
Abstract
LY6E is an antiviral restriction factor that inhibits coronavirus spike-mediated fusion, but the cell types in vivo that require LY6E for protection from respiratory coronavirus infection are unknown. Here we used a panel of seven conditional Ly6e knockout mice to define which Ly6e-expressing cells confer control of airway infection by murine coronavirus and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Loss of Ly6e in Lyz2-expressing cells, radioresistant Vav1-expressing cells and non-haematopoietic cells increased susceptibility to murine coronavirus. Global conditional loss of Ly6e expression resulted in clinical disease and higher viral burden after SARS-CoV-2 infection, but little evidence of immunopathology. We show that Ly6e expression protected secretory club and ciliated cells from SARS-CoV-2 infection and prevented virus-induced loss of an epithelial cell transcriptomic signature in the lung. Our study demonstrates that lineage confined rather than broad expression of Ly6e sufficiently confers resistance to disease caused by murine and human coronaviruses.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
The authors declare that the data supporting the findings of this study are available within the article and its Supplementary Information files or are available on request. The RNA-seq data discussed in this publication have been deposited in the Gene Expression Omnibus database (GSE209974). Source data are provided with this paper.
References
Schoggins, J. W. Interferon-stimulated genes: what do they all do? Annu. Rev. Virol. 6, 567–584 (2019).
Mao, M. et al. RIG-E, a human homolog of the murine Ly-6 family, is induced by retinoic acid during the differentiation of acute promyelocytic leukemia cell. Proc. Natl Acad. Sci. USA 93, 5910–5914 (1996).
Bacquin, A. et al. A cell fusion-based screening method identifies glycosylphosphatidylinositol-anchored protein Ly6e as the receptor for mouse endogenous retroviral envelope syncytin-A. J. Virol. 91, e00832–17 (2017).
Schupp, J. C. et al. Integrated single-cell atlas of endothelial cells of the human lung. Circulation 144, 286–302 (2021).
Tabula Muris, C. et al. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 562, 367–372 (2018).
Mar, K. B. et al. LY6E mediates an evolutionarily conserved enhancement of virus infection by targeting a late entry step. Nat. Commun. 9, 3603 (2018).
Lee, P. Y., Wang, J. X., Parisini, E., Dascher, C. C. & Nigrovic, P. A. Ly6 family proteins in neutrophil biology. J. Leukoc. Biol. 94, 585–594 (2013).
Saitoh, S. et al. Modulation of TCR-mediated signaling pathway by thymic shared antigen-1 (TSA-1)/stem cell antigen-2 (Sca-2). J. Immunol. 155, 5574–5581 (1995).
Mao, W., Hunt, H. D. & Cheng, H. H. Cloning and functional characterization of chicken stem cell antigen 2. Dev. Comp. Immunol. 34, 360–368 (2010).
Xu, X. et al. IFN-stimulated gene LY6E in monocytes regulates the CD14/TLR4 pathway but inadequately restrains the hyperactivation of monocytes during chronic HIV-1 infection. J. Immunol. 193, 4125–4136 (2014).
Classon, B. J. & Coverdale, L. Mouse stem cell antigen Sca-2 is a member of the Ly-6 family of cell surface proteins. Proc. Natl Acad. Sci. USA 91, 5296–5300 (1994).
Wu, L. et al. Mouse thymus dendritic cells: kinetics of development and changes in surface markers during maturation. Eur. J. Immunol. 25, 418–425 (1995).
Noda, S., Kosugi, A., Saitoh, S., Narumiya, S. & Hamaoka, T. Protection from anti-TCR/CD3-induced apoptosis in immature thymocytes by a signal through thymic shared antigen-1/stem cell antigen-2. J. Exp. Med. 183, 2355–2360 (1996).
Zammit, D. J. et al. Essential role for the lymphostromal plasma membrane Ly-6 superfamily molecule thymic shared antigen 1 in development of the embryonic adrenal gland. Mol. Cell. Biol. 22, 946–952 (2002).
Langford, M. B., Outhwaite, J. E., Hughes, M., Natale, D. R. C. & Simmons, D. G. Deletion of the Syncytin A receptor Ly6e impairs syncytiotrophoblast fusion and placental morphogenesis causing embryonic lethality in mice. Sci. Rep. 8, 3961 (2018).
Krishnan, M. N. et al. RNA interference screen for human genes associated with West Nile virus infection. Nature 455, 242–245 (2008).
Schoggins, J. W. et al. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature 472, 481–485 (2011).
Schoggins, J. W. et al. Pan-viral specificity of IFN-induced genes reveals new roles for cGAS in innate immunity. Nature 505, 691–695 (2014).
Hackett, B. A. & Cherry, S. Flavivirus internalization is regulated by a size-dependent endocytic pathway. Proc. Natl Acad. Sci. USA 115, 4246–4251 (2018).
Yu, J., Liang, C. & Liu, S. L. Interferon-inducible LY6E protein promotes HIV-1 infection. J. Biol. Chem. 292, 4674–4685 (2017).
Pfaender, S. et al. LY6E impairs coronavirus fusion and confers immune control of viral disease. Nat. Microbiol 5, 1330–1339 (2020).
Wickenhagen, A. et al. A prenylated dsRNA sensor protects against severe COVID-19. Science 374, eabj3624 (2021).
Danziger, O., Patel, R. S., DeGrace, E. J., Rosen, M. R. & Rosenberg, B. R. Inducible CRISPR activation screen for interferon-stimulated genes identifies OAS1 as a SARS-CoV-2 restriction factor. PLoS Pathog. 18, e1010464 (2022).
Mac Kain, A. et al. Identification of DAXX as a restriction factor of SARS-CoV-2 through a CRISPR/Cas9 screen. Nat. Commun. 13, 2442 (2022).
Abram, C. L., Roberge, G. L., Hu, Y. & Lowell, C. A. Comparative analysis of the efficiency and specificity of myeloid-Cre deleting strains using ROSA-EYFP reporter mice. J. Immunol. Methods 408, 89–100 (2014).
McCubbrey, A. L., Allison, K. C., Lee-Sherick, A. B., Jakubzick, C. V. & Janssen, W. J. Promoter specificity and efficacy in conditional and inducible transgenic targeting of lung macrophages. Front. Immunol. 8, 1618 (2017).
Joseph, C. et al. Deciphering hematopoietic stem cells in their niches: a critical appraisal of genetic models, lineage tracing, and imaging strategies. Cell Stem Cell 13, 520–533 (2013).
Bao, L. et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 583, 830–833 (2020).
Dinnon, K. H. 3rd et al. A mouse-adapted model of SARS-CoV-2 to test COVID-19 countermeasures. Nature 586, 560–566 (2020).
Leist, S. R. et al. A mouse-adapted SARS-CoV-2 induces acute lung injury and mortality in standard laboratory mice. Cell 183, 1070–1085 e1012 (2020).
Gu, H. et al. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 369, 1603–1607 (2020).
Imai, M. et al. Characterization of a new SARS-CoV-2 variant that emerged in Brazil. Proc. Natl Acad. Sci. USA 118, e2106535118 (2021).
Orthgiess, J. et al. Neurons exhibit Lyz2 promoter activity in vivo: Implications for using LysM-Cre mice in myeloid cell research. Eur. J. Immunol. 46, 1529–1532 (2016).
Gandhi, R. T., Lynch, J. B. & Del Rio, C. Mild or moderate Covid-19. N. Engl. J. Med. 383, 1757–1766 (2020).
Huang, K. et al. Q493K and Q498H substitutions in Spike promote adaptation of SARS-CoV-2 in mice. EBioMedicine 67, 103381 (2021).
Sun, S. et al. Characterization and structural basis of a lethal mouse-adapted SARS-CoV-2. Nat. Commun. 12, 5654 (2021).
Muruato, A. et al. Mouse-adapted SARS-CoV-2 protects animals from lethal SARS-CoV challenge. PLoS Biol. 19, e3001284 (2021).
Winkler, E. S. et al. SARS-CoV-2 infection of human ACE2-transgenic mice causes severe lung inflammation and impaired function. Nat. Immunol. 21, 1327–1335 (2020).
Zheng, J. et al. COVID-19 treatments and pathogenesis including anosmia in K18-hACE2 mice. Nature 589, 603–607 (2021).
Fumagalli, V. et al. Administration of aerosolized SARS-CoV-2 to K18-hACE2 mice uncouples respiratory infection from fatal neuroinvasion. Sci. Immunol. 7, eabl9929 (2022).
Salahudeen, A. A. et al. Progenitor identification and SARS-CoV-2 infection in human distal lung organoids. Nature 588, 670–675 (2020).
Ravindra, N. G. et al. Single-cell longitudinal analysis of SARS-CoV-2 infection in human airway epithelium identifies target cells, alterations in gene expression, and cell state changes. PLoS Biol. 19, e3001143 (2021).
Fiege, J. K. et al. Single cell resolution of SARS-CoV-2 tropism, antiviral responses, and susceptibility to therapies in primary human airway epithelium. PLoS Pathog. 17, e1009292 (2021).
Peng, Y. et al. Angiotensin-converting enzyme 2 in peripheral lung club cells modulates the susceptibility to SARS-CoV-2 in chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 322, L712–L721 (2022).
Fadista, J. et al. Shared genetic etiology between idiopathic pulmonary fibrosis and COVID-19 severity. EBioMedicine 65, 103277 (2021).
van Moorsel, C. H. M. et al. The MUC5B promoter polymorphism associates with severe COVID-19 in the European population. Front. Med. 8, 668024 (2021).
Verma, A. et al. A MUC5B gene polymorphism, rs35705950-T, confers protective effects against COVID-19 hospitalization but not severe disease or mortality. Am. J. Respir. Crit. Care Med 206, 1220–1229 (2022).
Initiative, C.-H. G. A first update on mapping the human genetic architecture of COVID-19. Nature 608, E1–E10 (2022).
Coley, S. E. et al. Recombinant mouse hepatitis virus strain A59 from cloned, full-length cDNA replicates to high titers in vitro and is fully pathogenic in vivo. J. Virol. 79, 3097–3106 (2005).
Rihn, S. J. et al. A plasmid DNA-launched SARS-CoV-2 reverse genetics system and coronavirus toolkit for COVID-19 research. PLoS Biol. 19, e3001091 (2021).
Brattelid, T. et al. Reference gene alternatives to Gapdh in rodent and human heart failure gene expression studies. BMC Mol. Biol. 11, 22 (2010).
Jhingran, A., Kasahara, S. & Hohl, T. M. Flow cytometry of lung and bronchoalveolar lavage fluid cells from mice challenged with fluorescent Aspergillus reporter (FLARE) conidia. Bio Protoc. 6, e1927 (2016).
Dietert, K. et al. Spectrum of pathogen- and model-specific histopathologies in mouse models of acute pneumonia. PLoS ONE 12, e0188251 (2017).
Choi, H. M. et al. Mapping a multiplexed zoo of mRNA expression. Development 143, 3632–3637 (2016).
Choi, H. M. T. et al. Third-generation in situ hybridization chain reaction: multiplexed, quantitative, sensitive, versatile, robust. Development 145, dev165753 (2018).
Hurskainen, M. et al. Single cell transcriptomic analysis of murine lung development on hyperoxia-induced damage. Nat. Commun. 12, 1565 (2021).
Richardson, R. B. et al. A CRISPR screen identifies IFI6 as an ER-resident interferon effector that blocks flavivirus replication. Nat. Microbiol 3, 1214–1223 (2018).
Shalem, O. et al. Genome-scale CRISPR–Cas9 knockout screening in human cells. Science 343, 84–87 (2014).
Acknowledgements
We thank members of the Schoggins lab for useful discussions. We also thank M. Aufiero and T. Hohl (Memorial Sloan Kettering Cancer Center) for advice and feedback for bone marrow chimera studies, the lab of L. Hooper (University of Texas Southwestern Medical Center (UTSW)) for CD11c-Cre and LysM-Cre transgenic mice, the labs of J. Pfeiffer (UTSW) and M. Baldridge (Washington University in St. Louis) for Ifnar−/− and Ifnlr−/− mice, respectively, the UTSW Animal Resource Center for training and animal husbandry, the UTSW Metabolic Phenotyping Core for analysis of serum samples for ALT levels and expertise, the UTSW Histo Pathology Core, the UTSW Preclinical Radiation Core Facility (supported by funding from CPRIT grant RP180770) and the UTSW Immunology Flow Cytometry Core. The authors also acknowledge the Quantitative Light Microscopy Core, a Shared Resource of the Harold C. Simmons Cancer Center, supported in part by an NCI Cancer Center Support Grant, 1P30 CA142543-01, and 1S10 RR029731-01 to K. Luby-Phelps.
Author information
Authors and Affiliations
Contributions
K.B.M. and J.W.S. designed the project. K.B.M., M.C.C.V.D., A.H.L. and J.W.S. performed in vivo and in vitro experiments with help from W.F. and N.W.H. A.I.W. performed HCR-RNA-FISH and immunofluorescence experiments. J.L.E. and J.W.S. performed and analysed HBE experiments. J.L.E. performed in vitro experiments. B.M.E. and J.M.S. contributed to pathology analysis. K.B.M. analysed remaining data and prepared figures. K.B.M. and J.W.S. wrote the manuscript. All authors reviewed and provided comments on the manuscript. This study was supported by grants from The Clayton Foundation (to J.W.S.) and NIH (AI158124 to J.W.S. and AI132751 to N.W.H.). J.W.S. holds an Investigators in the Pathogenesis of Infectious Disease Award from the Burroughs Wellcome Fund.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Microbiology thanks Yize Li, Olivier Schwartz and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1
a-f, Ly6efl/fl and Ly6eΔVav1 mice were intraperitoneally infected with 5,000 PFU MHV-A59 and assessed for survival (a), viral burden in brain, lung, spleen, and liver (b), viral burden in heart (c), viral burden in serum (d), serum alanine aminotransferase (e), and post-mortem spleen weight (f). In a, data represents means from n = 34 Ly6efl/fl and n = 23 Ly6eΔVav1; b, d-f, n = 8 Ly6efl/fl and n = 15 Ly6eΔVav1; c, n = 5 Ly6efl/fl and n = 10 Ly6eΔVav1. g, Flow cytometry gating strategy for sorting lymphocytes from the spleen for examining Ly6e gene expression in Ly6eΔCD4, Ly6eΔCD8a, and Ly6eΔCD19 mice relative to Ly6efl/fl littermates in Fig. 1g. h–l, mice were intraperitoneally infected with 5,000 PFU MHV-A59 and assessed for (h) survival (n = 47 Ly6efl/fl, n = 15 Ly6eΔLyz2, n = 14 Ly6eΔCD11c, n = 14 Ly6eΔCD4, n = 5 Ly6eΔCD8a, and n = 16 Ly6eΔCD19), viral burden in brain, lung, spleen, and liver (i), viral burden in serum (j), serum alanine aminotransferase (k), and post-mortem spleen weight (l). In i-l, data represents means from n = 30 Ly6efl/fl, n=13 Ly6eΔLysM, n = 14 Ly6eΔCD11c, n = 10 Ly6eΔCD4, n = 6 Ly6eΔCD8a, and n = 14 Ly6eΔCD19. Male and female mice were used at an approximately 1 to 1 ratio for these experiments. Statistical significance was determined by log-rank (Mantel-Cox) tests (a, h), two-sided Mann-Whitney test (b-d), two-sided unpaired t-test (e-f), Kruskal-Wallis test (i-j), and one-way ANOVA (k-l). Error bars represent mean ± standard deviation.
Extended Data Fig. 2
a, Flow cytometry gating strategy for sorting lung CD45+ cells and CD31+ cells from Ly6eΔVav1 mice for determining Ly6e gene expression as shown in Fig. 2c,d. b, Flow cytometry gating strategy for identifying donor (CD45.1+) and recipient (CD45.2+) immune cells. c, Relative composition of CD45.1+ and CD45.2+ immune cells of compartment total CD45+ cells in lung, spleen, and blood. d, Relative Ly6e mRNA levels in brain, heart, lung, liver, and spleen from Ly6e wildtype, heterozygous, and knockout mice. In c, data represents means from n = 3 Ly6efl/fl; CD45.2/.2, n = 3 Ly6efl/fl; CD45.1/.1, n = 3 Ly6efl→fl, n = 3 Ly6efl→ΔVav1; d, n = 3 Ly6e wildtype, n = 3 Ly6e heterozygous, n = 3 Ly6e knockout. Error bars represent mean ± standard deviation.
Extended Data Fig. 3
a, Ly6e wildtype and knockout mice were intranasally infected with 8,700 PFU SARS-CoV-2 and monitored daily for weight loss. Lung viral burden measured by quantitative PCR for Ly6e wildtype and knockout mice (b) or C57BL/6J, Ifnar−/−, and Ifnlr−/− mice (c) that were infected with 60,000 PFU SARS-CoV-2 and euthanized 3 days post-infection. d, Lung Ly6e expression from C57BL/6J, Ifnar−/−, and Ifnlr−/− mice that were mock treated with PBS or infected with 8,700 PFU SARS-CoV-2 and euthanized the next day. Expression is shown relative to PBS-treated C57BL/6J mice. Expression of Ly6e (e) and Mx1 (f) in lungs from C57BL/6J mice treated intranasally with recombinant human IFNλ or retro-orbitally with recombinant murine IFNβ relative to untreated (untr). In a, data represents means from n = 5 Ly6e wildtype, n = 5 Ly6e knockout mice; b, n = 13 Ly6e wildtype, n = 13 Ly6e knockout mice; c, n = 8 C57BL6/J, n = 8 Ifnar−/−, and n=8 Ifnlr−/−; d, n = 5 for each group and genotype; e-f, n = 6 untreated, n = 9 IFNλ-treated, n = 8 IFNβ-treated. g, Representative hematoxylin and eosin-stained lung sections used for analysis shown in Fig. 3e from Ly6e wildtype (n = 6) and Ly6e knockout (n = 6) mice euthanized 3 days after treated with PBS or intranasal infection with 60,000 PFU SARS-CoV-2. Example of automated image analysis of SARS-CoV-2 infected cells for Fig. 3g–h (h) and of CD45+ cells for Fig. 3i, j (i). In the corresponding markup images, cells without DAB marker (for example positive stain) are colored blue, and positive cells identified as weak positive, moderate positive, and strong positive for the DAB marker are colored as yellow, orange, and red respectively. Statistical significance was determined by two-sided unpaired t-test (a), two-sided Mann-Whitney test (b-c), one way ANOVA with Holm-Šídák′s multiple comparisons test (d–f). Error bars represent mean ± SEM in a and mean ± SD in b-f. Scale bars: 600 μm (g), 1 mm and 100 μm (h-i).
Extended Data Fig. 4
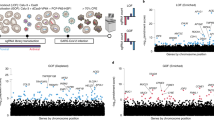
mRNA sequencing of lungs from Ly6e wildtype and Ly6e knockout mice that were intranasally infected with 60,000 PFU P.1 SARS-CoV-2 and euthanized 3 days post-infection. Volcano plot summarization of differentially expressed genes (DEGs) between uninfected Ly6e wildtype (n = 6) and Ly6e knockout (n = 4) mice (a), uninfected wildtype (n = 6) and SARS-CoV-2 infected wildtype (n = 7) mice (b), and uninfected knockout (n = 4) and SARS-CoV-2 infected knockout (n=7) mice (c). d, Z-score heatmap of the top 25 DEGs for data summarized in a. e, Gene Ontology (GO) enrichment analysis of transcriptomic data from SARS-CoV-2 infected Ly6e wildtype and Ly6e knockout mice. The abscissa is the ratio of the number of differential genes linked with the GO term to the total number of differential genes. The size of a point represents the number of genes annotated to a specific GO term, and the color from red to purple represents the significant level of the enrichment. Male and female mice were used at an approximately 1 to 1 ratio for these experiments. For a-c, P-values were determined using the negative binomial distribution model and adjusted for multiple hypothesis testing using the Benjamini and Hochberg’s approach for controlling the false discovery rate. P-values for e were determined using hypergeometric distribution and adjusted testing using the Benjamini and Hochberg’s approach for controlling the false discovery rate.
Extended Data Fig. 5
Differential expression of select genes highlighted in Fig. 4a between SARS-CoV-2-infected Ly6e wildtype mice and uninfected Ly6e wildtype mice (a) and SARS-CoV-2-infected Ly6e knockout mice and uninfected Ly6e knockout mice (b).
Extended Data Fig. 6
Mice were infected with 8,700 PFU P.1 SARS-CoV-2 and euthanized 1 day post-infection. Lung sections were probed for SARS-CoV-2 RNA (green) and Scgb1a1 mRNA (red) (a, c) or Scgb3a1 mRNA (red) (b, d). Lung sections from n = 3 Ly6e knockout mice were stained and imaged. Images shown are from two different mice than in Fig. 4. Scale bars: 100 µM (a-b), 25 µM (c-d).
Extended Data Fig. 7
Mice were infected with 8,700 PFU SARS-CoV-2 and euthanized 1 day post-infection. Lung sections were probed for SARS-CoV-2 RNA (green) and Sftpc mRNA (red) (a, c) or for SARS-CoV-2 nucleocapsid (green) and acetylated tubulin (red) (b, d). Lung sections from n = 3 Ly6e knockout mice were stained and imaged. Images shown are from different mice than in Fig. 4. Scale bars: 100 µM (a-b), 25 µM (c-d).
Extended Data Fig. 8
a, Flow cytometry gating strategy used in Fig. 4g–h for detecting SARS-CoV-2 nucleocapsid (N) in SCGB1A1-positive and acetylated tubulin-positive pulmonary epithelial cells. Representative plots are from uninfected Ly6e knockout mice.
Extended Data Fig. 9
a, Area under the curve analysis of SARS-CoV-2 infectivity in HBE-ACE2 cells from n = 3 independent experiments, b, Western blot of two independent preparations of control and LY6E knockout cells used in a. Statistical significance was determined by two-sided unpaired t-test (a). Error bars represent mean ± standard deviation.
Supplementary information
Supplementary Information
Supplementary Tables 1–5.
Source data
Source Data Fig. 1
Unprocessed western blot for Extended Dat Fig. 9.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Mar, K.B., Wells, A.I., Caballero Van Dyke, M.C. et al. LY6E is a pan-coronavirus restriction factor in the respiratory tract. Nat Microbiol 8, 1587–1599 (2023). https://doi.org/10.1038/s41564-023-01431-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41564-023-01431-w
This article is cited by
-
Bacterial-induced or passively administered interferon gamma conditions the lung for early control of SARS-CoV-2
Nature Communications (2023)
