schematic of the brain regions analysed in our study. b, Representative photographs of coronal sections from the indicated monkeys. Scale bar, 0.5 cm. c, Haematoxylin and eosin staining in coronal sections from the brains of the indicated monkeys. Results shown are representative of three independent experiments. Scale bar, 0.25 cm. d, Images of immunostaining for the neuronal nuclear antigen (NeuN) in the cortices of SIRT6−/− and wild-type newborn, and wild-type 2-month-old and 4-month-old fetal, monkeys. Results shown are representative of three independent experiments. Scale bar, 150 μm. e, Left, images of immunostaining for HOPX in SIRT6−/− and wild-type cortical tissue sections. Right, bar graph showing the diameter of the HOPX+ cells. The red arrows identify HOPX+ cells. n = 141. Scale bars, 100 μm. f, Left, images of PAX6 immunostaining in the hippocampi of SIRT6−/− and wild-type newborn monkeys, and wild-type fetuses. Right, bar graph showing the relative percentages of PAX6+ cells in brain sections. n = 3 slices per monkey. Scale bar, 100 μm. g, Left, haematoxylin and eosin staining of the cerebellum from the wild-type and SIRT6−/− newborn, and wild-type 2-month-old and 4-month-old fetal, monkeys. Right, the relative thicknesses of the molecular layers. The bottom panels show higher magnification images of the boxed areas in the corresponding top panels. Green lines indicate the thickness of the molecular layer of the cerebellum. Green arrows identify Purkinje neurons. n ≥ 156 images per genotype. Image was taken at about every 4 Purkinje neurons in wild-type newborn monkeys. Scale bar, 50 μm. h, Images of immunostaining (left) and quantification of the size (right) of PCP2+ cerebellar Purkinje neurons. PCP2+ cells were very scarce (only 14 were found) in the tissue(s) from the SIRT6−/− monkeys, n = 114 PCP2+ cells in wild-type monkeys. Arrows identify PCP2+ neurons. Scale bar, 50 μm. Red dashed lines identify the boundaries of the cerebral cortex. Grey dashed lines represent the average values for the 2-month-old and 4-month-old fetuses. White box indicates enlarged area. a.u., arbitrary units. Horizontal lines show the average values for each group (e, g, h). Data are mean ± s.e.m. (f); P values were determined by two-sided Student’s t-test or one-way ANOVA followed by Holm–Sidak’s multiple comparisons test.
Extended Data Fig. 7 SIRT6 deficiency resulted in a delay in muscle development.
a, Representative images of haematoxylin and eosin staining in the gastrocnemius and soleus muscles from the indicated monkeys, and quantification of the fibre diameter (n ≥ 60 fibres). Scale bar, 10 μm. b, Images (left) and quantification (right) of myosin ATPase staining at pH 4.4 and 10.6 in cross-sections of the gastrocnemius and soleus muscle tissue. n = 6 slices per monkey. Scale bar, 20 μm. c, Left, images of immunofluorescence staining for the slow-twitch marker myosin-slow and sarcolemma-bound protein dystrophin in the gastrocnemius and soleus muscle. Scale bar, 50 μm. Right, distributions of the types of muscle fibres were calculated as a percentage of myosin-slow+ fibres. n = 3 slices per monkey. d, Electron microscopy images of mitochondrial morphology showing dense mitochondrial cristae predominantly in infant wild-type monkeys. Each yellow dashed circle identifies a single mitochondrion. Grey dashed lines represent the average values for the 2-month-old and 4-month-old fetuses. Scale bar, 100 nm. n = 3 slices per monkey, ≥ 30 mitochondria per slices. Data are mean ± s.e.m.; P values were determined by one-way ANOVA followed by Holm–Sidak’s multiple comparisons test.
Extended Data Fig. 8 SIRT6−/− monkeys exhibit embryonic transcriptome features.
a, Principal component analyses of brain and muscle tissues from SIRT6+/+ newborn and fetal monkeys and SIRT6−/− monkeys, based on development-related genes as described in Methods. n = 2 independent experiments per tissue per monkey. b, Hierarchical clustering analysis based upon global DNA methylation of brain tissues from SIRT6+/+ newborn and fetal monkeys and SIRT6−/− monkeys, as described in Methods. c, An unsupervised t-SNE cluster analysis of the SIRT6−/− (red) and wild-type (grey) samples. n = 2 independent experiments per tissue per monkey. d, The Gene Ontology terms for genes in SIRT6−/− newborn monkeys and wild-type fetuses that were differentially expressed in the same manner as in the wild-type infants. e, Venn diagrams showing the overlap among differentially upregulated and downregulated genes (fold change (SIRT6−/−/SIRT6+/+) > 2 or < 0.5, false-discovery-rate-adjusted P < 0.05) in brains and muscles of the SIRT6−/− and wild-type infants, and the 2-month-old and 4-month-old wild-type fetuses and the wild-type infants. n = 2 independent experiments per tissue per monkey. f, Levels of H19 in muscle and brain tissues from the wild-type newborn (no. 2 and no. 3) and fetal monkeys and SIRT6−/− monkeys, as measured by qRT–PCR. For d, e, the average transcript levels in the tissues from SIRT6+/+ monkeys no. 2 and no. 3 were normalized to 1, and the relative expression level of each gene was coloured as shown in the bottom panel.
Extended Data Fig. 9 Deacetylation of H3K56 by SIRT6 regulates H19 expression.
a, Schematic showing the method used to generate SIRT6+/+ and SIRT6−/− NPCs, which were differentiated into post-mitotic neurons. b, Images of immunofluorescence staining of SIRT6+/+ and SIRT6−/− NPCs. Scale bar, 20 μm. c, Karyotyping analysis of NPCs. d, Measurements of the percentages of early passage NPCs at each cell cycle stage, n = 3 wells per condition. e, Western blots of the indicated proteins in the SIRT6+/+ and SIRT6−/− NPCs and in their neuronal differentiation products (neuronal dif.). f, Immunofluorescence staining for H3K56ac levels in SIRT6+/+ and SIRT6−/− neuronal differentiation products. Scale bar, 50 μm. g, Western blots showing H3K56ac levels in protein extracts from the cortices and muscle from the infant and 2-month-old and 4-month-old fetal SIRT6+/+ monkeys, and from SIRT6−/− newborn monkeys. h, ChIP–qPCR showing the enrichment of CTCF at the imprinting control region of H19 in the genomes of differentiated neuronal cells. n = 5 wells per condition. i, qPCR assessment of the H19 levels in neuronal derivatives transduced with the indicated lentiviral vector. n = 4 wells per condition. j, Images of immunofluorescence staining (left) and statistical analyses (right, bar graph) of the neuronal cells that overexpress H3(K56Q). n = 3 wells per condition. Scale bar, 50 μm. k, Western blots of levels of H3K56ac in protein extracts from SIRT6−/− NPCs that overexpress luciferase (control), inactive SIRT6 (SIRT6-HY) and wild-type SIRT6. l, Images of immunostaining for MAP2 and TuJ1 in the neuronal derivatives of wild-type NPCs that overexpress H19 (see Fig. 4e). Scale bar, 50 μm. m, Images of immunofluorescence staining (left) and statistical analyses (right, bar graph) of the neuronal cells with H19 shRNA vector (n = 4 wells per condition). Scale bar, 50 μm. Data are mean ± s.e.m., P values were determined by two-sided Student’s t-test (i, j and m) or one unpaired t-test followed by multiple comparisons using the Holm–Sidak method (d, i). For uncropped gels, see Supplementary Fig. 1.
Extended Data Fig. 10 A model of the mechanism by which SIRT6 regulates prenatal development in monkeys.
Top, SIRT6-null monkeys die soon after birth and exhibit retarded development that resembles that of 2- to 4-month-old fetuses. Notably, SIRT6 deficiency delays neuronal development, which was recapitulated in an in vitro study examining the differentiation of wild-type and SIRT6−/− human NPCs. Bottom, during neuronal differentiation SIRT6 is recruited to the imprinting control region of H19, where it deacetylates H3K56ac and thereby prevents CTCF-mediated transcription of the developmental repressor long non-coding RNA H19. SIRT6 depletion promotes the expression of H19 and arrests neuronal differentiation.
Supplementary information
Supplementary Figure 1
This file contains the uncropped gels for the western blots, including Source data for Fig. 4c. Pulldown of endogenous SIRT6 using an anti-SIRT6 antibody (CST). The SIRT6 band (red) resides at a MW between 50 and 37 KDa.
Supplementary Table 1
This file contains: Sheet 1 Summary of monkey cohorts used in the present study; Sheet 2 The results of Sanger sequencing for the targeted SIRT6 sites from cells that were individually transfected with one of the different sgRNAs. The PAM sites are shown in red uppercase letters and the insertions are shown in red lowercase letters. The numbers of inserted (+) and deleted nucleotides (Δ) are presented to the right of each read. The fraction on the far right of each read indicates the number of mutant sequenced reads (numerator) divided by the total number of sequenced reads (denominator) for each allele; Sheet 3 The results of Sanger sequencing for monkey embryos injected with Cas9 and SIRT6-targeting sgRNAs; Sheet 4 Genotyping analysis of monkey embryos injected with Cas9 and SIRT6-targeting sgRNAs. Upper panel, pre-implantation development of embryos injected with Cas9 and SIRT6 sgRNAs; lower panel, mutation rate and gender ratio in SIRT6 mutant monkey embryos; Sheet 5 Types of deletions in the SIRT6 gene identified by Sanger sequencing after PCR amplification of cDNAs from the organs of the three SIRT6–/– monkeys; Sheet 6 Organ mass (units: grams) in monkeys. P values were determined by two-sided Student’s t-test.
Supplementary Table 2
Evaluating potential off-target sites.
Supplementary Table 3
A list of differentially expressed genes.
Supplementary Table 4
A list of primer sequences.
Rights and permissions
About this article

Cite this article
Zhang, W., Wan, H., Feng, G. et al. SIRT6 deficiency results in developmental retardation in cynomolgus monkeys. Nature 560, 661–665 (2018). https://doi.org/10.1038/s41586-018-0437-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0437-z
This article is cited by
-
Analysis of the aging-related biomarker in a nonhuman primate model using multilayer omics
BMC Genomics (2024)
-
Exosomes derived from adipose tissue-derived stem cells alleviated H2O2-induced oxidative stress and endothelial-to-mesenchymal transition in human umbilical vein endothelial cells by inhibition of the mir-486-3p/Sirt6/Smad signaling pathway
Cell Biology and Toxicology (2024)
-
SIRT6 Improves Hippocampal Neurogenesis Following Prolonged Sleep Deprivation Through Modulating Energy Metabolism in Developing rats
Molecular Neurobiology (2024)
-
Epigenetics and environmental health
Frontiers of Medicine (2024)
-
CATI: an efficient gene integration method for rodent and primate embryos by MMEJ suppression
Genome Biology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}