
Abstract
Fibroblast growth factor 21 (FGF21) is a stress-inducible hormone that has important roles in regulating energy balance and glucose and lipid homeostasis through a heterodimeric receptor complex comprising FGF receptor 1 (FGFR1) and β-klotho. Administration of FGF21 to rodents or non-human primates causes considerable pharmacological benefits on a cluster of obesity-related metabolic complications, including a reduction in fat mass and alleviation of hyperglycaemia, insulin resistance, dyslipidaemia, cardiovascular disorders and non-alcoholic steatohepatitis (NASH). However, native FGF21 is unsuitable for clinical use owing to poor pharmacokinetic and biophysical properties. A large number of long-acting FGF21 analogues and agonistic monoclonal antibodies for the FGFR1–β-klotho receptor complexes have been developed. Several FGF21 analogues and mimetics have progressed to early phases of clinical trials in patients with obesity, type 2 diabetes mellitus and NASH. In these trials, the primary end points of glycaemic control have not been met, whereas substantial improvements were observed in dyslipidaemia, hepatic fat fractions and serum markers of liver fibrosis in patients with NASH. The complexity and divergence in pharmacology and pathophysiology of FGF21, interspecies variations in FGF21 biology, the possible existence of obesity-related FGF21 resistance and endogenous FGF21 inactivation enzymes represent major obstacles to clinical implementation of FGF21-based pharmacotherapies for metabolic diseases.
Key points
-
The discovery of fibroblast growth factor 21 (FGF21) as a potent agent for treatment of obesity and type 2 diabetes mellitus in animals has inspired the development of engineered FGF21 analogues and mimetics with improved potency and pharmacokinetic profiles.
-
The multiple metabolic effects of FGF21 are mediated by both its central and peripheral actions, and by its fine-tuning of inter-organ metabolic crosstalk.
-
In individuals with obesity and type 2 diabetes mellitus, FGF21 analogues alleviate dyslipidaemia and increase adiponectin levels, but have minimal effects on glycaemic control, thereby highlighting interspecies differences in the actions of FGF21.
-
In patients with non-alcoholic steatohepatitis, FGF21 analogues ameliorate hepatic steatosis, liver stiffness and biomarkers of liver fibrosis, whereas the long-term effects on histopathology and clinical outcomes of non-alcoholic steatohepatitis remain unknown.
-
In obesity, systemic and/or adipose depot-selective FGF21 resistance might exist and underpin insulin resistance, thus potentially compromising the therapeutic effects of FGF21 analogues.
-
Target-specific delivery of FGF21 analogues, as well as the development of tissue-selective FGF21 receptor agonists and FGF21 sensitizers, might help to improve the therapeutic efficacy and safety of FGF21-based pharmacotherapies.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others
References
Hui, Q., Jin, Z., Li, X., Liu, C. & Wang, X. FGF family: from drug development to clinical application. Int. J. Mol. Sci. 19, 1875 (2018).
Degirolamo, C., Sabba, C. & Moschetta, A. Therapeutic potential of the endocrine fibroblast growth factors FGF19, FGF21 and FGF23. Nat. Rev. Drug Discov. 15, 51–69 (2016).
Lewis, J. E., Ebling, F. J. P., Samms, R. J. & Tsintzas, K. Going back to the biology of FGF21: new insights. Trends Endocrinol. Metab. 30, 491–504 (2019).
Kharitonenkov, A. et al. FGF-21 as a novel metabolic regulator. J. Clin. Invest. 115, 1627–1635 (2005). This paper is the first to discover FGF21 as a potential antidiabetic factor.
Keuper, M., Haring, H. U. & Staiger, H. Circulating FGF21 levels in human health and metabolic disease. Exp. Clin. Endocrinol. Diabetes. https://doi.org/10.1055/a-0879-2968 (2019).
Coskun, T. et al. Fibroblast growth factor 21 corrects obesity in mice. Endocrinology 149, 6018–6027 (2008).
Kharitonenkov, A. et al. The metabolic state of diabetic monkeys is regulated by fibroblast growth factor-21. Endocrinology 148, 774–781 (2007).
Berglund, E. D. et al. Fibroblast growth factor 21 controls glycemia via regulation of hepatic glucose flux and insulin sensitivity. Endocrinology 150, 4084–4093 (2009).
Adams, A. C. et al. LY2405319, an engineered FGF21 variant, improves the metabolic status of diabetic monkeys. PLoS ONE 8, e65763 (2013).
Talukdar, S. et al. A long-acting FGF21 molecule, PF-05231023, decreases body weight and improves lipid profile in non-human primates and type 2 diabetic subjects. Cell Metab. 23, 427–440 (2016). This is the first clinical study of a long-acting FGF21 analogue in patients with T2DM.
Stanislaus, S. et al. A novel Fc-FGF21 with improved resistance to proteolysis, increased affinity toward beta-klotho, and enhanced efficacy in mice and cynomolgus monkeys. Endocrinology 158, 1314–1327 (2017).
Xu, J. et al. Fibroblast growth factor 21 reverses hepatic steatosis, increases energy expenditure, and improves insulin sensitivity in diet-induced obese mice. Diabetes 58, 250–259 (2009).
Xu, J. et al. Acute glucose-lowering and insulin-sensitizing action of FGF21 in insulin-resistant mouse models — association with liver and adipose tissue effects. Am. J. Physiol. Endocrinol. Metab. 297, E1105–E1114 (2009).
Huang, X. et al. Forced expression of hepatocyte-specific fibroblast growth factor 21 delays initiation of chemically induced hepatocarcinogenesis. Mol. Carcinog. 45, 934–942 (2006).
Ding, X. et al. Beta-klotho is required for fibroblast growth factor 21 effects on growth and metabolism. Cell Metab. 16, 387–393 (2012). This work is the first to prove that β-klotho is required for metabolic actions of FGF21 in vivo.
Fisher, F. M. et al. Fibroblast growth factor 21 limits lipotoxicity by promoting hepatic fatty acid activation in mice on methionine and choline-deficient diets. Gastroenterology 147, 1073–1083.e6 (2014).
Lee, J. H. et al. An engineered FGF21 variant, LY2405319, can prevent non-alcoholic steatohepatitis by enhancing hepatic mitochondrial function. Am. J. Transl Res. 8, 4750–4763 (2016).
Luo, Y., Krupinski, J., Gao, S., Charles, E. & Christian, R. BMS-986036, a PEGylated fibroblast growth factor 21 analogue, reduces fibrosis and Pro-C3 in a mouse model of non-alcoholic steatohepatitis. J. Hepatol. 68, S396–S397 (2018).
Lin, Z. et al. Fibroblast growth factor 21 prevents atherosclerosis by suppression of hepatic sterol regulatory element-binding protein-2 and induction of adiponectin in mice. Circulation 131, 1861–1871 (2015).
Jia, H. et al. Fibroblast growth factor 21 attenuates inflammation and oxidative stress in atherosclerotic rat via enhancing the Nrf1-ARE signaling pathway. Int. J. Clin. Exp. Pathol. 11, 1308–1317 (2018).
Wu, F. et al. FGF21 ameliorates diabetic cardiomyopathy by activating the AMPK–paraoxonase 1 signaling axis in mice. Clin. Sci. 131, 1877–1893 (2017).
Zhao, L. et al. Potent long-acting rhFGF21 analog for treatment of diabetic nephropathy in db/db and DIO mice. BMC Biotechnol. 17, 58 (2017).
Jin, L., Lin, Z. & Xu, A. Fibroblast growth factor 21 protects against atherosclerosis via fine-tuning the multiorgan crosstalk. Diabetes Metab. J. 40, 22–31 (2016).
Planavila, A., Redondo-Angulo, I. & Villarroya, F. FGF21 and cardiac physiopathology. Front. Endocrinol. 6, 133 (2015).
Brahma, M. K. et al. Fibroblast growth factor 21 is induced upon cardiac stress and alters cardiac lipid homeostasis. J. Lipid Res. 55, 2229–2241 (2014).
Planavila, A. et al. Fibroblast growth factor 21 protects against cardiac hypertrophy in mice. Nat. Commun. 4, 2019 (2013).
Planavila, A. et al. Fibroblast growth factor 21 protects the heart from oxidative stress. Cardiovasc. Res. 106, 19–31 (2015).
Wang, S., Wang, Y., Zhang, Z., Liu, Q. & Gu, J. Cardioprotective effects of fibroblast growth factor 21 against doxorubicin-induced toxicity via the SIRT1/LKB1/AMPK pathway. Cell Death Dis. 8, e3018 (2017).
Yan, X. et al. FGF21 deletion exacerbates diabetic cardiomyopathy by aggravating cardiac lipid accumulation. J. Cell Mol. Med. 19, 1557–1568 (2015).
Ruan, C. C. et al. A2A receptor activation attenuates hypertensive cardiac remodeling via promoting brown adipose tissue-derived FGF21. Cell Metab. 28, 476–489.e5 (2018).
Liu, S. Q. et al. Endocrine protection of ischemic myocardium by FGF21 from the liver and adipose tissue. Sci. Rep. 3, 2767 (2013).
Zhang, C. et al. Fibroblast growth factor 21 protects the heart from apoptosis in a diabetic mouse model via extracellular signal-regulated kinase 1/2-dependent signalling pathway. Diabetologia 58, 1937–1948 (2015).
Joki, Y. et al. FGF21 attenuates pathological myocardial remodeling following myocardial infarction through the adiponectin-dependent mechanism. Biochem. Bioph Res. Commun. 459, 124–130 (2015).
Wei, W. et al. Fibroblast growth factor 21 promotes bone loss by potentiating the effects of peroxisome proliferator-activated receptor gamma. Proc. Natl Acad. Sci. USA 109, 3143–3148 (2012).
Li, H. et al. Increased expression of FGF-21 negatively affects bone homeostasis in dystrophin/utrophin double knockout mice. J. Bone Min. Res. 35, 738–752 (2020).
Wang, X., Wei, W., Krzeszinski, J. Y., Wang, Y. & Wan, Y. A liver–bone endocrine relay by IGFBP1 promotes osteoclastogenesis and mediates FGF21-induced bone resorption. Cell Metab. 22, 811–824 (2015).
Charoenphandhu, N. et al. Fibroblast growth factor-21 restores insulin sensitivity but induces aberrant bone microstructure in obese insulin-resistant rats. J. Bone Min. Metab. 35, 142–149 (2017).
Andersen, B. et al. FGF21 decreases body weight without reducing food intake or bone mineral density in high-fat fed obese rhesus macaque monkeys. Int. J. Obes. 42, 1151–1160 (2018).
Ogawa, Y. et al. Beta-klotho is required for metabolic activity of fibroblast growth factor 21. Proc. Natl Acad. Sci. USA 104, 7432–7437 (2007).
Fon Tacer, K. et al. Research resource: comprehensive expression atlas of the fibroblast growth factor system in adult mouse. Mol. Endocrinol. 24, 2050–2064 (2010).
Petryszak, R. et al. Expression atlas update — an integrated database of gene and protein expression in humans, animals and plants. Nucleic Acids Res. 44, D746–D752 (2016).
Veniant, M. M. et al. FGF21 promotes metabolic homeostasis via white adipose and leptin in mice. PLoS ONE 7, e40164 (2012).
Dollet, L. et al. FGF21 improves the adipocyte dysfunction related to seipin deficiency. Diabetes 65, 3410–3417 (2016).
Adams, A. C. et al. The breadth of FGF21’s metabolic actions are governed by FGFR1 in adipose tissue. Mol. Metab. 2, 31–37 (2012).
Wu, A. L. et al. Amelioration of type 2 diabetes by antibody-mediated activation of fibroblast growth factor receptor 1. Sci. Transl Med. 3, 113ra126 (2011).
Adams, A. C., Cheng, C. C., Coskun, T. & Kharitonenkov, A. FGF21 requires beta-klotho to act in vivo. PLoS ONE 7, e49977 (2012).
BonDurant, L. D. et al. FGF21 regulates metabolism through adipose-dependent and -independent mechanisms. Cell Metab. 25, 935–944 (2017). This work reveals the importance of both adipose actions and central actions of FGF21 in regulating metabolism.
Ge, X. et al. Fibroblast growth factor 21 induces glucose transporter-1 expression through activation of the serum response factor/Ets-like protein-1 in adipocytes. J. Biol. Chem. 286, 34533–34541 (2011).
Lewis, J. E. et al. Whole-body and adipose tissue-specific mechanisms underlying the metabolic effects of fibroblast growth factor 21 in the Siberian hamster. Mol. Metab. 31, 45–54 (2020).
Schlein, C. et al. FGF21 lowers plasma triglycerides by accelerating lipoprotein catabolism in white and brown adipose tissues. Cell Metab. 23, 441–453 (2016). This study is the first report showing the roles of adipose tissues in the lipid-lowering effects of FGF21.
Arner, P. et al. FGF21 attenuates lipolysis in human adipocytes — a possible link to improved insulin sensitivity. FEBS Lett. 582, 1725–1730 (2008).
Li, X. et al. Inhibition of lipolysis may contribute to the acute regulation of plasma FFA and glucose by FGF21 in ob/ob mice. FEBS Lett. 583, 3230–3234 (2009).
Inagaki, T. et al. Endocrine regulation of the fasting response by PPARalpha-mediated induction of fibroblast growth factor 21. Cell Metab. 5, 415–425 (2007). This study establishes FGF21 as a crucial hormone in the body’s adaptation to fasting.
Geng, L. et al. Exercise alleviates obesity-induced metabolic dysfunction via enhancing FGF21 sensitivity in adipose tissues. Cell Rep. 26, 2738–2752 (2019).
Hotta, Y. et al. Fibroblast growth factor 21 regulates lipolysis in white adipose tissue but is not required for ketogenesis and triglyceride clearance in liver. Endocrinology 150, 4625–4633 (2009).
Lin, Z. et al. Adiponectin mediates the metabolic effects of FGF21 on glucose homeostasis and insulin sensitivity in mice. Cell Metab. 17, 779–789 (2013). This work uncovers the important role of adiponectin as a downstream effector of FGF21 mediating its metabolic benefits in mice.
Holland, W. L. et al. An FGF21–adiponectin–ceramide axis controls energy expenditure and insulin action in mice. Cell Metab. 17, 790–797 (2013).
Zinker, B. et al. Effects of a PEGylated fibroblast growth factor 21 variant on steatosis, inflammation, and fibrosis in a mouse model of nonalcoholic steatohepatitis. J. Hepatol. 68, S390 (2018).
Yamauchi, T. et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 7, 941–946 (2001).
Gaich, G. et al. The effects of LY2405319, an FGF21 analog, in obese human subjects with type 2 diabetes. Cell Metab. 18, 333–340 (2013). This is the first clinical study of a short-acting FGF21 analogue in patients with T2DM.
Sanyal, A. et al. Pegbelfermin (BMS-986036), a PEGylated fibroblast growth factor 21 analogue, in patients with non-alcoholic steatohepatitis: a randomised, double-blind, placebo-controlled, phase 2a trial. Lancet 392, 2705–2717 (2018). This is the first study reporting the beneficial effects of an FGF21 analogue on improving NASH in humans.
Bao, L. et al. A long-acting FGF21 alleviates hepatic steatosis and inflammation in a mouse model of non-alcoholic steatohepatitis partly through an FGF21–adiponectin–IL17A pathway. Br. J. Pharmacol. 175, 3379–3393 (2018).
Fisher, F. M. et al. FGF21 regulates PGC-1alpha and browning of white adipose tissues in adaptive thermogenesis. Genes. Dev. 26, 271–281 (2012).
Huang, Z. et al. The FGF21–CCL11 axis mediates beiging of white adipose tissues by coupling sympathetic nervous system to type 2 immunity. Cell Metab. 26, 493–508 (2017). This paper identifies an important mechanism whereby FGF21 induces beiging of white adipocytes.
Kwon, M. M., O’Dwyer, S. M., Baker, R. K., Covey, S. D. & Kieffer, T. J. FGF21-mediated improvements in glucose clearance require uncoupling protein 1. Cell Rep. 13, 1521–1527 (2015).
Veniant, M. M. et al. Pharmacologic effects of FGF21 are independent of the “browning” of white adipose tissue. Cell Metab. 21, 731–738 (2015).
Samms, R. J. et al. Discrete aspects of FGF21 in vivo pharmacology do not require UCP1. Cell Rep. 11, 991–999 (2015).
Liu, X. et al. Paradoxical resistance to diet-induced obesity in UCP1-deficient mice. J. Clin. Invest. 111, 399–407 (2003).
Keipert, S. et al. Endogenous FGF21-signaling controls paradoxical obesity resistance of UCP1-deficient mice. Nat. Commun. 11, 624 (2020).
Chen, M. Z. et al. FGF21 mimetic antibody stimulates UCP1-independent brown fat thermogenesis via FGFR1/beta-klotho complex in non-adipocytes. Mol. Metab. 6, 1454–1467 (2017).
Liang, Q. et al. FGF21 maintains glucose homeostasis by mediating the cross talk between liver and brain during prolonged fasting. Diabetes 63, 4064–4075 (2014).
Bookout, A. L. et al. FGF21 regulates metabolism and circadian behavior by acting on the nervous system. Nat. Med. 19, 1147–1152 (2013). This work provides the first evidence that FGF21 directly acts on the brain in mice.
Hultman, K. et al. The central fibroblast growth factor receptor/beta klotho system: comprehensive mapping in Mus musculus and comparisons to nonhuman primate and human samples using an automated in situ hybridization platform. J. Comp. Neurol. 527, 2069–2085 (2019).
Talukdar, S. et al. FGF21 regulates sweet and alcohol preference. Cell Metab. 23, 344–349 (2016).
von Holstein-Rathlou, S. et al. FGF21 mediates endocrine control of simple sugar intake and sweet taste preference by the liver. Cell Metab. 23, 335–343 (2016).
Yilmaz, U., Tekin, S., Demir, M., Cigremis, Y. & Sandal, S. Effects of central FGF21 infusion on the hypothalamus–pituitary–thyroid axis and energy metabolism in rats. J. Physiol. Sci. 68, 781–788 (2018).
Sarruf, D. A. et al. Fibroblast growth factor 21 action in the brain increases energy expenditure and insulin sensitivity in obese rats. Diabetes 59, 1817–1824 (2010).
Owen, B. M. et al. FGF21 acts centrally to induce sympathetic nerve activity, energy expenditure, and weight loss. Cell Metab. 20, 670–677 (2014).
Lan, T. et al. FGF19, FGF21, and an FGFR1/beta-klotho-activating antibody act on the nervous system to regulate body weight and glycemia. Cell Metab. 26, 709–718.e3 (2017).
Douris, N. et al. Central fibroblast growth factor 21 browns white fat via sympathetic action in male mice. Endocrinology 156, 2470–2481 (2015).
Markan, K. R. et al. Circulating FGF21 is liver derived and enhances glucose uptake during refeeding and overfeeding. Diabetes 63, 4057–4063 (2014).
Gong, Q. et al. Fibroblast growth factor 21 improves hepatic insulin sensitivity by inhibiting mammalian target of rapamycin complex 1 in mice. Hepatology 64, 425–438 (2016).
Potthoff, M. J. et al. FGF21 induces PGC-1alpha and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proc. Natl Acad. Sci. USA 106, 10853–10858 (2009).
Liu, Y. et al. Fibroblast growth factor 21 deficiency exacerbates chronic alcohol-induced hepatic steatosis and injury. Sci. Rep. 6, 31026 (2016).
Zarei, M. et al. Hepatic regulation of VLDL receptor by PPARbeta/delta and FGF21 modulates non-alcoholic fatty liver disease. Mol. Metab. 8, 117–131 (2018).
Ye, D. et al. Fibroblast growth factor 21 protects against acetaminophen-induced hepatotoxicity by potentiating peroxisome proliferator-activated receptor coactivator protein-1alpha-mediated antioxidant capacity in mice. Hepatology 60, 977–989 (2014).
Kim, S. H. et al. Fibroblast growth factor 21 participates in adaptation to endoplasmic reticulum stress and attenuates obesity-induced hepatic metabolic stress. Diabetologia 58, 809–818 (2015).
Feingold, K. R. et al. FGF21 is increased by inflammatory stimuli and protects leptin-deficient ob/ob mice from the toxicity of sepsis. Endocrinology 153, 2689–2700 (2012).
Xu, P. et al. Fibroblast growth factor 21 attenuates hepatic fibrogenesis through TGF-β/smad2/3 and NF-κB signaling pathways. Toxicol. Appl. Pharmacol. 290, 43–53 (2016).
Kurosu, H. et al. Tissue-specific expression of beta-klotho and fibroblast growth factor (FGF) receptor isoforms determines metabolic activity of FGF19 and FGF21. J. Biol. Chem. 282, 26687–26695 (2007).
Fisher, F. M. et al. Integrated regulation of hepatic metabolism by fibroblast growth factor 21 (FGF21) in vivo. Endocrinology 152, 2996–3004 (2011).
Zhao, L. et al. Paracrine-endocrine FGF chimeras as potent therapeutics for metabolic diseases. EBioMedicine 48, 462–477 (2019).
Foltz, I. N. et al. Treating diabetes and obesity with an FGF21-mimetic antibody activating the βklotho/FGFR1c receptor complex. Sci. Transl Med. 4, 162ra153 (2012). This study is the first to show that an agonistic antibody specifically activating the FGF21 receptor complex can mimic the therapeutic benefits of FGF21.
Huang, J. et al. Development of a novel long-acting antidiabetic FGF21 mimetic by targeted conjugation to a scaffold antibody. J. Pharmacol. Exp. Ther. 346, 270–280 (2013).
Gillum, M. P. & Potthoff, M. J. FAP finds FGF21 easy to digest. Biochem. J. 473, 1125–1127 (2016).
Dunshee, D. R. et al. Fibroblast activation protein cleaves and inactivates fibroblast growth factor 21. J. Biol. Chem. 291, 5986–5996 (2016).
Kharitonenkov, A. et al. Rational design of a fibroblast growth factor 21-based clinical candidate, LY2405319. PLoS ONE 8, e58575 (2013).
Hecht, R. et al. Rationale-based engineering of a potent long-acting FGF21 analog for the treatment of type 2 diabetes. PLoS ONE 7, e49345 (2012).
Camacho, R. C., Zafian, P. T., Achanfuo-Yeboah, J., Manibusan, A. & Berger, J. P. Pegylated Fgf21 rapidly normalizes insulin-stimulated glucose utilization in diet-induced insulin resistant mice. Eur. J. Pharmacol. 715, 41–45 (2013).
Charles, E. D. et al. Pegbelfermin (BMS-986036), PEGylated FGF21, in patients with obesity and type 2 diabetes: results from a randomized phase 2 study. Obesity 27, 41–49 (2019).
Weng, Y. et al. Pharmacokinetics (PK), pharmacodynamics (PD) and integrated PK/PD modeling of a novel long acting FGF21 clinical candidate PF-05231023 in diet-induced obese and leptin-deficient obese mice. PLoS ONE 10, e0119104 (2015).
Thompson, W. C., Zhou, Y., Talukdar, S. & Musante, C. J. PF-05231023, a long-acting FGF21 analogue, decreases body weight by reduction of food intake in non-human primates. J. Pharmacokinet. Pharmacodyn. 43, 411–425 (2016).
Lee, S. et al. Structures of beta-klotho reveal a ‘zip code’-like mechanism for endocrine FGF signalling. Nature 553, 501–505 (2018). This paper is the first to reveal the crystal structure of the FGF21 coreceptor β-klotho.
Smith, R. et al. FGF21 can be mimicked in vitro and in vivo by a novel anti-FGFR1c/beta-klotho bispecific protein. PLoS ONE 8, e61432 (2013).
Min, X. et al. Agonistic beta-klotho antibody mimics fibroblast growth factor 21 (FGF21) functions. J. Biol. Chem. 293, 14678–14688 (2018).
Kolumam, G. et al. Sustained brown fat stimulation and insulin sensitization by a humanized bispecific antibody agonist for fibroblast growth factor receptor 1/beta-klotho complex. EBioMedicine 2, 730–743 (2015).
Shi, S. Y. et al. A biparatopic agonistic antibody that mimics fibroblast growth factor 21 ligand activity. J. Biol. Chem. 293, 5909–5919 (2018).
Ritchie, M., Hanouneh, I. A., Noureddin, M., Rolph, T. & Alkhouri, N. Fibroblast growth factor (FGF)-21 based therapies: a magic bullet for nonalcoholic fatty liver disease (NAFLD)? Expert. Opin. Investig. Drugs 29, 197–204 (2020).
Kim, A. M. et al. Once-weekly administration of a long-acting fibroblast growth factor 21 analogue modulates lipids, bone turnover markers, blood pressure and body weight differently in obese people with hypertriglyceridaemia and in non-human primates. Diabetes Obes. Metab. 19, 1762–1772 (2017).
Depaoli, A. et al. NGM313, a novel activator of beta-klotho/FGFR1c, improves insulin resistance and reduces hepatic fat in obese, nondiabetic subjects [abstract 140-LB]. Diabetes 68 (Suppl. 1) 140-LB (2019).
Dutchak, P. A. et al. Fibroblast growth factor-21 regulates PPARgamma activity and the antidiabetic actions of thiazolidinediones. Cell 148, 556–567 (2012).
Badman, M. K., Koester, A., Flier, J. S., Kharitonenkov, A. & Maratos-Flier, E. Fibroblast growth factor 21-deficient mice demonstrate impaired adaptation to ketosis. Endocrinology 150, 4931–4940 (2009).
Li, H. et al. Fibroblast growth factor 21 increases insulin sensitivity through specific expansion of subcutaneous fat. Nat. Commun. 9, 272 (2018).
Chen, W. et al. Growth hormone induces hepatic production of fibroblast growth factor 21 through a mechanism dependent on lipolysis in adipocytes. J. Biol. Chem. 286, 34559–34566 (2011).
Lundberg, J. et al. Influence of growth hormone on circulating fibroblast growth factor 21 levels in humans. J. Intern. Med. 274, 227–232 (2013).
Badman, M. K. et al. Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 5, 426–437 (2007).
Christodoulides, C., Dyson, P., Sprecher, D., Tsintzas, K. & Karpe, F. Circulating fibroblast growth factor 21 is induced by peroxisome proliferator-activated receptor agonists but not ketosis in man. J. Clin. Endocrinol. Metab. 94, 3594–3601 (2009).
Fazeli, P. K. et al. FGF21 and the late adaptive response to starvation in humans. J. Clin. Invest. 125, 4601–4611 (2015).
Galman, C. et al. The circulating metabolic regulator FGF21 is induced by prolonged fasting and PPARalpha activation in man. Cell Metab. 8, 169–174 (2008).
Li, P., Zhao, M., Qi, X., Zhu, X. & Dai, J. Downregulation of klotho beta is associated with invasive ductal carcinoma progression. Oncol. Lett. 14, 7443–7448 (2017).
van Marken Lichtenbelt, W. D. et al. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 360, 1500–1508 (2009).
Lee, P., Greenfield, J. R., Ho, K. K. & Fulham, M. J. A critical appraisal of the prevalence and metabolic significance of brown adipose tissue in adult humans. Am. J. Physiol. Endocrinol. Metab. 299, E601–E606 (2010).
Zhang, X. et al. Serum FGF21 levels are increased in obesity and are independently associated with the metabolic syndrome in humans. Diabetes 57, 1246–1253 (2008). This is the first clinical study showing paradoxical increase of circulating FGF21 in individuals with obesity and patients with metabolic syndrome.
Fisher, F. M. et al. Obesity is a fibroblast growth factor 21 (FGF21)-resistant state. Diabetes 59, 2781–2789 (2010).
Konner, A. C. & Bruning, J. C. Selective insulin and leptin resistance in metabolic disorders. Cell Metab. 16, 144–152 (2012).
Yang, W. et al. Exercise ameliorates the FGF21–adiponectin axis impairment in diet-induced obese mice. Endocr. Connect. 8, 596–604 (2019).
Patel, V. et al. Novel insights into the cardio-protective effects of FGF21 in lean and obese rat hearts. PLoS ONE 9, e87102 (2014).
So, W. Y. et al. High glucose represses beta-klotho expression and impairs fibroblast growth factor 21 action in mouse pancreatic islets: involvement of peroxisome proliferator-activated receptor gamma signaling. Diabetes 62, 3751–3759 (2013).
Asrih, M., Altirriba, J., Rohner-Jeanrenaud, F. & Jornayvaz, F. R. Ketogenic diet impairs FGF21 signaling and promotes differential inflammatory responses in the liver and white adipose tissue. PLoS ONE 10, e0126364 (2015).
Samms, R. J., Cheng, C. C., Kharitonenkov, A., Gimeno, R. E. & Adams, A. C. Overexpression of beta-klotho in adipose tissue sensitizes male mice to endogenous FGF21 and provides protection from diet-induced obesity. Endocrinology 157, 1467–1480 (2016).
Diaz-Delfin, J. et al. TNF-alpha represses beta-klotho expression and impairs FGF21 action in adipose cells: involvement of JNK1 in the FGF21 pathway. Endocrinology 153, 4238–4245 (2012).
Fu, T. et al. MicroRNA-34a inhibits beige and brown fat formation in obesity in part by suppressing adipocyte fibroblast growth factor 21 signaling and SIRT1 function. Mol. Cell Biol. 34, 4130–4142 (2014).
Pan, Y., Feng, T., Chan, Y.-C., Xu, A. & Lam, K. OR01-3 microRNA-34a-mediated FGF21 resistance in the adipose tissue contributes to insulin resistance and hypoadiponectinemia in diet-induced obesity. J. Endocr. Soc. https://doi.org/10.1210/js.2019-OR01-3 (2019).
Pan, Y. et al. Adipocyte-secreted exosomal microRNA-34a inhibits M2 macrophage polarization to promote obesity-induced adipose inflammation. J. Clin. Invest. 129, 834–849 (2019).
Moyers, J. S. et al. Molecular determinants of FGF-21 activity — synergy and cross-talk with PPARgamma signaling. J. Cell Physiol. 210, 1–6 (2007).
Hale, C. et al. Lack of overt FGF21 resistance in two mouse models of obesity and insulin resistance. Endocrinology 153, 69–80 (2012).
Li, H. et al. High serum level of fibroblast growth factor 21 is an independent predictor of non-alcoholic fatty liver disease: a 3-year prospective study in China. J. Hepatol. 58, 557–563 (2013).
Gallego-Escuredo, J. M. et al. Opposite alterations in FGF21 and FGF19 levels and disturbed expression of the receptor machinery for endocrine FGFs in obese patients. Int. J. Obes. 39, 121–129 (2015).
Hong, E. S. et al. Plasma fibroblast growth factor 21 levels increase with ectopic fat accumulation and its receptor levels are decreased in the visceral fat of patients with type 2 diabetes. BMJ Open. Diabetes Res. Care 7, e000776 (2019).
Liu, Q. et al. Improved FGF21 sensitivity and restored FGF21 signaling pathway in high-fat diet/streptozotocin-induced diabetic rats after duodenal-jejunal bypass and sleeve gastrectomy. Front. Endocrinol. 10, 566 (2019).
Zeng, K. et al. Diet polyphenol curcumin stimulates hepatic Fgf21 production and restores its sensitivity in high-fat-diet-fed male mice. Endocrinology 158, 277–292 (2017).
Yie, J. et al. FGF21 N- and C-termini play different roles in receptor interaction and activation. FEBS Lett. 583, 19–24 (2009).
Micanovic, R. et al. Different roles of N- and C- termini in the functional activity of FGF21. J. Cell Physiol. 219, 227–234 (2009).
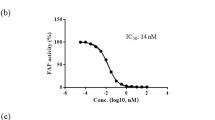
Zhen, E. Y., Jin, Z., Ackermann, B. L., Thomas, M. K. & Gutierrez, J. A. Circulating FGF21 proteolytic processing mediated by fibroblast activation protein. Biochem. J. 473, 605–614 (2016). This work is the first to identify FAP as a circulating protease that cleaves and inactivates the endogenous FGF21.
Coppage, A. L. et al. Human FGF-21 is a substrate of fibroblast activation protein. PLoS ONE 11, e0151269 (2016).
Sanchez-Garrido, M. A. et al. Fibroblast activation protein (FAP) as a novel metabolic target. Mol. Metab. 5, 1015–1024 (2016).
Bachovchin, D. A. et al. A high-throughput, multiplexed assay for superfamily-wide profiling of enzyme activity. Nat. Chem. Biol. 10, 656–663 (2014).
Panaro, B. L. et al. Fibroblast activation protein is dispensable for control of glucose homeostasis and body weight in mice. Mol. Metab. 19, 65–74 (2019).
Samms, R. J. et al. FGF21 is an insulin-dependent postprandial hormone in adult humans. J. Clin. Endocrinol. Metab. 102, 3806–3813 (2017).
Minard, A. Y. et al. mTORC1 is a major regulatory node in the FGF21 signaling network in adipocytes. Cell Rep. 17, 29–36 (2016).
Hamson, E. J., Keane, F. M., Tholen, S., Schilling, O. & Gorrell, M. D. Understanding fibroblast activation protein (FAP): substrates, activities, expression and targeting for cancer therapy. Proteom. Clin. Appl. 8, 454–463 (2014).
Acknowledgements
The authors acknowledge the support of the Hong Kong Research Grants Council/Area of Excellence (AoE/M/707-18), the Collaborative Research Fund (C7037-17W), the General Research Fund (17125317 and 17127518) and the Science and Technology Major Project of Hunan Province (2017SK1020).
Author information
Authors and Affiliations
Contributions
A.X. and L.G. researched data for the article, made substantial contributions to discussions of the content and wrote the article. A.X., L.G. and K.S.L.L. carried out a review and editing of the manuscript before submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information
Nature Reviews Endocrinology thanks D. Mangelsdorf, E. Maratos-Flier and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Glossary
- Type 2 immune responses
-
Type 2 T helper cell immune responses are characterized by the recruitment and activation of mast cells, basophils and eosinophils, and goblet cell hyperplasia to produce IL-4, IL-5 and IL-13 in tissues.
- Fragment antigen-binding region
-
(Fab). A region on an antibody that binds to antigens that is composed of one constant and one variable domain of each of the heavy and the light chain.
- Fragment crystallizable region
-
(Fc). The tail region of an antibody that interacts with cell surface receptors called Fc receptors and some proteins of the complement system.
- Avimer
-
A type of artificial antibody mimetic specifically binding to certain antigens via multiple binding sites.
- Phage display
-
A laboratory technique for the study of protein–protein, protein–peptide and protein–DNA interactions that uses bacteriophages (viruses that infect bacteria) to connect proteins with the genetic information that encodes them, which enables antibody maturation in vitro and revolutionizes antibody drug discovery.
- XenoMouse
-
A genetically engineered strain of mice that produce fully humanized antibodies for therapeutic applications.
- Knobs-into-holes
-
Originally proposed as a model for the packing of amino acid side chains between adjacent α-helices, this technology is now being used as an effective design strategy for engineering antibody heavy chain homodimers for heterodimerization.
- Biparatopic molecule
-
A bispecific molecule that recognizes two different epitopes on the same antigen.
Rights and permissions
About this article

Cite this article
Geng, L., Lam, K.S.L. & Xu, A. The therapeutic potential of FGF21 in metabolic diseases: from bench to clinic. Nat Rev Endocrinol 16, 654–667 (2020). https://doi.org/10.1038/s41574-020-0386-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41574-020-0386-0
This article is cited by
-
Skin advanced glycation end-products as indicators of the metabolic profile in diabetes mellitus: correlations with glycemic control, liver phenotypes and metabolic biomarkers
BMC Endocrine Disorders (2024)
-
Advances in secondary prevention mechanisms of macrovascular complications in type 2 diabetes mellitus patients: a comprehensive review
European Journal of Medical Research (2024)
-
Intellectual disability and autism in propionic acidemia: a biomarker-behavioral investigation implicating dysregulated mitochondrial biology
Molecular Psychiatry (2024)
-
Exerkines and cardiometabolic benefits of exercise: from bench to clinic
EMBO Molecular Medicine (2024)
-
Inter-organ crosstalk during development and progression of type 2 diabetes mellitus
Nature Reviews Endocrinology (2024)
